Biosynthesis of Medium- to Long-Chain α,ω-Diols from Free Fatty Acids Using CYP153A Monooxygenase, Carboxylic Acid Reductase, and E. coli Endogenous Aldehyde Reductases

 ,
, 

Abstract
1. Introduction
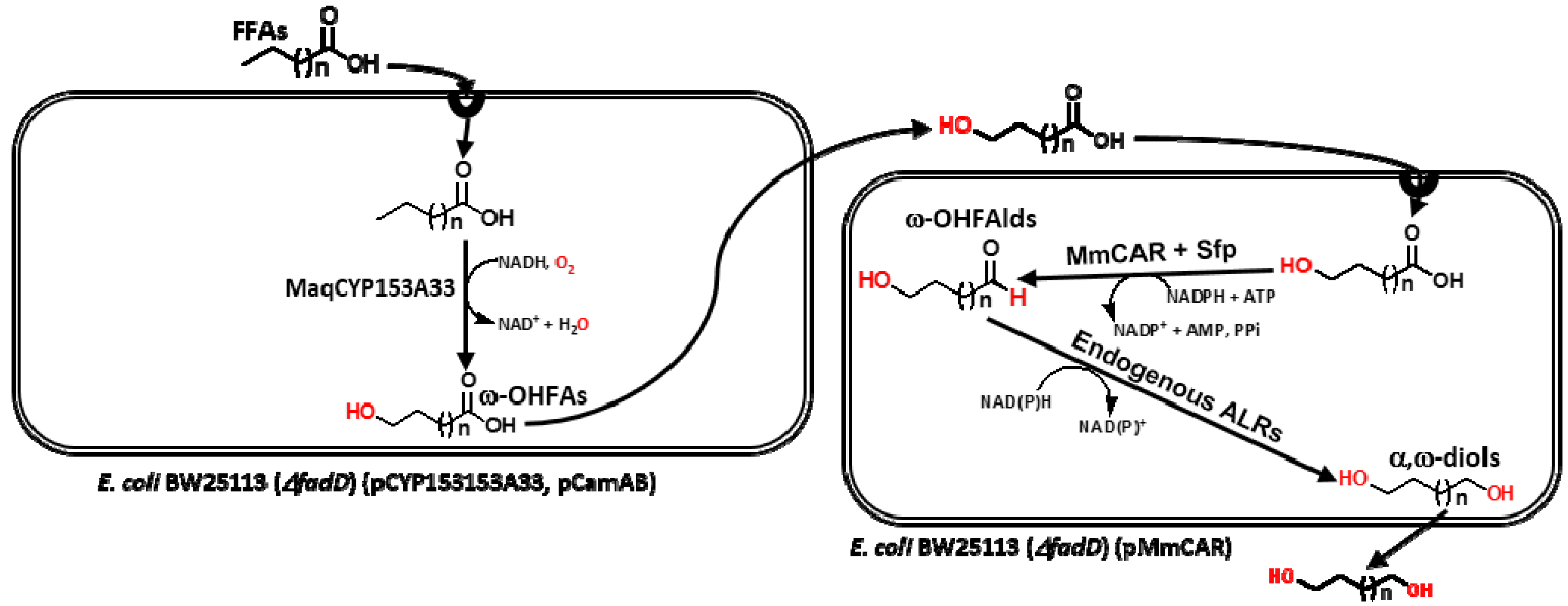
2. Results and Discussion
2.1. Establishment of MmCAR Reaction System
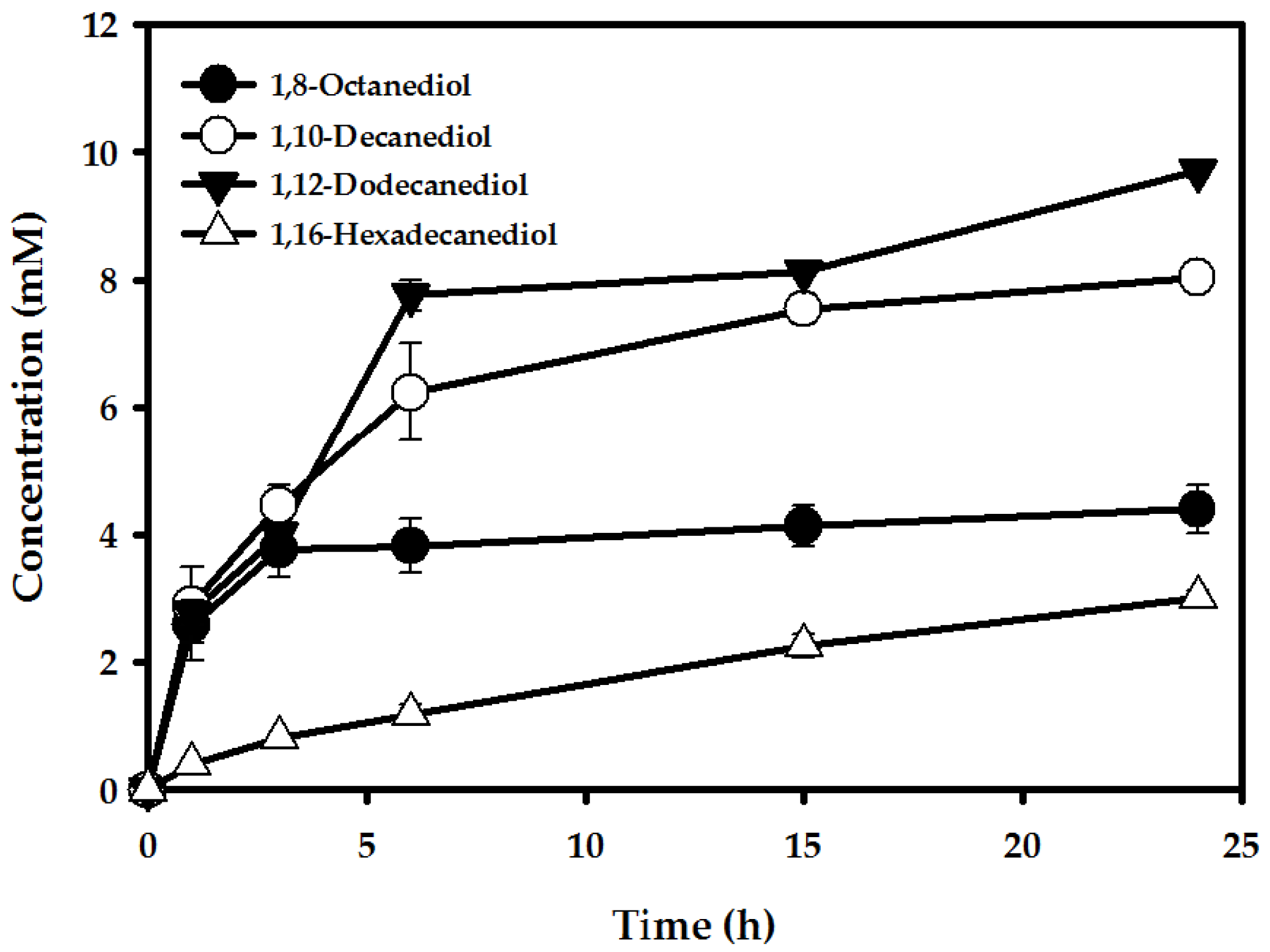
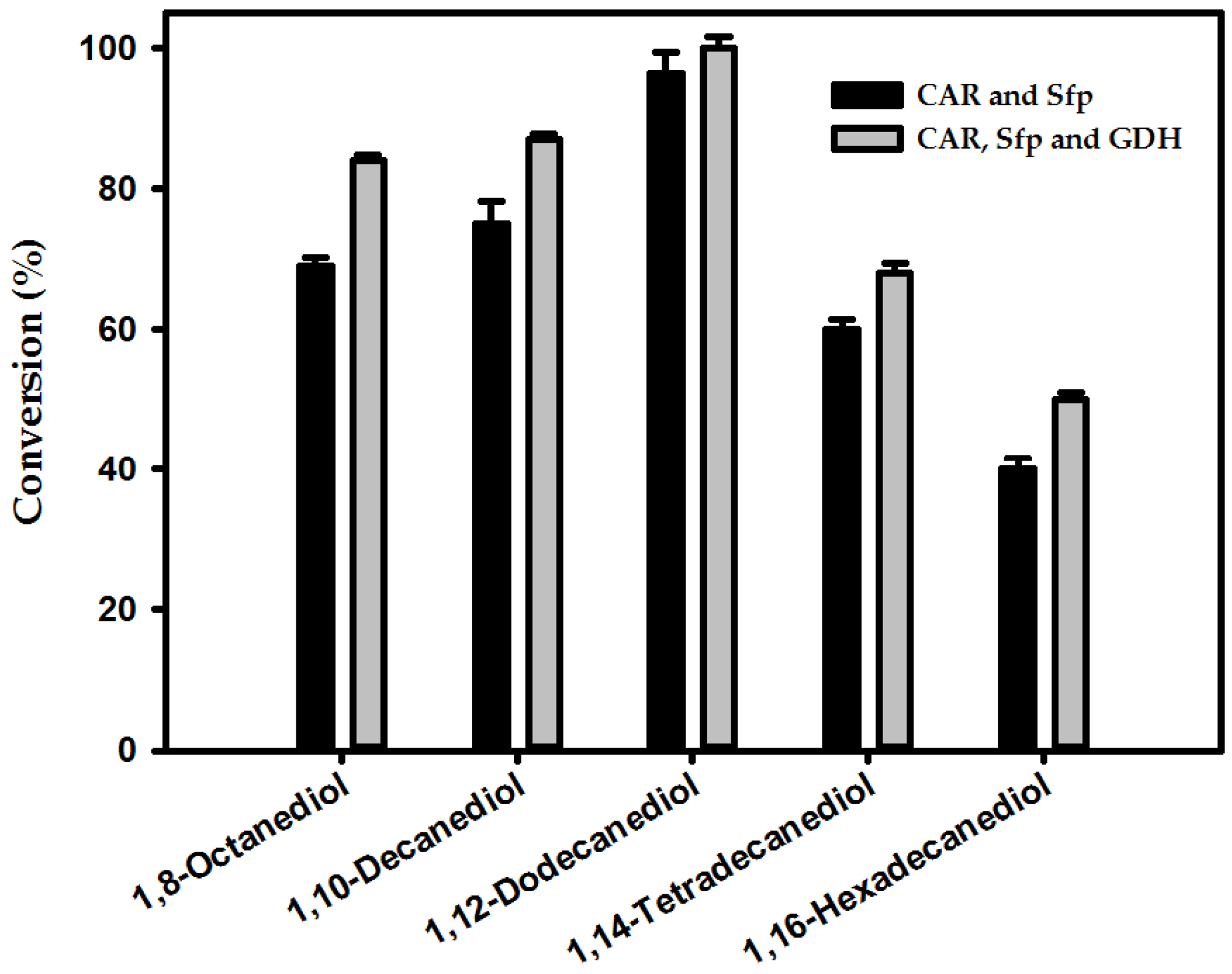
2.2. Production of α,ω-Diols from ω-OHFAs
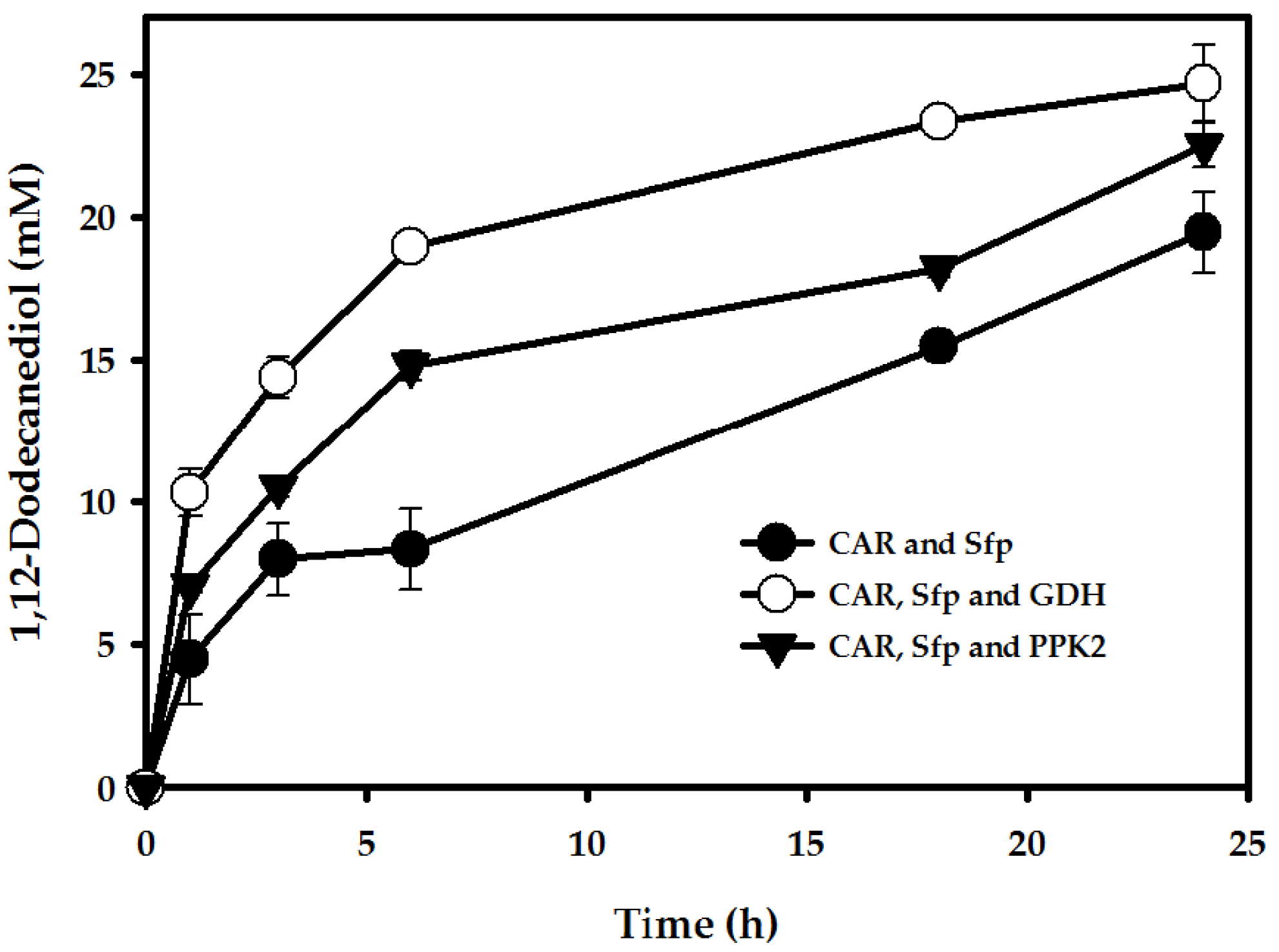
2.3. Enhancement of 1,12-Dodecanediol Production
2.3.1. pH Optimization
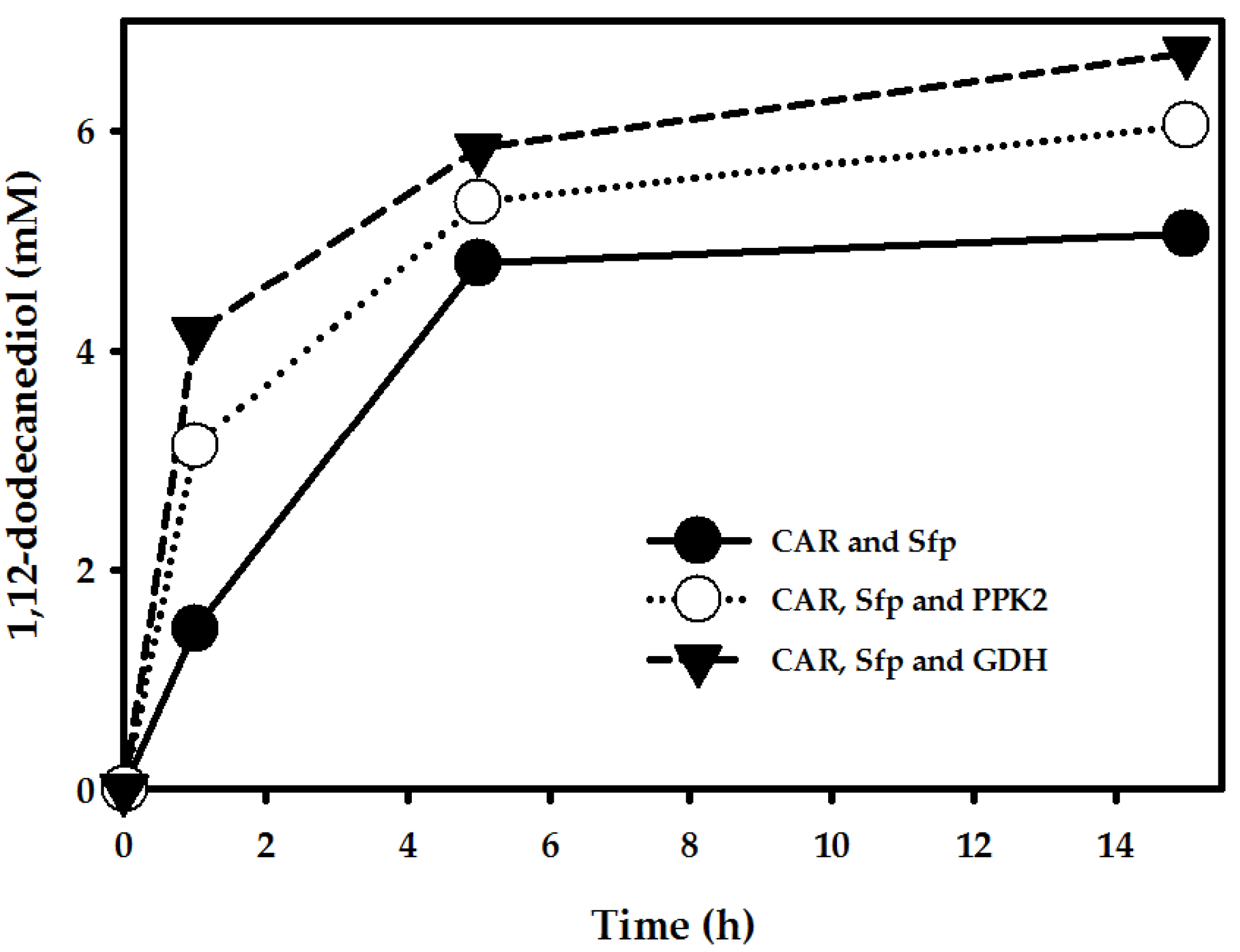
2.3.2. Cofactor Regeneration
2.4. Production of α,ω-Diols from FFAs
2.4.1. Production of α,ω-Diols by One-Pot, One-Step Reaction
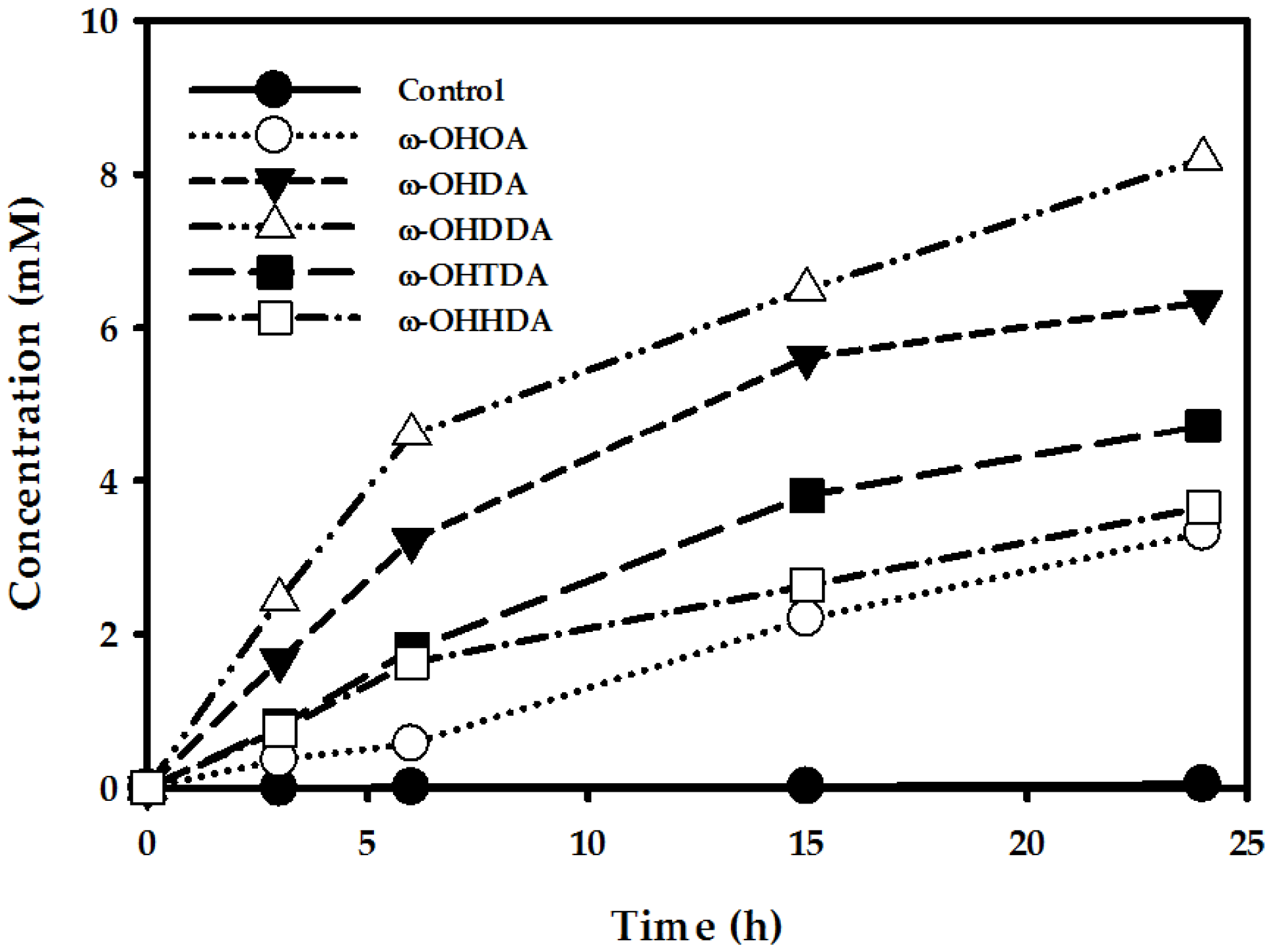
2.4.2. Production of α,ω-Diols Using Biotransformed ω-OHFAs
Production of Various ω-OHFAs (C8–C16) by CYP153A Monooxygenase
Production of α,ω-Diols Using Biotransformed ω-OHFAs
3. Materials and Methods
3.1. Chemicals
3.2. Plasmid Construction and Gene Manipulation
3.3. Protein Expression and Biotransformation
3.4. Analysis of Compounds by Gas Chromatography
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kawaguchi, H.; Ogino, C.; Kondo, A. Microbial conversion of biomass into bio-based polymers. Bioresour. Technol. 2017, 245, 1664–1673. [Google Scholar] [CrossRef] [PubMed]
- Lligadas, G.; Ronda, J.C.; Galià, M.; Cádiz, V. Oleic and undecylenic acids as renewable feedstocks in the synthesis of polyols and polyurethanes. Polymers 2010, 2, 440–453. [Google Scholar] [CrossRef]
- Gandini, A. Polymers from renewable resources: A challenge for the future of macromolecular materials. Macromolecules 2008, 41, 9491–9504. [Google Scholar] [CrossRef]
- Lligadas, G.; Ronda, J.C.; Galià, M.; Cádiz, V. Renewable polymeric materials from vegetable oils: A perspective. Mater. Today 2013, 16, 337–343. [Google Scholar] [CrossRef]
- Sharma, V.; Kundu, P.P. Condensation polymers from natural oils. Prog. Polym. Sci. 2008, 33, 1199–1215. [Google Scholar] [CrossRef]
- Madhavan, A.; Sindhu, R.; Binod, P.; Sukumaran, R.K.; Pandey, A. Strategies for design of improved biocatalysts for industrial applications. Bioresour. Technol. 2017, 245, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Si, T.; Zhao, H. Biocatalyst development by directed evolution. Bioresour. Technol. 2012, 115, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Liu, W.; Zou, H.; Cheng, T.; Tian, N.; Xian, M. Microbial production of short chain diols. Microb. Cell Fact. 2014, 13, 165. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, L.; Liu, Q.; Zhang, H.; Cao, Y.; Xian, M.; Liu, H. High-specificity synthesis of novel monomers by remodeled alcohol hydroxylase. BMC Biotechnol. 2016, 16, 61. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Narikawa, T.; Sumisa, F.; Arisawa, A.; Takeda, K.; Kato, J. Production of α,ω-alkanediols using Escherichia coli expressing a cytochrome P450 from Acinetobacter sp. OC4. Biosci. Biotechnol. Biochem. 2006, 70, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Scheller, U.; Zimmer, T.; Becher, D.; Schauer, F.; Schunck, W.-H. Oxygenation cascade in conversion of n-alkanes to α, ω-dioic acids catalyzed by cytochrome P450 52A3. J. Biol. Chem. 1998, 273, 32528–32534. [Google Scholar] [CrossRef] [PubMed]
- Shiio, I.; Uchio, R. Microbial production of long-chain dicarboxylic acids from n-alkanes. Agric. Biol. Chem. 1971, 35, 2033–2042. [Google Scholar] [CrossRef]
- Schaffer, S.; Haas, T. Biocatalytic and fermentative production of α,ω-bifunctional polymer precursors. Org. Process. Res. Dev. 2014, 18, 752–766. [Google Scholar] [CrossRef]
- Scheps, D.; Honda Malca, S.; Hoffmann, H.; Nestl, B.M.; Hauer, B. Regioselective ω-hydroxylation of medium-chain n-alkanes and primary alcohols by CYP153 enzymes from Mycobacterium marinum and Polaromonas sp. strain JS666. Org. Biomol. Chem. 2011, 9, 6727–6733. [Google Scholar] [CrossRef] [PubMed]
- Sabra, W.; Groeger, C.; Zeng, A.-P. Microbial cell factories for diol production. In Bioreactor Engineering Research and Industrial Applications I: Cell Factories; Ye, Q., Bao, J., Zhong, J.-J., Eds.; Springer: Berlin/Heidelberg, Geramny, 2016; pp. 165–197. [Google Scholar]
- Honda Malca, S.; Scheps, D.; Kuhnel, L.; Venegas-Venegas, E.; Seifert, A.; Nestl, B.M.; Hauer, B. Bacterial CYP153A monooxygenases for the synthesis of omega-hydroxylated fatty acids. Chem. Commun. 2012, 48, 5115–5117. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.K.; Turner, N.J.; Jones, P.R. Carboxylic acid reductase is a versatile enzyme for the conversion of fatty acids into fuels and chemical commodities. Proc. Natl. Acad. Sci. USA 2013, 110, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Fatma, Z.; Jawed, K.; Mattam, A.J.; Yazdani, S.S. Identification of long chain specific aldehyde reductase and its use in enhanced fatty alcohol production in E. coli. Metab. Eng. 2016, 37, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, G.M.; Atsumi, S. Toward aldehyde and alkane production by removing aldehyde reductase activity in Escherichia coli. Metab. Eng. 2014, 25, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Finnigan, W.; Thomas, A.; Cromar, H.; Gough, B.; Snajdrova, R.; Adams, J.P.; Littlechild, J.A.; Harmer, N.J. Characterization of carboxylic acid reductases as enzymes in the toolbox for synthetic chemistry. ChemCatChem 2017, 9, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Khusnutdinova, A.N.; Flick, R.; Popovic, A.; Brown, G.; Tchigvintsev, A.; Nocek, B.; Correia, K.; Joo, J.C.; Mahadevan, R.; Yakunin, A.F. Exploring bacterial carboxylate reductases for the reduction of bifunctional carboxylic acids. Biotechnol. J. 2017. [Google Scholar] [CrossRef] [PubMed]
- Venkitasubramanian, P.; Daniels, L.; Rosazza, J.P. Reduction of carboxylic acids by Nocardia aldehyde oxidoreductase requires a phosphopantetheinylated enzyme. J. Biol. Chem. 2007, 282, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Lundemo, M.T.; Notonier, S.; Striedner, G.; Hauer, B.; Woodley, J.M. Process limitations of a whole-cell P450 catalyzed reaction using a CYP153A-CPR fusion construct expressed in Escherichia coli. Appl. Microbiol. Biotechnol. 2016, 100, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Durairaj, P.; Lee, D.; Ahsan, M.M.; Yun, H. Improved nadph regeneration for fungal cytochrome P450 monooxygenase by co-expressing bacterial glucose dehydrogenase in resting-cell biotransformation of recombinant yeast. J. Microbiol. Biotechnol. 2016, 26, 2076–2086. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.; Park, B.G.; Ahsan, M.M.; Kim, J.; Yun, H.; Choi, K.-Y.; Kim, B.-G. Production of ω-hydroxy palmitic acid using CYP153A35 and comparison of cytochrome P450 electron transfer system in vivo. Appl. Microbiol. Biotechnol. 2016, 100, 10375–10384. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J. Molecular Cloning: A Laboratory Manual/Joseph Sambrook, David W. Russell; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Hanahan, D. Studies on transformation of escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Ahsan, M.M.; Jeon, H.; Nadarajan, S.P.; Chung, T.; Yoo, H.-W.; Kim, B.-G.; Patil, M.D.; Yun, H. Biosynthesis of the Nylon 12 monomer, ω-aminododecanoic acid with novel CYP153A, AlkJ, and ω-TA enzymes. Biotechnol. J. 2017. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmids/Strains | Description | Reference |
|---|---|---|
| Plasmids | ||
| pET24ma | P15A ori lacI T7 promoter, KmR | [25] |
| pCYP153A33 | pET24ma encoding for MaqCYP153A33 | [25] |
| pETDuet-1 | pBR322 ori lacI T7 promoter, AmpR | Novagen |
| pCamAB | pETDuet-1 encoding for CamA and CamB | [25] |
| pMmCAR | pETDuet-1 encoding for MmCAR | This study |
| pCAR-GDH | pETDuet-1 encoding for MmCAR and GDH | This study |
| pCAR-PK2 | pETDuet-1 encoding for MmCAR and PPK2 | This study |
| pSFP | pET24ma encoding for Sfp | This study |
| E. coli strains | ||
| DH5α | F−, endA1, glnV44, thi-1, recA1, relA1, gyrA96 deoR, supE44, Φ80dlacZ∆M15 ∆(lacZYA−-argF)-U169, hsdR17(rK− mK+), λ− | [27] |
| BW25113(DE3) | rrnB3 ΔlacZ4787 hsdR514 Δ(araBAD)567 Δ(rhaBAD)568 rph-1 λ(DE3) | [25] |
| DL | BW25113(DE3) ΔfadD | [28] |
| A33AB | DL carrying pCYP153A33 and pCamAB | [28] |
| CAR | DL carrying pCAR | This study |
| CS | DL carrying pCAR and pSFP | This study |
| CSG | DL carrying pCAR-GDH and pSFP | This study |
| CSK | DL carrying pCAR-PK2 and pSFP | This study |
| PCR oligonucleotides | Sequence (5′-3′) | |
| CAR_F (Bam HI) | ATTAGGATCCCATGTCGCCAATCACGCGTG | Cloned into pETDuet1 |
| CAR_R (Hindd III) | TAATAAGCTTGAGCAGGCCGAGTAGGCG | |
| SFP_F (Nde I) | ATTACATATGAAAATCTACGGCATCTAC | Cloned into pET24ma |
| SFP_R (Xho I) | TAATCTCGAGTTACAGCAGTTCTTCATA | |
| PPK2_F (Nde I) | AAGGAGATATACATATGCCGATGGTTGCTGCAG | Cloned into pETDuet1 |
| PPK2_R (Kpn I) | TTACCAGACTCGAGGGTACCTTAAGATTCAACAACCAGAGC | |
| GDH_F (NdeI) | AAGGAGATATACATATGTATCCGGATTTAAAAGGA | Cloned into pETDuet1 |
| GDH_R (Kpn I) | TTACCAGACTCGAGGGTACCTTAACCGCGGCCTGCCTGGAA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahsan, M.M.; Sung, S.; Jeon, H.; Patil, M.D.; Chung, T.; Yun, H. Biosynthesis of Medium- to Long-Chain α,ω-Diols from Free Fatty Acids Using CYP153A Monooxygenase, Carboxylic Acid Reductase, and E. coli Endogenous Aldehyde Reductases. Catalysts 2018, 8, 4. https://doi.org/10.3390/catal8010004
Ahsan MM, Sung S, Jeon H, Patil MD, Chung T, Yun H. Biosynthesis of Medium- to Long-Chain α,ω-Diols from Free Fatty Acids Using CYP153A Monooxygenase, Carboxylic Acid Reductase, and E. coli Endogenous Aldehyde Reductases. Catalysts. 2018; 8(1):4. https://doi.org/10.3390/catal8010004
Chicago/Turabian StyleAhsan, Md Murshidul, Sihyong Sung, Hyunwoo Jeon, Mahesh D. Patil, Taeowan Chung, and Hyungdon Yun. 2018. "Biosynthesis of Medium- to Long-Chain α,ω-Diols from Free Fatty Acids Using CYP153A Monooxygenase, Carboxylic Acid Reductase, and E. coli Endogenous Aldehyde Reductases" Catalysts 8, no. 1: 4. https://doi.org/10.3390/catal8010004
APA StyleAhsan, M. M., Sung, S., Jeon, H., Patil, M. D., Chung, T., & Yun, H. (2018). Biosynthesis of Medium- to Long-Chain α,ω-Diols from Free Fatty Acids Using CYP153A Monooxygenase, Carboxylic Acid Reductase, and E. coli Endogenous Aldehyde Reductases. Catalysts, 8(1), 4. https://doi.org/10.3390/catal8010004