
Purification and Characteristics of β-Amylase from Soybean Whey Wastewater


Abstract
1. Introduction
2. Results and Discussion
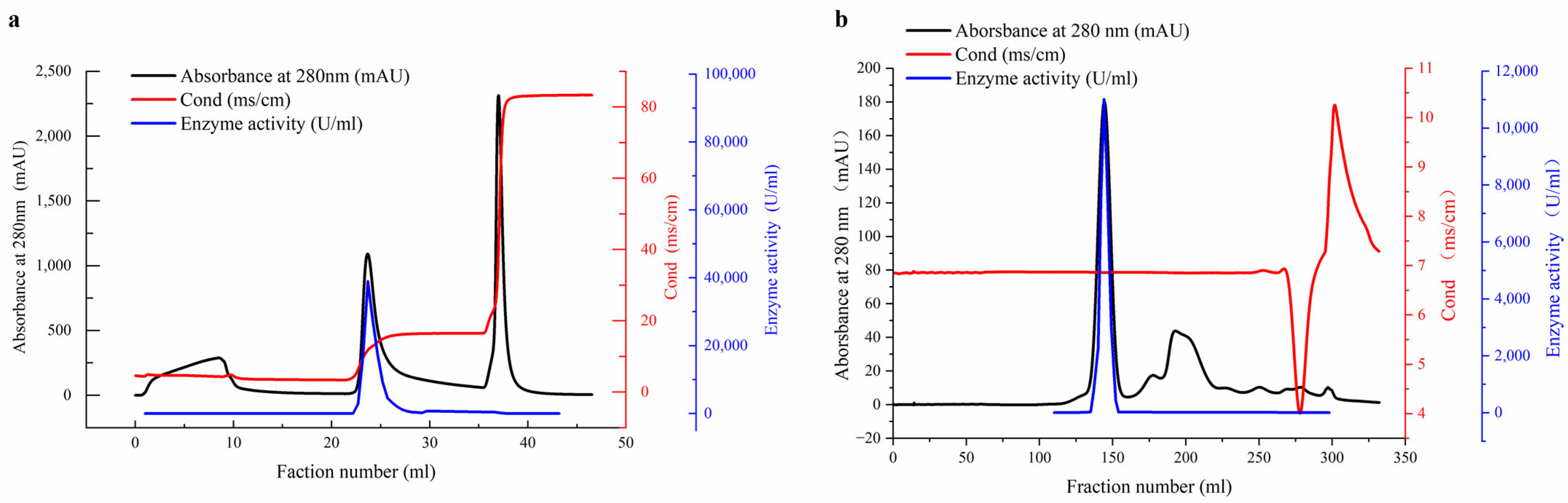
2.1. Purification of β-Amylase
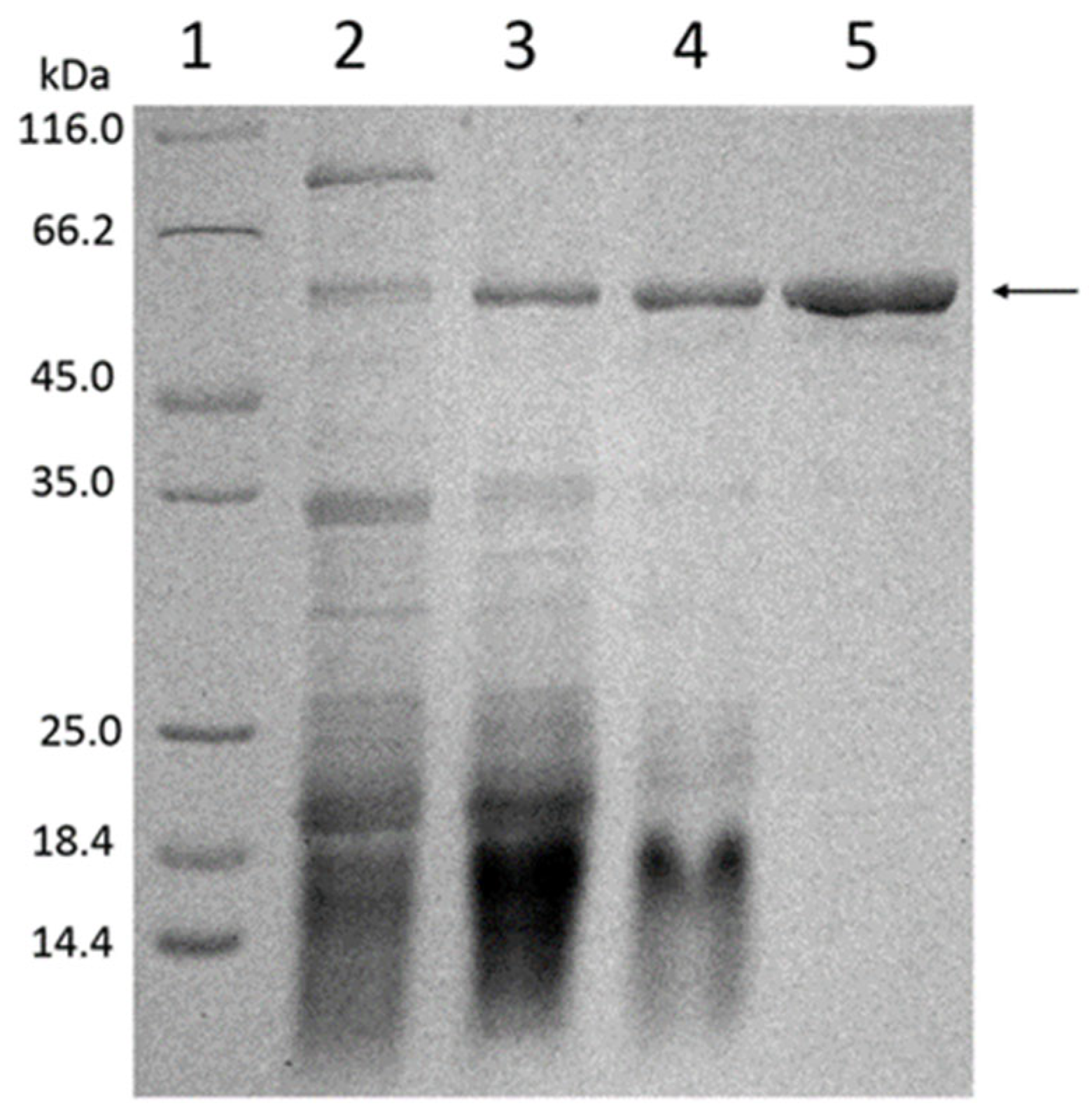
2.2. Molecular Mass Determination
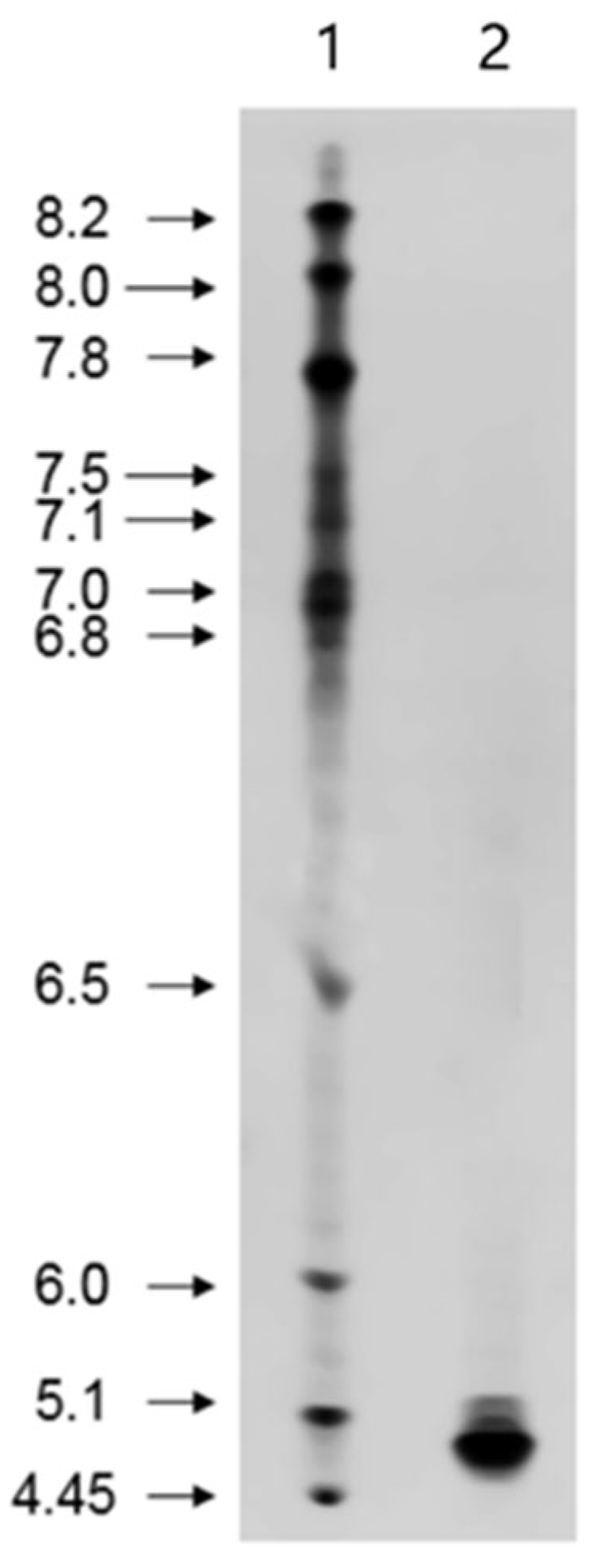
2.3. Isoelectric Point Determination
2.4. Effect of Temperature on Activity and Stability of Purified β-Amylase
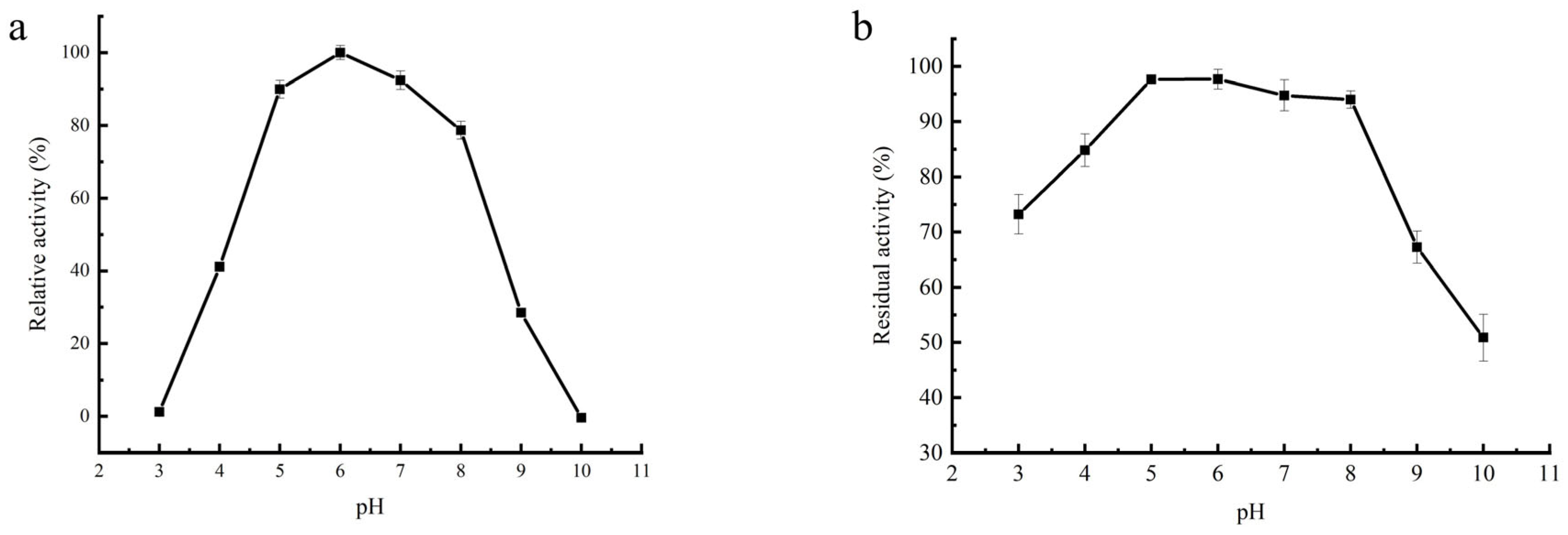
2.5. Effect of pH on Activity and Stability of Purified β-Amylase
2.6. Effects of Metal Ions and Surface-Active Agents on β-Amylase Activity
2.7. Substrate Specificity and Kinetics Studies
3. Materials and Methods
3.1. Materials and Reagents
3.2. Enzyme Activity Assay
3.3. Protein Determination
3.4. Extraction of the Crude β-Amylase
3.5. Purification of the β-Amylase
3.6. SDS-PAGE for Purity and Molecular Mass Determination
3.7. Isoelectric Focusing of Purified β-Amylase
3.8. Effect of Temperature on β-Amylase Activity and Stability
3.9. Effect of pH on β-Amylase Activity and Stability
3.10. Effects of Metal Ions, Inhibitors, and Surfactants on β-Amylase Activity
3.11. Substrate Specificity
3.12. Determination of Kinetic Parameters
3.13. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hu, M.; Gao, Y.; Wen, W.; Zhang, P.; Zhang, F.; Fan, B.; Wang, F.; Li, S. The Aggregation Behavior between Soybean Whey Protein and Polysaccharides of Diverse Structures and Their Implications in Soybean Isoflavone Delivery. Food Chem. 2024, 439, 138061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, C.; Wang, J.; Wen, Y.; Li, H.; Liu, X. The Investigation of Soybean Protein Isolates and Soybean Peptides Assisting Lactobacillus plantarum K25 to Inhibit Escherichia coli. Curr. Res. Food Sci. 2024, 8, 100662. [Google Scholar] [CrossRef] [PubMed]
- Chua, J.-Y.; Liu, S.-Q. Soy Whey: More than Just Wastewater from Tofu and Soy Protein Isolate Industry. Trends Food Sci. Technol. 2019, 91, 24–32. [Google Scholar] [CrossRef]
- Liu, C.; Sun, F.; Tian, Y.; Jiang, L.; Wang, Z.; Zhou, L. Recovery of Soy Whey Protein from Soy Whey Wastewater at Various Cavitation Jet Pretreatment Time and Their Structural and Emulsifying Properties. Food Chem. X 2024, 21, 101122. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, H.; Tian, J.; Shi, J.; Linhardt, R.J.; Ye, T.D.X.; Chen, S. Recovery of High Value-added Nutrients from Fruit and Vegetable Industrial Wastewater. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1388–1402. [Google Scholar] [CrossRef]
- Sorgentini, D.A.; Wagner, J.R. Comparative Study of Structural Characteristics and Thermal Behavior of Whey and Isolate Soybean Proteins. J. Food Biochem. 1999, 23, 489–507. [Google Scholar] [CrossRef]
- Wang, Y.; Serventi, L. Sustainability of Dairy and Soy Processing: A Review on Wastewater Recycling. J. Clean. Prod. 2019, 237, 117821. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, S.; Lu, C.; Zhang, Y.; Zhao, H. Enrichment of Trypsin Inhibitor from Soybean Whey Wastewater Using Different Precipitating Agents and Analysis of Their Properties. Molecules 2024, 29, 2613. [Google Scholar] [CrossRef]
- Nieto-Veloza, A.; Zhong, Q.; Kim, W.-S.; D’Souza, D.; Krishnan, H.B.; Dia, V.P. Utilization of Tofu Processing Wastewater as a Source of the Bioactive Peptide Lunasin. Food Chem. 2021, 362, 130220. [Google Scholar] [CrossRef]
- Jiang, C.; Wu, Z.; Li, R.; Liu, Q. Technology of Protein Separation from Whey Wastewater by Two-Stage Foam Separation. Biochem. Eng. J. 2011, 55, 43–48. [Google Scholar] [CrossRef]
- Li, H.; Liu, J.; Li, D.; Wang, H. Study on Separation and Purification of Genistein in the Soybean Residue Using Macroporous Resin Adsorption. Ind. Eng. Chem. Res. 2012, 51, 44–49. [Google Scholar] [CrossRef]
- Liu, W.; Wu, Z.L.; Wang, Y.J.; Zhao, Y.L.; Liu, W.C.; Yu, Y. Recovery of Isoflavones from the Soy Whey Wastewater Using Two-Stage Batch Foam Fractionation. Ind. Eng. Chem. Res. 2013, 52, 13761–13767. [Google Scholar] [CrossRef]
- Guan, X.; Wang, Q.; Lin, B.; Sun, M.; Zheng, Q.; Huang, J.; Lai, G. Structural Characterization of a Soluble Polysaccharide SSPS1 from Soy Whey and Its Immunoregulatory Activity in Macrophages. Int. J. Biol. Macromol. 2022, 217, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Ying, Z.; Li, W.; Li, H.; Liu, X. Trypsin Inhibitor from Soybean Whey Wastewater: Isolation, Purification and Stability. Appl. Sci. 2022, 12, 10084. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, M.; Lv, Y.; Tian, S.; Li, N.; Wu, Z. Foam Fractionation for Recovering Whey Soy Protein from Whey Wastewater: Strengthening Foam Drainage Using a Novel Internal Component with Superhydrophobic Surface. J. Taiwan Inst. Chem. Eng. 2017, 78, 39–44. [Google Scholar] [CrossRef]
- Baba, T.; Kainuma, K. Partial Hydrolysis of Sweet-Potato Starch with β-Amylase. Agric. Biol. Chem. 1987, 51, 1365–1371. [Google Scholar] [CrossRef]
- Qi, J.-C.; Zhang, G.-P.; Zhou, M.-X. Protein and Hordein Content in Barley Seeds as Affected by Nitrogen Level and Their Relationship to β-Amylase Activity. J. Cereal Sci. 2006, 43, 102–107. [Google Scholar] [CrossRef]
- Folasade, M.O.; Folasade, M.A. Production, Purification and Partial Characterization of Moderately Thermostable β-Amylase from Bacillus polymyxa BWB-01. Curr. Biotechnol. 2015, 4, 187–196. [Google Scholar]
- Vajravijayan, S.; Pletnev, S.; Mani, N.; Pletneva, N.; Nandhagopal, N.; Gunasekaran, K. Structural Insights on Starch Hydrolysis by Plant β-Amylase and Its Evolutionary Relationship with Bacterial Enzymes. Int. J. Biol. Macromol. 2018, 113, 329–337. [Google Scholar] [CrossRef]
- Agrawal, D.C.; Dwevedi, A.; Kayastha, A.M. Biochemical and Thermodynamic Characterization of de Novo Synthesized β-Amylase from Fenugreek. Int. J. Biol. Macromol. 2019, 130, 786–797. [Google Scholar] [CrossRef]
- Niu, C.; Zheng, F.; Li, Y.; Liu, C.; Li, Q. Process Optimization of the Extraction Condition of β-amylase from Brewer’s Malt and Its Application in the Maltose Syrup Production. Biotechnol. Appl. Biochem. 2018, 65, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Amylase Research Society of Japan. Handbook of Amylases and Related Enzymes: Their Sources, Isolation Methods, Properties and Applications; Pergamon: Oxford, UK, 1988. [Google Scholar]
- Obe, D.A.; Fatoki, T.H. In Silico Evaluation of the Structural Dynamics β-Amylase from Sweet Potato (Ipomoea batatas). Asian J. Biotechnol. Bioresour. Technol. 2021, 7, 1–10. [Google Scholar] [CrossRef]
- Ma, Y.; Stewart, D.C.; Eglinton, J.K.; Logue, S.J.; Langridge, P.; Evans, D.E. Comparative Enzyme Kinetics of Two Allelic Forms of Barley (Hordeum vulgare L.) β-Amylase. J. Cereal Sci. 2000, 31, 335–344. [Google Scholar] [CrossRef]
- Li, R.; Ji, X.; Zhu, Y.; Zhang, Y.; Guan, J. Precipitation of Proteins from Soybean Whey Wastewater by Successive Foaming and Defoaming. Chem. Eng. Process. Process Intensif. 2018, 128, 124–131. [Google Scholar] [CrossRef]
- Dicko, M.H.; Searle-van Leeuwen, M.J.F.; Beldman, G.; Ouedraogo, O.G.; Hilhorst, R.; Traoré, A.S. Purification and Characterization of β-Amylase from Curculigo pilosa. Appl. Microbiol. Biotechnol. 1999, 52, 802–805. [Google Scholar] [CrossRef]
- Van Damme, E.J.M.; Hu, J.; Barre, A.; Hause, B.; Baggerman, G.; Rougé, P.; Peumans, W.J. Purification, Characterization, Immunolocalization and Structural Analysis of the Abundant Cytoplasmic β-amylase from Calystegia sepium (Hedge Bindweed) Rhizomes. Eur. J. Biochem. 2001, 268, 6263–6273. [Google Scholar] [CrossRef] [PubMed]
- Kolawole, A.O.; Ajele, J.O.; Sirdeshmukh, R. Purification and Characterization of Alkaline-Stable β-Amylase in Malted African Finger Millet (Eleusine coracana) Seed. Process Biochem. 2011, 46, 2178–2186. [Google Scholar] [CrossRef]
- Viksø-Nielsen, A.; Christensen, T.M.I.E.; Bojko, M.; Marcussen, J. Purification and Characterization of β-amylase from Leaves of Potato (Solanum tuberosum). Physiol. Plant. 1997, 99, 190–196. [Google Scholar] [CrossRef]
- Lin, J.; Lin, Y.-S.; Kuo, S.-T.; Jiang, C.-M.; Wu, M.-C. Purification of Soybean Amylase by Superparamagnetic Particles. Food Chem. 2009, 117, 94–98. [Google Scholar] [CrossRef]
- Lizotte, P.A.; Henson, C.A.; Duke, S.H. Purification and Characterization of Pea Epicotyl β-Amylase. Plant Physiol. 1990, 92, 615–621. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, G.; Zhang, T.; Duan, C.; Liu, Z.; Qin, L. Properties of β-Amylase from China Soybean Core Collection. J. Chin. Cereals Oils Assoc. 2006, 21, 88–92. [Google Scholar]
- Yoon, S.-H.; Robyt, J.F. Activation and Stabilization of 10 Starch-Degrading Enzymes by Triton X-100, Polyethylene Glycols, and Polyvinyl Alcohols. Enzyme Microb. Technol. 2005, 37, 556–562. [Google Scholar] [CrossRef]
- Kumar, R.S.S.; Vishwanath, K.S.; Singh, S.A.; Rao, A.G.A. Entrapment of α-Amylase in Alginate Beads: Single Step Protocol for Purification and Thermal Stabilization. Process Biochem. 2006, 41, 2282–2288. [Google Scholar] [CrossRef]
- Gertler, A.; Birk, Y. Purification and Characterization of a β-Amylase from Soya Beans. Biochem. J. 1965, 95, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Nirmala, M.; Muralikrishna, G. Three α-Amylases from Malted Finger Millet (Ragi, Eleusine Coracana, Indaf-15)—Purification and Partial Characterization. Phytochemistry 2003, 62, 21–30. [Google Scholar] [CrossRef]
- Okolo, B.N.; Ire, F.S.; Ezeogu, L.I.; Anyanwu, C.U.; Odibo, F.J. Purification and Some Properties of a Novel Raw Starch-Digesting Amylase from Aspergillus carbonarius. J. Sci. Food Agric. 2001, 81, 329–336. [Google Scholar] [CrossRef]
- Bush, D.S.; Sticher, L.; Van Huystee, R.; Wagner, D.; Jones, R.L. The Calcium Requirement for Stability and Enzymatic Activity of Two Isoforms of Barley Aleurone α-Amylase. J. Biol. Chem. 1989, 264, 19392–19398. [Google Scholar] [CrossRef]
- Xie, F.; Quan, S.; Liu, D.; Ma, H.; Li, F.; Zhou, F.; Chen, G. Purification and Characterization of a Novel α-Amylase from a Newly Isolated Bacillus methylotrophicus Strain P11-2. Process Biochem. 2014, 49, 47–53. [Google Scholar] [CrossRef]
- Arnesen, S.; Havn Eriksen, S.; Olsen, J.; Jensen, B. Increased Production of α-Amylase from Thermomyces lanuginosus by the Addition of Tween 80. Enzyme Microb. Technol. 1998, 23, 249–252. [Google Scholar] [CrossRef]
- Oberoi, R.; Beg, Q.K.; Puri, S.; Saxena, R.K.; Gupta, R. Characterization and Wash Performance Analysis of an SDS-Stable Alkaline Protease from a Bacillus sp. World J. Microbiol. Biotechnol. 2001, 17, 493–497. [Google Scholar] [CrossRef]
- Matsui, H.; Chiba, S.; Shimomura, T. Purification and Some Properties of Active β-Amylase in Rice. Agric. Biol. Chem. 1977, 41, 841–847. [Google Scholar] [CrossRef]
- Chia, T.; Thorneycroft, D.; Chapple, A.; Messerli, G.; Chen, J.; Zeeman, S.C.; Smith, S.M.; Smith, A.M. A Cytosolic Glucosyltransferase Is Required for Conversion of Starch to Sucrose in Arabidopsis Leaves at Night. Plant J. 2004, 37, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Biazus, J.P.M.; Souza, R.R.D.; Márquez, J.E.; Franco, T.T.; Santana, J.C.C.; Tambourgi, E.B. Production and Characterization of Amylases from Zea mays Malt. Braz. Arch. Biol. Technol. 2009, 52, 991–1000. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Vesterberg, O.; Svensson, H. Isoelectric Fractionation, Analysis, and Characterization of Ampholytes in Natural pH Gradients. Acta Chem. Scand. 1966, 20, 820–834. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Steps | Total Protein (mg) | Total Activity (Units) | Specific Activity (Units/mg) | Purification (Fold) | Yield (%) |
|---|---|---|---|---|---|
| Crude extract | 494 ± 52 | 890,100 ± 2540 | 1800 ± 20 | 1.0 | 100.0 |
| Ethanol precipitation | 73 ± 3 | 528,400 ± 360 | 7200 ± 30 | 4.0 | 59.4 |
| Hitrap QFF | 14 ± 1 | 297,100 ± 80 | 20,890 ± 560 | 11.6 | 33.4 |
| HiPrepTM 26/60 SephacrylTM S-100 HR | 5 ± 0 | 154,200 ± 160 | 29,700 ± 110 | 16.5 | 17.3 |
| Reagent | Concentration (mM) | Relative Activity (%) |
|---|---|---|
| None | - | 100 |
| MnCl2 | 10 | 93.43 ± 4.07 |
| FeCl3 | 10 | 91.12 ± 2.12 |
| MgCl2 | 10 | 104.90 ± 2.67 |
| LiCl2 | 10 | 106.57 ± 2.77 |
| BaCl2 | 10 | 96.58 ± 3.03 |
| CuSO4 | 10 | 38.40 ± 2.79 |
| CaCl2 | 10 | 96.58 ± 3.03 |
| KCl | 10 | 101.76 ± 6.24 |
| ZnCl2 | 10 | 106.75 ± 4.29 |
| NaCl | 10 | 100.74 ± 3.46 |
| Reagent | Concentration (%) | Relative Activity (%) |
|---|---|---|
| None | - | 100 |
| EDTA | 10 | 91.33 ± 4.32 |
| PMSF | 10 | 152.14 ± 3.30 |
| β-mercaptoethanol | 10 | 11.71 ± 1.39 |
| Tween-20 | 2 | 158.45 ± 0.86 |
| Tween-40 | 2 | 147.99 ± 2.20 |
| Tween-60 | 2 | 150.54 ± 5.17 |
| Tween-80 | 2 | 149.84 ± 1.63 |
| TritonX-100 | 2 | 158.13 ± 3.11 |
| SDS | 2 | 20.49 ± 0.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, B.; Jin, C.; Guan, Y.; Chang, Z.; Liu, Q.; Gao, H. Purification and Characteristics of β-Amylase from Soybean Whey Wastewater. Catalysts 2024, 14, 909. https://doi.org/10.3390/catal14120909
Yao B, Jin C, Guan Y, Chang Z, Liu Q, Gao H. Purification and Characteristics of β-Amylase from Soybean Whey Wastewater. Catalysts. 2024; 14(12):909. https://doi.org/10.3390/catal14120909
Chicago/Turabian StyleYao, Bowei, Congli Jin, Yanyan Guan, Zhongyi Chang, Qian Liu, and Hongliang Gao. 2024. "Purification and Characteristics of β-Amylase from Soybean Whey Wastewater" Catalysts 14, no. 12: 909. https://doi.org/10.3390/catal14120909
APA StyleYao, B., Jin, C., Guan, Y., Chang, Z., Liu, Q., & Gao, H. (2024). Purification and Characteristics of β-Amylase from Soybean Whey Wastewater. Catalysts, 14(12), 909. https://doi.org/10.3390/catal14120909