
Efficient Synthesis of γ-Aminobutyric Acid from Monosodium Glutamate Using an Engineered Glutamate Decarboxylase Active at a Neutral pH

 and
and 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
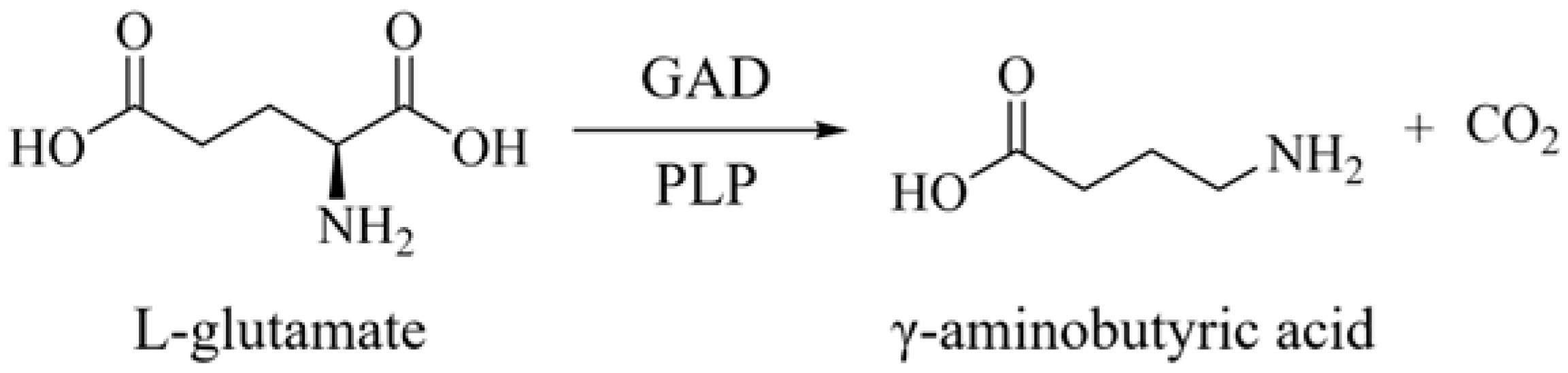
1. Introduction
2. Results
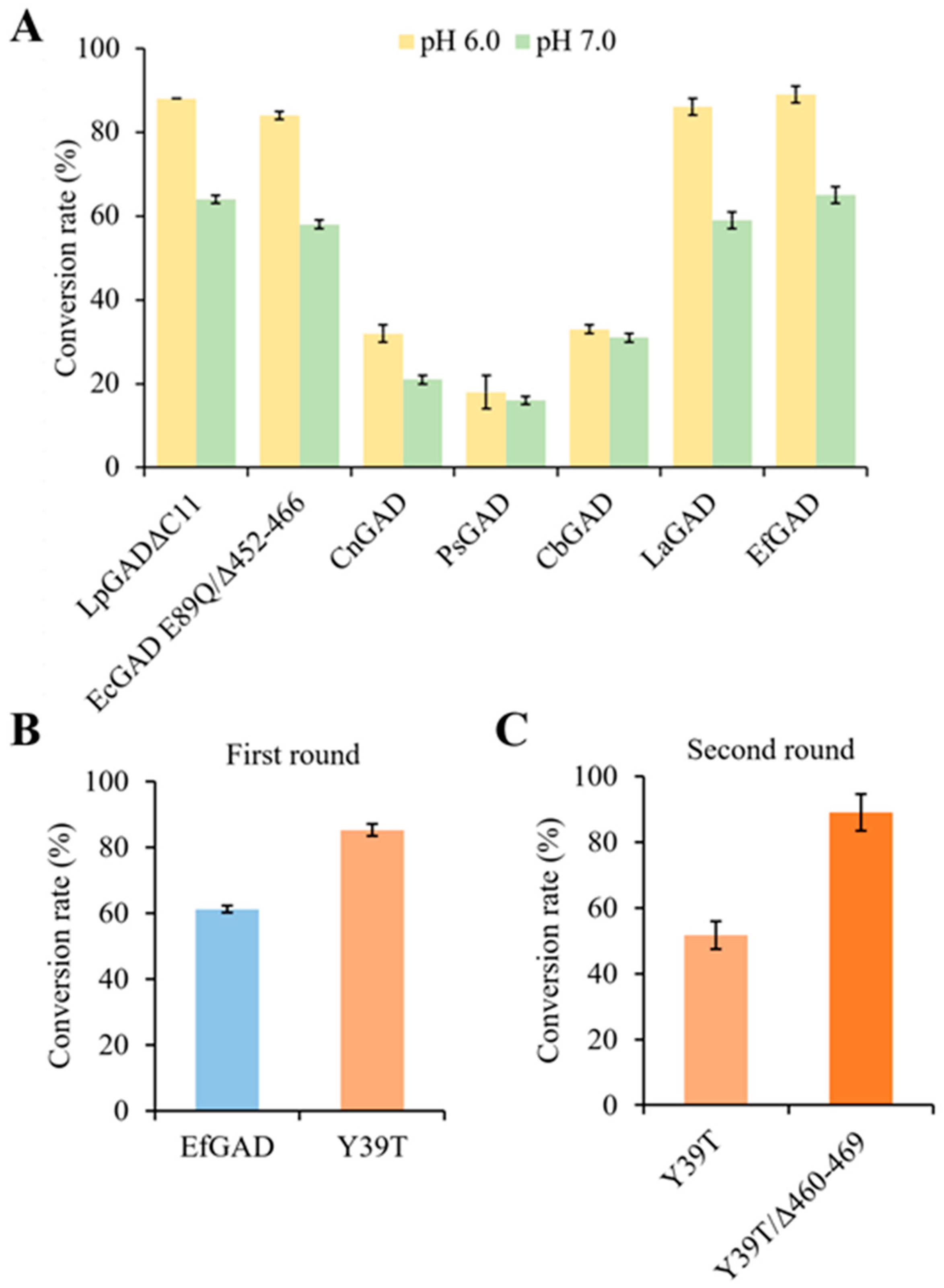
2.1. Screening Natural GADs with High Activity at a Neutral pH
2.2. EfGAD Engineering for Improved Catalytic Performance at a Neutral pH
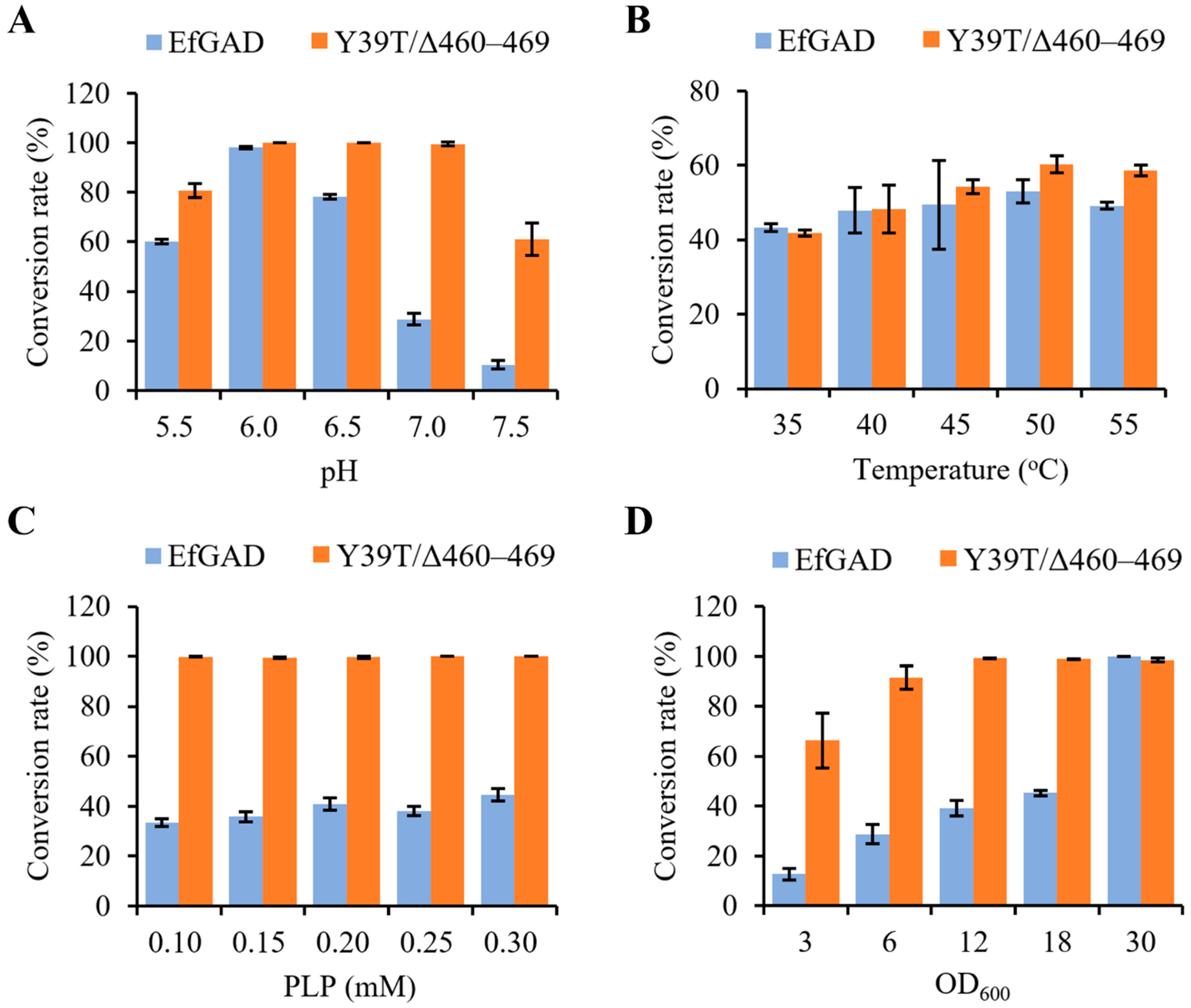
2.3. Effect of Reaction Conditions on MSG Bioconversion Catalyzed by EfGAD and Its Mutant
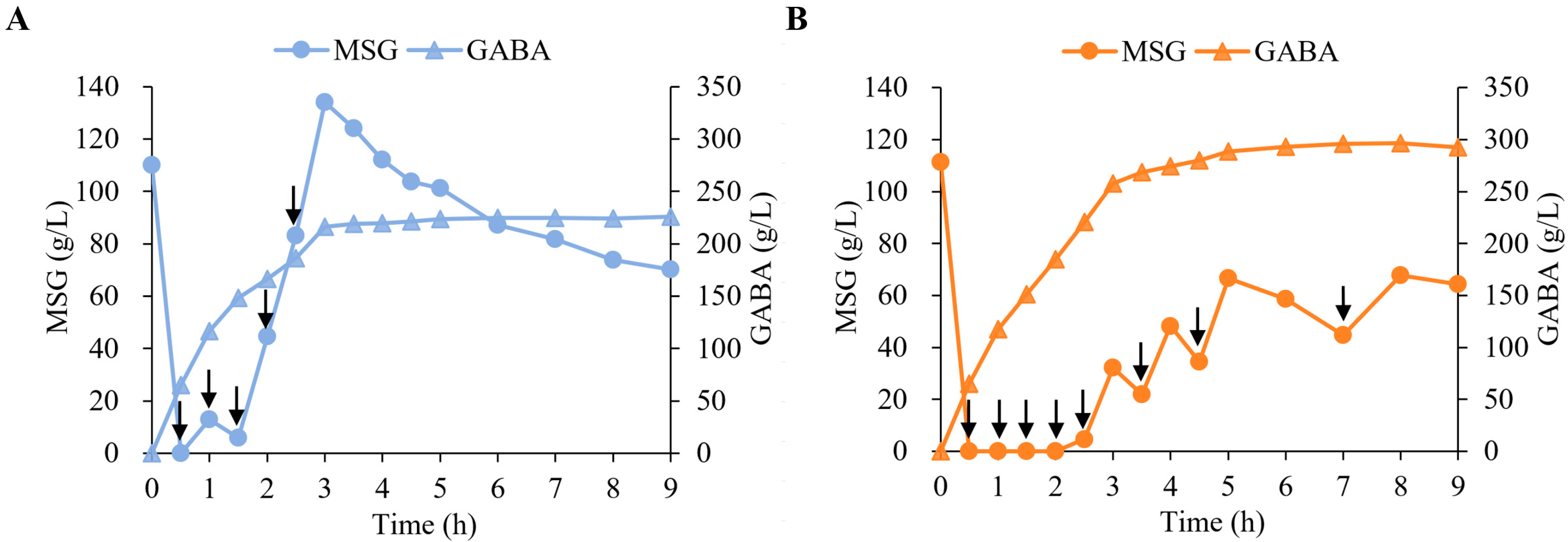
2.4. Evaluation of EfGAD and Y39T/Δ460–469 for GABA Production in 5 L Fermenters
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, and Chemicals
4.2. Cloning Glutamate Decarboxylase-Encoding Gene
4.3. ZS Prediction-Guided Engineering of EfGAD
4.4. Expression of GADs
4.5. Conversion of MSG to GABA in Tubes Catalyzed by GADs
4.6. Effect of Reaction Conditions on the Catalytic Performance of GADs
4.7. Whole-Cell Biotransformation in 5 L Fermenters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yuan, H.; Wang, H.; Fidan, O.; Qin, Y.; Xiao, G.; Zhan, J. Identification of new glutamate decarboxylases from Streptomyces for efficient production of γ-aminobutyric acid in engineered Escherichia coli. J. Med. Biol. Eng. 2019, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Park, Y.-L.; Choi, T.-R.; Kim, H.J.; Song, H.-S.; Han, Y.-H.; Lee, S.M.; Park, S.L.; Lee, H.S.; Bhatia, S.K.; et al. Production of γ-aminobutyric acid from monosodium glutamate using Escherichia coli whole-cell biocatalysis with glutamate decarboxylase from Lactobacillus brevis KCTC 3498. Korean J. Chem. Eng. 2020, 37, 2225–2231. [Google Scholar] [CrossRef]
- Yarabbi, H.; Mortazavi, S.A.; Yavarmanesh, M.; Javadmanesh, A. Molecular cloning, gene overexpression and characterization of glutamate decarboxylase from Enterococcus faecium DO. LWT 2021, 148, 111699. [Google Scholar] [CrossRef]
- Lyu, C.; Yao, L.; Zhu, Q.; Mei, J.; Cao, Y.; Hu, S.; Zhao, W.; Huang, J.; Mei, L.; Yao, S.; et al. Reconstruction of the glutamate decarboxylase system in Lactococcus lactis for biosynthesis of food-grade γ-aminobutyric acid. Appl. Microbiol. Biotechnol. 2021, 105, 4127–4140. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.-D.; Li, X.; Shi, H.-L.; Jia, Y.-Y.; Dong, Z.-X.; Jiao, Z.-J.; Wang, L.-F.; Xu, J.-H.; Yao, L.-G.; Kan, Y.-C. Efficient expression of novel glutamate decarboxylases and high level production of γ-aminobutyric acid catalyzed by engineered Escherichia coli. Int. J. Biol. Macromol. 2020, 160, 372–379. [Google Scholar] [CrossRef]
- Choi, J.W.; Yim, S.S.; Lee, S.H.; Kang, T.J.; Park, S.J.; Jeong, K.J. Enhanced production of gamma-aminobutyrate (GABA) in recombinant Corynebacterium glutamicum by expressing glutamate decarboxylase active in expanded pH range. Microb. Cell Factories. 2015, 14, 21. [Google Scholar] [CrossRef]
- Yang, H.; Xing, R.; Hu, L.; Liu, S.; Li, P. Accumulation of γ-aminobutyric acid by Enterococcus avium 9184 in scallop solution in a two-stage fermentation strategy. Microb. Biotechnol. 2015, 9, 478–485. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, L.; Gao, Q.; Yu, S.M.; Li, L.; Gao, N.F. The two-step biotransformation of monosodium glutamate to GABA by Lactobacillus brevis growing and resting cells. Appl. Microbiol. Biotechnol. 2012, 94, 1619–1627. [Google Scholar] [CrossRef]
- Wen, J.; Sun, W.; Leng, G.; Li, D.; Feng, C.; Tian, Z.; Wang, X. Enhanced fermentative γ-aminobutyric acid production by a metabolic engineered Corynebacterium glutamicum. Biotechnol. Bioprocess Eng. 2024, 29, 129–140. [Google Scholar] [CrossRef]
- Thu Ho, N.A.; Hou, C.Y.; Kim, W.H.; Kang, T.J. Expanding the active pH range of Escherichia coli glutamate decarboxylase by breaking the cooperativeness. J. Biosci. Bioeng. 2013, 115, 154–158. [Google Scholar] [CrossRef]
- Song, Z.; Zhang, Q.; Wu, W.; Pu, Z.; Yu, H. Rational design of enzyme activity and enantioselectivity. Front. Bioeng. Biotechnol. 2023, 11, 1129149. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Liu, B.; Shen, Y.; Jing, K.; Savage, T.R. Protein engineering design from directed evolution to de novo synthesis. Biochem. Eng. J. 2021, 174, 108096. [Google Scholar] [CrossRef]
- Qu, G.; Li, A.; Acevedo-Rocha, C.G.; Sun, Z.; Reetz, M.T. The crucial role of methodology development in directed evolution of selective enzymes. Angew. Chem. Int. Ed. 2020, 59, 13204–13231. [Google Scholar] [CrossRef] [PubMed]
- Siedhoff, N.E.; Schwaneberg, U.; Davari, M.D. Machine learning-assisted enzyme engineering. Meth. Enzymol. 2020, 643, 281–315. [Google Scholar]
- Kouba, P.; Kohout, P.; Haddadi, F.; Bushuiev, A.; Samusevich, R.; Sedlar, J.; Damborsky, J.; Pluskal, T.; Sivic, J.; Mazurenko, S. Machine learning-guided protein engineering. ACS Catal. 2023, 13, 13863–13895. [Google Scholar] [CrossRef]
- Yang, K.K.; Wu, Z.; Arnold, F.H. Machine-learning-guided directed evolution for protein engineering. Nat. Methods 2019, 16, 687–694. [Google Scholar] [CrossRef]
- Kang, L.; Tan, P.; Hong, L. Enzyme engineering in the age of artificial intelligence. ACS Synth. Biol. 2023, 4, 524–534. [Google Scholar]
- Wittmann, B.J.; Johnston, K.E.; Wu, Z.; Arnold, F.H. Advances in machine learning for directed evolution. Curr. Opin. Struct. Biol. 2021, 69, 11–18. [Google Scholar] [CrossRef]
- Wittmann, B.J.; Yue, Y.; Arnold, F.H. Informed training set design enables efficient machine learning-assisted directed protein evolution. Cell Syst. 2021, 12, 1026–1045. [Google Scholar] [CrossRef]
- Meier, J.; Rao, R.; Verkuil, R.; Liu, J.; Sercu, T.; Rives, A. Language models enable zero-shot prediction of the effects of mutations on protein function. Adv. Neural inf. Process. Syst. 2021, 34, 29287–29303. [Google Scholar]
- Mansoor, S.; Baek, M.; Juergens, D.; Watson, J.L.; Baker, D. Zero-shot mutation effect prediction on protein stability and function using RoseTTAFold. Protein Sci. 2023, 32, e4780. [Google Scholar] [CrossRef]
- Cheng, P.; Mao, C.; Tang, J.; Yang, S.; Cheng, Y.; Wang, W.; Gu, Q.; Han, W.; Chen, H.; Li, S.; et al. Zero-shot prediction of mutation effects with multimodal deep representation learning guides protein engineering. Cell Res. 2024, 34, 630–647. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Deng, H.; He, J.; Keasling, J.D.; Luo, X. UniKP: A unified framework for the prediction of enzyme kinetic parameters. Nat. Commun. 2023, 14, 8211. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yuan, L.; Lu, H.; Li, G.; Chen, Y.; Engqvist, M.K.M.; Kerkhoven, E.J.; Nielsen, J. Deep learning-based kcat prediction enables improved enzyme-constrained model reconstruction. Nat. Catal. 2022, 5, 662–672. [Google Scholar] [CrossRef]
- Sun, J.; Zhu, T.; Cui, Y.; Wu, B. Structure-based self-supervised learning enables ultrafast prediction of stability changes upon mutation at the protein universe scale. bioRxiv 2023. [Google Scholar] [CrossRef]
- Yang, L.; Liang, X.; Zhang, N.; Lu, L. STAR: A web server for assisting directed protein evolution with machine learning. ACS Omega 2023, 8, 44751–44756. [Google Scholar] [CrossRef]
- Firnberg, E.; Labonte, J.W.; Gray, J.J.; Ostermeier, M. A comprehensive, high-resolution map of a gene’s fitness landscape. Mol. Biol. Evol. 2014, 31, 1581–1592. [Google Scholar] [CrossRef]
- Romero, P.A.; Arnold, F.H. Exploring protein fitness landscapes by directed evolution. Nat. Rev. Mol. Cell Biol. 2009, 10, 866–876. [Google Scholar] [CrossRef]
- Yu, K.; Lin, L.; Hu, S.; Huang, J.; Mei, L. C-terminal truncation of glutamate decarboxylase from Lactobacillus brevis CGMCC 1306 extends its activity toward near-neutral pH. Enzyme Microb. Technol. 2012, 50, 263–269. [Google Scholar] [CrossRef]
- Shin, S.-M.; Kim, H.; Joo, Y.; Lee, S.-J.; Lee, Y.-J.; Lee, S.J.; Lee, D.-W. Characterization of glutamate decarboxylase from Lactobacillus plantarum and its C-terminal function for the pH dependence of activity. J. Agric. Food Chem. 2014, 62, 12186–12193. [Google Scholar] [CrossRef]
- Capitani, G.; Biase, D.D.; Aurizi, C.; Gut, H.; Bossa, F.; Grütter, M.G. Crystal structure and functional analysis of Escherichia coli glutamate decarboxylase. EMBO J. 2003, 22, 4027–4037. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, L.; Zheng, Y.; Cheng, R.; Liu, J.; Zheng, Z.; Yang, H.; Zhao, J. Efficient Synthesis of γ-Aminobutyric Acid from Monosodium Glutamate Using an Engineered Glutamate Decarboxylase Active at a Neutral pH. Catalysts 2024, 14, 905. https://doi.org/10.3390/catal14120905
Miao L, Zheng Y, Cheng R, Liu J, Zheng Z, Yang H, Zhao J. Efficient Synthesis of γ-Aminobutyric Acid from Monosodium Glutamate Using an Engineered Glutamate Decarboxylase Active at a Neutral pH. Catalysts. 2024; 14(12):905. https://doi.org/10.3390/catal14120905
Chicago/Turabian StyleMiao, Lulu, Yingjia Zheng, Runxi Cheng, Jianzhong Liu, Zhinan Zheng, Hong Yang, and Jing Zhao. 2024. "Efficient Synthesis of γ-Aminobutyric Acid from Monosodium Glutamate Using an Engineered Glutamate Decarboxylase Active at a Neutral pH" Catalysts 14, no. 12: 905. https://doi.org/10.3390/catal14120905
APA StyleMiao, L., Zheng, Y., Cheng, R., Liu, J., Zheng, Z., Yang, H., & Zhao, J. (2024). Efficient Synthesis of γ-Aminobutyric Acid from Monosodium Glutamate Using an Engineered Glutamate Decarboxylase Active at a Neutral pH. Catalysts, 14(12), 905. https://doi.org/10.3390/catal14120905