
State of the Art on the Microbial Production of Industrially Relevant Organic Acids

Abstract
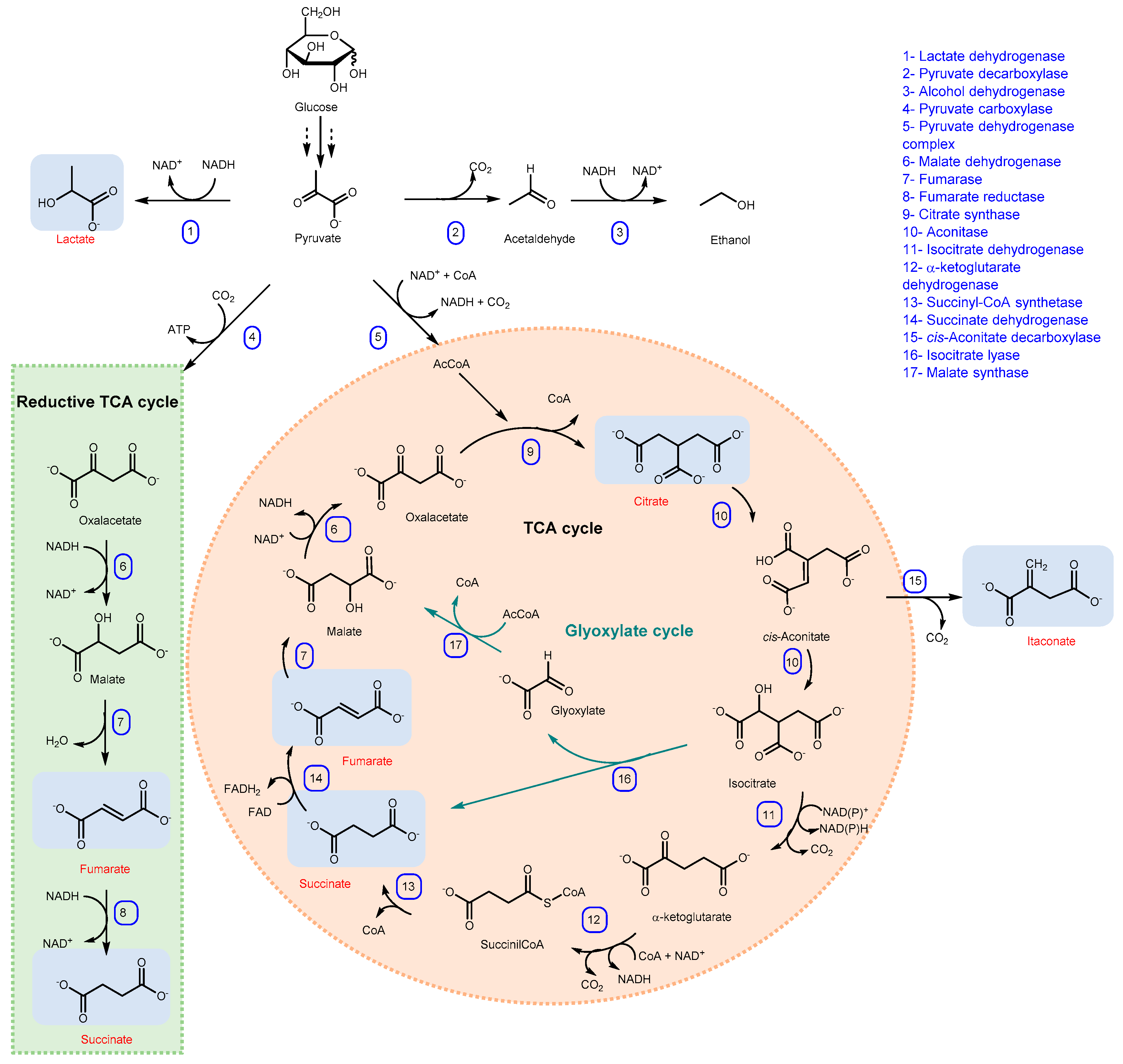
1. Introduction
2. From First Evidence and Studies to Current Pilot or Commercial-Scale Production
2.1. Citric Acid (C6H8O7, 192.124 g·mol−1, pKa1 3.13, pKa2 4.76, pKa3 6.39)

{kind=link}
{kind=link}
| Microorganism | Substrate | Citric Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| A. niger MH 15-15 | Glucose | 31 | Shake flask | [38] |
| A. niger MH 15-15 | Sucrose | 53 | Shake flask | [38] |
| A. niger | Corn stover | 136.3 | Simultaneous saccharification and fermentation | [39] |
| A. niger GMCC 5751 | Liquefied corn | 151.7 | Batch | [40] |
| Adapted A. niger | Corn powder and citric acid effluent | 157.3 | Batch | [41] |
| Mutant A. niger MO-25 | Beet molasses and chicken feather peptone | 68.8 | Shake flask | [28] |
| Mutant A. niger UMIP 2564 | Sucrose | 96.3 | Shake flask | [29] |
| Mutant A. niger UMIP 2564 | Beet molasses | 98.3 | Shake flask | [29] |
| Mutant A. niger UMIP 2564 | Cane molasses | 91.2 | Shake flask | [29] |
| A. niger 831f | Liquefied cassava | 162.7 | Repeat-fed batch | [42] |
| Y. lipolytica NRRL YB-423 | Glycerol | 21.6 | Shake flask | [43] |
2.2. Fumaric Acid (C4H4O4, 116.07 g·mol−1, pKa1 3.03, pKa2 4.44)
2.3. Itaconic Acid (C5H6O4, 130.10 g·mol−1, pKa1 3.85, pKa2 5.45)
| Microorganism | Substrate | Itaconic Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| A. terreus DSM 23081 | Glucose | 160 | Fed-batch | [107] |
| A. terreus NRRL 1960 | d-Xylose | 53.97 | Batch | [119] |
| A. terreus NRRL 1960 | Bleached eucalyptus pulp | 37.5 | Shake flask | [120] |
| U. maydis MB215 | Glucose | 44.5 | Batch | [109] |
| Mutant U. maydis MB215 | Glucose | 80 | Fed-batch | [121] |
| Mutant U. maydis MB215 | Glucose | 74.9 | Continuous feeding | [117] |
| U. vetiveriae TZ1 | Glycerol | 34.7 | Batch | [122] |
| A. terreus CICC40205 | Wheat bran hydrolysate | 34.2 | Shake flask | [123] |
| A. terreus M69 | Corn stover hydrolysate | 33.6 | Shake flask | [124] |
2.4. Lactic Acid (C3H6O3, 90.07948 g·mol−1, pKa1 3.78)
| Microorganism | Substrate | Lactic Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| Mutant P. kudriavzevii | Glucose | 154 (d-Lactic acid) | Fed-batch | [153] |
| Evolved L. delbrueckii S-NL31 | Soybean meal | 112.3 (d-Lactic acid) | Fed-batch | [161] |
| Evolved and mutated S. cerevisiae | Glucose | 82.6 (d-Lactic acid) | Fed-batch | [162] |
| Mutant S. cerevisiae | Glucose | 40 (d-Lactic acid) | Fed-batch | [163] |
| Mutant E. coli | Glycerol | 115 (d-Lactic acid) | Fed-batch | [164] |
| L. delbrueckii ssp. bulgaricus | Orange peel waste | 45 (d-Lactic acid) | Separate hydrolysis and fermentation | [165] |
| L. delbrueckii | Molasses and corn steep liquor | 162 (d-Lactic acid) | Fed-batch | [166] |
| B. coagulans A107 | Defatted rice bran (DRB) | 75.9 (l-Lactic acid) | Batch | [137] |
| L. brevis ATCC 367 and L. plantarum ATCC 21028 | Corn stover | 24 (l-Lactic acid) 31.2 (l-Lactic acid) | Simultaneous sequential | [167] |
| L. casei | Sugarcane molasses | 120.23 (NS) | Batch | [168] |
| Mutant S. cerevisiae | Glucose | 142 (l-Lactic acid) | Fed-batch | [169] |
| R. oryzae LA-UN-1 | Glucose | 162 (l-Lactic acid) | Fed-batch | [170] |
| L. rhamnosus DUT1908 | Starchy biomass | 108 (l-Lactic acid) | One step liquefaction, saccharification and fermentation | [171] |
| Mutated and evolved L. paracasei NCBIO01 | Glucose | 202 (l-Lactic acid) 9.06 (d-Lactic acid) | Open fermentation | [172] |
| B. coagulans and L. rhamnosus | Cassava bagasse | 113 (l-Lactic acid) | Simultaneous saccharification and co-fermentation | [173] |
| Mutant L. plantarum NCIMB 8826 | Raw corn starch | 50 (l-Lactic acid) | Batch | [174] |
| L. paracasei subsp. paracasei2 | Food waste | 34 (l-Lactic acid) | Batch | [175] |
2.5. Succinic Acid (C4H6O4, 118.09 g·mol−1, pKa1 4.2, pKa2 5.6)
| Microorganism | Substrate | Succinic Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| Isolate AKR177 | Pure glycerol | 117 | Fed-batch | [190] |
| Isolate AKR177 | Crude glycerol | 86.9 | Fed-batch | [190] |
| A. succinogenes CCTCC M2012036 | Sugarcane bagasse | 39.9 70.81 | Batch Fed-batch | [191] |
| Mutant E. coli MH28 | Glycerol | 84.3 | Batch | [98] |
| Mutant S. cerevisiae SUC-297 | Glucose | 43 | Fed-batch | [185] |
| Mutant B. succiniciproducens LU15224 | Glycerol and maltose | 69.8 | Batch | [192] |
| Mutant Y. lipolytica | Crude Glycerol | 209.7 | Fed-batch | [193] |
| Mutant E. coli | Glucose | 99.2 | Dual-phase | [188] |
| Mutant Y. lipolytica | Glucose | 101.4 | Fed-batch | [194] |
| Mutant Y. lipolytica | Glycerol | 110.7 | Fed-batch | [195] |
| Evolved Y. lipolytica | Mixed food waste | 71.6 | Fed-batch | [196] |
3. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Branduardi, P. Closing the loop: The power of microbial biotransformations from traditional bioprocesses to biorefineries, and beyond. Microb. Biotechnol. 2021, 14, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass Volume I—Results of Screening for Potential Candidates from Sugars and Synthesis Gas; National Renewable Energy Lab: Golden, CO, USA, 2004; Volume 2.
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—The US Department of Energy’s “top 10” revisited. Green Chem. 2010, 12, 539–555. [Google Scholar] [CrossRef]
- Becker, J.; Lange, A.; Fabarius, J.; Wittmann, C. Top value platform chemicals: Bio-based production of organic acids. Curr. Opin. Biotechnol. 2015, 36, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Song, C.W.; Shin, J.H.; Lee, S.Y. Biorefineries for the production of top building block chemicals and their derivatives. Metab. Eng. 2015, 28, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Research, G.V. Organic Acids Market Analysis, by Product Type (Formic Acid, Citric Acid, Others), by Source Type (Biomass, Chemical Synthesis, Others), by Application (Food & Beverages Industry, Pharmaceutical Industry, Others), Forecasts To 2028. Available online: https://www.reportsanddata.com/report-detail/organic-acids-market (accessed on 14 December 2021).
- Van Dien, S. From the first drop to the first truckload: Commercialization of microbial processes for renewable chemicals. Curr. Opin. Biotechnol. 2013, 24, 1061–1068. [Google Scholar] [CrossRef]
- Wehrs, M.; Tanjore, D.; Eng, T.; Lievense, J.; Pray, T.R.; Mukhopadhyay, A. Engineering robust production microbes for large-scale cultivation. Trends Microbiol. 2019, 27, 524–537. [Google Scholar] [CrossRef]
- Scheele, C.W. Sammtliche Physische and Chemischewerke; Mayer & Müller: Berlin, Germany, 1784; Volume 2. [Google Scholar]
- Papagianni, M. Advances in citric acid fermentation by Aspergillus niger: Biochemical aspects, membrane transport and modeling. Biotechnol. Adv. 2007, 25, 244–263. [Google Scholar] [CrossRef]
- Grimoux, E.; Adams, P. Synthese de lácide citrique. C. R. Hebd. Seances Acad. Sci. 1880, 90, 1252. [Google Scholar]
- Wehmer, C.F.W. Beiträge zur Kenntnis Einheimischer Pilze; Hahn’sche Buchhandlung: Hannover/Jena, Germany, 1893. [Google Scholar]
- Currie, J.N. The citric acid fermentation of Aspergillus niger. J. Biol. Chem. 1917, 31, 15–37. [Google Scholar] [CrossRef]
- Apelblat, A. Citric Acid; Springer International Publishing: Cham, Switzerland, 2014; ISBN 978-3-319-11232-9. [Google Scholar]
- Judah, G. Pfizer’s Work on Penicillin for World War II becomes a National Historic Chemical Landmark. Available online: https://www.acs.org/content/acs/en/pressroom/newsreleases/2008/june/pfizers-work-on-penicillin-for-world-war-ii-becomes-a-national-historic-chemical-landmark.html (accessed on 13 December 2021).
- Mores, S.; de Souza Vandenberghe, L.P.; Júnior AI, M.; de Carvalho, J.C.; de Mello AF, M.; Pandey, A.; Soccol, C.R. Citric acid bioproduction and downstream processing: Status, opportunities, and challenges. Bioresour. Technol. 2021, 320, 124426. [Google Scholar] [CrossRef]
- Ciriminna, R.; Meneguzzo, F.; Delisi, R.; Pagliaro, M. Citric acid: Emerging applications of key biotechnology industrial product. Chem. Cent. J. 2017, 11, 1–9. [Google Scholar] [CrossRef]
- Savi, L.K.; Dias, M.C.G.C.; Carpine, D.; Waszczynskyj, N.; Ribani, R.H.; Haminiuk, C.W.I. Natural deep eutectic solvents (NADES) based on citric acid and sucrose as a potential green technology: A comprehensive study of water inclusion and its effect on thermal, physical and rheological properties. Int. J. Food Sci. Technol. 2019, 54, 898–907. [Google Scholar] [CrossRef]
- Amato, A.; Becci, A.; Beolchini, F. Citric acid bioproduction: The technological innovation change. Crit. Rev. Biotechnol. 2020, 40, 199–212. [Google Scholar] [CrossRef]
- Behera, B.C.; Mishra, R.; Mohapatra, S. Microbial citric acid: Production, properties, application, and future perspectives. Food Front. 2021, 2, 62–76. [Google Scholar] [CrossRef]
- Erdem, C.; Papanikolaou, S.; Fickers, P.; Agirman, B.; Erten, H. Citric Acid Production by Yarrowia lipolytica. In Non-Conventional Yeasts: From Basic Research to Application; Sibirny, A., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 91–117. [Google Scholar]
- Yang, L.; Lübeck, M.; Lübeck, P.S. Aspergillus as a versatile cell factory for organic acid production. Fungal Biol. Rev. 2017, 31, 33–49. [Google Scholar] [CrossRef]
- Perlman, D. Mycological production of citric acid: The submerged culture method. Econ. Bot. 1949, 3, 360. [Google Scholar] [CrossRef]
- Gil, R.G.; Correa, H.S.; Contreras Larios, J.L.; González-Brambila, M.M. A biotechnological process for obtaining citric acid through paper cellulose aerobic bioreaction. Int. J. Chem. React. Eng. 2020, 18, 1–10. [Google Scholar] [CrossRef]
- Roukas, T.; Kotzekidou, P. Pomegranate peel waste: A new substrate for citric acid production by Aspergillus niger in solid-state fermentation under non-aseptic conditions. Environ. Sci. Pollut. Res. 2020, 27, 13105–13113. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, E.; Kontogiannopoulos, K.N.; Assimopoulou, A.N.; Mantzouridou, F.T. Feasibility of multi-hydrolytic enzymes production from optimized grape pomace residues and wheat bran mixture using Aspergillus niger in an integrated citric acid-enzymes production process. Bioresour. Technol. 2020, 309, 123317. [Google Scholar] [CrossRef]
- Papadaki, E.; Mantzouridou, F.T. Citric acid production from the integration of spanish-style green olive processing wastewaters with white grape pomace by Aspergillus niger. Bioresour. Technol. 2019, 280, 59–69. [Google Scholar] [CrossRef]
- Ozdal, M.; Kurbanoglu, E.B. Citric acid production by Aspergillus niger from agro-industrial by-products: Molasses and chicken feather peptone. Waste Biomass Valorization 2019, 10, 631–640. [Google Scholar] [CrossRef]
- Lotfy, W.A.; Ghanem, K.M.; El-Helow, E.R. Citric acid production by a novel Aspergillus niger isolate: I. Mutagenesis and cost reduction studies. Bioresour. Technol. 2007, 98, 3464–3469. [Google Scholar] [CrossRef] [PubMed]
- Adeoye, A.O.; Lateef, A.; Gueguim-Kana, E.B. Optimization of citric acid production using a mutant strain of Aspergillus niger on cassava peel substrate. Biocatal. Agric. Biotechnol. 2015, 4, 568–574. [Google Scholar] [CrossRef]
- Pel, H.J.; De Winde, J.H.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; De Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cao, Z.; Hou, L.; Yin, L.; Wang, D.; Gao, Q.; Wu, Z.; Wang, D. The opposite roles of agdA and glaA on citric acid production in Aspergillus niger. Appl. Microbiol. Biotechnol. 2016, 100, 5791–5803. [Google Scholar] [CrossRef]
- Xue, X.; Bi, F.; Liu, B.; Li, J.; Zhang, L.; Zhang, J.; Gao, Q.; Wang, D. Improving citric acid production of an industrial Aspergillus niger CGMCC 10142: Identification and overexpression of a high-affinity glucose transporter with different promoters. Microb. Cell Fact. 2021, 20, 1–13. [Google Scholar] [CrossRef]
- Steiger, M.G.; Rassinger, A.; Mattanovich, D.; Sauer, M. Engineering of the citrate exporter protein enables high citric acid production in Aspergillus niger. Metab. Eng. 2019, 52, 224–231. [Google Scholar] [CrossRef]
- Sun, X.; Wu, H.; Zhao, G.; Li, Z.; Wu, X.; Liu, H.; Zheng, Z. Morphological regulation of Aspergillus niger to improve citric acid production by chsC gene silencing. Bioprocess Biosyst. Eng. 2018, 41, 1029–1038. [Google Scholar] [CrossRef]
- Hou, L.; Liu, L.; Zhang, H.; Zhang, L.; Zhang, L.; Zhang, J.; Gao, Q.; Wang, D. Functional analysis of the mitochondrial alternative oxidase gene (aox1) from Aspergillus niger CGMCC 10142 and its effects on citric acid production. Appl. Microbiol. Biotechnol. 2018, 102, 7981–7995. [Google Scholar] [CrossRef] [PubMed]
- De Jongh, W.A.; Nielsen, J. Enhanced citrate production through gene insertion in Aspergillus niger. Metab. Eng. 2008, 10, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Brooks, J.D.; Maddox, I.S. The effect of the sugar source on citric acid production by Aspergillus niger. Appl. Microbiol. Biotechnol. 1984, 19, 393–397. [Google Scholar] [CrossRef]
- Hou, W.; Bao, J. Simultaneous saccharification and aerobic fermentation of high titer cellulosic citric acid by filamentous fungus Aspergillus niger. Bioresour. Technol. 2018, 253, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, J.; Cao, Z.; Wang, Y.; Gao, Q.; Zhang, J.; Wang, D. Inhibition of oxidative phosphorylation for enhancing citric acid production by Aspergillus niger. Microb. Cell Fact. 2015, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhao, S.; Tan, F.; Li, H.; Chu, R.; Wang, X.; Sun, H.; Zhang, M. A novel green production process of citric acid on the pilot scale by directly recycling its extraction effluent. J. Clean. Prod. 2020, 277, 124068. [Google Scholar] [CrossRef]
- Yu, B.; Zhang, X.; Sun, W.; Xi, X.; Zhao, N.; Huang, Z.; Ying, Z.; Liu, L.; Liu, D.; Niu, H.; et al. Continuous citric acid production in repeated-fed batch fermentation by Aspergillus niger immobilized on a new porous foam. J. Biotechnol. 2018, 276–277, 1–9. [Google Scholar] [CrossRef]
- Levinson, W.E.; Kurtzman, C.P.; Kuo, T.M. Characterization of Yarrowia lipolytica and related species for citric acid production from glycerol. Enzyme Microb. Technol. 2007, 41, 292–295. [Google Scholar] [CrossRef]
- Vandenberghe, L.P.S.; Rodrigues, C.; de Carvalho, J.C.; Medeiros, A.B.P.; Soccol, C.R. Production and application of citric acid. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2017; pp. 557–575. ISBN 9780444636621. [Google Scholar]
- Vandenberghe, L.P.S.; Soccol, C.R.; Prado, F.C.; Pandey, A. Comparison of citric acid production by solid-state fermentation in flask, column, tray, and drum bioreactors. Appl. Biochem. Biotechnol. 2004, 118, 293–304. [Google Scholar] [CrossRef]
- Straathof, A.J.J. The proportion of downstream costs in fermentative production processes. In Comprehensive Biotechnology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 1, pp. 811–814. [Google Scholar]
- Kertes, A.S.; King, C.J. Extraction chemistry of fermentation product carboxylic acids. Biotechnol. Bioeng. 2009, 103, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Araújo, E.M.R.; Coelho, F.E.B.; Balarini, J.C.; Miranda, T.L.S.; Salum, A. Solvent extraction of citric acid with different organic phases. Adv. Chem. Eng. Sci. 2017, 7, 304–324. [Google Scholar] [CrossRef]
- Djas, M.; Henczka, M. Reactive extraction of citric acid using supercritical carbon dioxide. J. Supercrit. Fluids 2016, 117, 59–63. [Google Scholar] [CrossRef]
- Pourcelly, G. Citric acid recovery by electrodialysis. In Encyclopedia of Membranes; Drioli, E., Giorno, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Delgado Dobladez, J.A.; Águeda Maté, V.I.; Uribe Santos, D.L.; Torrellas, S.Á.; Larriba, M. Citric acid purification by simulated moving bed adsorption with methanol as desorbent. Sep. Sci. Technol. 2019, 54, 930–942. [Google Scholar] [CrossRef]
- Wang, J.; Cui, Z.; Li, Y.; Cao, L.; Lu, Z. Techno-economic analysis and environmental impact assessment of citric acid production through different recovery methods. J. Clean. Prod. 2020, 249, 119315. [Google Scholar] [CrossRef]
- Anastassiadis, S.; Morgunov, I.G.; Kamzolova, S.V.; Finogenova, T.V. Citric acid production patent review. Recent Pat. Biotechnol. 2008, 2, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Citric Acid Market: Global Industry Trends, Share, Size, Growth, Opportunity and Forecast 2021–2026. Available online: https://www.imarcgroup.com/citric-acid-manufacturing-plant (accessed on 29 January 2022).
- Ilica, R.A.; Kloetzer, L.; Galaction, A.I.; Caşcaval, D. Fumaric acid: Production and separation. Biotechnol. Lett. 2019, 41, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, N.; Cao, Y.; Jin, C.; Li, F.; Cai, C.; Yao, J. Effects of fumaric acid supplementation on methane production and rumen fermentation in goats fed diets varying in forage and concentrate particle size. J. Anim. Sci. Biotechnol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Reda, F.M.; Ismail, I.E.; Attia, A.I.; Fikry, A.M.; Khalifa, E.; Alagawany, M. Use of fumaric acid as a feed additive in quail’s nutrition: Its effect on growth rate, carcass, nutrient digestibility, digestive enzymes, blood metabolites, and intestinal microbiota. Poult. Sci. 2021, 100, 101493. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; Straathof, A.J.J.; Zijlmans, T.W.; Van Gulik, W.M.; Van Der Wielen, L.A.M. Fumaric acid production by fermentation. Appl. Microbiol. Biotechnol. 2008, 78, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Moharregh-Khiabani, D.; Linker, R.A.; Gold, R.; Stangel, M. Fumaric acid and its esters: An emerging treatment for multiple sclerosis. Curr. Neuropharmacol. 2009, 7, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Smith, D. Fumaric acid esters for psoriasis: A systematic review. Ir. J. Med. Sci. 2017, 186, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, S.; Iino, T.; Sato, S.; Nakahara, T.; Mukataka, S. Improvement of production rate and yield of fumaric acid from maleic acid by heat treatment of Pseudomonas alcaligenes strain XD-1. Biochem. Eng. J. 2003, 13, 7–13. [Google Scholar] [CrossRef]
- Goto, M.; Nara, T.; Tokumaru, I.; Fugono, N.; Uchida, Y.; Terasawa, M.; Yukawa, H. Method of Producing Fumaric Acid. U.S. Patent US5783428, 21 July 1998. [Google Scholar]
- Abe, A.; Oda, Y.; Asano, K.; Sone, T. Rhizopus delemar is the proper name for Rhizopus oryzae fumaric-malic acid producers. Mycologia 2007, 99, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Kenealy, W.; Zaady, E.; Du Preez, J.C. Biochemical aspects of fumaric acid accumulation by Rhizopus arrhizus. Appl. Environ. Microbiol. 1986, 52, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.T.; Zhang, K.; Zhang, B.; Huang, H. Fumaric Acid. Compr. Biotechnol. Second Ed. 2011, 3, 163–177. [Google Scholar] [CrossRef]
- Zhang, B.; Skory, C.D.; Yang, S.T. Metabolic engineering of Rhizopus oryzae: Effects of overexpressing pyc and pepc genes on fumaric acid biosynthesis from glucose. Metab. Eng. 2012, 14, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yang, S.T. Metabolic engineering of Rhizopus oryzae: Effects of overexpressing fumR gene on cell growth and fumaric acid biosynthesis from glucose. Process Biochem. 2012, 47, 2159–2165. [Google Scholar] [CrossRef]
- Song, C.W.; Kim, D.I.; Choi, S.; Jang, J.W.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of fumaric acid. Biotechnol. Bioeng. 2013, 110, 2025–2034. [Google Scholar] [CrossRef]
- Li, N.; Zhang, B.; Wang, Z.; Tang, Y.J.; Chen, T.; Zhao, X. Engineering Escherichia coli for fumaric acid production from glycerol. Bioresour. Technol. 2014, 174, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Chen, X.; Liu, L.; Jiang, L. Fumaric acid production in Saccharomyces cerevisiae by simultaneous use of oxidative and reductive routes. Bioresour. Technol. 2013, 148, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, P.; Liu, L. Modular optimization of multi-gene pathways for fumarate production. Metab. Eng. 2016, 33, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, J.; Song, W.; Zhang, L.; Wang, H.; Liu, L. Fumaric acid production by Torulopsis glabrata: Engineering the urea cycle and the purine nucleotide cycle. Biotechnol. Bioeng. 2015, 112, 156–167. [Google Scholar] [CrossRef]
- Sebastian, J.; Hegde, K.; Kumar, P.; Rouissi, T.; Brar, S.K. Bioproduction of fumaric acid: An insight into microbial strain improvement strategies. Crit. Rev. Biotechnol. 2019, 39, 817–834. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.B.; Ng, T.K. Fermentation Process for Carboxylic Acids. U.S. Patent US4877731A, 31 October 1989. [Google Scholar]
- Sebastian, J.; Rouissi, T.; Brar, S.K. Miscanthus sp. Perennial lignocellulosic biomass as feedstock for greener fumaric acid bioproduction. Ind. Crops Prod. 2022, 175, 114248. [Google Scholar] [CrossRef]
- Li, X.; Zhou, J.; Ouyang, S.; Ouyang, J.; Yong, Q. Fumaric acid production from alkali-pretreated corncob by fed-batch simultaneous saccharification and fermentation combined with separated hydrolysis and fermentation at high solids loading. Appl. Biochem. Biotechnol. 2017, 181, 573–583. [Google Scholar] [CrossRef]
- Das, R.K.; Brar, S.K.; Verma, M. A fermentative approach towards optimizing directed biosynthesis of fumaric acid by Rhizopus oryzae 1526 utilizing apple industry waste biomass. Fungal Biol. 2015, 119, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Ma, J.; Wang, M.; Wang, W.; Deng, L.; Nie, K.; Yue, X.; Wang, F.; Tan, T. Food waste fermentation to fumaric acid by Rhizopus arrhizus rh7-13. Appl. Biochem. Biotechnol. 2016, 180, 1524–1533. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, S.; Xu, Q.; Gao, M.; Huang, H. Production of fumaric acid by simultaneous saccharification and fermentation of starchy materials with 2-deoxyglucose-resistant mutant strains of Rhizopus oryzae. Bioresour. Technol. 2012, 107, 363–367. [Google Scholar] [CrossRef]
- Wu, X.; Liu, Q.; Deng, Y.; Chen, X.; Zheng, Z.; Jiang, S.; Li, X. Production of fumaric acid by bioconversion of corncob hydrolytes using an improved Rhizopus oryzae strain. Appl. Biochem. Biotechnol. 2018, 184, 553–569. [Google Scholar] [CrossRef]
- Moresi, M.; Parente, E.; Petruccioli, M.; Federici, F. Fumaric acid production from hydrolysates of starch-based substrates. J. Chem. Technol. Biotechnol. 1992, 54, 283–290. [Google Scholar] [CrossRef]
- Deng, F.; Aita, G.M. fumaric acid production by Rhizopus oryzae ATCC®® 20344tm from lignocellulosic syrup. Bioenergy Res. 2018, 11, 330–340. [Google Scholar] [CrossRef]
- Scholz, S.A.; Graves, I.; Minty, J.J.; Lin, X.N. Production of cellulosic organic acids via synthetic fungal consortia. Biotechnol. Bioeng. 2017, 115, 1096–1100. [Google Scholar] [CrossRef]
- Zhou, Y.; Nie, K.; Zhang, X.; Liu, S.; Wang, M.; Deng, L.; Wang, F.; Tan, T. Production of fumaric acid from biodiesel-derived crude glycerol by Rhizopus arrhizus. Bioresour. Technol. 2014, 163, 48–53. [Google Scholar] [CrossRef]
- Das, R.K.; Brar, S.K. Enhanced fumaric acid production from brewery wastewater and insight into the morphology of Rhizopus oryzae 1526. Appl. Biochem. Biotechnol. 2014, 172, 2974–2988. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Du, G.; Hua, Z.; Zhou, J.; Chen, J. Optimization of fumaric acid production by Rhizopus delemar based on the morphology formation. Bioresour. Technol. 2011, 102, 9345–9349. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, A.; Androutsopoulos, N.; Patsalou, M.; Koutinas, M.; Kopsahelis, N.; Machado De Castro, A.; Papanikolaou, S.; Koutinas, A.A. Biotechnological production of fumaric acid: The effect of morphology of Rhizopus arrhizus NRRL 2582. Fermentation 2017, 3, 33. [Google Scholar] [CrossRef]
- Zhang, K.; Yu, C.; Yang, S.T. Effects of soybean meal hydrolysate as the nitrogen source on seed culture morphology and fumaric acid production by Rhizopus oryzae. Process Biochem. 2015, 50, 173–179. [Google Scholar] [CrossRef]
- Zhou, Y.; Du, J.; Tsao, G.T. Mycelial pellet formation by Rhizopus oryzae ATCC 20344. Appl. Biochem. Biotechnol. 2000, 84, 779–789. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; Van Gulik, W.M.; Marang, L.; Van der Wielen, L.A.M.; Straathof, A.J.J. Development of a low pH fermentation strategy for fumaric acid production by Rhizopus oryzae. Enzyme Microb. Technol. 2011, 48, 39–47. [Google Scholar] [CrossRef]
- Das, R.K.; Brar, S.K.; Verma, M. effects of different metallic nanoparticles on germination and morphology of the fungus Rhizopus oryzae 1526 and changes in the production of fumaric acid. Bionanoscience 2015, 5, 217–226. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, S.; Jin, Y.; Yue, X.; Deng, L.; Wang, F.; Tan, T. Production of fumaric acid by immobilized Rhizopus arrhizus RH 7-13-9# on loofah fiber in a stirred-tank reactor. Bioresour. Technol. 2017, 244, 929–933. [Google Scholar] [CrossRef]
- Naude, A.; Nicol, W. Improved continuous fumaric acid production with immobilised Rhizopus oryzae by implementation of a revised nitrogen control strategy. N. Biotechnol. 2018, 44, 13–22. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, H.; Liu, X.; Zhu, H.; Fan, T.; Deng, L.; Wang, F. A high efficient method for simultaneous fermentation and separation of fumaric acid with a fixed bed ion exchange column. Biochem. Eng. J. 2020, 160, 107610. [Google Scholar] [CrossRef]
- Zhou, Y.; Du, J.; Tsao, G. Comparison of fumaric acid production by Rhizopus oryzae using different neutralizing agents. Bioprocess Biosyst. Eng. 2002, 25, 179–181. [Google Scholar] [CrossRef]
- Rhodes, R.A.; Lagoda, A.A.; Misenheimer, T.J.; Smith, M.L.; Anderson, R.F.; Jackson, R.W. production of fumaric acid in 20-liter fermentors. Appl. Microbiol. 1962, 10, 9–15. [Google Scholar] [CrossRef]
- Guettler, M.V.; Hanchar, R.J.; Rumler, D.S.; Kleff, S. Fermentation Method using a Magnesium Compound Containing Oxygen. U.S. Patent 8,431,372, 30 April 2010. [Google Scholar]
- Yocum, R.R.; Hermann, T.; Yu, X. Fermentation of Glycerol to Organic Acids 2013. WO2013/015770Al, 31 January 2013. [Google Scholar]
- Yocum, R.R.; Dole, S.; Pero, J.G. Production of Organic Acids by Fermentation at Low pH. U.S. Patent US20150240270A1, 28 September 2015. [Google Scholar]
- Cordes, T.; Michelucci, A.; Hiller, K. Itaconic Acid: The surprising role of an industrial compound as a mammalian antimicrobial metabolite. Annu. Rev. Nutr. 2015, 35, 451–473. [Google Scholar] [CrossRef]
- Gopaliya, D.; Kumar, V.; Khare, S.K. Recent advances in itaconic acid production from microbial cell factories. Biocatal. Agric. Biotechnol. 2021, 36, 102130. [Google Scholar] [CrossRef]
- Teleky, B.E.; Vodnar, D.C. Biomass-derived production of itaconic acid as a building block in specialty polymers. Polymers 2019, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- Cunha da Cruz, J.; Machado de Castro, A.; Camporese Sérvulo, E.F. World market and biotechnological production of itaconic acid. 3 Biotech. 2018, 8. [Google Scholar] [CrossRef]
- Bafana, R.; Pandey, R.A. New approaches for itaconic acid production: Bottlenecks and possible remedies. Crit. Rev. Biotechnol. 2018, 38, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Weastra Determination of market potential for selected platform chemicals: Itaconic acid, Succinic acid, 2,5-Furandicarboxylic acid. BioConSepT 2011, 1–173.
- Okabe, M.; Lies, D.; Kanamasa, S.; Park, E.Y. Biotechnological production of itaconic acid and its biosynthesis in Aspergillus terreus. Appl. Microbiol. Biotechnol. 2009, 84, 597–606. [Google Scholar] [CrossRef]
- Krull, S.; Hevekerl, A.; Kuenz, A.; Prüße, U. Process development of itaconic acid production by a natural wild type strain of Aspergillus terreus to reach industrially relevant final titers. Appl. Microbiol. Biotechnol. 2017, 101, 4063–4072. [Google Scholar] [CrossRef]
- Saha, B.C. Emerging biotechnologies for production of itaconic acid and its applications as a platform chemical. J. Ind. Microbiol. Biotechnol. 2017, 44, 303–315. [Google Scholar] [CrossRef]
- Maassen, N.; Panakova, M.; Wierckx, N.; Geiser, E.; Zimmermann, M.; Bölker, M.; Klinner, U.; Blank, L.M. Influence of carbon and nitrogen concentration on itaconic acid production by the smut fungus Ustilago maydis. Eng. Life Sci. 2014, 14, 129–134. [Google Scholar] [CrossRef]
- Kuenz, A.; Gallenmüller, Y.; Willke, T.; Vorlop, K.D. Microbial production of itaconic acid: Developing a stable platform for high product concentrations. Appl. Microbiol. Biotechnol. 2012, 96, 1209–1216. [Google Scholar] [CrossRef]
- Wierckx, N.; Agrimi, G.; Lübeck, P.S.; Steiger, M.G.; Mira, N.P.; Punt, P.J. Metabolic specialization in itaconic acid production: A tale of two fungi. Curr. Opin. Biotechnol. 2020, 62, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, T.; Sugisawa, T.; Ishidori, T.; Nakahara, T.; Sugiyama, J. Itaconic acid fermentation by a yeast belonging to the genus Candida. Agric. Biol. Chem. 1981, 45, 475–479. [Google Scholar] [CrossRef]
- Levinson, W.E.; Kurtzman, C.P.; Kuo, T.M. Production of itaconic acid by Pseudozyma antarctica NRRL Y-7808 under nitrogen-limited growth conditions. Enzyme Microb. Technol. 2006, 39, 824–827. [Google Scholar] [CrossRef]
- Winskillm, N. Tricarboxylic acid cycle activity in relation to itaconic acid biosynthesis by Aspergillus terreus. Microbiology 1983, 129, 2877–2883. [Google Scholar] [CrossRef][Green Version]
- Zhao, M.; Lu, X.; Zong, H.; Li, J.; Zhuge, B. Itaconic acid production in microorganisms. Biotechnol. Lett. 2018, 40, 455–464. [Google Scholar] [CrossRef]
- Geiser, E.; Przybilla, S.K.; Friedrich, A.; Buckel, W.; Wierckx, N.; Blank, L.M.; Bölker, M. Ustilago maydis produces itaconic acid via the unusual intermediate trans-aconitate. Microb. Biotechnol. 2016, 9, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Tehrani, H.H.; Ernst, P.; Blank, L.M.; Wierckx, N. An optimized Ustilago maydis for itaconic acid production at maximal theoretical yield. J. Fungi 2021, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Vila-Santa, A.; Liu, N.; Prozorov, T.; Xie, D.; Faria, N.T.; Ferreira, F.C.; Mira, N.P.; Shao, Z. Metabolic engineering of an acid-tolerant yeast strain Pichia kudriavzevii for itaconic acid production. Metab. Eng. Commun. 2020, 10, e00124. [Google Scholar] [CrossRef]
- Kolláth, I.S.; Molnár, Á.P.; Soós, Á.; Fekete, E.; Sándor, E.; Kovács, B.; Kubicek, C.P.; Karaffa, L. Manganese deficiency is required for high itaconic acid production from D-xylose in Aspergillus terreus. Front. Microbiol. 2019, 10, 1589. [Google Scholar] [CrossRef]
- Kerssemakers, A.A.J.; Doménech, P.; Cassano, M.; Yamakawa, C.K.; Dragone, G.; Mussatto, S.I. Production of itaconic acid from cellulose pulp: Feedstock feasibility and process strategies for an efficient microbial performance. Energies 2020, 13, 1654. [Google Scholar] [CrossRef]
- Demir, H.T.; Bezirci, E.; Becker, J.; Tehrani, H.H.; Nikerel, E.; Wierck, N.; Türker, M. High level production of itaconic acid at low pH by Ustilago maydis with fed-batch fermentation. Bioprocess Biosyst. Eng. 2021, 44, 749–758. [Google Scholar] [CrossRef]
- Zambanini, T.; Hosseinpour Tehrani, H.; Geiser, E.; Merker, D.; Schleese, S.; Krabbe, J.; Buescher, J.M.; Meurer, G.; Wierckx, N.; Blank, L.M. Efficient itaconic acid production from glycerol with Ustilago vetiveriae TZ1. Biotechnol. Biofuels 2017, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, Q.; Deng, Y.; Li, J.; Chen, X.; Gu, Y.; Lv, X.; Zheng, Z.; Jiang, S.; Li, X. Production of itaconic acid by biotransformation of wheat bran hydrolysate with Aspergillus terreus CICC40205 mutant. Bioresour. Technol. 2017, 241, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, G.; Zhang, J.; Balan, V.; Bao, J. Itaconic acid fermentation using activated charcoal-treated corn stover hydrolysate and process evaluation based on Aspen plus model. Biomass Convers. Biorefinery 2020, 10, 463–470. [Google Scholar] [CrossRef]
- Xie, H.; Ma, Q.; Wei, D.; Wang, F. Metabolic engineering of an industrial Aspergillus niger strain for itaconic acid production. 3 Biotech 2020, 10, 113. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xu, H.; Jiang, J.; Zhang, N.; Xie, J.; Wei, M.; Zhao, J. Production of itaconic acid through microbiological fermentation of inexpensive materials. J. Bioresour. Bioprod. 2019, 4, 135–142. [Google Scholar] [CrossRef]
- Klement, T.; Büchs, J. Itaconic acid—A biotechnological process in change. Bioresour. Technol. 2013, 135, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Ghaffar, T.; Irshad, M.; Anwar, Z.; Aqil, T.; Zulifqar, Z.; Tariq, A.; Kamran, M.; Ehsan, N.; Mehmood, S. Recent trends in lactic acid biotechnology: A brief review on production to purification. J. Radiat. Res. Appl. Sci. 2014, 7, 222–229. [Google Scholar] [CrossRef]
- Komesu, A.; de Oliveira, J.A.R.; da Silva Martins, L.H.; Maciel, M.R.W.; Maciel Filho, R. Lactic acid production to purification: A review. BioResources 2017, 12, 4364–4383. [Google Scholar] [CrossRef]
- Meng, K.; Zhang, G.; Ding, C.; Zhang, T.; Yan, H.; Zhang, D.; Fang, T.; Liu, M.; You, Z.; Yang, C.; et al. Recent advances on purification of lactic acid. Chem. Rec. 2020, 20, 1236–1256. [Google Scholar] [CrossRef] [PubMed]
- Farooq, U.; Anjum, F.M.; Zahoor, T.; Sajjad-Ur-Rahman; Randhawa, M.A.; Ahmed, A.; Akram, K. Optimization of lactic acid production from cheap raw material: Sugarcane molasses. Pakistan J. Bot. 2012, 44, 333–338. [Google Scholar]
- Ajala, E.O.; Olonade, Y.O.; Ajala, M.A.; Akinpelu, G.S. Lactic acid production from lignocellulose–A review of major challenges and selected solutions. ChemBioEng Rev. 2020, 7, 38–49. [Google Scholar] [CrossRef]
- Rawoof, S.A.A.; Kumar, P.S.; Vo, D.V.N.; Devaraj, K.; Mani, Y.; Devaraj, T.; Subramanian, S. Production of optically pure lactic acid by microbial fermentation: A review. Environ. Chem. Lett. 2021, 19, 539–556. [Google Scholar] [CrossRef]
- Jem, K.J.; Tan, B. The development and challenges of poly (lactic acid) and poly (glycolic acid). Adv. Ind. Eng. Polym. Res. 2020, 3, 60–70. [Google Scholar] [CrossRef]
- Naser, A.Z.; Deiab, I.; Darras, B.M. Poly(lactic acid) (PLA) and polyhydroxyalkanoates (PHAs), green alternatives to petroleum-based plastics: A review. RSC Adv. 2021, 11, 17151–17196. [Google Scholar] [CrossRef]
- Chung, M.R.W.Y.; Tan, I.S.; Foo, H.C.Y.; Lam, M.K.; Lim, S. Potential of macroalgae-based biorefinery for lactic acid production from exergy aspect. Biomass Convers. Biorefinery 2021, 1–31. [Google Scholar] [CrossRef]
- Alexandri, M.; Neu, A.K.; Schneider, R.; López-Gómez, J.P.; Venus, J. Evaluation of various Bacillus coagulans isolates for the production of high purity L-lactic acid using defatted rice bran hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 1321–1329. [Google Scholar] [CrossRef]
- Maas, R.H.W.; Bakker, R.R.; Jansen, M.L.A.; Visser, D.; De Jong, E.; Eggink, G.; Weusthuis, R.A. Lactic acid production from lime-treated wheat straw by Bacillus coagulans: Neutralization of acid by fed-batch addition of alkaline substrate. Appl. Microbiol. Biotechnol. 2008, 78, 751–758. [Google Scholar] [CrossRef]
- Awasthi, D.; Wang, L.; Rhee, M.S.; Wang, Q.; Chauliac, D.; Ingram, L.O.; Shanmugam, K.T. Metabolic engineering of Bacillus subtilis for production of D-lactic acid. Biotechnol. Bioeng. 2018, 115, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, J.; Aravindan, R.; Viruthagiri, T. Recent trends in the production, purification and application of lactic acid. Chem. Biochem. Eng. Q. 2008, 22, 245–264. [Google Scholar]
- Abedi, E.; Hashemi, S.M.B. Lactic acid production–Producing microorganisms and substrates sources-state of art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef] [PubMed]
- Holyoak, C.D.; Stratford, M.; Mcmullin, Z.; Cole, M.B.; Crimmins, K.; Brown, A.J.P.; Coote, P.J. Activity of the plasma membrane H+-ATPase and optimal glycolytic flux are required for rapid adaptation and growth of Saccharomyces cerevisiae in the presence of the weak-acid preservative sorbic acid. Appl. Environ. Microbiol. 1996, 62, 3158–3164. [Google Scholar] [CrossRef]
- Porro, D.; Bianchi, M.M.; Brambilla, L.; Menghini, R.; Bolzani, D.; Carrera, V.; Lievense, J.; Liu, C.; Ranzi, B.M.; Frontali, L.; et al. replacement of a metabolic pathway for large-scale production of lactic acid from engineered yeasts. Appl. Environ. Microbiol. 1999, 65, 4211–4215. [Google Scholar] [CrossRef] [PubMed]
- Alves De Oliveira, R.; Alexandri, M.; Komesu, A.; Venus, J.; Vaz Rossell, C.E.; Maciel Filho, R. Current advances in separation and purification of second-generation lactic acid. Sep. Purif. Rev. 2020, 49, 159–175. [Google Scholar] [CrossRef]
- Tian, X.; Chen, H.; Liu, H.; Chen, J. Recent advances in lactic acid production by lactic acid bacteria. Appl. Biochem. Biotechnol. 2021, 193, 4151–4171. [Google Scholar] [CrossRef] [PubMed]
- Pleissner, D.; Demichelis, F.; Mariano, S.; Fiore, S.; Navarro Gutiérrez, I.M.; Schneider, R.; Venus, J. Direct production of lactic acid based on simultaneous saccharification and fermentation of mixed restaurant food waste. J. Clean. Prod. 2017, 143, 615–623. [Google Scholar] [CrossRef]
- Zhou, X.; Ye, L.; Wu, J.C. Efficient production of l-lactic acid by newly isolated thermophilic Bacillus coagulans WCP10-4 with high glucose tolerance. Appl. Microbiol. Biotechnol. 2013, 97, 4309–4314. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Xu, P. Efficient production of l-lactic acid using co-feeding strategy based on cane molasses/glucose carbon sources. Bioresour. Technol. 2014, 153, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Porro, D.; Brambilla, L.; Ranzi, B.M.; Martegani, E.; Alberghina, L. Development of metabolically engineered Saccharomyces cerevisiae cells for the production of lactic acid. Biotechnol. Prog. 1995, 11, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Baptista, S.L.; Costa, C.E.; Cunha, J.T.; Soares, P.O.; Domingues, L. Metabolic engineering of Saccharomyces cerevisiae for the production of top value chemicals from biorefinery carbohydrates. Biotechnol. Adv. 2021, 47, 107697. [Google Scholar] [CrossRef]
- Sauer, M.; Porro, D.; Mattanovich, D.; Branduardi, P. 16 Years research on lactic acid production with yeast. Biotechnol. Genet. Eng. Rev. 2010, 21, 229–256. [Google Scholar] [CrossRef] [PubMed]
- Abbott, D.A.; Zelle, R.M.; Pronk, J.T.; van Maris, A.J.A. Metabolic engineering of Saccharomyces cerevisiae for production of carboxylic acids: Current status and challenges. FEMS Yeast Res. 2009, 9, 1123–1136. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Bae, J.H.; Ko, H.J.; Lee, S.H.; Sung, B.H.; Han, J.I.; Sohn, J.H. Low-pH production of D-lactic acid using newly isolated acid tolerant yeast Pichia kudriavzevii NG7. Biotechnol. Bioeng. 2018, 115, 2232–2242. [Google Scholar] [CrossRef]
- Eş, I.; Mousavi Khaneghah, A.; Barba, F.J.; Saraiva, J.A.; Sant’Ana, A.S.; Hashemi, S.M.B. Recent advancements in lactic acid production—A review. Food Res. Int. 2018, 107, 763–770. [Google Scholar] [CrossRef]
- Grand View Research Lactic Acid Market Size Worth $5.02 Billion by 2028. Available online: https://www.grandviewresearch.com/press-release/global-lactic-acid-and-poly-lactic-acid-market (accessed on 14 December 2021).
- Srivastava, D.G. Lactic acid production for pharmaceutical applications by Lactobacillus delbrueckii immobilized on chitosan and polyethylene glycol stabilized calcium alginate beads: Process optimization using response surface methodology. World J. Pharm. Res. 2017, 6, 1737–1762. [Google Scholar] [CrossRef][Green Version]
- Li, Y.; Bhagwat, S.S.; Cortés-Penã, Y.R.; Ki, D.; Rao, C.V.; Jin, Y.S.; Guest, J.S. Sustainable lactic acid production from lignocellulosic biomass. ACS Sustain. Chem. Eng. 2021, 9, 1341–1351. [Google Scholar] [CrossRef]
- Wasewar, K.L. Separation of lactic acid: Recent advances. Chem. Biochem. Eng. Q. 2005, 19, 159–172. [Google Scholar]
- Li, C.; Gao, M.; Zhu, W.; Wang, N.; Ma, X.; Wu, C.; Wang, Q. Recent advances in the separation and purification of lactic acid from fermentation broth. Process Biochem. 2021, 104, 142–151. [Google Scholar] [CrossRef]
- Lee, H.D.; Lee, M.Y.; Hwang, Y.S.; Cho, Y.H.; Kim, H.W.; Park, H.B. Separation and purification of lactic acid from fermentation broth using membrane-integrated separation Processes. Ind. Eng. Chem. Res. 2017, 56, 8301–8310. [Google Scholar] [CrossRef]
- Liang, S.; Jiang, W.; Song, Y.; Zhou, S.F. Improvement and metabolomics-based analysis of d -lactic acid production from agro-industrial wastes by Lactobacillus delbrueckii submitted to adaptive laboratory evolution. J. Agric. Food Chem. 2020, 68, 7660–7669. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Kwon, E.Y.; Bae, S.J.; Cho, B.R.; Kim, S.Y.; Hahn, J.S. Improvement of D-lactic acid production in Saccharomyces cerevisiae under acidic conditions by evolutionary and rational metabolic engineering. Biotechnol. J. 2017, 12, 1–7. [Google Scholar] [CrossRef]
- Watcharawipas, A.; Sae-Tang, K.; Sansatchanon, K.; Sudying, P.; Boonchoo, K.; Tanapongpipat, S.; Kocharin, K.; Runguphan, W. Systematic engineering of Saccharomyces cerevisiae for D-lactic acid production with near theoretical yield. FEMS Yeast Res. 2021, 21, 1–15. [Google Scholar] [CrossRef]
- Wang, Y.D.; Liao, J.Y.; Chiang, C.J.; Chao, Y.P. A simple strategy to effectively produce D-lactate in crude glycerol-utilizing Escherichia coli. Biotechnol. Biofuels 2019, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, D.; Tortajada, M.; Ramón, D.; Rojas, A. Production of D-lactic acid by the fermentation of orange peel waste hydrolysate by lactic acid bacteria. Fermentation 2020, 6, 1. [Google Scholar] [CrossRef]
- Beitel, S.M.; Coelho, L.F.; Contiero, J. Efficient conversion of agroindustrial waste into D(-) lactic acid by Lactobacillus delbrueckii using fed-batch fermentation. Biomed Res. Int. 2020, 2020, 4194052. [Google Scholar] [CrossRef]
- Zhang, Y.; Vadlani, P.V. Lactic acid production from biomass-derived sugars via co-fermentation of Lactobacillus brevis and Lactobacillus plantarum. J. Biosci. Bioeng. 2015, 119, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Panesar, P.S.; Saini, M.S. Optimization of process parameters and estimation of kinetic parameters for lactic acid production by Lactobacillus casei MTCC 1423. Biomass Convers. Biorefinery 2019, 9, 253–266. [Google Scholar] [CrossRef]
- Song, J.Y.; Park, J.S.; Kang, C.D.; Cho, H.Y.; Yang, D.; Lee, S.; Cho, K.M. Introduction of a bacterial acetyl-CoA synthesis pathway improves lactic acid production in Saccharomyces cerevisiae. Metab. Eng. 2016, 35, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Sun, X.; Zhu, H.; Jiang, R.; Luo, X.; Yin, L. An optimized fed-batch culture strategy integrated with a one-step fermentation improves L-lactic acid production by Rhizopus oryzae. World J. Microbiol. Biotechnol. 2018, 34, 74. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, H.; Yang, Y.; Zhou, X.; Xiu, Z. High-efficient l-lactic acid production from inedible starchy biomass by one-step open fermentation using thermotolerant Lactobacillus rhamnosus DUT1908. Bioprocess Biosyst. Eng. 2021, 44, 1935–1941. [Google Scholar] [CrossRef]
- Tian, X.; Liu, X.; Zhang, Y.; Chen, Y.; Hang, H.; Chu, J.; Zhuang, Y. Metabolic engineering coupled with adaptive evolution strategies for the efficient production of high-quality L-lactic acid by Lactobacillus paracasei. Bioresour. Technol. 2021, 323, 124549. [Google Scholar] [CrossRef]
- Chen, H.; Chen, B.; Su, Z.; Wang, K.; Wang, B.; Wang, Y.; Si, Z.; Wu, Y.; Cai, D.; Qin, P. Efficient lactic acid production from cassava bagasse by mixed culture of Bacillus coagulans and Lactobacillus rhamnosus using stepwise pH controlled simultaneous saccharification and co-fermentation. Ind. Crops Prod. 2020, 146, 112175. [Google Scholar] [CrossRef]
- Okano, K.; Uematsu, G.; Hama, S.; Tanaka, T.; Noda, H.; Kondo, A.; Honda, K. Metabolic engineering of Lactobacillus plantarum for direct L-lactic acid production from raw corn starch. Biotechnol. J. 2018, 13, 1–6. [Google Scholar] [CrossRef]
- Gao, M.; Ma, X.; Song, N.; Wang, Q.; Wu, C. A newly isolated strain, Lactobacillus paracasei subsp. paracasei 2, produces L-lactic acid from pilot-scale fermentation of food waste under sterile and nonsterile conditions. J. Chem. Technol. Biotechnol. 2020, 95, 3193–3201. [Google Scholar] [CrossRef]
- Li, C.; Ong, K.L.; Cui, Z.; Sang, Z.; Li, X.; Patria, R.D.; Qi, Q.; Fickers, P.; Yan, J.; Lin, C.S.K. Promising advancement in fermentative succinic acid production by yeast hosts. J. Hazard. Mater. 2021, 401, 123414. [Google Scholar] [CrossRef]
- Song, H.; Lee, S.Y. Production of succinic acid by bacterial fermentation. Enzyme Microb. Technol. 2006, 39, 352–361. [Google Scholar] [CrossRef]
- Putri, D.N.; Sahlan, M.; Montastruc, L.; Meyer, M.; Negny, S.; Hermansyah, H. Progress of fermentation methods for bio-succinic acid production using agro-industrial waste by Actinobacillus succinogenes. Energy Rep. 2020, 6, 234–239. [Google Scholar] [CrossRef]
- E4tech; RE-CORD; WUR. “From the Sugar Platform to Biofuels and Biochemicals”. Final Report for the European Commission; Contract No. ENER/C2/423-2012/SI2.673791; European Commission: Luxembourg, 2015. [Google Scholar]
- Pateraki, C.; Patsalou, M.; Vlysidis, A.; Kopsahelis, N.; Webb, C.; Koutinas, A.A.; Koutinas, M. Actinobacillus succinogenes: Advances on succinic acid production and prospects for development of integrated biorefineries. Biochem. Eng. J. 2016, 112, 285–303. [Google Scholar] [CrossRef]
- Dessie, W.; Xin, F.; Zhang, W.; Jiang, Y.; Wu, H.; Ma, J.; Jiang, M. Opportunities, challenges, and future perspectives of succinic acid production by Actinobacillus succinogenes. Appl. Microbiol. Biotechnol. 2018, 102, 9893–9910. [Google Scholar] [CrossRef] [PubMed]
- Cok, B.; Tsiropoulos, I.; Roes, A.L.; Patel, M.K. Succinic acid production derived from carbohydrates: An energy and greenhouse gas assessment of a platform chemical toward a bio-based economy. Biofuels Bioprod. Bioref 2014, 8, 16–29. [Google Scholar] [CrossRef]
- Ito, Y.; Hirasawa, T.; Shimizu, H. Metabolic engineering of Saccharomyces cerevisiae to improve succinic acid production based on metabolic profiling. Biosci. Biotechnol. Biochem. 2014, 78, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.K.; Wang, G.Y.; Zeng, J.; Zhang, J.A. Improved succinate production by metabolic engineering. Biomed Res. Int. 2013, 2013, 538790. [Google Scholar] [CrossRef] [PubMed]
- Van De Graaf, M.J.; Valianpoer, F.; Fiey, G.; Delattre, L.; Schulten, E.A.M. Process for the Crystallization of Succinic Acid 2015. US20150057425A1, 26 February 2015. [Google Scholar]
- Ahn, J.H.; Jang, Y.S.; Lee, S.Y. Production of succinic acid by metabolically engineered microorganisms. Curr. Opin. Biotechnol. 2016, 42, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Stylianou, E.; Pateraki, C.; Ladakis, D.; Damala, C.; Vlysidis, A.; Latorre-Sánchez, M.; Coll, C.; Lin, C.S.K.; Koutinas, A. Bioprocess development using organic biowaste and sustainability assessment of succinic acid production with engineered Yarrowia lipolytica strain. Biochem. Eng. J. 2021, 174, 108099. [Google Scholar] [CrossRef]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Succinate production in dual-phase Escherichia coli fermentations depends on the time of transition from aerobic to anaerobic conditions. J. Ind. Microbiol. Biotechnol. 2002, 28, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Salma, A.; Abdallah, R.; Fourcade, F.; Amrane, A.; Djelal, H. A New Approach to produce succinic acid through a co-culture system. Appl. Biochem. Biotechnol. 2021, 193, 2872–2892. [Google Scholar] [CrossRef] [PubMed]
- Kuenz, A.; Hoffmann, L.; Goy, K.; Bromann, S.; Prüße, U. High-level production of succinic acid from crude glycerol by a wild type organism. Catalysts 2020, 10, 470. [Google Scholar] [CrossRef]
- Chen, P.; Tao, S.; Zheng, P. Efficient and repeated production of succinic acid by turning sugarcane bagasse into sugar and support. Bioresour. Technol. 2016, 211, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Schroder, H.; Haefner, S.; Von Abendroth, G.; Hollmann, R.; Raddatz, A.; Ernst, H.; Gurski, H. Microbial Ssccinic Acid Producers and Purification of Succinic Acid. U.S. Patent 8,673,598, 18 March 2014. [Google Scholar]
- Li, C.; Gao, S.; Yang, X.; Lin, C.S.K. Green and sustainable succinic acid production from crude glycerol by engineered Yarrowia lipolytica via agricultural residue based in situ fibrous bed bioreactor. Bioresour. Technol. 2018, 249, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Cui, Z.; Zhu, Z.; Liu, Y.; Tang, Y.J.; Hou, J.; Qi, Q. Engineering of Yarrowia lipolytica transporters for high-efficient production of biobased succinic acid from glucose. Biotechnol. Biofuels 2021, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Gao, C.; Li, J.; Hou, J.; Lin, C.S.K.; Qi, Q. Engineering of unconventional yeast Yarrowia lipolytica for efficient succinic acid production from glycerol at low pH. Metab. Eng. 2017, 42, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ong, K.L.; Yang, X.; Lin, C.S.K. Bio-refinery of waste streams for green and efficient succinic acid production by engineered Yarrowia lipolytica without pH control. Chem. Eng. J. 2019, 371, 804–812. [Google Scholar] [CrossRef]
- Porro, D.; Branduardi, P.; Sauer, M.; Mattanovich, D. Old obstacles and new horizons for microbial chemical production. Curr. Opin. Biotechnol. 2014, 30, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Crater, J.S.; Lievense, J.C. Scale-up of industrial microbial processes. FEMS Microbiol. Lett. 2018, 365, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Soares-Silva, I.; Ribas, D.; Sousa-Silva, M.; Azevedo-Silva, J.; Rendulić, T.; Casal, M. Membrane transporters in the bioproduction of organic acids: State of the art and future perspectives for industrial applications. FEMS Microbiol. Lett. 2020, 367, 1–15. [Google Scholar] [CrossRef]
- Lu, J.; Lv, Y.; Qian, X.; Jiang, Y.; Wu, M.; Zhang, W.; Zhou, J.; Dong, W.; Xin, F.; Jiang, M. Current advances in organic acid production from organic wastes by using microbial co-cultivation systems. Biofuels Bioprod. Biorefining 2020, 14, 481–492. [Google Scholar] [CrossRef]
- Steiger, M.G.; Mattanovich, D.; Sauer, M. Microbial organic acid production as carbon dioxide sink. FEMS Microbiol. Lett. 2017, 364, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Branduardi, P.; Sauer, M. Microbial carbon dioxide fixation: New tricks for an old game. FEMS Microbiol. Lett. 2018, 365, 1–7. [Google Scholar] [CrossRef] [PubMed]


| 2004 | 2010 |
|---|---|
| 1,4-Diacids (Succinic, fumaric and malic acids) | 3-Hydroxypropionic acid/aldehyde |
| 2,5-Furan dicarboxylic | Bio-hydrocarbons |
| 3-Hydroxybutyrolactone | Ethanol |
| 3-Hydroxy propionic acid | Furans |
| Aspartic acid | Glycerol and derivatives |
| Glucaric acid | Lactic acid |
| Aylitol/Arabitol | Levulinic acid |
| Glutamic acid | Succinic acid |
| Itaconic acid | Sorbitol |
| Levulinic acid | Xylitol |
| Glycerol | |
| Sorbitol |
| Microorganism | Substrate | Fumaric Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| R. arrhizus NRRL 1526 | Glucose | 130 | Batch | [74] |
| R. arrhizus NRRL 1526 | Perennial grasses | 8.9 | Shake flask | [75] |
| R. oryzae ATCC20344 | Lignocellulosic Syrup | 34.2 | Shake flask | [82] |
| R. arrhizus RH7-13 | Food Waste | 32.7 | Shake flask | [78] |
| R. oryzae NRRL 1526 | Brewery wastewater | 31.3 | Shake flask | [85] |
| R. arrhizus RH-07-13 | Glucose and glycerol | 22.8 | Shake flask | [84] |
| R. arrhizus RH-7−13-9 | Glucose | 17.5 | Simultaneous fermentation and separation | [94] |
| Mutant E. coli E2 | Glycerol | 41.5 | Fed-batch | [69] |
| Mutant S. cerevisiae | Glucose | 33.1 | Shake flask | [71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Lorenzo, R.D.; Serra, I.; Porro, D.; Branduardi, P. State of the Art on the Microbial Production of Industrially Relevant Organic Acids. Catalysts 2022, 12, 234. https://doi.org/10.3390/catal12020234
Di Lorenzo RD, Serra I, Porro D, Branduardi P. State of the Art on the Microbial Production of Industrially Relevant Organic Acids. Catalysts. 2022; 12(2):234. https://doi.org/10.3390/catal12020234
Chicago/Turabian StyleDi Lorenzo, Raffaella Desirè, Immacolata Serra, Danilo Porro, and Paola Branduardi. 2022. "State of the Art on the Microbial Production of Industrially Relevant Organic Acids" Catalysts 12, no. 2: 234. https://doi.org/10.3390/catal12020234
APA StyleDi Lorenzo, R. D., Serra, I., Porro, D., & Branduardi, P. (2022). State of the Art on the Microbial Production of Industrially Relevant Organic Acids. Catalysts, 12(2), 234. https://doi.org/10.3390/catal12020234