
Laccase and Its Mutant Displayed on the Bacillus subtilis Spore Coat for Oxidation of Phenolic Compounds in Organic Solvents

Abstract
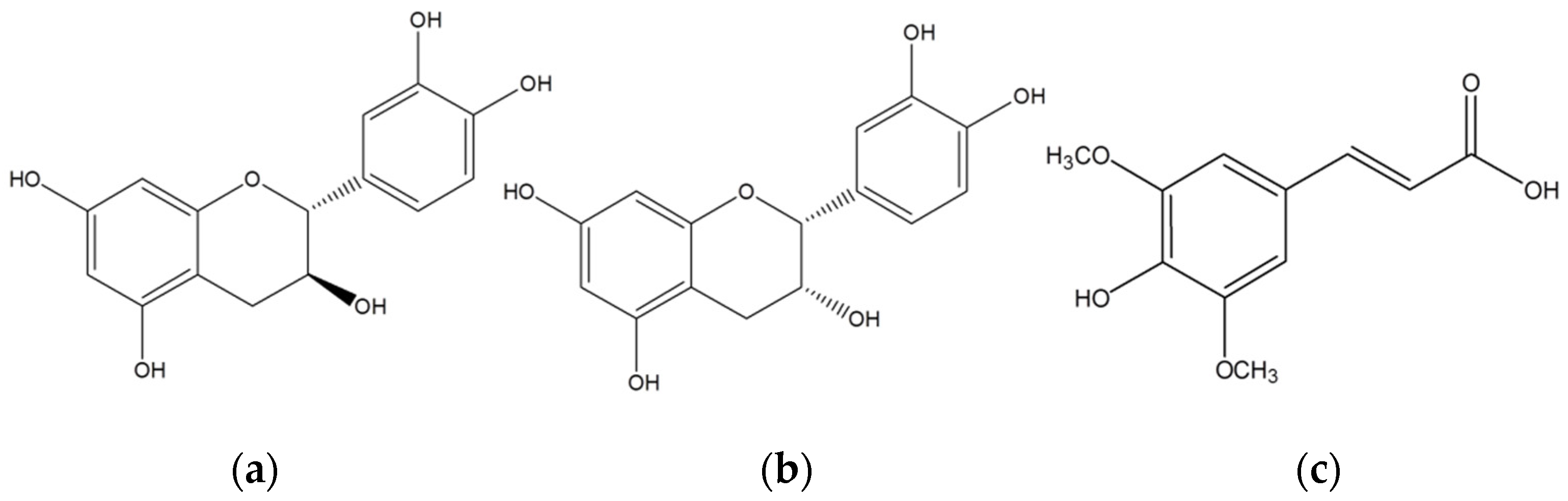
1. Introduction
2. Results
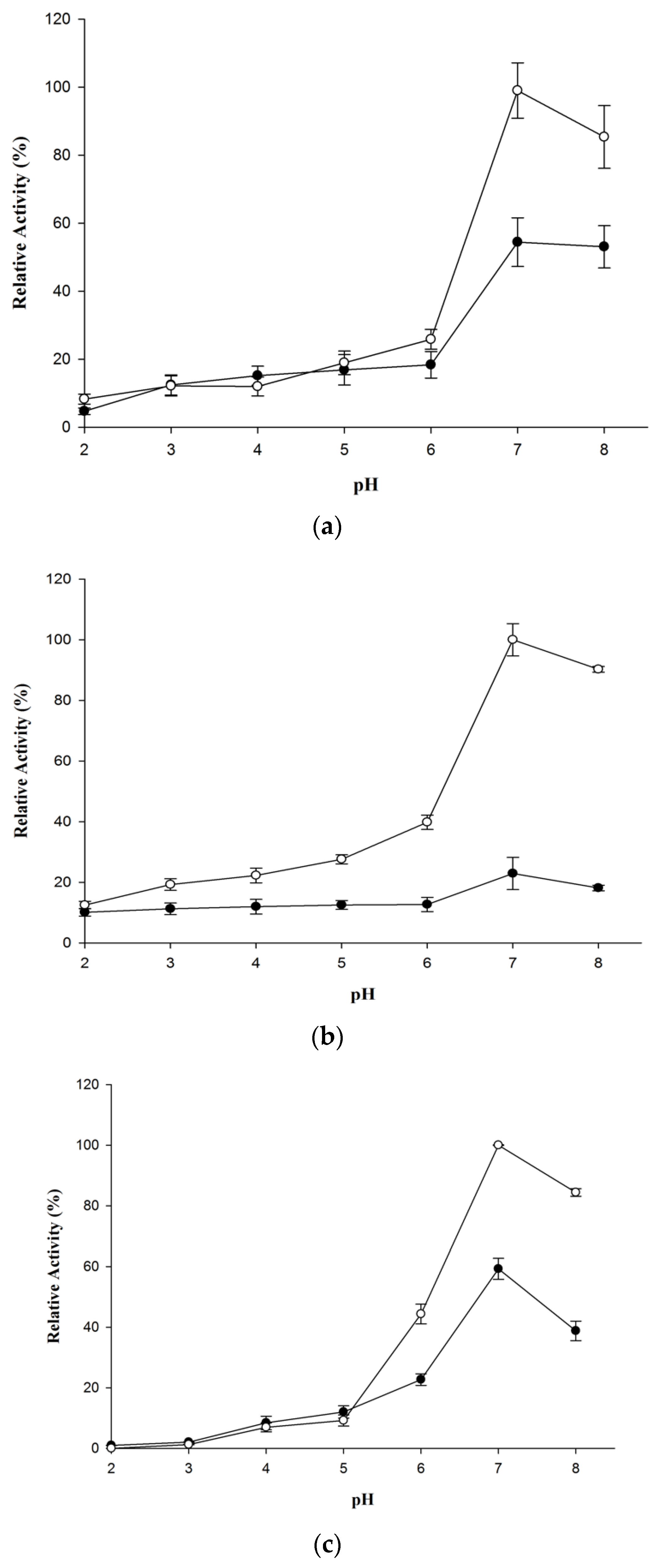
2.1. Wt-CotA and T480A-CotA Activity Versus pH
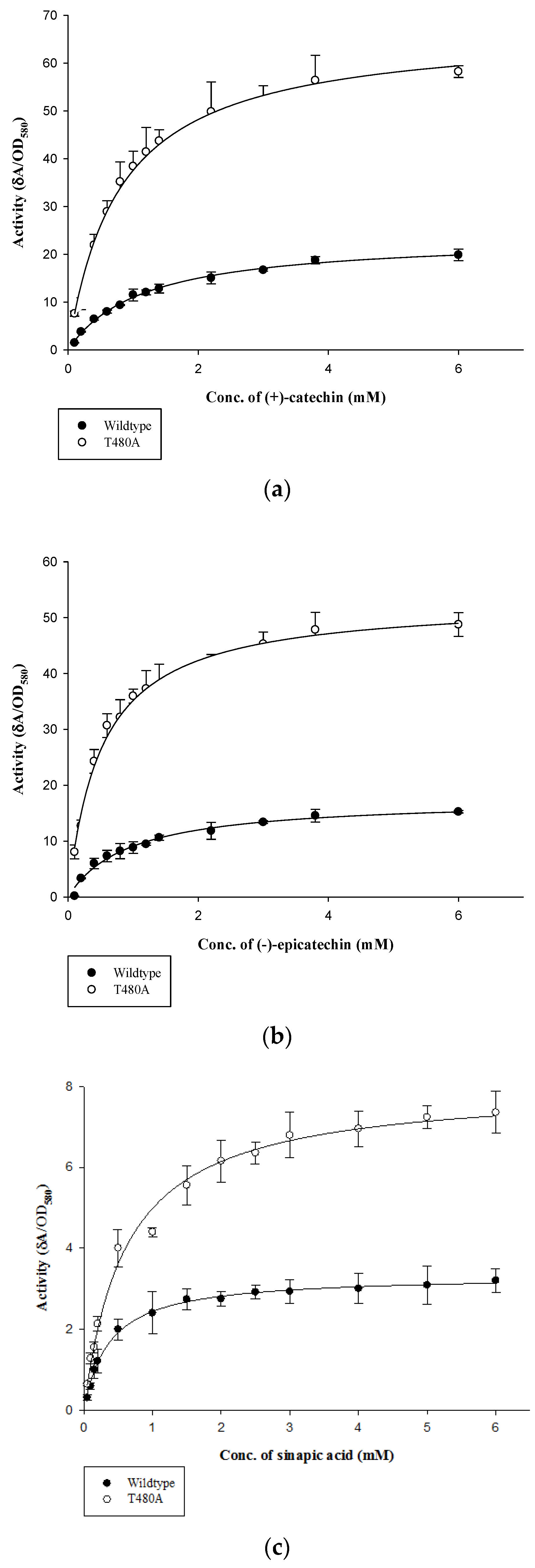
2.2. Determination of Kinetic Parameters
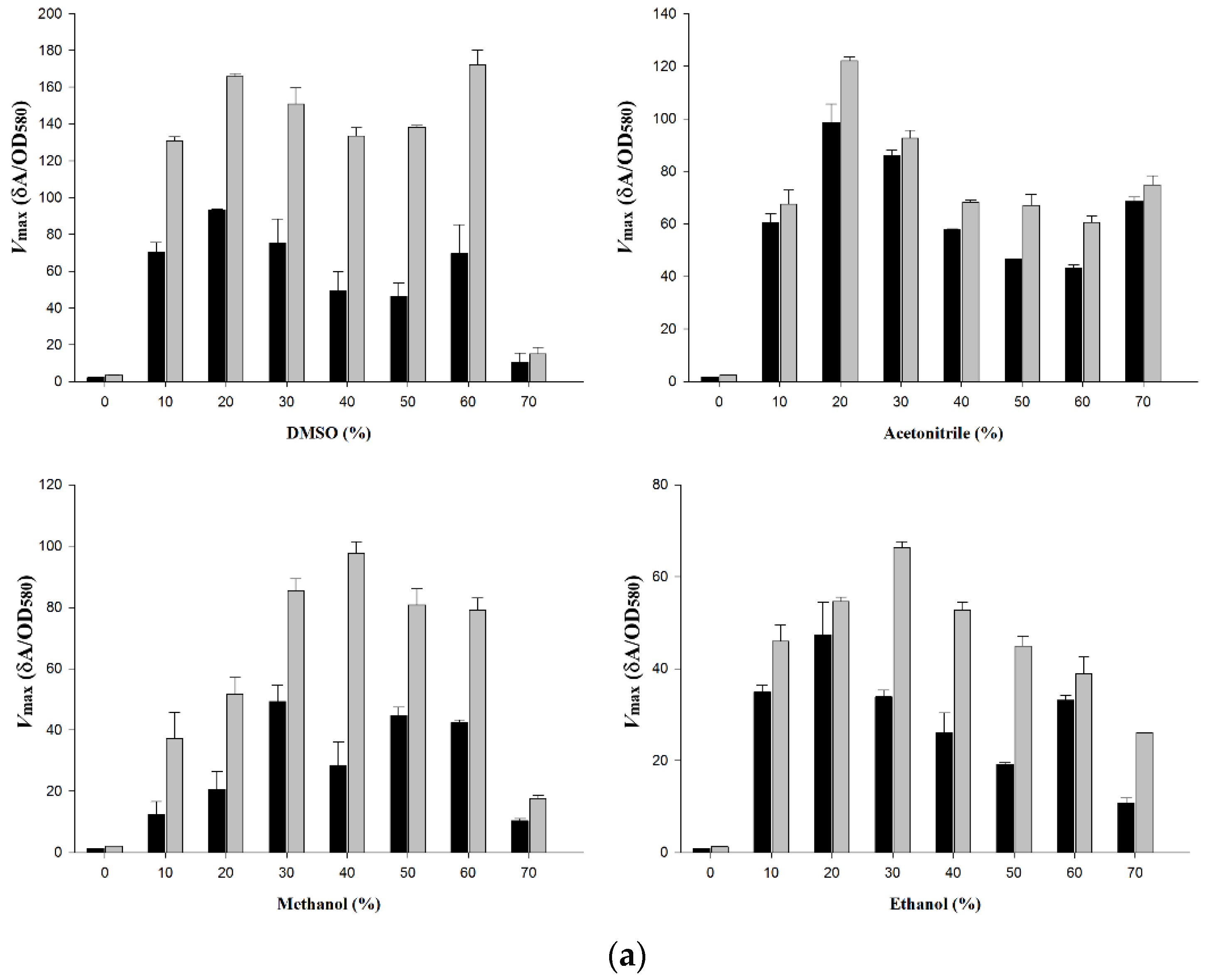
2.3. Activity of wt-CotA and T480A-CotA in Organic Solvent
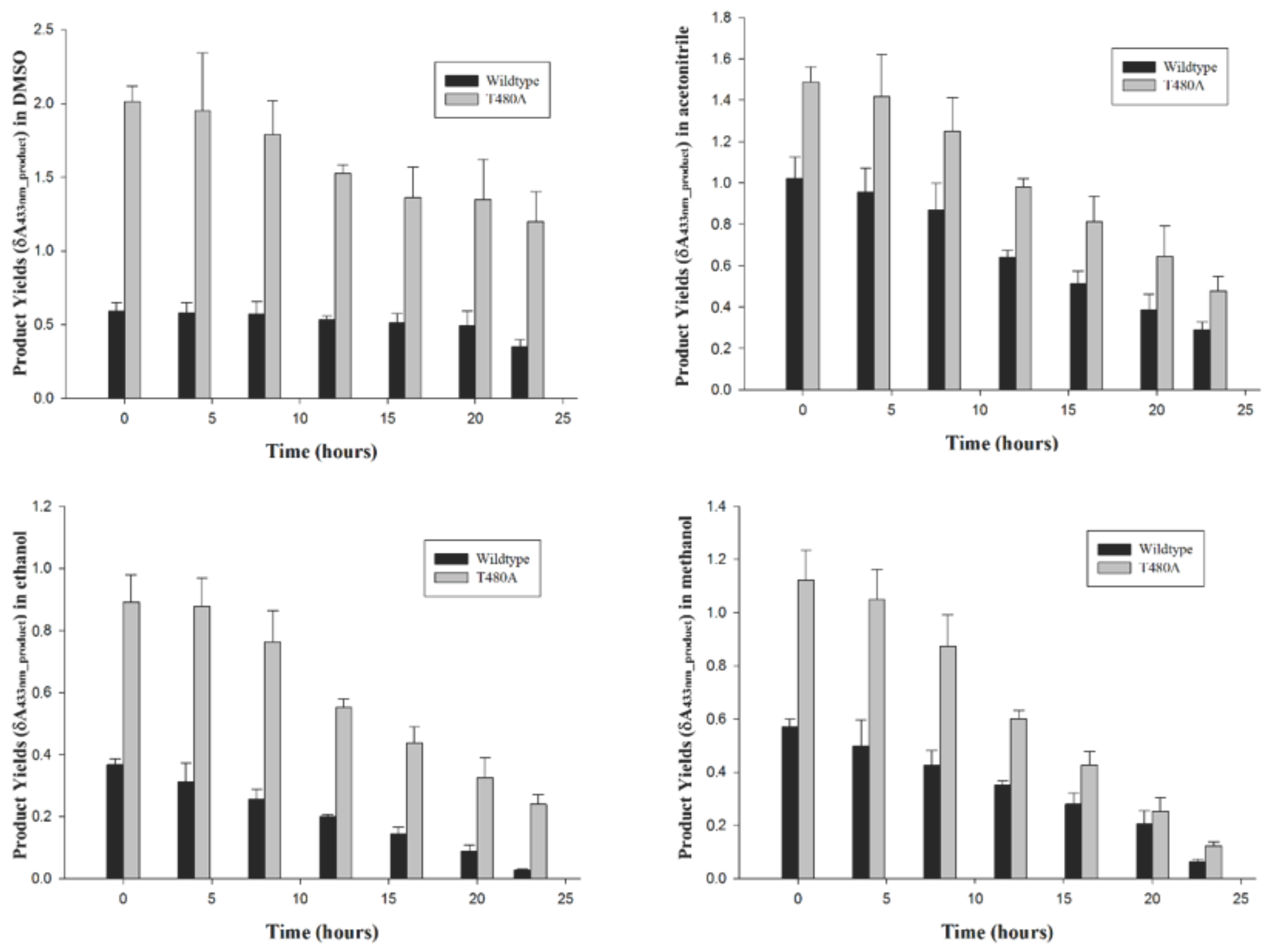
2.4. Wt-CotA and T480A-CotA Recycling in Organic Solvents
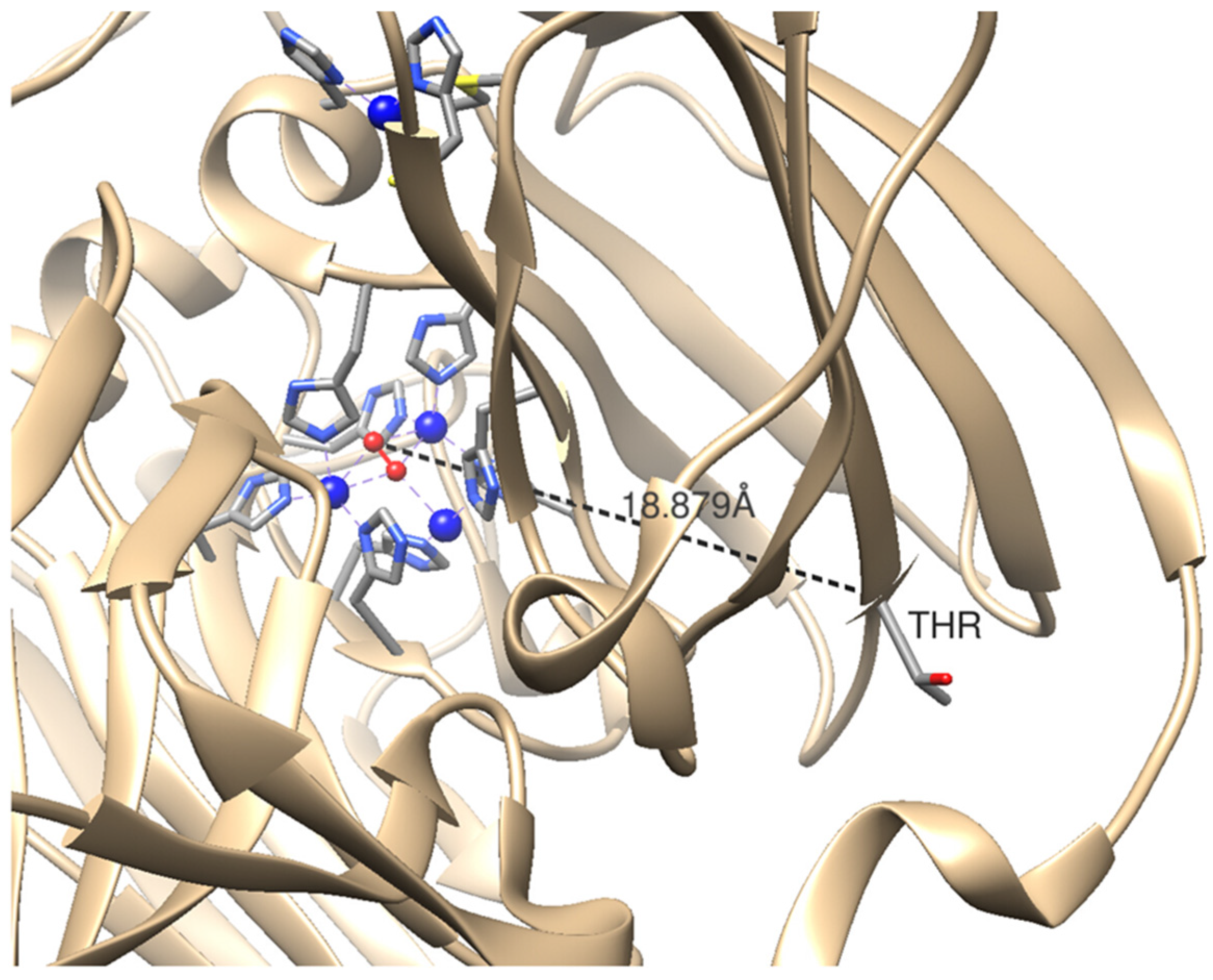
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Wt-CotA and T480A-CotA Activity Versus pH
4.3. Determination of Kinetic Parameters of wt-CotA and T480A-CotA
4.4. Wt-CotA and T480A-CotA Activity in Organic Solvents
4.5. Wt-CotA and T480A-CotA Recycling in Organic Solvents
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a. Vmax (δA/OD580) of wt-CotA and T480A-CotA in 0–70% Organic Solvents for Substrate (+)-Catechin | ||||||||
| Conc. of Organic Solvents | Vmax (δA/OD580) | |||||||
| DMSO | Acetonitrile | Methanol | Ethanol | |||||
| wt | T480A | wt | T480A | wt | T480A | wt | T480A | |
| 0% | 2.3 ± 0.1 | 3.5 ± 0.2 | 1.6 ± 0.1 | 2.5 ± 0.1 | 1.32 ± 0.2 | 2.01 ± 0.1 | 0.9 ± 0.0 | 1.33 ± 0.1 |
| 10% | 70.4 ± 5.3 | 130.9 ± 2.3 | 60.5 ± 3.3 | 67.6 ± 5.3 | 12.4 ± 4.0 | 36.6 ± 8.4 | 34.9 ± 1.4 | 46.0 ± 3.4 |
| 20% | 93.5 ± 0.2 | 165.7 ± 1.7 | 98.7 ± 6.9 | 122 ± 1.6 | 20.3 ± 5.8 | 50.9 ± 5.4 | 47.4 ± 7 | 54.6 ± 1.0 |
| 30% | 75.3 ± 13.1 | 150.9 ± 8.7 | 86.0 ± 2.1 | 92.7 ± 2.9 | 48.6 ± 5.2 | 84.2 ± 3.8 | 33.8 ± 1.5 | 66.3 ± 1.4 |
| 40% | 49.1 ± 10.5 | 133.6 ± 4.3 | 57.7 ± 0.3 | 68.2 ± 0.9 | 28.1 ± 7.5 | 96.1 ± 3.9 | 26.1 ± 4.3 | 52.7 ± 1.7 |
| 50% | 46.3 ± 7.1 | 138.3 ± 0.8 | 46.6 ± 0.1 | 67.0 ± 4.2 | 44.0 ± 2.7 | 79.6 ± 5.2 | 19.2 ± 0.4 | 44.9 ± 2.2 |
| 60% | 69.7 ± 15.6 | 172.2 ± 8.0 | 43.3 ± 1.2 | 60.6 ± 2.6 | 41.8 ± 0.9 | 77.9 ± 4.0 | 33.2 ± 1.0 | 38.9 ± 3.7 |
| 70% | 10.5 ± 5.0 | 15.1 ± 3.5 | 68.6 ± 1.5 | 74.8 ± 3.5 | 10.2 ± 0.7 | 17.4 ± 1.0 | 10.8 ± 1.0 | 26.0 ± 0.1 |
| b. Vmax (δA/OD580) of wt-CotA and T480A-CotA in 0–70% Organic Solvents for Substrate (−)-Epicatechin | ||||||||
| Conc. of Organic Solvents | Vmax (δA/OD580) | |||||||
| DMSO | Acetonitrile | Methanol | Ethanol | |||||
| wt | T480A | wt | T480A | wt | T480A | Wt | T480A | |
| 0% | 2.9 ± 0.1 | 4.4 ± 0.2 | 1.7 ± 0.1 | 2.7 ± 0.1 | 1.6 ± 0.1 | 2.4 ± 0.1 | 1.0 ± 0.0 | 1.5 ± 0.1 |
| 10% | 9.2 ± 0.5 | 22.6 ± 6.9 | 16.2 ± 1.7 | 42.0 ± 3.4 | 19.3 ± 0.4 | 56.4 ± 9.6 | 11.7 ± 1.4 | 22.3 ± 3.2 |
| 20% | 31.0 ± 2.7 | 126.6 ± 1.6 | 96.8 ± 1.1 | 133 ± 1.9 | 25.4 ± 4.2 | 63.6 ± 3.8 | 22.2 ± 2.8 | 29.0 ± 1.0 |
| 30% | 57.4 ± 5.9 | 154.7 ± 2.3 | 74.5 ± 3.6 | 77.4 ± 1.0 | 43.6 ± 3.0 | 90.7 ± 3.6 | 44.9 ± 1.4 | 76.1 ± 1.3 |
| 40% | 67.8 ± 11.1 | 119.6 ± 7.8 | 15.6 ± 0.1 | 67.8 ± 0.6 | 14.4 ± 4.4 | 95.1 ± 9.9 | 19.4 ± 2.9 | 71.5 ± 1.6 |
| 50% | 91.2 ± 7.8 | 111.0 ± 4.3 | 13.4 ± 0.1 | 75.3 ± 2.8 | 18.7 ± 2.6 | 58.2 ± 3.4 | 9.6 ± 0.4 | 59.9 ± 2.1 |
| 60% | 213.6 ± 7.5 | 220.4 ± 2.0 | 63.5 ± 0.6 | 83.7 ± 1.7 | 15.4 ± 3.6 | 60.9 ± 3.5 | 20.1 ± 1.0 | 39.3 ± 3.5 |
| 70% | 7.4 ± 4.3 | 20.1 ± 5.0 | 8.7 ± 0.8 | 36.4 ± 2.3 | 10.9 ± 0.6 | 11.6 ± 0.9 | 1.3 ± 1.0 | 10.3 ± 0.1 |
| c. Vmax (δA/OD580) of wt-CotA and T480A-CotA in 0–70% Organic Solvents for Substrate Sinapic Acid | ||||||||
| Conc. of Organic Solvents | Vmax (δA/OD580) | |||||||
| DMSO | Acetonitrile | Methanol | Ethanol | |||||
| wt | T480A | wt | T480A | wt | T480A | Wt | T480A | |
| 0% | 0.2 ± 0.0 | 0.3 ± 0.0 | 0.3 ± 0.0 | 0.5 ± 0.0 | 0.1 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.3 ± 0.0 |
| 10% | 2.5 ± 0.4 | 9.6 ± 0.2 | 6.2 ± 0.6 | 12.3 ± 1.0 | 3.3 ± 0.4 | 9.6 ± 1.0 | 4.4 ± 0.0 | 15.6 ± 0.9 |
| 20% | 5.1 ± 0.0 | 12.3 ± 0.1 | 9.3 ± 1.3 | 14.2 ± 0.3 | 3.1 ± 0.6 | 10.8 ± 0.6 | 7.1 ± 0.4 | 17.0 ± 0.2 |
| 30% | 5.1 ± 1.0 | 13.5 ± 0.7 | 13.0 ± 0.4 | 23.9 ± 0.6 | 1.8 ± 0.6 | 11.1 ± 0.4 | 3.0 ± 0.4 | 15.3 ± 0.4 |
| 40% | 3.2 ± 0.8 | 8.9 ± 0.3 | 12.1 ± 0.0 | 22.4 ± 0.2 | 1.7 ± 0.8 | 7.9 ± 0.4 | 3.4 ± 0.6 | 12.6 ± 0.4 |
| 50% | 1.8 ± 0.5 | 8.7 ± 0.0 | 8.7 ± 0.0 | 19.7 ± 0.8 | 0.4 ± 0.3 | 0.6 ± 0.1 | 2.8 ± 0.1 | 6.7 ± 0.6 |
| 60% | 1.4 ± 1.2 | 4.1 ± 0.6 | 5.1 ± 0.2 | 7.2 ± 0.5 | 0.2 ± 0.1 | 0.4 ± 0.2 | 1.0 ± 0.3 | 2.7 ± 0.9 |
| 70% | 1.0 ± 0.4 | 3.4 ± 0.3 | 0.7 ± 0.3 | 1.2 ± 0.7 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.3 ± 0.3 | 0.5 ± 0.0 |
| Time (Hours) | Product Yields (δA433nm_product) | |||||||
|---|---|---|---|---|---|---|---|---|
| DMSO | Acetonitrile | Methanol | Ethanol | |||||
| wt | T480A | wt | T480A | wt | T480A | wt | T480A | |
| 0 | 0.59 | 2.01 | 1.02 | 1.49 | 0.37 | 0.89 | 0.57 | 1.12 |
| 4 | 0.58 | 1.95 | 0.96 | 1.42 | 0.31 | 0.88 | 0.5 | 1.05 |
| 8 | 0.57 | 1.79 | 0.87 | 1.25 | 0.26 | 0.76 | 0.43 | 0.87 |
| 12 | 0.53 | 1.53 | 0.64 | 0.98 | 0.20 | 0.55 | 0.35 | 0.6 |
| 16 | 0.51 | 1.36 | 0.51 | 0.81 | 0.14 | 0.44 | 0.28 | 0.43 |
| 20 | 0.49 | 1.35 | 0.39 | 0.64 | 0.09 | 0.32 | 0.21 | 0.25 |
| 23 | 0.35 | 1.20 | 0.29 | 0.48 | 0.03 | 0.24 | 0.06 | 0.12 |
References
- Kumar, R.A.; Clark, D.S. High-throughput screening of biocatalytic activity: Applications in drug discovery. Curr. Opin. Chem. Biol. 2006, 10, 162–168. [Google Scholar] [CrossRef]
- Porter, J.L.; Rusli, R.A.; Ollis, D.L. Directed Evolution of Enzymes for Industrial Biocatalysis. ChemBioChem 2016, 17, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, R. Biocatalysis-key to sustainable industrial chemistry. Curr. Opin. Biotechnol. 2010, 21, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Klibanov, A. Why are enzymes less active in organic solvents than in water? Trends Biotechnol. 1997, 15, 97–101. [Google Scholar] [CrossRef]
- Doukyu, N.; Ogino, H. Organic solvent-tolerant enzymes. Biochem. Eng. J. 2010, 48, 270–282. [Google Scholar] [CrossRef]
- Stepankova, V.; Bidmanova, S.; Koudelakova, T.; Prokop, Z.; Chaloupkova, R.; Damborsky, J. Strategies for stabilization of enzymes in organic solvents. ACS Catal. 2013, 3, 2823–2836. [Google Scholar] [CrossRef]
- Dicosimo, R.; McAuliffe, J.; Poulose, A.J.; Bohlmann, G. Industrial use of immobilized enzymes. Chem. Soc. Rev. 2013, 42, 6437–6474. [Google Scholar] [CrossRef] [PubMed]
- Liese, A.; Hilterhaus, L. Evaluation of immobilized enzymes for industrial applications. Chem. Soc. Rev. 2013, 42, 6236–6249. [Google Scholar] [CrossRef]
- Binay, B.; Alagöz, D.; Yildirim, D.; Çelik, A.; Tükel, S.S. Highly stable and reusable immobilized formate dehydrogenases: Promising biocatalysts for in situ regeneration of NADH. Beilstein J. Org. Chem. 2016, 12, 271–277. [Google Scholar] [CrossRef]
- Mitsou, E.; Xenakis, A.; Zoumpanioti, M. Oxidation catalysis by enzymes in microemulsions. Catalysts 2017, 7, 52. [Google Scholar] [CrossRef]
- Qiu, X.; Wang, Y.; Xue, Y.; Li, W.; Hu, Y. Laccase immobilized on magnetic nanoparticles modified by amino-functionalized ionic liquid via dialdehyde starch for phenolic compounds biodegradation. Chem. Eng. J. 2020, 391. [Google Scholar] [CrossRef]
- Shao, X.; Ni, H.; Lu, T.; Jiang, M.; Li, H.; Huang, X.; Li, L. An improved system for the surface immobilisation of proteins on Bacillus thuringiensis vegetative cells and spores through a new spore cortex-lytic enzyme anchor. New Biotechnol. 2012, 29, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Bessette, P.H.; Åslund, F.; Beckwith, J.; Georgiou, G. Efficient folding of proteins with multiple disulfide bonds in the Escherichia coli cytoplasm. Proc. Natl. Acad. Sci. USA 1999, 96, 13703–13708. [Google Scholar] [CrossRef]
- Chen, G.; Driks, A.; Tawfiq, K.; Mallozzi, M.; Patil, S. Bacillus anthracis and Bacillus subtilis spore surface properties and transport. Colloids Surf. B Biointerfaces 2010, 76, 512–518. [Google Scholar] [CrossRef]
- Moeller, R.; Setlow, P.; Reitz, G.; Nicholson, W.L. Roles of small, acid-soluble spore proteins and core water content in survival of Bacillus subtilis spores exposed to environmental solar UV radiation. Appl. Environ. Microbiol. 2009, 75, 5202–5208. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Lee, F.S.; Farinas, E.T. Bacillus subtilis spore display of laccase for evolution under extreme conditions of high concentrations of organic solvent. ACS Combi. Sci. 2014, 16, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.; Dworkin, J. Recent progress in Bacillus subtilis sporulation. FEMS Microbiol. Rev. 2012, 36, 131–148. [Google Scholar] [CrossRef]
- Gupta, N.; Farinas, E.T. Directed evolution of CotA laccase for increased substrate specificity using Bacillus subtilis spores. Protein Eng. Des. Sel. 2010, 23, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, H.; Parvez, S.; Rehman, H.; Banerjee, B.D.; Raisuddin, S. Catechin as an antioxidant in liver mitochondrial toxicity: Inhibition of tamoxifen-induced protein oxidation and lipid peroxidation. J. Biochem. Mol. Toxicol. 2007, 21, 110–117. [Google Scholar] [CrossRef]
- Sangwan, N.S.; Shanker, S.; Sangwan, R.S.; Kumar, S. Plant-derived products as antimutagens. Phytother. Res. 1998, 12, 389–399. [Google Scholar] [CrossRef]
- Suzuki, J.I.; Isobe, M.; Morishita, R.; Nagai, R. Effects of green tea catechin consumption on cardiovascular diseases. In Tea Consumption and Health; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2012; pp. 87–99. [Google Scholar]
- Wang, Y.F.; Shao, S.H.; X u, P.; Yang, X.Q.; Qian, L.S. Catechin-enriched green tea extract as a safe and effective agent for antimicrobial and anti-inflammatory treatment. Afr. J. Pharm. Pharmacol. 2011, 5, 1452–1461. [Google Scholar] [CrossRef]
- Han, Z.X.; Rana, M.M.; Liu, G.F.; Gao, M.J.; Li, D.X.; Wu, F.G.; Li, X.B.; Wan, X.C.; Wei, S. Green tea flavour determinants and their changes over manufacturing processes. Food Chem. 2016, 212, 739–748. [Google Scholar] [CrossRef]
- Papp, A.; Winnewisser, W.; Geiger, E.; Briem, F. Influence of (+)-catechin and ferulic acid on formation of beer haze and their removal through different polyvinylpolypyrollidone-types. J. Inst. Brew. 2001, 107, 55–60. [Google Scholar] [CrossRef]
- Kurisawa, M.; Chung, J.E.; Uyama, H.; Kobayashi, S. Laccase-catalyzed Synthesis and Antioxidant Property of Poly(catechin). Macromol. Biosci. 2003, 3, 758–764. [Google Scholar] [CrossRef]
- Madeira Junior, J.V., Jr.; Teixeira, C.B.; Macedo, G.A. Biotransformation and bioconversion of phenolic compounds obtainment: An overview. Crit. Rev. Biotechnol. 2015, 35, 75–81. [Google Scholar] [CrossRef]
- Krygier, K.; Sosulski, F.; Hogge, L. Free, esterified, and insoluble-bound phenolic acids. 2. Composition of phenolic acids in rapeseed flour and hulls. J. Agric. Food Chem. 1982, 30, 334–336. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, L.; Pu, H.; Li, C.; Hu, Q. Profile and distribution of soluble and insoluble phenolics in Chinese rapeseed (Brassica napus). Food Chem. 2012, 135, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Naczk, M.; Amarowicz, R.; Sullivan, A.; Shahidi, F. Current research developments on polyphenolics of rapeseed/canola: A review. Food Chem. 1998, 62, 489–502. [Google Scholar] [CrossRef]
- Shahidi, F.; Naczk, M. An overview of the phenolics of canola and rapeseed: Chemical, sensory and nutritional significance. J. Am. Oil Chem. Soc. 1992, 69, 917–924. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Hao, Z.; Xi, R.; Cai, Y.; Liao, X. Hydrogen peroxide-resistant CotA and YjqC of bacillus altitudinis spores are a promising biocatalyst for catalyzing reduction of sinapic acid and sinapine in rapeseed meal. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Solomon, E.I. Electron transfer and reaction mechanism of laccases. Cell. Mol. Life Sci. 2015, 72, 869–883. [Google Scholar] [CrossRef]
- Solomon, E.I.; Heppner, D.E.; Johnston, E.M.; Ginsbach, J.W.; Cirera, J.; Qayyum, M.; Kieber-Emmons, M.T.; Kjaergaard, C.H.; Hadt, R.G.; Tian, L. Copper active sites in biology. Chem. Rev. 2014, 114, 3659–3853. [Google Scholar] [CrossRef] [PubMed]
- Kües, U. Fungal enzymes for environmental management. Curr. Opin. Biotechnol. 2015, 33, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Pardo, I.; Camarero, S. Laccase engineering by rational and evolutionary design. Cell. Mol. Life Sci. 2015, 72, 897–910. [Google Scholar] [CrossRef]
- Santhanam, N.; Vivanco, J.M.; Decker, S.R.; Reardon, K.F. Expression of industrially relevant laccases: Prokaryotic style. Trends Biotechnol. 2011, 29, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Shraddha; Shekher, R.; Sehgal, S.; Kamthania, M.; Kumar, A. Laccase: Microbial sources, production, purification, and potential biotechnological applications. Enzyme Res. 2011, 2011. [Google Scholar] [CrossRef]
- Ma, H.L.; Kermasha, S.; Gao, J.M.; Borges, R.M.; Yu, X.z. Laccase-catalyzed oxidation of phenolic compounds in organic media. J. Mol. Catal. B Enzym. 2009, 57, 89–95. [Google Scholar] [CrossRef]
- Osman, A.M.; Wong, K.K.Y.; Fernyhough, A. The laccase/ABTS system oxidizes (+)-catechin to oligomeric products. Enzym. Microb. Technol. 2007, 40, 1272–1279. [Google Scholar] [CrossRef]






| Substrate | Laccase | Km 1 | Vmax 2 | Vmax/Km |
|---|---|---|---|---|
| (+)-catechin | Wildtype | 1.15 ± 0.06 | 23.7 ± 0.49 | 20.6 |
| T480A | 0.79 ± 0.05 | 67.2 ± 1.54 | 85 | |
| (−)-epicatechin | Wildtype | 0.94 ± 0.1 | 17.6 ± 0.69 | 18.7 |
| T480A | 0.51 ± 0.03 | 53.2 ± 0.99 | 104.3 | |
| Sinapic acid | Wildtype | 0.36 ± 0.02 | 3.33 ± 0.04 | 9.3 |
| T480A | 0.61 ± 0.05 | 8.00 ± 0.18 | 13.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, S.; Farinas, E.T. Laccase and Its Mutant Displayed on the Bacillus subtilis Spore Coat for Oxidation of Phenolic Compounds in Organic Solvents. Catalysts 2021, 11, 606. https://doi.org/10.3390/catal11050606
Sheng S, Farinas ET. Laccase and Its Mutant Displayed on the Bacillus subtilis Spore Coat for Oxidation of Phenolic Compounds in Organic Solvents. Catalysts. 2021; 11(5):606. https://doi.org/10.3390/catal11050606
Chicago/Turabian StyleSheng, Silu, and Edgardo T. Farinas. 2021. "Laccase and Its Mutant Displayed on the Bacillus subtilis Spore Coat for Oxidation of Phenolic Compounds in Organic Solvents" Catalysts 11, no. 5: 606. https://doi.org/10.3390/catal11050606
APA StyleSheng, S., & Farinas, E. T. (2021). Laccase and Its Mutant Displayed on the Bacillus subtilis Spore Coat for Oxidation of Phenolic Compounds in Organic Solvents. Catalysts, 11(5), 606. https://doi.org/10.3390/catal11050606