Biosynthesis of Putrescine from L-arginine Using Engineered Escherichia coli Whole Cells


Abstract

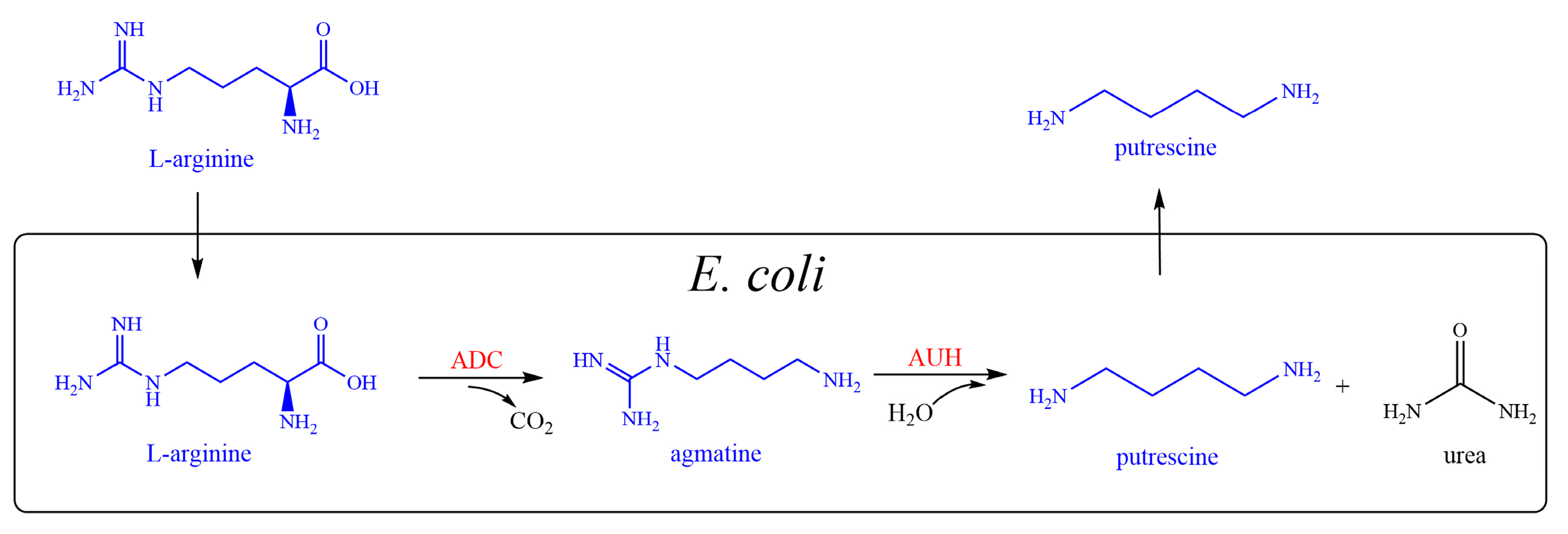
1. Introduction
2. Results
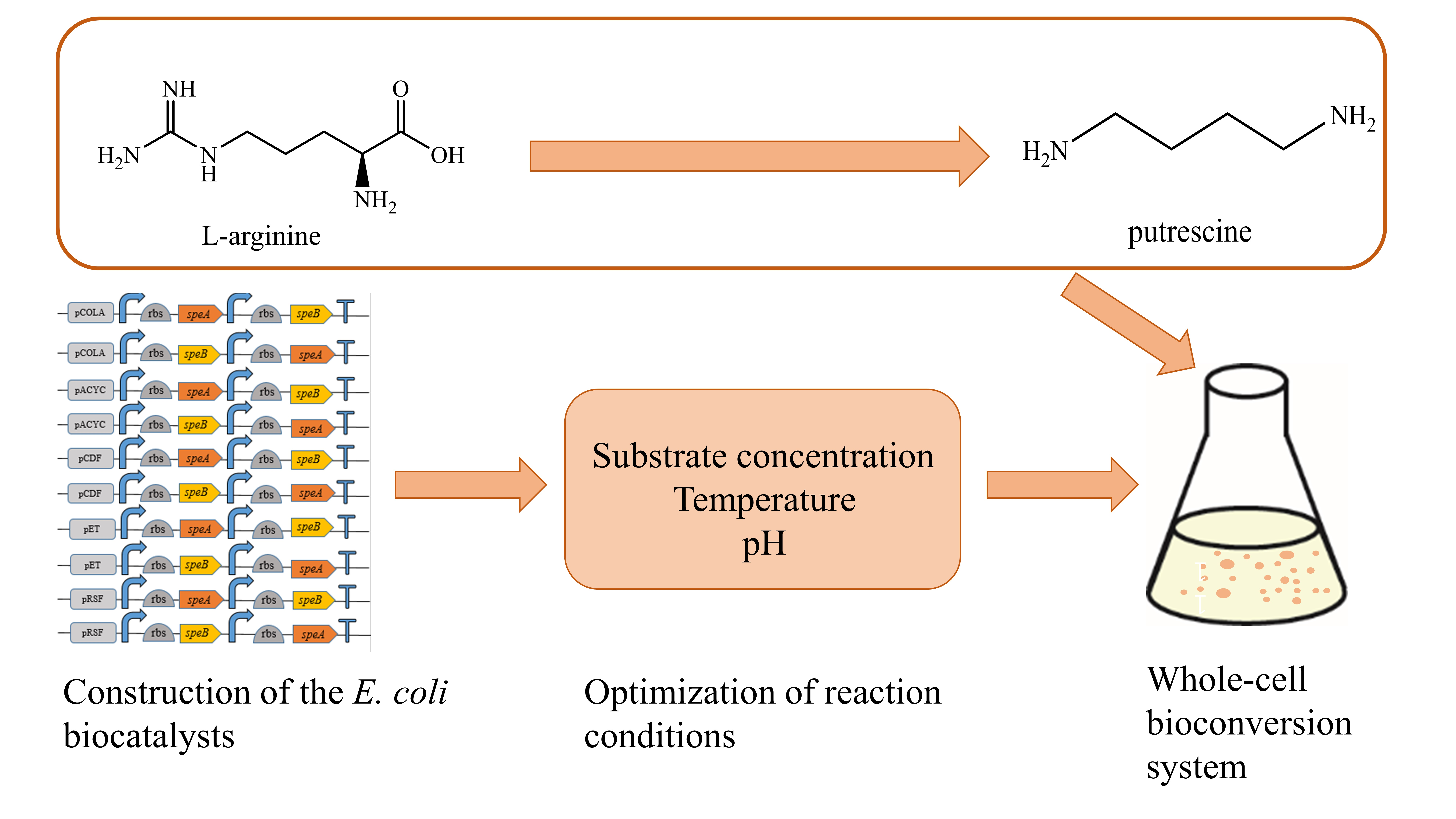
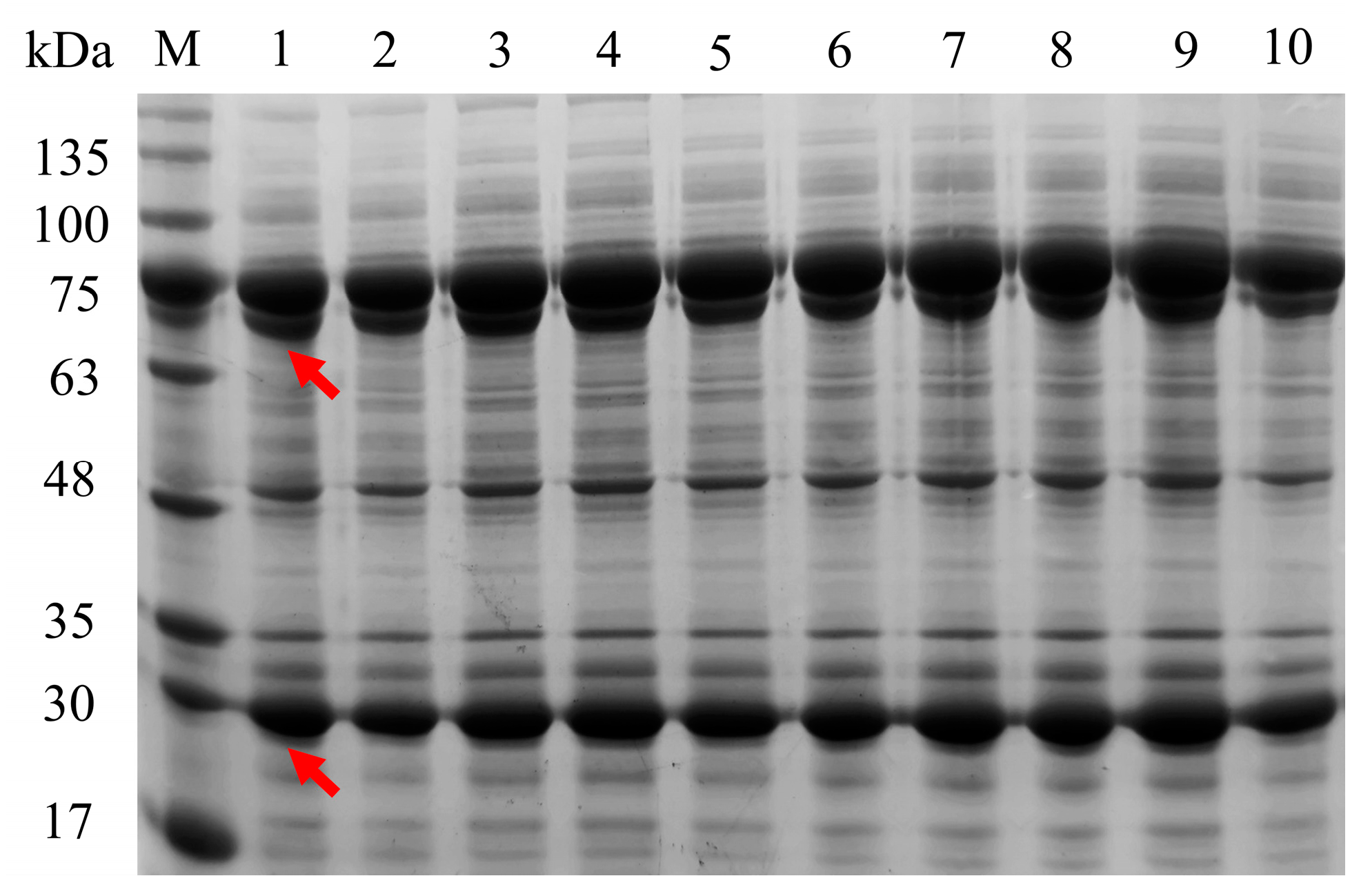
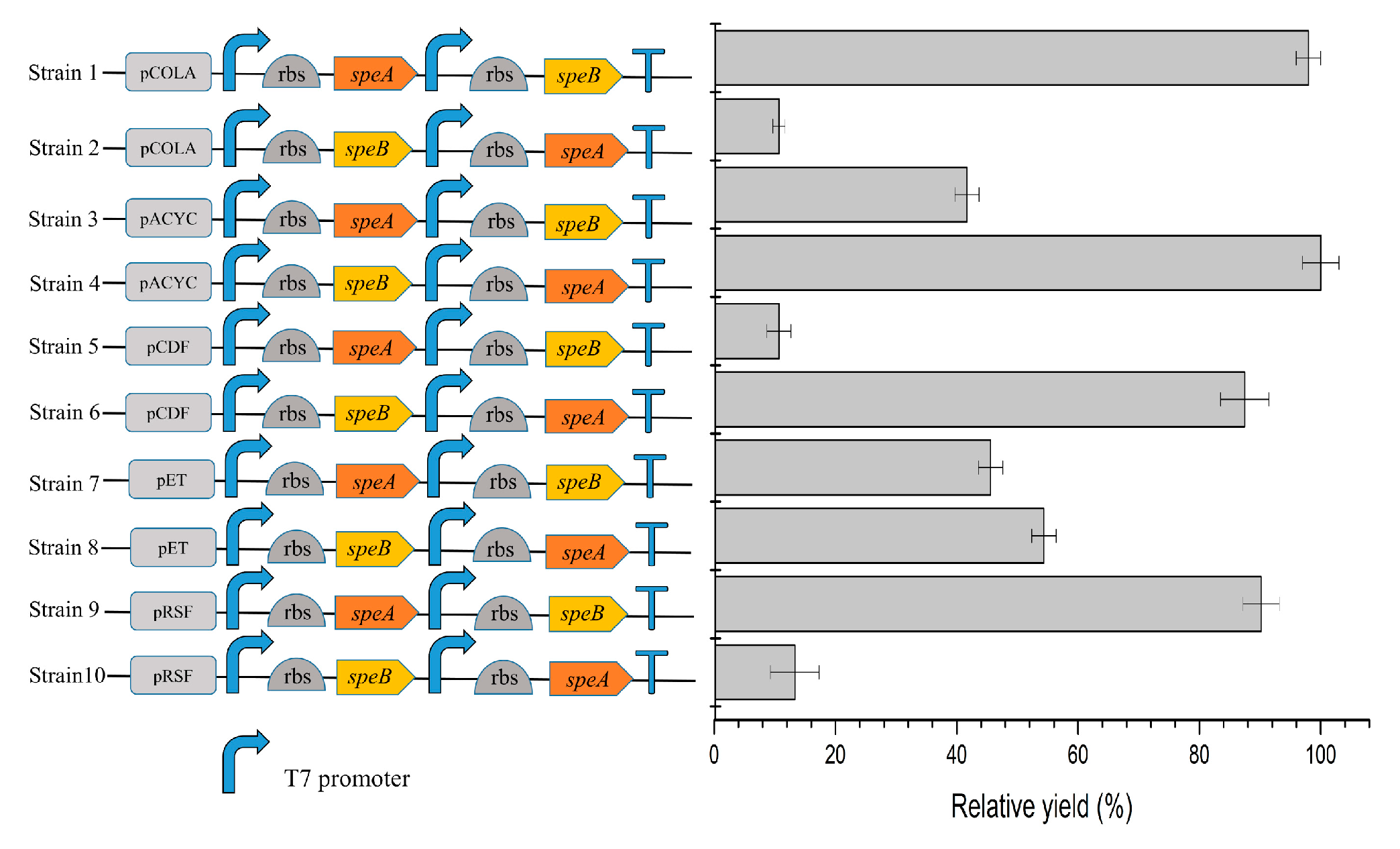
2.1. Construction of E. coli Whole-Cell Biocatalysts
2.2. Comparison of Different Strains for Putrescine Productiond
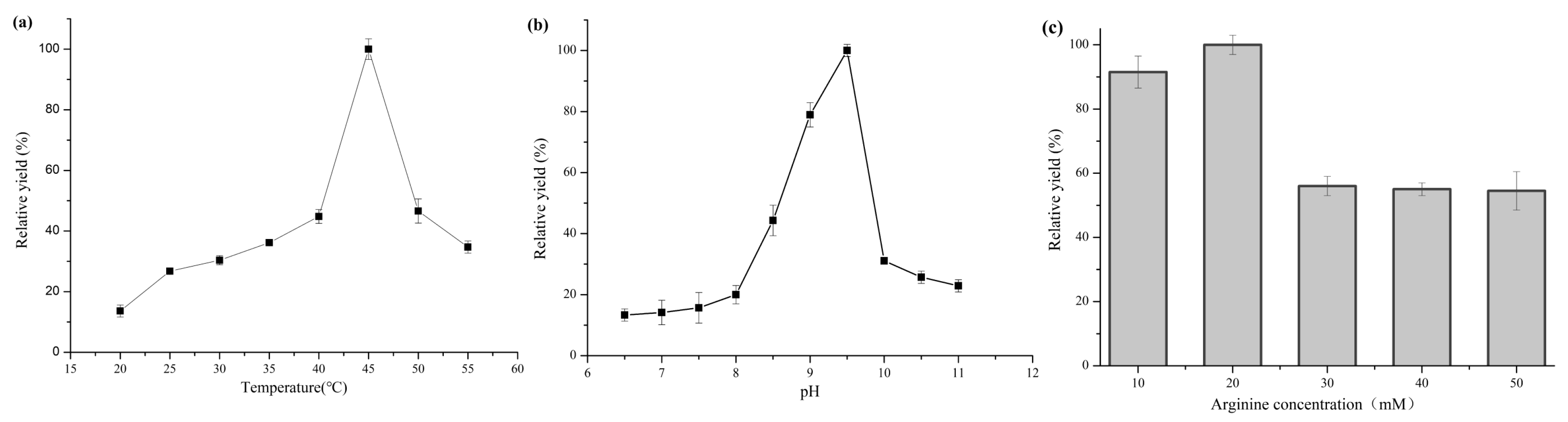
2.3. Optimization of Whole-Cell Biotransformation Conditions
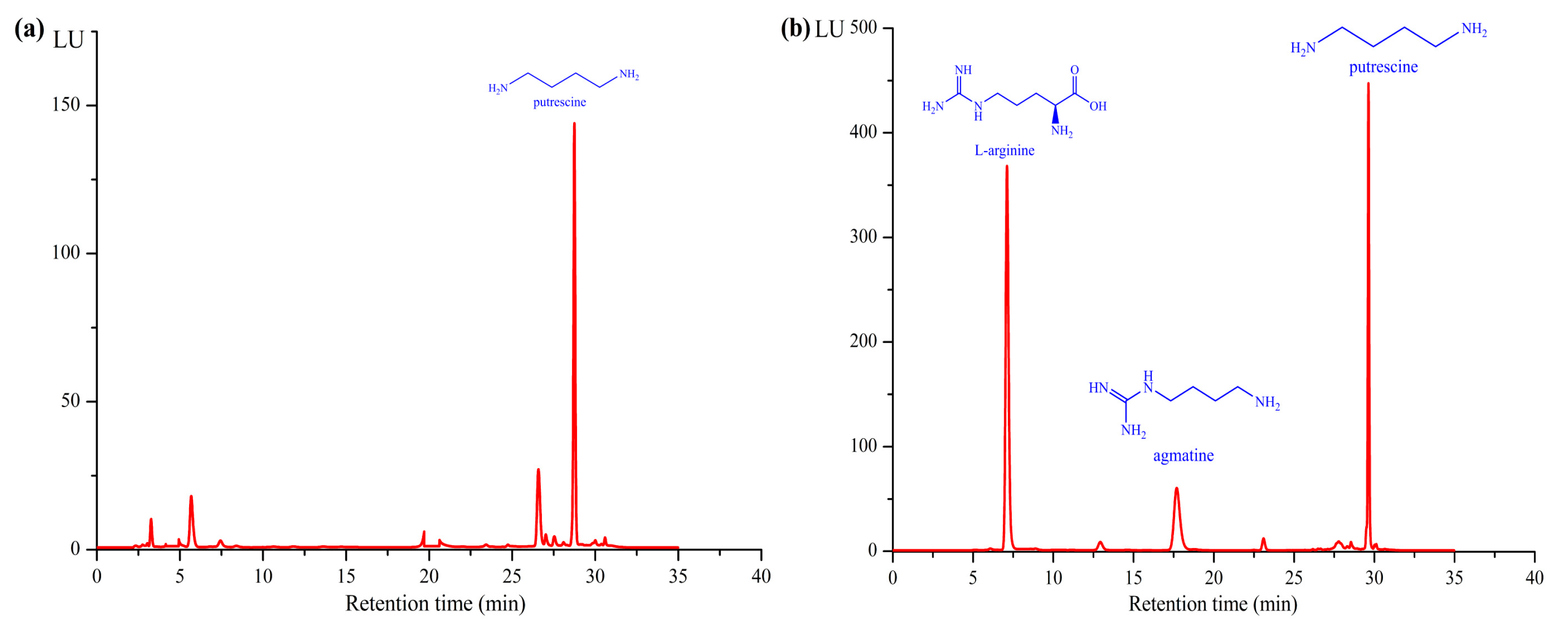
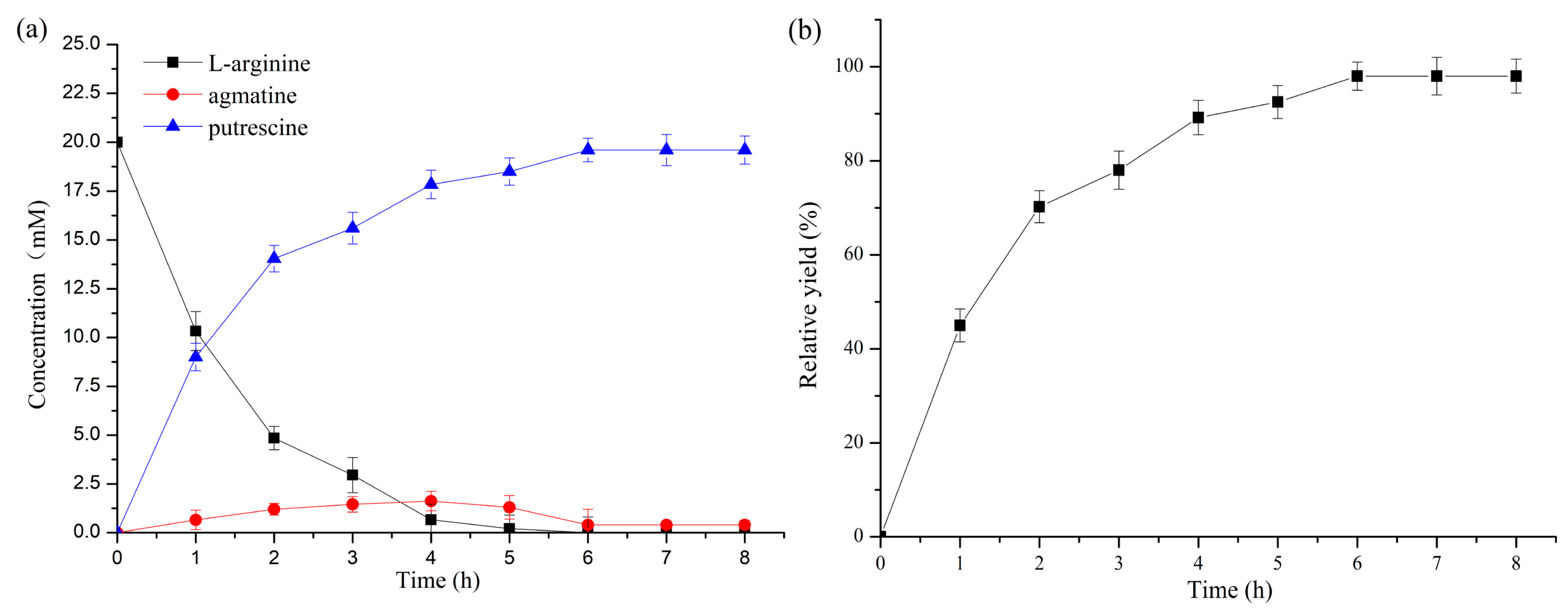
2.4. Time Course of Putrescine Production
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, Other Materials
4.2. Pathway and Plasmid Construction
4.3. Culture Conditions and Preparation of Whole-Cell Biocatalysts
4.4. SDS-PAGE Analysis
4.5. Whole-Cell Biocatalysts and Optimization of Reaction Conditions
4.6. Analytical Methods
Author Contributions
Funding
Conflicts of Interest
References
- Sugiyama, Y.; Nakamura, A.; Matsumoto, M.; Kanbe, A.; Sakanaka, M.; Higashi, K.; Igarashi, K.; Katayama, T.; Suzuki, H.; Kurihara, S. A novel putrescine exporter sapBCDF of Escherichia coli. J. Biol. Chem. 2016, 291, 26343–26351. [Google Scholar] [CrossRef] [PubMed]
- Wallace, H.M.; Fraser, A.V.; Hughes, A. A perspective of polyamine metabolism. Biochem. J. 2003, 376, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E.; McCann, P.P. Polyamine metabolism and function. Am. J. Physiol.-Cell Physiol. 1982, 243, C212–C221. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zheng, J.; Wu, X.; Xu, X.; Jia, G.; Zhao, H.; Chen, X.; Wu, C.; Tian, G.; Wang, J. Putrescine enhances intestinal immune function and regulates intestinal bacteria in weaning piglets. Food Funct. 2019, 10, 4134–4142. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Nara, M.; Sakanaka, M.; Gotoh, A.; Kitakata, A.; Okuda, S.; Kurihara, S. Comprehensive analysis of polyamine transport and biosynthesis in the dominant human gut bacteria: Potential presence of novel polyamine metabolism and transport genes. Int. J. Biochem. Cell Biol. 2017, 93, 52–61. [Google Scholar] [CrossRef]
- Chen, D.D.; Shao, Q.S.; Yin, L.H.; Younis, A.; Zheng, B.S. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant. Sci. 2019, 9, 13. [Google Scholar] [CrossRef]
- Yuan, Y.H.; Zhong, M.; Du, N.S.; Shu, S.; Sun, J.; Guo, S.R. Putrescine enhances salt tolerance of cucumber seedlings by regulating ion homeostasis. Environ. Exp. Bot. 2019, 165, 70–82. [Google Scholar] [CrossRef]
- Zheng, Q.L.; Zuo, J.H.; Gu, S.T.; Gao, L.P.; Hu, W.Z.; Wang, Q.; Jiang, A.L. Putrescine treatment reduces yellowing during senescence of broccoli (Brassica oleracea L. var. italica). Postharvest Biol. Technol. 2019, 152, 29–35. [Google Scholar] [CrossRef]
- Scott, E.; Peter, F.; Sanders, J. Biomass in the manufacture of industrial products—The use of proteins and amino acids. Appl. Microbiol. Biotechnol. 2007, 75, 751–762. [Google Scholar] [CrossRef]
- Qian, Z.G.; Xia, X.X.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of putrescine: A four carbon diamine. Biotechnol. Bioeng. 2009, 104, 651–662. [Google Scholar] [CrossRef]
- Yamanobe, T.; Kurihara, Y.; Uehara, H.; Komoto, T. Structure and characterization of nylon 46. J. Mol. Struct. 2007, 829, 80–87. [Google Scholar] [CrossRef]
- Choi, H.; Kyeong, H.-H.; Choi, J.M.; Kim, H.-S. Rational design of ornithine decarboxylase with high catalytic activity for the production of putrescine. Appl. Microbiol. Biotechnol. 2014, 98, 7483–7490. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Wendisch, V.F.J.A.M. Putrescine production by engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2010, 88, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Eberhardt, D.; Wendisch, V.F. Improving putrescine production by Corynebacterium glutamicum by fine-tuning ornithine transcarbamoylase activity using a plasmid addiction system. Appl. Microbiol. Biotechnol. 2012, 95, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.Q.D.; Schneider, J.; Reddy, G.K.; Wendisch, V.F. Fermentative production of the diamine putrescine: System metabolic engineering of Corynebacterium glutamicum. Metabolites 2015, 5, 211–231. [Google Scholar] [CrossRef]
- Nguyen, A.Q.D.; Schneider, J.; Wendisch, V.F. Elimination of polyamine N-acetylation and regulatory engineering improved putrescine production by Corynebacterium glutamicum. J. Biotechnol. 2015, 201, 75–85. [Google Scholar] [CrossRef]
- Meiswinkel, T.M.; Gopinath, V.; Lindner, S.N.; Nampoothiri, K.M.; Wendisch, V.F. Accelerated pentose utilization by Corynebacterium glutamicum for accelerated production of lysine, glutamate, ornithine and putrescine. Microb. Biotechnol. 2013, 6, 131–140. [Google Scholar] [CrossRef]
- Del Rio, B.; Linares, D.M.; Ladero, V.; Redruello, B.; Fernandez, M.; Martin, M.C.; Alvarez, M.A. Putrescine production via the agmatine deiminase pathway increases the growth of Lactococcus lactis and causes the alkalinization of the culture medium. Appl. Microbiol. Biotechnol. 2015, 99, 897–905. [Google Scholar] [CrossRef]
- Srinivasan, P.; Smolke, C.D. Engineering a microbial biosynthesis platform for de novo production of tropane alkaloids. Nat. Commun. 2019, 10, 15. [Google Scholar] [CrossRef]
- France, S.P.; Hepworth, L.J.; Turner, N.J.; Flitsch, S.L. Constructing biocatalytic cascades: In vitro and in vivo approaches to de novo multi-enzyme pathways. ACS Catal. 2017, 7, 710–724. [Google Scholar] [CrossRef]
- Li, C.; Jia, P.; Bai, Y.; Fan, T.P.; Zheng, X.; Cai, Y. Efficient synthesis of hydroxytyrosol from l-3,4-dihydroxyphenylalanine using engineered Escherichia coli whole cells. J. Agric. Food Chem. 2019, 67, 6867–6873. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Jiang, J.; Bai, Y.; Fan, T.-p.; Zhao, Y.; Zheng, X.; Cai, Y. Biosynthesis of D-danshensu from L-DOPA using engineered Escherichia coli whole cells. Appl. Microbiol. Biotechnol. 2019, 103, 6097–6105. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Zhou, C.; Liu, R.; Wu, X.; Wu, D.; Hu, X.; Ding, Y. Expression and purification of recombinant arginine decarboxylase (speA) from Escherichia coli. Mol. Biol. Rep. 2010, 37, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Satishchandran, C.; Boyle, S.M. Purification and properties of agmatine ureohydrolyase, a putrescine biosynthetic enzyme in Escherichia coli. J. Bacteriol. 1986, 165, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Wu, J.; Zhan, X.; Zhu, L.; Zheng, Z.; Gao, M. Phosphoenolpyruvate-supply module in Escherichia coli improves N-acetyl-d-neuraminic acid biocatalysis. Biotechnol. Lett. 2017, 39, 227–234. [Google Scholar] [CrossRef]
- Bentley, W.E.; Mirjalili, N.; Andersen, D.C.; Davis, R.H.; Kompala, D.S. Plasmid-encoded protein: The principal factor in the “metabolic burden” associated with recombinant bacteria. Biotechnol. Bioeng. 2009, 102, 1284–1297. [Google Scholar] [CrossRef]
- Jones, K.L.; Kim, S.-W.; Keasling, J.D. Low-copy plasmids can perform as well as or better than high-copy plasmids for metabolic engineering of bacteria. Metab. Eng. 2000, 2, 328–338. [Google Scholar] [CrossRef]
- Morris, D.R.; Koffron, K.L. Putrescine biosynthesis in Escherichia coli. regulation through pathway selection. J. Biol. Chem. 1969, 244, 6094–6099. [Google Scholar]
- Wu, J.; Du, G.; Zhou, J.; Chen, J. Metabolic engineering of Escherichia coli for (2S)-pinocembrin production from glucose by a modular metabolic strategy. Metab. Eng. 2013, 16, 48–55. [Google Scholar] [CrossRef]
- Hou, Y.; Hossain, G.S.; Li, J.; Shin, H.D.; Du, G.; Chen, J.; Liu, L. Metabolic engineering of cofactor flavin adenine dinucleotide (FAD) synthesis and regeneration in Escherichia coli for production of alpha-keto acids. Biotechnol. Bioeng. 2017, 114, 1928–1936. [Google Scholar] [CrossRef]
- Wu, W.H.; Morris, D.R. Biosynthetic arginine decarboxylase from Escherichia coli. purification and properties. J. Biol. Chem. 1973, 248, 1687–1695. [Google Scholar] [PubMed]
- Forouhar, F.; Lew, S.; Seetharaman, J.; Xiao, R.; Acton, T.B.; Montelione, G.T.; Tong, L.A. Structures of bacterial biosynthetic arginine decarboxylases. Acta Crystallogr. F-Struct. Biol. Cryst. Commun. 2010, 66, 1562–1566. [Google Scholar] [CrossRef] [PubMed]
- Ginesy, M.; Belotserkovsky, J.; Enman, J.; Isaksson, L.; Rova, U. Metabolic engineering of Escherichia coli for enhanced arginine biosynthesis. Microb. Cell. Fact. 2015, 14, 29. [Google Scholar] [CrossRef] [PubMed]
- Najmi, Z.; Ebrahimipour, G.; Franzetti, A.; Banat, I.M. In situ downstream strategies for cost-effective bio/surfactant recovery. Biotechnol. Appl. Biochem. 2018, 65, 523–532. [Google Scholar] [CrossRef]
- Yildirim, H.K.; Uren, A.; Yucel, U. Evaluation of biogenic amines in organic and non-organic wines by HPLC OPA derivatization. Food Technol. Biotechnol. 2007, 45, 62–68. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences, 5′-3′ |
|---|---|
| Site1-speA-F | TTAAGTATAAGAAGGAGATATACATATGAGCACCTTAGGTCATCAATACG |
| Site1-speA-R | TTAAGCATTATGCGGCCGCAAGCTTTTACTCATCTTCAAGATAAGTATAAC |
| Site1-speB-F | TCACCACAGCCAGGATCCGAATTCGATGAGCACCTTAGGTCATCAATACG |
| Site1-speB-R | TTTCTTTACCAGACTCGAGGGTACCTTACTCGCCCTTTTTCGCCG |
| Site2-speA-F | TTAAGTATAAGAAGGAGATATACATATGTCTGACGACATGTCTATGGGT |
| Site2-speA-R | TTAAGCATTATGCGGCCGCAAGCTTTTACTCGCCCTTTTTCGCCGC |
| Site2-speB-F | TTAAGCATTATGCGGCCGCAAGCTTTTACTCATCTTCAAGATAAGTATAAC |
| Site2-speB-R | TTTCTTTACCAGACTCGAGGGTACCTTACTCATCTTCAAGATAAGTATAACC |
| Strains/Plasmids | Description | Source |
|---|---|---|
| Plasmids | ||
| pCOLADuet-1 | double T7 promoters, COLA ori, KanR | Novagen |
| pACYCDuet-1 | double T7 promoters, P15A ori, ChlR | Novagen |
| pCDFDuet-1 | double T7 promoters, CDF13 ori, SmR | Novagen |
| pETDuet-1 | double T7 promoters, pBR322 ori, AmpR | Novagen |
| pRSFDuet-1 | double T7 promoters, RSF ori, KanR | Novagen |
| pCOLADuet-speA-speB | pCOLADuet-1 carrying speA and speB | this study |
| pCOLADuet-speB-speA | pCOLADuet-1 carrying speB and speA | this study |
| pACYCDuet-speA-speB | pACYCDuet-1 carrying speA and speB | this study |
| pACYCDuet-speB-speA | pACYCDuet-1 carrying speB and speA | this study |
| pCDFDuet-speA-speB | pCDFDuet-1 carrying speA and speB | this study |
| pCDFDuet-speB-speA | pCDFDuet-1 carrying speB and speA | this study |
| pETDuet-speA-speB | pETDuet-1 carrying speA and speB | this study |
| pETDuet-speB-speA | pETDuet-1 carrying speB and speA | this study |
| pRSFDuet-speA-speB | pRSFDuet-1 carrying speA and speB | this study |
| pRSFDuet-speB-speA | pRSFDuet-1 carrying speB and speA | this study |
| Strains | ||
| strain 1 | E. coli BL21 (DE3)/pACYCDuet-speA-speB | this study |
| strain 2 | E. coli BL21 (DE3)/pACYCDuet-speB-speA | this study |
| strain 3 | E. coli BL21 (DE3)/pCOlADuet- speA-speB | this study |
| strain 4 | E. coli BL21 (DE3)/pCOlADuet- speB-speA | this study |
| strain 5 | E. coli BL21 (DE3)/pETDuet- speA-speB | this study |
| strain 6 | E. coli BL21 (DE3)/pETDuet- speB-speA | this study |
| strain 7 | E. coli BL21 (DE3)/pCDFDuet- speA-speB | this study |
| strain 8 | E. coli BL21 (DE3)/pCDFDuet- speB-speA | this study |
| strain 9 | E. coli BL21 (DE3)/pRSFDuet- speA-speB | this study |
| strain 10 | E. coli BL21 (DE3)/pRSFDuet- speB-speA | this study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hui, H.; Bai, Y.; Fan, T.-P.; Zheng, X.; Cai, Y. Biosynthesis of Putrescine from L-arginine Using Engineered Escherichia coli Whole Cells. Catalysts 2020, 10, 947. https://doi.org/10.3390/catal10090947
Hui H, Bai Y, Fan T-P, Zheng X, Cai Y. Biosynthesis of Putrescine from L-arginine Using Engineered Escherichia coli Whole Cells. Catalysts. 2020; 10(9):947. https://doi.org/10.3390/catal10090947
Chicago/Turabian StyleHui, Hongjie, Yajun Bai, Tai-Ping Fan, Xiaohui Zheng, and Yujie Cai. 2020. "Biosynthesis of Putrescine from L-arginine Using Engineered Escherichia coli Whole Cells" Catalysts 10, no. 9: 947. https://doi.org/10.3390/catal10090947
APA StyleHui, H., Bai, Y., Fan, T.-P., Zheng, X., & Cai, Y. (2020). Biosynthesis of Putrescine from L-arginine Using Engineered Escherichia coli Whole Cells. Catalysts, 10(9), 947. https://doi.org/10.3390/catal10090947