1. Introduction
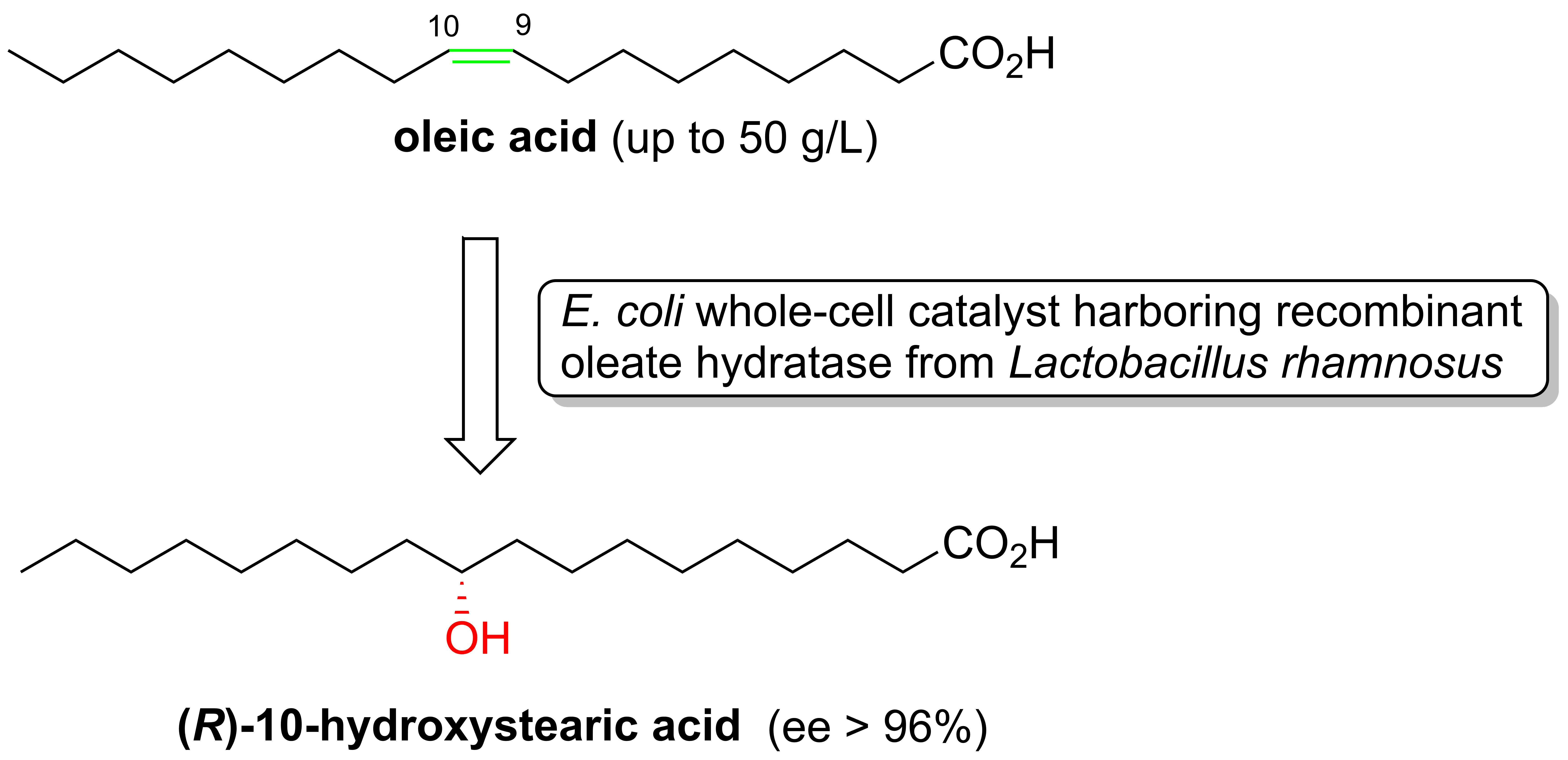
The enzymatic addition of water to the double bond of free fatty acids was first described, in 1962, by Wallen et al. [
1] in the course of a study on the ability of the
Pseudomonas sp. strain 3266 to catalyze the conversion of oleic acid
1 (OA) into 10-hydroxystearic acid
4 (10-HSA) (
Figure 1).
Afterward, several microorganisms were identified as possessing this particular capability [
2,
3,
4,
5,
6,
7], including different probiotics species [
8]. This hydration reaction is catalyzed by the enzymes oleate hydratase (EC 4.2.1.53), whose first isolation and characterization was performed by Hager [
9], almost half a century after the publication of Wallen’s research. Since the enzyme identification, a large number of new oleate hydratases (OLHs) have been discovered [
10,
11,
12,
13,
14,
15,
16,
17,
18,
19,
20,
21,
22,
23,
24,
25,
26,
27,
28,
29,
30,
31,
32]. These recent studies, besides describing the new enzymes’ structures, reported on their catalytic activity either in terms of substrate specificity or the stereoselectivity of the hydration reaction.
The physiological function of this particular enzymatic class is still unclear. In fact, only a few studies have speculated on the protective role of these enzymes against potentially toxic unsaturated fatty acids [
10,
20,
33,
34].
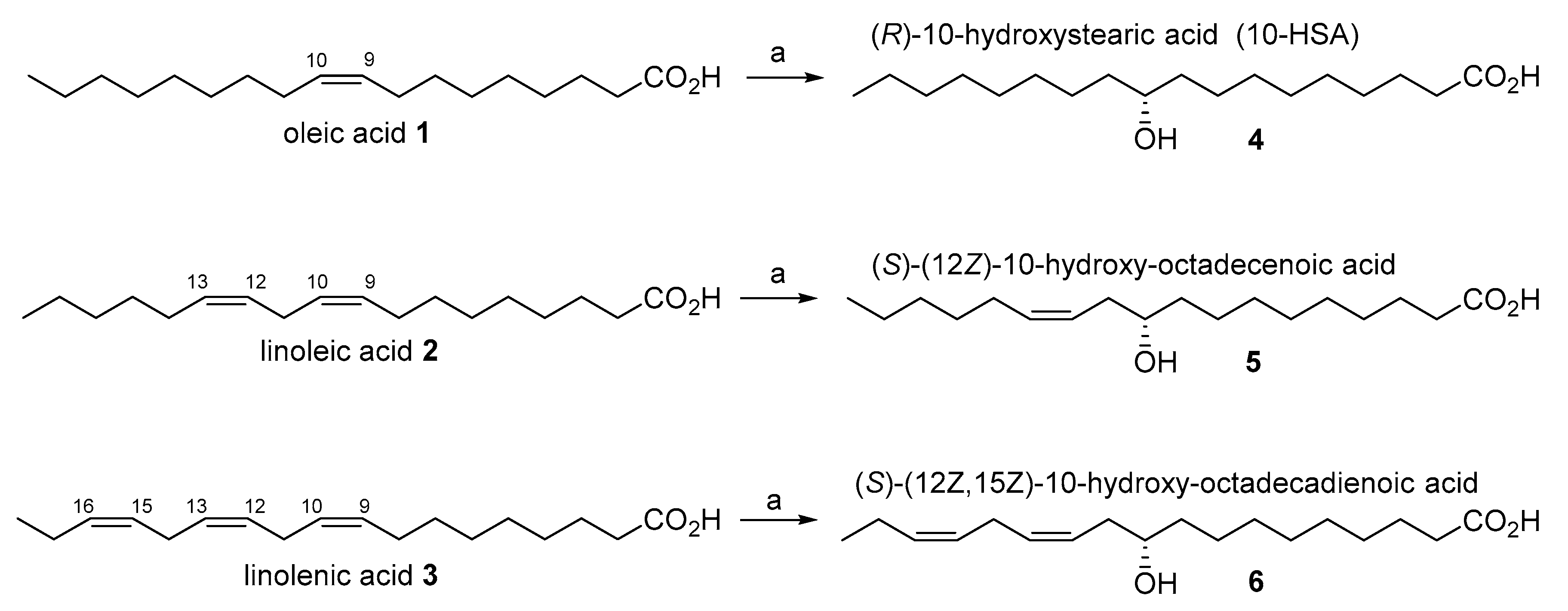
The most common unsaturated fatty acids (UFAs), shown in
Figure 1, are oleic (
1), linoleic (
2) and linolenic (
3) acid, and they are the main components (as triglycerides) of the vegetable oils used for human consumption.
From an applicative point of view, hydroxy fatty acids (HFAs) are important compounds used for many industrial applications, such as starting materials for biodegradable polymers, lubricants, emulsifiers, drugs, cosmetic ingredients, and flavors [
2,
13,
30,
35,
36,
37,
38,
39,
40,
41].
For example, 12-hydroxystearic acid (12-HSA), as well as 10-HSA, is mostly used as a thickening agent for the production of lubricants and skincare preparations [
35,
36,
37,
38,
39]; and ricinoleic acid is commonly used as starting material for the production of surfactants, resins and polymers (nylon-11) [
40,
41].
Moreover, HFAs and some of their derivatives also possess relevant biological activities. In fact, hydroxy fatty acids are plant self-defense substances [
42] and are important in the control of allergy, inflammation and immunity [
43,
44]. Recently, particular esters of hydroxy fatty acids possessing anti-diabetic and anti-inflammatory activities were also identified in bioactive lipids (FAHFAs) [
45].
From a synthetic point of view, a number of chemical reactions can be used to hydrate the easily available UFAs 1–3. Unfortunately, these transformations lack stereo- and regioselectivity, which afford complex mixtures of isomers. In the simplest case, the chemical hydration of oleic acid leads to a mixture of two racemic isomers, namely, 9-HSA and 10-HSA. Of course, the chemical hydration of polyunsaturated fatty acids leads to a much more complex mixtures of HFAs, including the possibility of the formation of diols and triols.
Otherwise, fatty acid hydratases are often highly selective and are able to introduce the hydroxyl group in a specific position of the UFAs carbon chain. Few studies have been carried out about the feasibility of using these enzymes from an applicative point of view [
21].
We have previously demonstrated the versatility of the probiotic bacteria
Lactobacillus rhamnosus ATCC 53103 [
6], which is able to hydrate oleic, linoleic and linolenic acid with high stereoselectivity and in good yields. More specifically, the latter strain catalyzes the hydration reaction exclusively affording the corresponding 10-HFA derivatives, without formation of other regioisomers or polyhydroxylated fatty acids. In addition, oleic, linoleic and linolenic acid are transformed into (
R)-10-hydroxystearic acid (
4), (
S)-(12
Z)-10-hydroxy-octadecenoic acid (
5) and (
S)-(12
Z,15
Z)-10-hydroxy-octadecadienoic acid (
6), respectively, in very high enantiomeric purity (ee > 95%).
It is worth noting that the aforementioned
Lactobacillus strain hydrates the three UFAs with identical stereoselectivity. The descriptor switch from (
R) form of hydroxystearic acid to the (
S) form of (12
Z)-10-hydroxy-octadecenoic and (12
Z,15
Z)-10-hydroxy-octadecadienoic acid is due only to a change of substituent priority, according to the Cahn-Ingold-Prelog rules. Most likely, the oleate hydratase(s) produced by
Lactobacillus rhamnosus accepts as substrates different unsaturated fatty acids, which must have a (Z) Δ
9–10 double bond as the sole mandatory requirement. It is reasonable that the biocatalytic activity of this
Lactobacillus strain was due to the expression of an oleate hydratase previously identified from the bacteria genome of the same strain [
15].
In order to enlarge our knowledge in using OLHs as suitable catalyst in organic synthesis, we decided to clone and overexpress the latter enzyme in a heterologous host, using specific primers designed from the deposited sequence of this enzyme. In addition, we studied the best expression condition using Escherichia coli BL21(DE3) as a host system. Then, the obtained biocatalyst was exploited in order to devise a reliable experimental procedure for the hydration of oleic acid.
Overall, this study deals with two topics. The first concerns the experimental sequence of gene amplification, cloning in a heterologous host, protein overexpression and hydratase purification. The second focuses on the study of the catalytic properties of the obtained enzyme. More specifically, we evaluated both the activity and the regio- and stereoselectivity related to the oleic acid hydration reaction, using the obtained oleate hydratase as biocatalyst.
2. Results and Discussion
2.1. Gene Identification and Cloning System
Oleate hydratase from
Lactobacillus rhamnosus ATCC 53103 was selected due to the positive results obtained with this strain in converting OA into 10-HSA [
6]. In addition, this particular microorganism was well characterized from a genetic point of view because it is one of the most common probiotic strains used for human health. In a previous work by Yang et al. [
15], a myosin-cross-reactive antigen of
L. rhamnosus ATCC 53103 was classified as oleate hydratase (OLH), but no studies were carried out about the possibility of using the enzyme as a suitable catalyst for biotechnological applications. Therefore, we started from the deposited sequence (GenBank: WP_005714981), and we cloned the enzyme in
Escherichia coli BL21(DE3) as described in the materials and method section. The system used is based on the inducible T7 expression system with an improvement control over gene expression.
The pETite C-His vectors facilitate enzyme-free cloning of target genes and are provided in a linearized form. Following the amplification of the target gene with appropriate primers, the polymerase chain reaction (PCR) product is mixed with the pETite vector and transformed directly into chemically competent HI-Control 10G cells for its amplification.
2.2. Expression Condition of Recombinant OLH
In order to understand the best expression conditions of the enzyme, we evaluated the two most common induction systems, which are based on the use of IPTG (isopropyl-β-D-1-thiogalactopyranoside) and on the use of an autoinduction medium.
IPTG induction presents a number of distinct advantages, especially in small-scale experiments. Unlike lactose and other galactosides, IPTG is not metabolized by the cells, and this ensures that the level of induction remains constant following its addition to the growth medium. The main drawback related to the use of IPTG lies in its very strong induction effect, which should be modulated in order to avoid a too quick protein expression that could end-up in the production of misfolded enzymes.
Otherwise, an autoinduction protein expression system could be useful from an industrial point of view because it permits a faster and easier management of the cell culture, even though it is hard to fine-tune the level of expression.
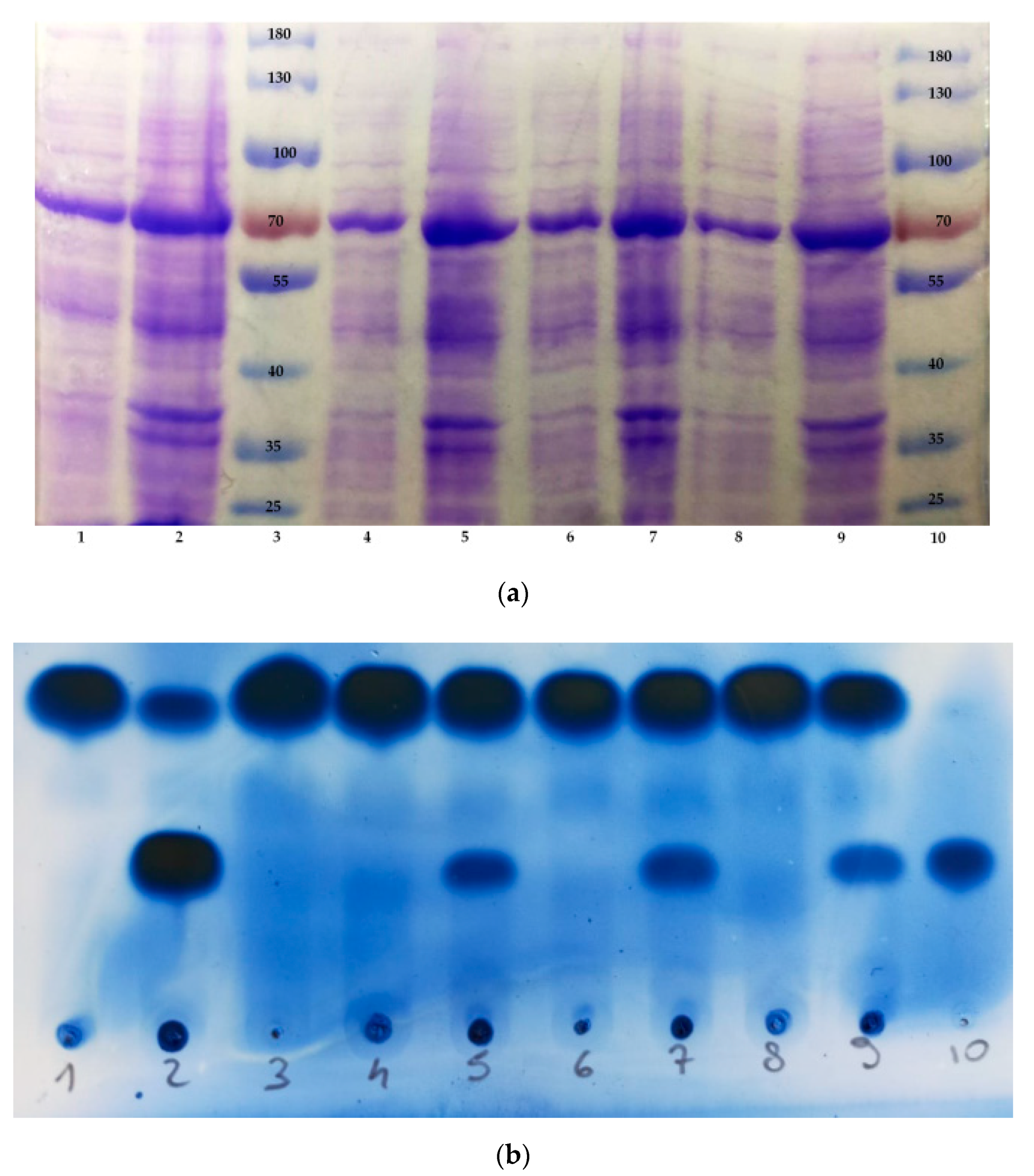
In order to identify the best induction conditions, we set up a series of experiments in which we compared the relative concentration and activity of the OLH produced by recombinant E. coli cells under different experimental conditions. Accordingly, we performed four expression experiments: three in which we used IPTG as an inducer at three different concentrations (0.1, 0.5 and 1 mM, respectively) and one in which the recombinant E. coli cells were grown in the lactose-based autoinduction medium ZYM-5052. The cells were collected by centrifugation, and each one of the four samples was suspended in a suitable buffer solution, sonicated and then centrifuged again to obtain a clear lysate solution and a cellular debris pellet.
The sodium dodecyl sulphate—polyacrylamide gel electrophoresis (SDS-PAGE) analysis of the obtained eight samples (
Figure 2a) confirms the presence of an overexpression band due to a protein with molecular weight comprised between 70 and 55 KDa. Since the OLH previously identified in
Lactobacillus rhamnosus ATCC 53103 shows a molecular weight of 64,444 Da, the SDS-PAGE analysis gave proof of the successful expression of the hydratase in
E. coli cells. Another noteworthy observation concerns the efficiency of the enzyme production under the different expression conditions. Overall, the SDS-PAGE analysis did not show strong evidence in term of differences in protein expression when using either IPTG or the ZYM-5052 medium. In fact, the hydratase is detectable in all the samples deriving from the different expression conditions used in this work.
Completely different results were obtained when testing the enzyme activity. The same eight samples analyzed by SDS-PAGE were used to catalyze the hydration of oleic acid in order to afford 10-HSA. The reaction conversion was monitored by thin layer chromatography (TLC) analysis (
Figure 2b). We assume that after a given time span, using the same starting concentration of oleic acid and using the same amount of biocatalyst, the measured 10-HSA/OA ratio is proportional to the reaction rate and thus to the enzyme activity. We observed that the soluble hydratase, contained in the clear lysate samples, was almost completely inactive (lanes 1, 4, 6, 8), whereas the cells debris samples possessed OLH activity, which was dependent on the kind of expression system. More specifically, the activity of the enzyme contained in the debris derived from cells grown in ZYM 5052 medium (lane 2) was much higher than that observed from cells induced with IPTG (lanes 5, 7, 9). In addition, the increasing IPTG concentration seemed neither to affect the protein production (
Figure 2a, lanes 4–9) nor its activity (
Figure 2b, lanes 5, 7, 9), as demonstrated by the very similar concentration and activity of the produced hydratase.
Considering these results, we decided to use ZYM-5052 as medium of choice for OLH expression. Once this point had been established, we examined the potential of the obtained biocatalyst in preparative organic chemistry.
2.3. Use of the Recombinant OLH as Biocatalyst for the Hydration of Oleic Acid
As described above, the most suitable way to obtain the active form of the OLH from Lactobacillus rhamnosus is based on growing the recombinant E. coli cells in ZYM 5052 medium. Hence, we decided to evaluate the best way to use the obtained biocatalyst for the synthesis of 10-(R)-hydroxystearic acid starting from OA. There are different opportunities and different purity grades of the catalyst depending on the aim of the users. After induction, the required hydratase is retained inside whole bacterial cells. Therefore, at least three different kind of OLH-based biocatalysts can be employed. The first is the whole-cell catalyst, which consists of the use of isolated bacterial cells, possibly treated with reagents that help to break-up the cell membranes and the stabilization of the enzyme consequently released. A second method is based on the ultrasonication of the bacteria and the separation of the soluble protein fraction from the cellular debris. The obtained clear solution contains the OLH and can be used as such to catalyze OA hydration reaction. Lastly, the purification of the latter protein mixture can afford pure OLH, which is the third kind of biocatalyst we evaluated.
OLH is a flavin adenine dinucleotide (FAD) dependent enzyme, in which FAD is noncovalently bound to the conserved N-terminal nucleotide and works as a structure stabilizer. This consideration is extremely important because the deficiency of this cofactor can lead to a completely inactive form of the enzyme [
17,
27].
For this reason, the use of recombinant E. coli BL21(DE3) as resting or still growing cells, as well as the employment of the soluble fraction of sonicated cells, could be a good way to avoid the FAD loss from the biocatalyst, thus preserving the native form of the enzyme.
We also isolated the pure enzyme through Ni Sepharose 6 Fast Flow agarose resin (Ni-NTA) chromatography, but we observed protein precipitation after dialysis. In addition, the purified OLH was completely inactive and the catalytic activity was restored only after the addition of an equivalent mole of FAD in the reaction mixture.
In this context, we decided to evaluate the OLH activity of the three above-described biocatalysts. More specifically, we evaluated the efficiency of the hydration reaction using OA as a substrate. We employed both a resting cell catalyst, a soluble crude extract catalyst derived from the ultrasonication of the recombinant E. coli cells and pure isolated OLH. To this end, we performed a number of experiments in which OA was treated with the above-described biocatalysts. Reaction time, temperature, pH and shaking speed were kept identical for all trials. For the resting cells catalyst, each experiment made use of the same cell concentration. Accordingly, for the reactions performed using an enzyme solution, we added an amount of catalyst that was the same as that obtainable by sonication of the biomass used in the whole-cell experiments.
The formation of 10-HSA was quantified by GC-MS analysis, and we assumed that the measured transformation yields were proportional to the enzymatic activity.
Table 1 shows the results obtained from the biotransformation experiments.
Our results demonstrated that the isolated form of OLH is not suitable for biotechnological purposes, at least from a preparative point of view. As discussed above, the purified OLH is completely inactive. Even after addition of FAD, the catalytic activity remained very low, allowing only 10% of OA hydration. Similarly, the soluble crude extract catalyst derived from the ultrasonication of the recombinant bacterial cells was confirmed to possess hydratase activity, but 10-HSA was not produced in yields higher than 15%. This probably means that OLH is poorly soluble or quickly loses its activity ability when outside the cells. These hypotheses are also corroborated by the SDS-PAGE analysis described above.
Otherwise, the use of recombinant E. coli BL21(DE3) as whole-cell biocatalyst (5% w/v) allows for obtaining higher amounts of 10-HSA. According to our findings, the biotransformation conditions seem to be crucial for the hydration reaction. In these cases, it is important to improve the oleic acid solubility in water and enhance the permeability of the membranes to the substrate. For these reasons, we added to the reaction mixture a co-solvent (ethanol) and a nonionic surfactant (Triton™ X-100) or glycerol. In both cases, we noted an increased activity of the enzyme.
We set a number of preliminary experiments in which we employed a constant concentration of ethanol (2% v/v) combined with the addition either of the surfactant or of the glycerol at different concentrations. We established that the higher hydratase activity was reached with the combination ethanol (2% v/v)—TritonTM X-100 (0.5% v/v) or with ethanol (2% v/v)—glycerol (10% v/v). Using smaller or bigger amounts of these co-solvents led to a significant reduction of the enzyme activity, probably due to the membrane barrier effect (for lower concentration of co-solvents) and to altered solvents-catalyst interactions (for higher concentration of co-solvents).
As described in
Table 1, the use of sonicated bacterial cells afforded 10-HSA in 30% yield. The same cells, treated with (Triton™ X-100) and ethanol (2%
v/
v), allowed doubling of the conversion yield (60%), whereas the use of glycerol and ethanol led to almost complete hydration of oleic acid (98% yield).
Interestingly, the whole-cell biocatalyst used in the latter experimental conditions is poorly inhibited both by the substrate and by the product, tolerating high substrate load. In separated experiments (not reported in
Table 1), we were able to run hydration reactions using a starting OA concentration of 50 g/L, and we observed only a modest reduction of the enzymatic activity (60% conversion of OA into 10-HSA after a reaction time of 3 days).
According to these findings, we selected the combination of the whole-cells of recombinant E. coli BL21(DE3) with ethanol (2% v/v) and glycerol (10% v/v) as the most efficient and reliable biocatalyst for the hydration of oleic acid.
2.4. Optimal pH and Temperature for the OLH-Catalysed Production of 10-HSA from OA
Another important topic of our work concerns the determination of the optimum pH and temperature of OLH-catalyzed hydration. We evaluated these parameters by performing the biotransformation of OA into 10-HSA using the whole-cell procedure described in
Section 3.4 of this paper.
In general, recombinant E. coli BL21(DE3) cells (5% w/v) were resuspended in a 50 mM potassium phosphate buffer of pH 6.6 with 3 g/L of oleic acid and in the presence of ethanol (2% v/v) and glycerol (10% v/v). The biotransformations were incubated at different temperatures under shaking and analyzed after 24 h to assess the 10-HSA yields.
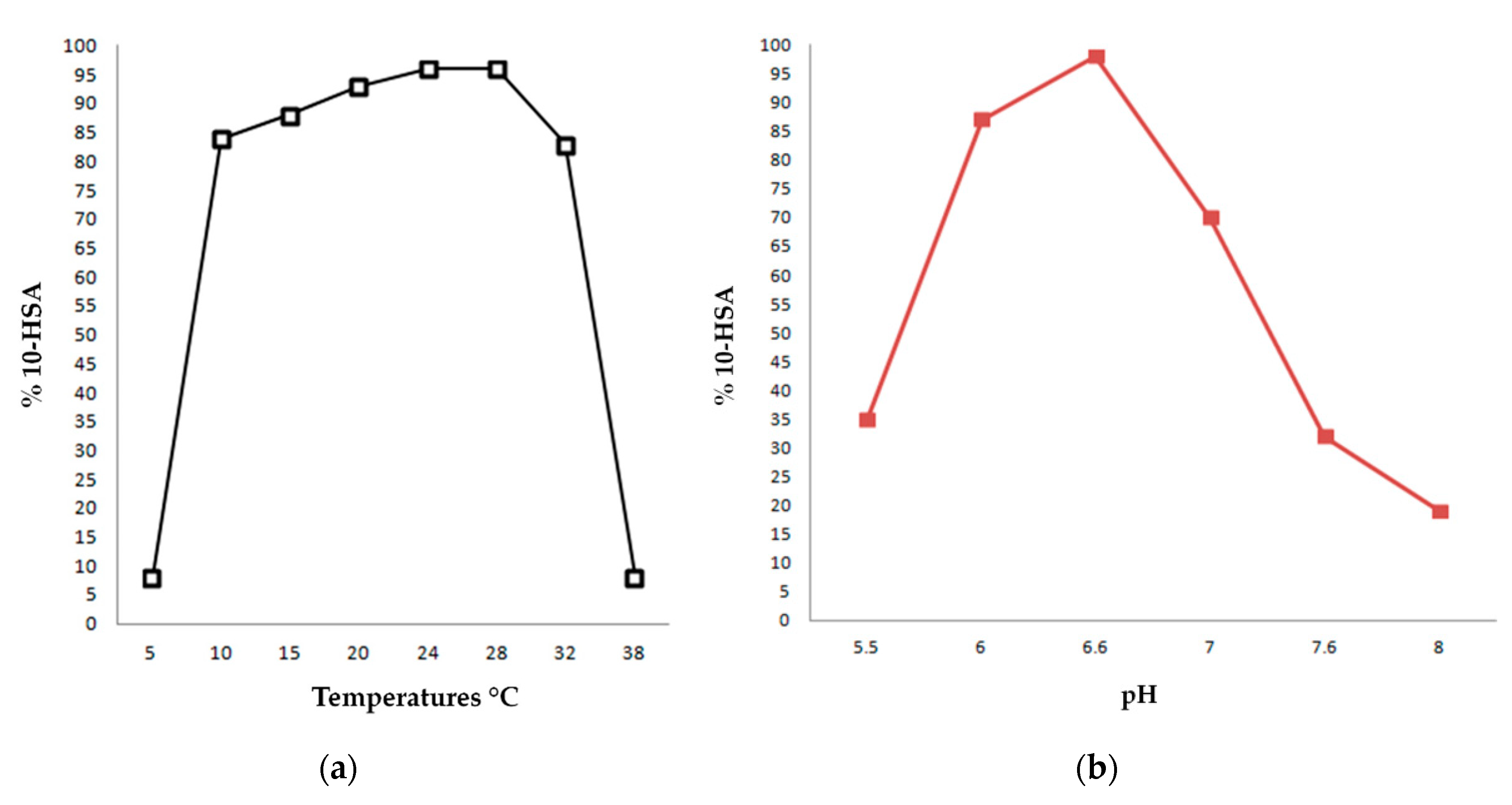
The results of our experiments are collected in
Figure 3a and show that the enzyme maintains high activity performances also at low temperatures, while at 37 °C, its activity decreases markedly.
Overall, we observed that 10-HSA was obtained with yields superior to 90% in a temperature range between 20 °C and 30 °C, with a maximum at 28 °C. This result is apparently in contrast with the fact that Lactobacillus rhamnosus ATCC 53103 is a probiotic microorganism isolated from the human intestine and with an optimum growth temperature of 37 °C. However, in recombinant bacterial cells, high temperatures could promote non-specific intra/inter molecular interactions that could alter the native structure and the functionality of the enzyme.
As far as the optimum pH concerns, we carried out the same experiment described above but incubating all the reactions at a temperature of 28 °C and modifying the composition of the potassium phosphate buffer in each trial in order to set them at desired pH.
Figure 3b reports the results of this study, showing that the enzyme possesses an optimum activity at pH 6.6, while the performances decrease drastically at pHs lower than 6 and higher than 7.5.
In conclusion, the data suggest that the best conditions for the hydration catalyzed by the recombinant oleate hydratase from Lactobacillus rhamnosus ATCC 53103 are in a temperature range between 24 and 28 °C and at pH 6.6.
2.5. Enantioselectivity of OLH Reaction
In order to define the stereoselectivity related to the hydration reaction, we measured the enantiomeric purity of the obtained 10-HSA according to the Rosazza’s procedure [
46].
This analytical method is based on the nuclear magnetic resonance analysis (1H-NMR) of the diastereoisomeric (S)-O-acetylmandelate esters of the corresponding methyl-10-hydroxystearate. The analysis of the 10-HSA samples prepared using the recombinant oleate hydratase from Lactobacillus rhamnosus ATCC 53103 indicated that the hydration reaction of the OA affords the (R)-10-hydroxy-enantiomer with high enantiomeric purity (ee > 96%).
These experiments confirmed our previous results obtained using the wild type of
L. rhamnosus (ATCC 53103) [
6], in which the produced (
R)-10-hydroxystearic acid possessed ee > 95%. Overall, these finding established that the cloned enzyme retains all the hydratase catalytic capability of the parent bacterium.
3. Materials and Methods
3.1. Materials and General Methods
All air and moisture sensitive reactions were carried out using dry solvents and under a static atmosphere of nitrogen. All solvents and reagents, including oleic acid (94%, lot. MKBZ2615V), kanamycin sulphate, Isopropyl-β-D-thiogalactoside (IPTG), Triton™X-100, Flavin adenine dinucleotide disodium salt hydrate (FAD, purity: ≥95% HPLC) were of commercial quality and were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Riboflavin was purchased from Health Leads UK Ltd., (Horeb, UK).
(
S)-
O-acetyl mandelic acid was prepared starting from (
S)-mandelic acid and using acetic anhydride, pyridine and cat. dimethylaminopyridine (DMAP), as described previously [
47].
A reference standard sample of (
R)-10-hydroxystearic acid (ee > 95%) was prepared by
Lactobacillus rhamnosus mediated hydration of oleic acid [
6].
A reference standard sample of 10-(
R)-hydroxystearic acid, showing 21% ee, was prepared by baker’s yeast-mediated hydration of oleic acid [
5].
3.2. Bacterial Strains and Gene Isolation
3.2.1. Microorganisms and Growth Media
Lactobacillus rhamnosus (ATCC 53103, trade name Kaleidon 60), was purchased from Malesci Spa (Bagno a Ripoli, Italy).
This particular strain was grown in MRS medium under anaerobic conditions, 37 °C and 110 rpm for 2 days.
MRS medium composition: casein peptone (10 g/L), meat extract (10 g/L), yeast extract (5 g/L), glucose (20 g/L), Tween 80 (1 mL/L), K2HPO4 (2 g/L), NaOAc (5 g/L), ammonium citrate dibasic (2 g/L), MgSO4·7H2O (0.2 g/L) MnSO4·H2O (0.05 g/L), L-cysteine 0.1% (w/w), sodium thioglycolate (2 g/L).
HI-Control E. coli BL21(DE3) chemically competent cells were cultivated following the instruction of the supplier Lucigen (Middleton, WI, USA).
The growth and expression media used in this work are LB and ZYM-5052 implemented with 20 mg/L of riboflavin (FAD precursor).
ZYM-5052 medium composition: ZYM-0, 50X 5052 solution (20 mL/L) and 1000X trace solution (200 µL).
The bulk composition of ZYM medium (ZYM-0) is casein peptone (10 g/L), yeast extract (5 g/L), Na2HPO4 (3.6 g/L), KH2PO4 (3.5 g/L), NH4Cl (2.7g/L), Na2SO4 (0.7 g/L) and MgSO4 (0.25 g/L). pH was adjusted to 7.0. ZYM-0 is then autoclaved for 15 min at 121 °C.
Trace solution (1000X) composition: FeCl3 (50 mM), CaCl2 (20 mM), MnCl2 (10 mM), ZnSO4 (10 mM), CoCl2 (2 mM), CuCl2 (2 mM), NiCl2 (2 mM), Na2MoO4 (2 mM), Na2SeO3 (2 mM), H3BO3 (2 mM).
50X 5052 solution composition: glycerol (250 g/L), glucose (25 g/L), α-lactose (100 g/L). Once prepared, the solution was filtered on Sartorius™ Minisart™ Plus Syringe Filters 0.2 µm pore size and stored at 4 °C.
LB composition: NaCl (5 g/L), tryptone (10 g/L), yeast extract (5 g/L). pH was adjusted to 7.0.
3.2.2. Oleate Hydratase from L. rhamnosus ATCC 53103: Gene Isolation and Cloning
Chromosomal DNA of
L. rhamnosus ATCC 53103 was obtained by growing the strain in MRS medium at 37 °C with 110 rpm for 36 h under anaerobic conditions. The DNA extraction was performed by using the E.Z.N.A. Bacterial DNA kit, Omega/VWR, D3350-02. The PCR amplification of the gene coding for oleate hydratase (GenBank: WP_005714981) was performed using primers designed for the subsequent cloning in the pETite C-His Kan vector in frame with C-term His Tag sequence (
Table 2).
PCR amplifications were carried out in 50 µL reaction mixtures containing 1 µg genomic DNA, primers (1 µM each), dNTPs (0.2 mM each), 2 U of Pfu polymerase and 5 µl of buffer containing MgCl2. All PCR reagents were from Genespin (Milan, Italy).
PCR conditions were as follows: 95 °C for 3 min, followed by 35 cycles at 95 °C for 30″, primers Tm-5 °C (55 °C) for 30″, 72 °C for 1′, and then 72 °C for 10′.
Purification of PCR products from agarose gel (0.7% (
w/
v)) was carried out by using the Wizard
® SV Gel and PCR Clean-Up System (Promega, WI, USA). The amplified fragments were subsequently cloned in the pETite C-His Kan plasmid (
Figure 4) using the Expresso T7 Cloning and Expression kit from Lucigen (Middleton, WI, USA) and following the manufacturer′s instructions.
The resulting plasmids were purified by using the Plasmid Mini kit II (Omega, Biel/Bienne, Switzerland; VWR, Radnor, PA, USA) and sequenced on both strands by Biofab Research (Rome, Italy) using primers pETite T7 Forward and pETite reverse (Lucigen Expresso T7 Cloning and Expression kit).
Recombinant pETite plasmids were then used for the transformation of chemically competent E. coli BL21(DE3) cells (Lucigen, Middleton, WI, USA).
3.3. Protein Expression and Purification
Recombinant E. coli BL21(DE3) cells, prepared as described above, were inoculated in a 1 L Erlenmeyer flask containing LB medium (100 mL) supplemented with kanamycin (30 µg/mL) and grown at 37 °C with 220 rpm overnight.
At this point, we tested the protein expression using ZYM-5052 and a fresh LB medium, both containing 30 µg/mL of kanamycin. The temperature tested was 20 °C.
Starting from the overnight culture described above, we inoculated 100 mL of medium in 1 L Erlenmeyer flasks in order to have an OD600 cell density of 0.2. The protein expression procedure is different depending on the use of LB medium or ZYM-5052 medium.
We kept ZYM-5052 cultures at 20 °C with 180 rpm for 24 h, while the cultures inoculated in LB medium were incubated at 37 °C with 220 rpm till the OD600 cell density reached 0.6. For the latter experiments, gene expression was then induced by the addition of IPTG (final concentration 0.1; 0.5; 1 mM), and the cultures were transferred to 20 °C and left overnight under shaking (180 rpm).
The cells were then recovered by centrifugation (8000 rpm, 15 min, 4 °C) and resuspended in 20 mM potassium phosphate buffer (10% w/v), pH 7.0, containing 500 mM NaCl and 20 mM imidazole. At this point, cells were disrupted by ultrasonication (6 cycles, 20″ON/40″ OFF pulses in ice).
Clear lysates were obtained by centrifugation (11,000 rpm, 30 min) and the presence of soluble proteins was assessed by SDS-PAGE (12% T, 2.6% C).
Protein purification was performed by incubation of clear cell lysates with Ni Sepharose 6 Fast Flow agarose resin (Ni-NTA) (GE Healthcare, Milan, Italy) under mild shaking for 90 min at 4 °C. The resin was then loaded onto a glass column (10 × 110 mm) and washed with 20 mL of wash buffer (20 mM imidazole, 500 mM NaCl, 20 mM potassium phosphate buffer). Elution of His-tagged proteins was achieved by using a 3-step gradient (10 mL washing buffer containing 100, 200 and 300 mM imidazole, respectively). Purified protein samples were dialyzed for 16 h at 4 °C against 50 mM potassium phosphate buffer, pH 6.6 (2 L), then stored at −80 °C.
Protein concentration was determined by using the Bio-Rad Protein Assay according to the method of Bradford, and SDS-PAGE analyses (12% T, 2.6% C) were performed to assess protein purity. Molecular weight protein standards were from Bio-Rad (Karlsruhe, Germany), and gels were stained with Coomassie Brilliant Blue for protein detection.
In general, 2 g of wet cells are required to obtain 30 mg of pure oleate hydratase.
3.4. Determination of Biotransformation Optimum Conditions
These experiments were performed using whole cells of recombinant
E. coli BL21(DE3), obtained as described below (
Section 3.5.1). A cell’s suspension (5%
w/
v) in 50 mM potassium phosphate buffer, at the required pH (from 5.5 to 8.0), was treated with oleic acid, glycerol and ethanol in order to obtain a final concentration of these compounds corresponding to 3 g/L, 10%
v/
v and 2%
v/
v, respectively. Each trial was shaken at 150 rpm for 24 h. After this time, the reaction was acidified to pH 3 by addition of diluted HCl and then filtered on a celite pad. The aqueous phases were then extracted three times with ethyl acetate, the combined organic layers were washed with brine and dried on Na
2SO
4 and the solvent was removed under reduced pressure. The 10-HSA/OA ratio was measured by GC-MS analysis, as described below (
Section 3.6.2).
In order to assess the optimal temperature for the OLH-catalyzed production of 10-HSA from OA, we performed eight hydration trials at different temperatures, using the general experimental conditions described above and setting the pH at 6.6 value. The experiments performed at 5, 10, 15, 20, 24, 28, 32 and 37 °C afforded 10-HSA in 8%, 84%, 88%, 93%, 96%, 96%, 83% and 8% yields, respectively. These results are collected in
Figure 3a.
In order to assess the optimal pH for the OLH-catalyzed production of 10-HSA from OA, we performed six hydration trials at different pH levels, using the general experimental conditions described above and setting the temperature at 28 °C. The experiments performed at pH 5.5, 6.0, 6.6, 7.0, 7.6 and 8.0 afforded 10-HSA in 35%, 87%, 98%, 70%, 32% and 19% yields, respectively. These results are collected in
Figure 3b.
3.5. Biotransformation Experiments
3.5.1. Whole-Cell Catalyst
Recombinant E. coli BL21(DE3) cells prepared in ZYM-5052 medium at 20 °C with 180 rpm for 24 h were recovered by centrifugation (8000 rpm, 15 min, 4 °C). The pellet was stored at −20 °C for at least one day.
A suspension of the latter cells (5%
w/
v) in 50 mM potassium phosphate buffer (pH 6.6) was treated with oleic acid (3 g/L) dissolved in EtOH (final EtOH concentration: 2%
v/
v). Different samples of the obtained mixture were treated with other co-solvent or non-ionic surfactant in order to improve the solubility of the enzyme and of the substrate. More specifically, we tested different concentrations of the surfactant Triton™ X-100 and of the co-solvent glycerol. Each trial was kept at 28 °C, shaking at 150 rpm for 24 h. After this time, the reaction was acidified to pH 3 by addition of diluted HCl and then filtered on a celite pad. The aqueous phases were then extracted three times with ethyl acetate, the combined organic layers were washed with brine and dried on Na
2SO
4 and the solvent was removed under reduced pressure. The crude biotransformation mixtures were derivatized, and the 10-HSA/OA ratio was measured by GC-MS analysis, as described below (
Section 3.6.2). As a result of these experiments, we assessed that the higher hydratase activity is obtained with a concentration of Triton™ X-100 of 0.5%
v/
v or, using a glycerol concentration, of 10% v/v. These two trials afforded 10-HSA in 60% and 98% yield, respectively (
Table 1).
Using smaller or bigger amounts of these two compounds, we observed a significant reduction of the enzyme activity.
3.5.2. Clear Cell Lysate Catalyst
Recombinant E. coli BL21(DE3) prepared in ZYM-5052 at 20 °C with 180 rpm for 24 h was recovered by centrifugation (8000 rpm, 15 min, 4 °C). The pellet was stored at −20 °C for at least one day.
At this point, it was resuspended (5% w/v) in 50 mM potassium phosphate buffer, pH 6.6, and sonicated as described above. The soluble protein fraction was separated from the cell debris by centrifugation (14,000 rpm, 30 min), and clear lysates were used as such for the biotransformation experiments, adding to the solution oleic acid (3 g/L), EtOH (2% v/v) and glycerol (10% v/v). The reaction mixture was kept at 28 °C with 150 rpm for 24 h.
After this time, the reaction was acidified to pH 3 by the addition of diluted HCl. The aqueous phases were then extracted three times with ethyl acetate, the combined organic layers were washed with brine and dried on Na
2SO
4 and the solvent was removed under reduced pressure. The crude biotransformation mixtures were derivatized, and the 10-HSA/OA ratio was measured by GC-MS analysis, as described below (
Section 3.6.2). The above-described trial afforded 10-HSA in 15% yield (
Table 1).
3.5.3. Purified OLH
Purified OLH samples (1.5 mg in 3 mL of 50 mM potassium phosphate buffer, pH 6.6, see
Section 3.3 for details) were treated with oleic acid, glycerol and ethanol in order to obtain a final concentration of these compounds corresponding to 3 g/L, 10%
v/
v and 2%
v/
v, respectively.
In another experiment, the same reaction mixture was treated with FAD (1.1 molar equivalents respect the enzyme) in order to evaluate its role in the activity of the enzyme.
The two reaction mixtures were kept at 28 °C with 150 rpm for 24 h.
After this time, the reactions were acidified to pH 3 by addition of diluted HCl. The aqueous phases were then extracted three times with ethyl acetate, the combined organic layers were washed with brine and dried on Na
2SO
4 and the solvent was removed under reduced pressure. The crude biotransformation mixtures were derivatized, and the 10-HSA/OA ratio was measured by GC-MS analysis, as described below (
Section 3.6.2). The above-described trials afforded 10-HSA in 0% and 10% yields (
Table 1).
3.6. Analytical Methods and Characterization of the Products Deriving from the Biotransformation Experiments
3.6.1. Instruments and Analytic Condition
Nuclear Magnetic Resonance spectroscopy (NMR): 1H- and 13C-NMR Spectra and DEPT experiments: CDCl3 solutions at RT using a Bruker-AC-400 spectrometer (Billerica, MA, USA) at 400, 100, and 100 MHz, respectively; 13C spectra are proton decoupled; chemical shifts in ppm relative to internal SiMe4 (=0 ppm).
TLC: Merck silica gel 60 F254 plates (Merck Millipore, Milan, Italy).
Column chromatography: silica gel.
GC-MS analyses: A HP-6890 gas chromatograph equipped with a 5973 mass detector and using a HP-5MS column (30 m × 0.25 mm, 0.25 μm film thickness; Hewlett Packard, Palo Alto, CA, USA) was used with the following temp. program: 120° (3 min)—12°/min—195° (10 min)—12°/min—300° (10 min); carrier gas: He; constant flow 1 mL/min; split ratio: 1/30; tR given in min.
3.6.2. GC-MS Analyses
The biotransformations of oleic acid to give 10-hydroxystearic acid were monitored by means of GC-MS analysis. To this end, the biotransformation mixture was acidified at pH 4 and filtered on celite. The aqueous phase was then extracted three times with ethyl acetate, and the combined organic layer was washed with brine and dried on Na2SO4. The solvent was then removed under reduced pressure, and the residue was treated at 0 °C with an excess of an ethereal solution of freshly prepared diazomethane. As soon as the evolution of nitrogen ceased, the solvent was eliminated and the residue was treated at RT with a 1:1 mixture of pyridine/acetic anhydride (4 mL for about 100 mg of residue) and DMAP (10 mg). After five hours, the excess of reagents was removed in vacuo and the residue was analyzed by GC-MS.
Oleic acid methyl ester: tR 18.95
GC-MS (EI): m/z (%) = 296 [M+] (7), 264 (49), 235 (6), 222 (30), 180 (19), 166 (10), 152 (12), 137 (17), 123 (26), 110 (32), 97 (62), 83 (68), 69 (79), 55 (100).
Methyl 10-acetoxystearate: tR 24.47
GC-MS (EI): m/z (%) = 313 [M+-MeCO] (6), 296 [M+-AcOH] (3), 281 (17), 264 (31), 243 (11), 222 (9), 201 (100), 169 (64), 157 (16), 125 (21), 97 (18), 83 (19), 69 (21), 55 (27).
3.6.3. Determination of the Absolute Configuration and of the Optical Purity of 10-Hydroxystearic Acid
The enantiomeric composition of the isolated 10-hydroxystearic acid sample, obtained from the biotransformation experiments, was determined by
1H-NMR analysis according to the Rosazza procedure [
46]. Hence, each one of the hydroxy acid samples (100 mg, 0.33 mmol) was treated with an excess of an ethereal solution of freshly prepared diazomethane. As soon as the evolution of nitrogen ceased, the solvent was eliminated and the resulting methyl ester was dissolved in dry CH
2Cl
2 (5 mL) treated with (
S)-
O-acetylmandelic acid (130 mg, 0.67 mmol), DCC (140 mg, 0.68 mmol) and DMAP (10 mg), stirring at RT for 6 h. The reaction was then quenched by the addition of water and diethyl ether (60 mL). The formed dicyclohexylurea was removed by filtration on celite and the organic phase was washed with aq. NaHCO
3, brine and dried on Na
2SO
4. The solvent was then removed under reduced pressure and the residue was roughly purified by chromatography, collecting every fraction containing the fatty acid mandelates. The
1H-NMR analysis of the obtained (
S)-
O-acetylmandelates allowed the determination of the absolute configuration of the starting hydroxy acid as well as the measurement of their enantiomeric purity.
The above described analytical procedure was applied to a sample of 10-HSA obtained by hydration of OA, using the recombinant OLH from
L. rhamnosus ATCC 53103 as a catalyst. We assessed an enantiomeric excess higher than 96% (
Supplementary Materials).
4. Conclusions
This work established the experimental procedures for the production of recombinant OLH from Lactobacillus rhamnosus by overexpression in Escherichia coli BL21(DE3). The study demonstrated that the obtained enzyme catalyzes the hydration of OA with a very high stereoselectivity, affording enantiopure (R)-10-HSA. We established that the optimal biotransformation conditions were pH 6.6 in phosphate buffer, at a temperature of 28 °C.
From an applicative point of view, the best biotransformation conditions were shown when using recombinant OLH as a whole-cell catalyst. We devised a reliable procedure for the hydration of OA by means of whole cells of recombinant E. coli as catalysts and using glycerol and ethanol as co-solvents. Under these conditions, high substrate concentrations (up to 50 g/L of OA) could be applied, thus resulting in a good compromise between catalyst performances and production costs. In fact, the enzyme purification leads both to the loss of the catalyst itself and to the reduction of its activity, which must be restored by the addition of FAD.
Finally, it is worthy of note that besides hydrating oleic acid, Lactobacillus rhamnosus is able to transform linoleic and linolenic acid into their corresponding 10-hydroxy-derivatives. Therefore, the evaluation of the substrate specificity of the recombinant OLH obtained by this work could give new insights into the biotechnological potential of this enzyme. Further studies on this topic are now in progress and will be reported on in due course.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}