Evolution of Cooperation for Multiple Mutant Configurations on All Regular Graphs with N ≤ 14 Players


Abstract
1. Introduction
2. Evolution of Cooperation
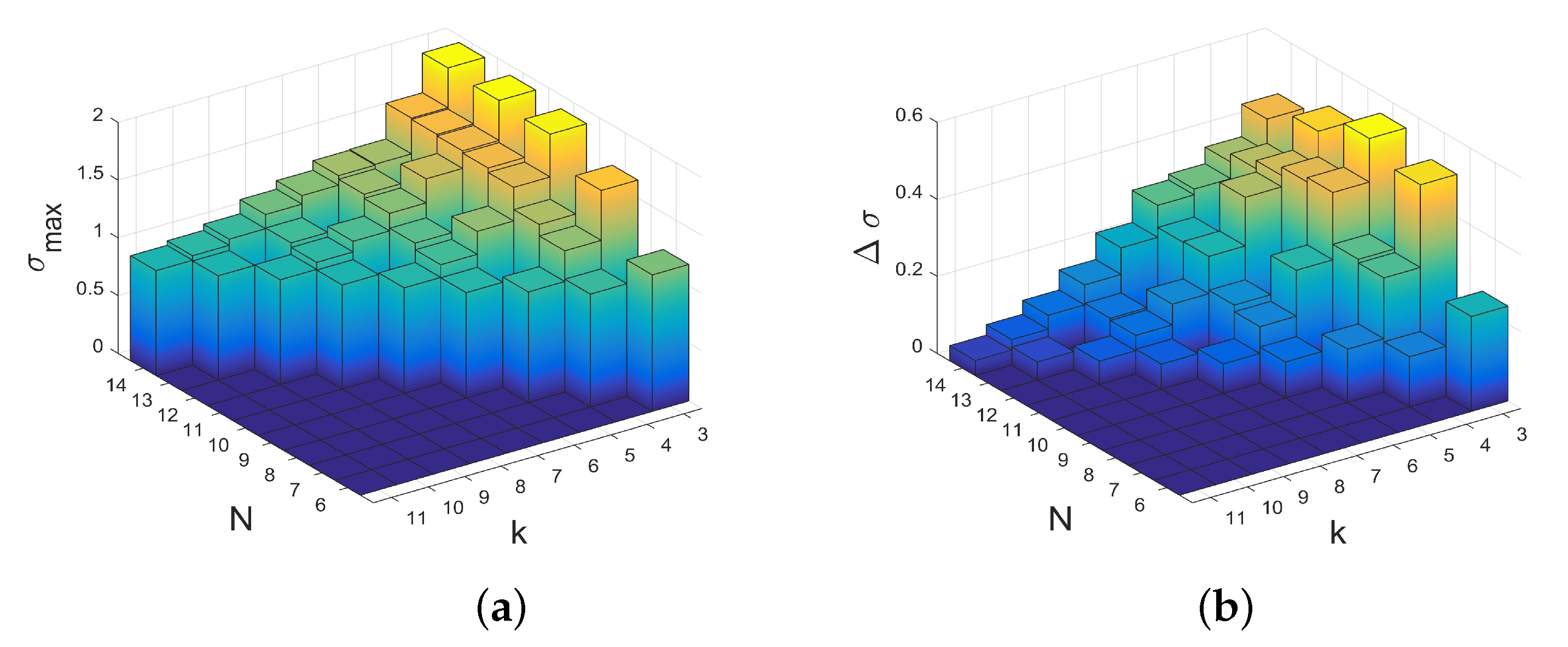
2.1. Upper and Lower Bounds on the Structure Coefficients
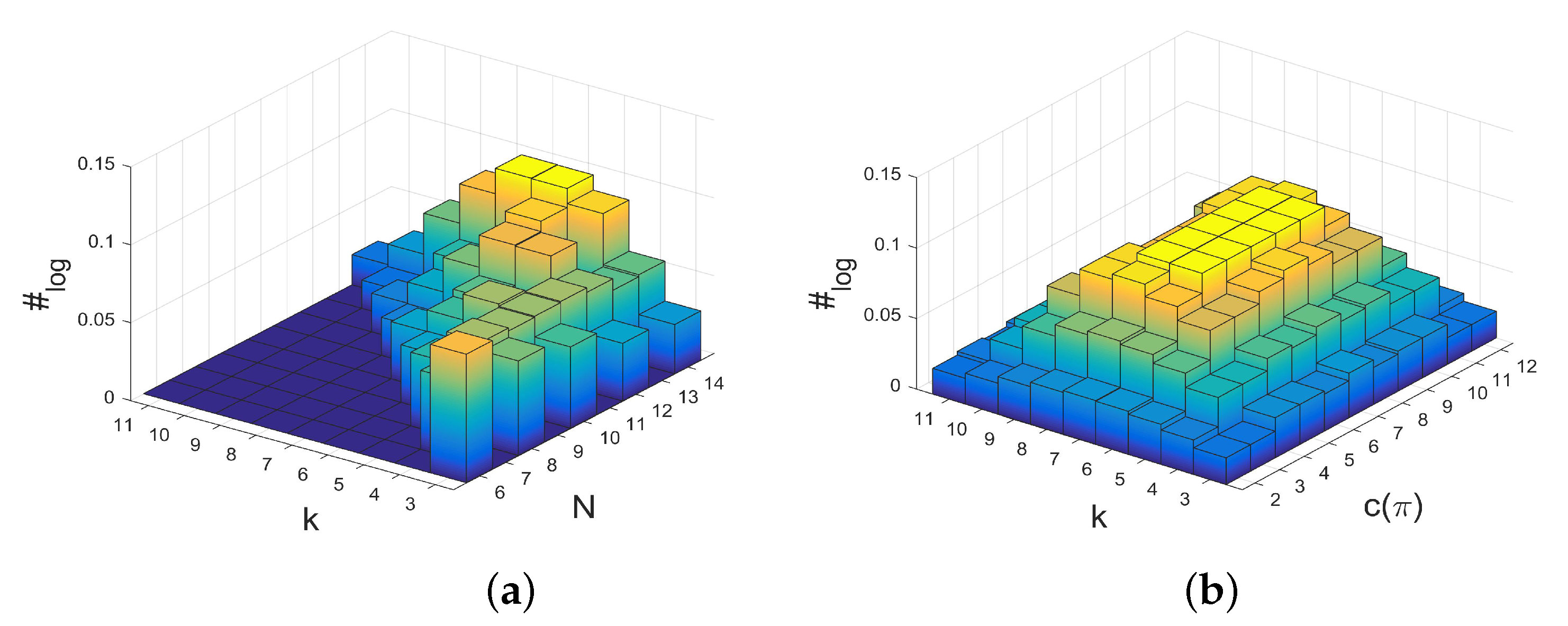
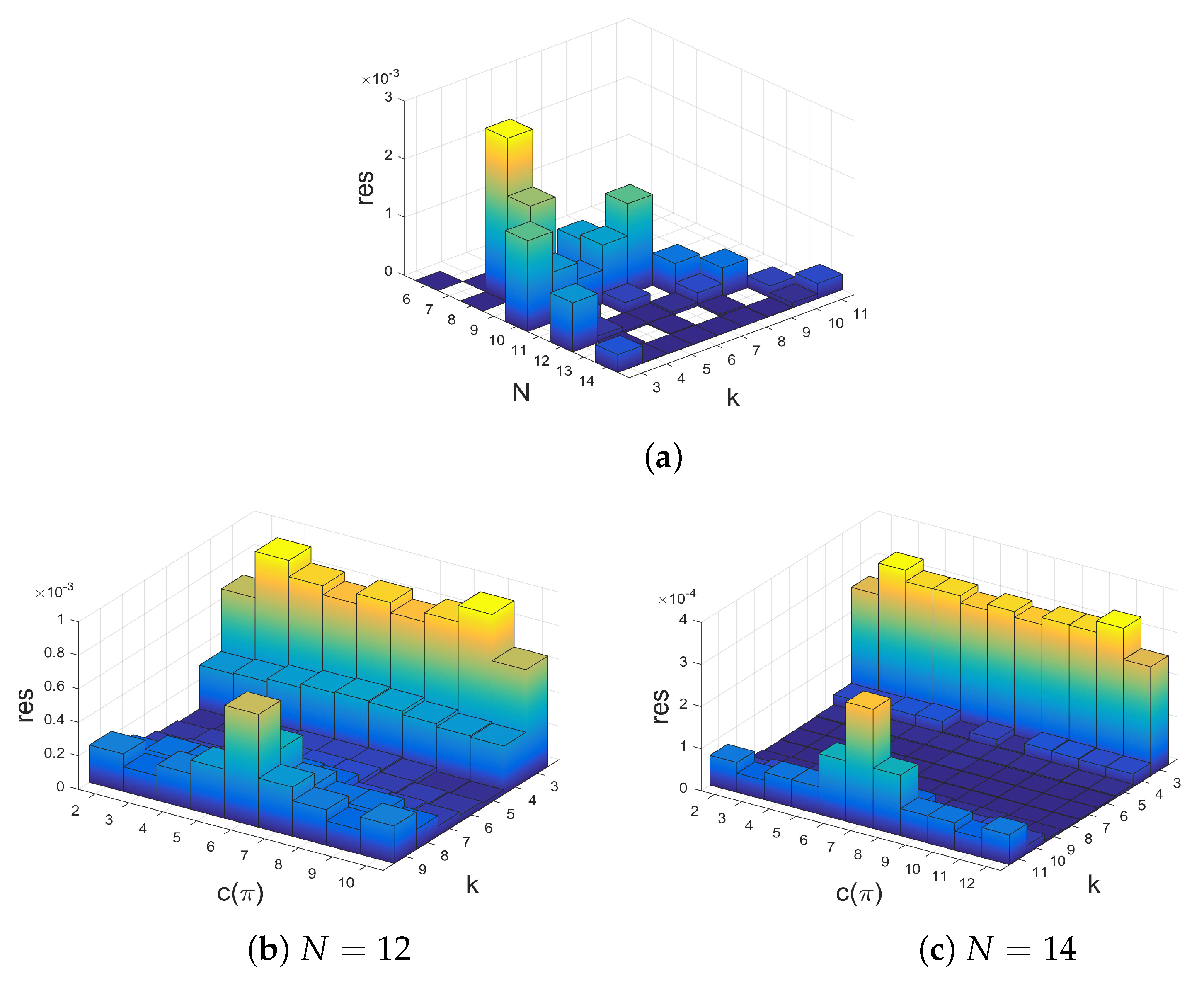
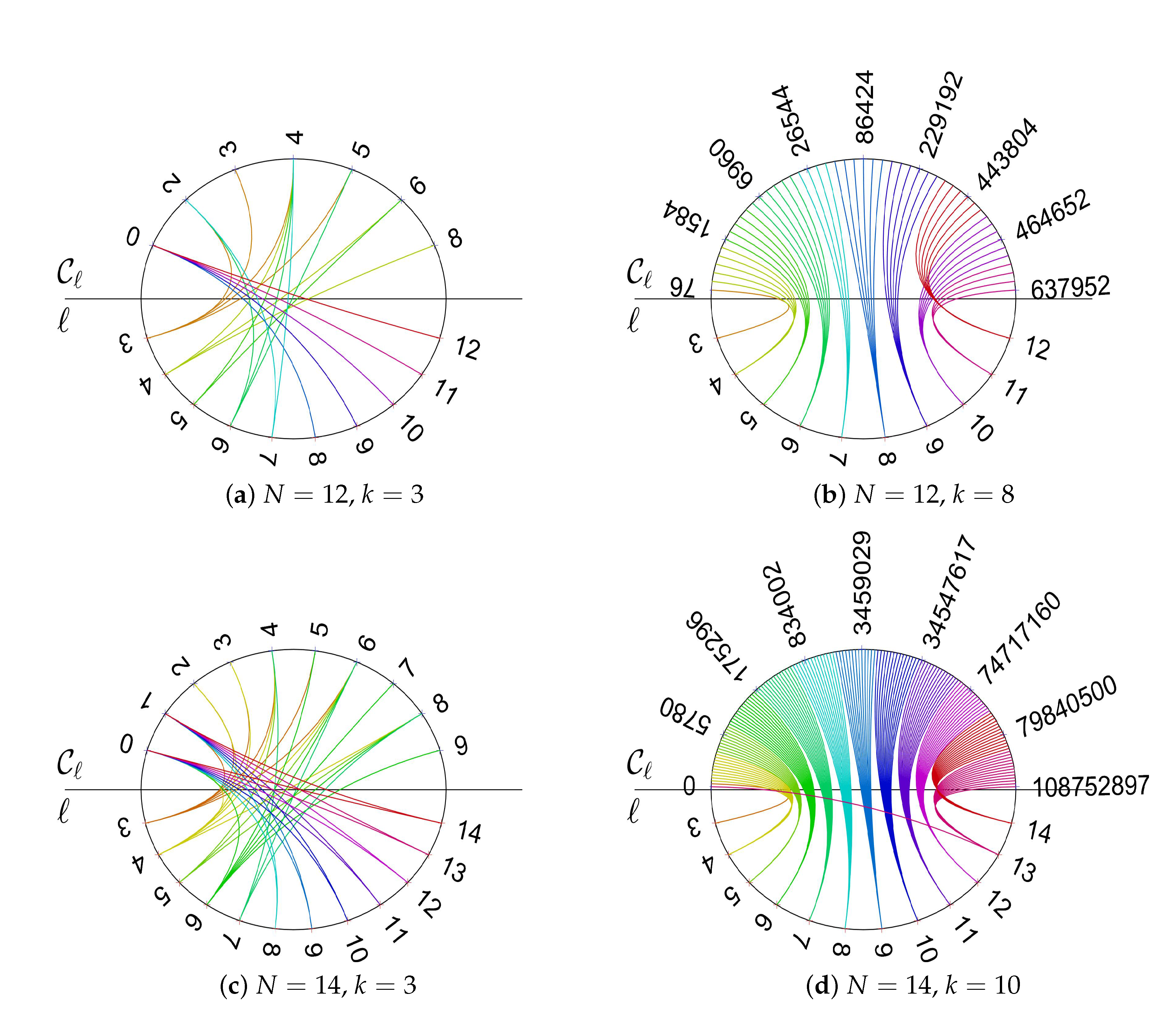
2.2. Relationships between Structure Coefficients and Graph Cycles
3. Discussion and Conclusions
- a.
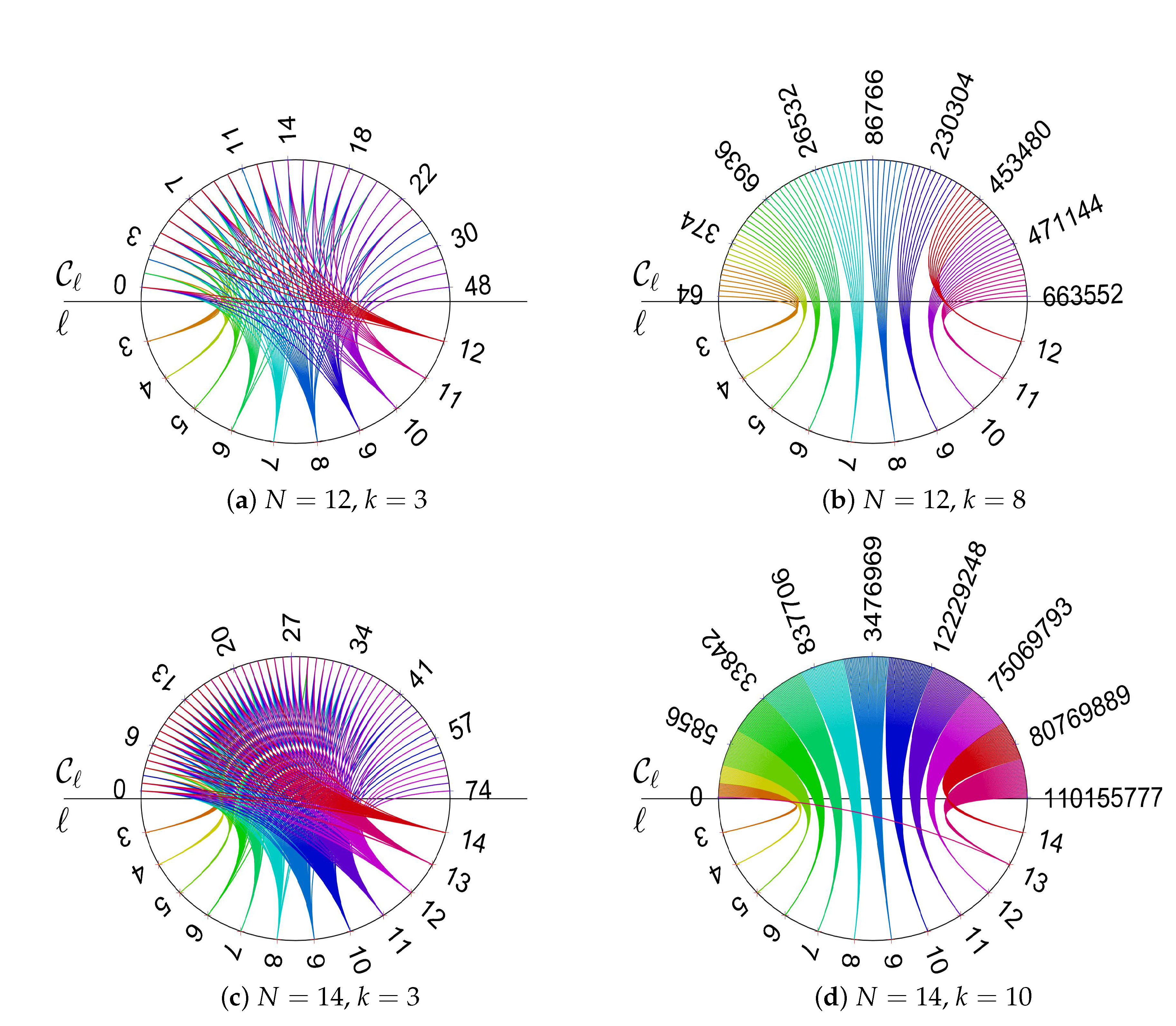
- There is an approximately linear relationship between maximal structure coefficients and the count of cycles of the interaction graph with a certain length. Moreover, the number of -graphs grows much slower for a rising number of players than the number of k-regular graphs on N vertices. Thus, graphs with maximal structure coefficients get rare for the number of players N getting large.
- b.
- The values of the structure coefficients are larger for a small number of coplayers (that is for graphs with a small degree) than for larger numbers of coplayers. It is maximal for , which is cubic graphs. This is also the case for the largest difference between maximal and minimal structure coefficients. Thus, for regular evolutionary graphs describing the interactions between players, the results for players suggest that a smaller number of coplayers is particularly prone to promote cooperation if a favorable graph is selected. The selection of graphs does matter less for a larger number of coplayers. The -graphs with small numbers of coplayers k not only have the largest maximal structure coefficients, they are also characterized by the absence of cycles with a length above a certain limit, see examples in the collection of -graphs in Appendix C.
- c.
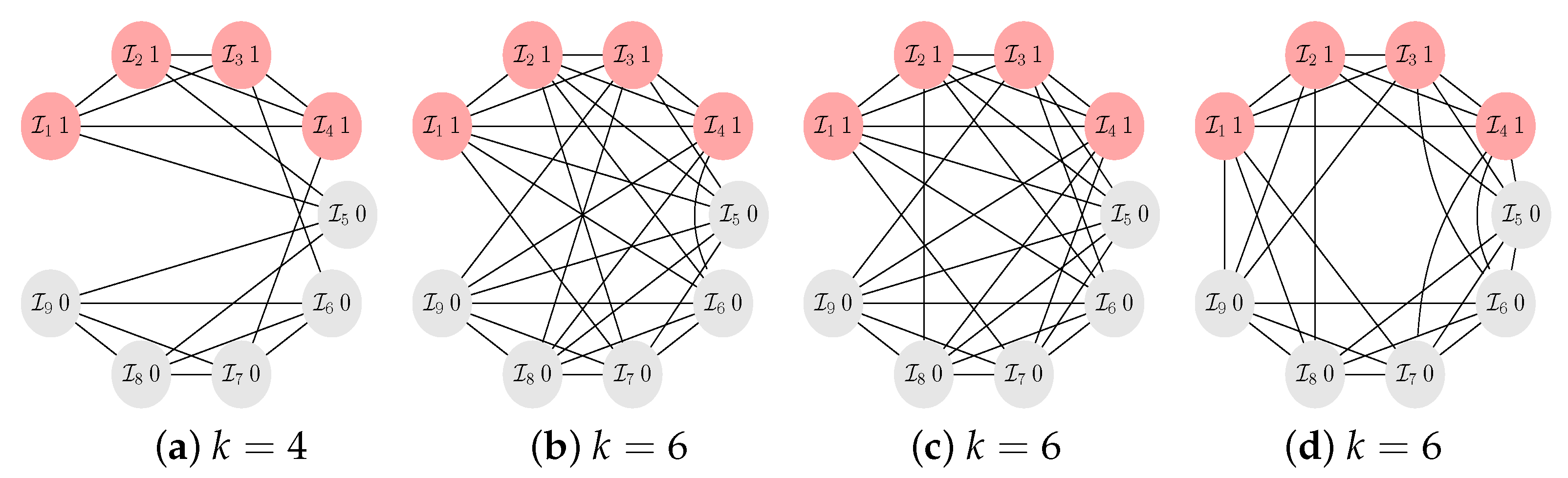
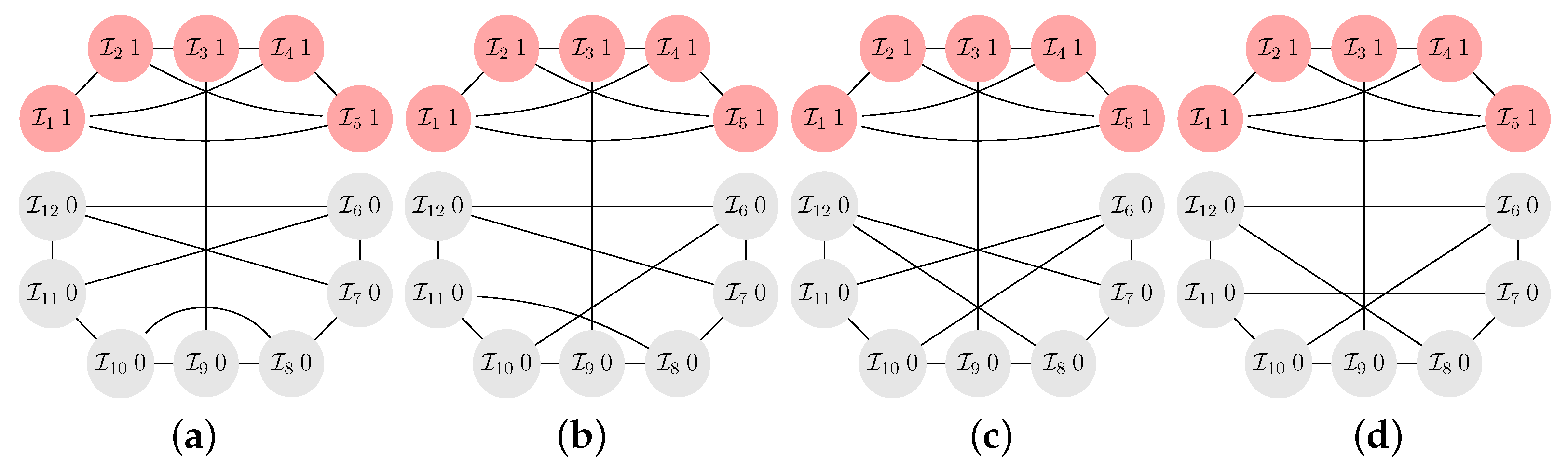
- There are not only no long cycles in -graphs with small k. The graphs are also structured into blocks that are connected by cut vertices and/or hinge vertices. A cut vertex is a vertex whose removal disconnects the graph, while a hinge vertex is a vertex whose removal makes the distance longer between at least two other vertices of the graphs [25,26]. For instance, for and , the vertices occupied by the players and , see Figure A5, are cut vertices, while for and , see Figure A4b, the vertices occupied by the players and are hinge vertices as their removal would make the distance between and longer. The blocks are occupied by clusters of cooperators. The clusters can be seen to serve as a mutant family that invades the remaining graph. As vertices with players of opposing strategies are connected by cut and/or hinge vertices there is only a small number of migration paths (or even just a single path) for the cooperators and/or defectors. A similar observation has been reported for evolutionary games on lattice grids [18,20], see also the discussion in Section 2.2. To summarize: the results suggest that -graphs for small numbers of coplayers have some distinct graph–theoretical properties. Searching for these properties in a given graph may inform the design of interactions graphs that are either particularly prone to cooperation or particularly opposed to it.
- d.
- The property of missing long cycles is also a possible explanation as to why regular graphs with a small degree differ substantially from graphs with a larger degree in terms of promoting cooperation in evolutionary games. A larger degree makes it impossible to have blocks that are connected by only a few edges. As the number of edges increases linearly with the degree by and each vertex has the same number of edges, there is an ample supply on connections. These results imply that the connectivity properties of the interaction graph play an important role in the emergence of cooperation. It may be interesting to see if these connectivity issues may possibly also show in algebraic graph measures, for instance, algebraic connectivity expressed by the Fiedler vector.
- e.
- The paper discusses the evolution of cooperation for all regular graphs with players with death–birth (DB) updating and weak selection. Thus, it may be interesting to hypothesize about the relevance of the results for non-regular graphs, stronger levels of selection and other types of strategy updating. Recently, the relationships between the graph structure and fixation properties have been clarified substantially for a single mutant [6,7,8,9]. These results suggest that regular graphs have some similarities to Erdös–Rényi graphs, whereas other types of graphs, for instance, cycles, trees, stars or comet-kites are much more different. Thus, it might be possible that the results given in this paper for regular graphs may similarly apply to Erdös–Rényi graphs. Particularly interesting in this context may be the relationships between the connectivity of the graphs and the promotion of cooperation. Furthermore, it has been shown that extrapolating results from weak to intermediate and strong selection is not always possible and depends highly on game characteristics, population size and spatial heterogeneity of the network [5,27,28]. However, a comparison between fixation probabilities is rather robust for varying intensity of selection and a single cooperator [27]. Thus, the results obtained using structure coefficients may still be valid beyond weak selection. Finally, for weak selection, many fitness-based updating schemes and pairwise comparison with a Fermi function have similar fixation properties if the fitness can be approximated as a positive constant [29,30]. Thus, the results obtained for DB given in this paper might also have relevance to pairwise comparisons. On the other hand, it has also been shown that for an increasing level of selection intensity fitness-based models and pairwise comparison models of evolutionary games are typically different [31]. These brief comments about the relevance of the results for non-regular graphs, stronger levels of selection and other types of strategy updating must be treated with due caution as they are informed by results for a single mutant. Thus, further work is needed to clarify these relationships and see if they are also valid for multiple mutants (or any arrangement of cooperators and defectors).
- f.
- The results are given in this paper show a clear dependency between the long-term prevalence of cooperation in evolutionary games on regular graphs and some of their graph–theoretical properties, which generally confirm previous findings on clusters of cooperators in games on lattice grids [18,19,20,21], on pairs of mutants on a circle graph () [32], and on short cooperator path lengths on some selected regular graphs with and , among them the Frucht, the Tietze and the Franklin graph [14]. However, apart from statements about the prevalence of cooperation, there are also other quantifiers of evolutionary dynamics that are highly relevant. In other words, some of the difficulty in the given approach for evaluating the emergence of cooperation in evolutionary games on graphs arises from structure coefficients merely treating a comparison of fixation probabilities. The condition indicates that the fixation probability of cooperation is higher than the fixation probability of defection. This, however, does not entail the values of these probabilities. However, structure coefficients can be calculated with polynomial time complexity [12], while computing fixation probabilities is generally intractable due to an exponential time complexity [33,34,35]. In other words, by using the approach involving structure coefficients, we exchange computational tractability for obtaining just a comparison of fixation probabilities instead of their exact values. Moreover, the difference in the descriptive power of the structure coefficients as compared to the fixation probabilities is salient in another way. Most likely, there is a rather complex relationship between structure coefficients and fixation probability, which is illustrated by the example of a single cooperator for which the structure coefficient does precisely not imply unique values of the fixation probability of cooperation. For a single cooperator, we get a single value of the structure coefficient, but fixation probabilities vary over initial configurations as shown for the Frucht and for the Tietze graph [36].
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BD | Birth Death updating |
| DB | Death Birth updating |
Appendix A. Configurations, Regular Graphs, and Structure Coefficients
Appendix B. Isomorphic Graphs, Isomorphic Configurations, and Cycle Counts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
|---|---|---|---|---|---|---|---|---|---|
| 3 | 2 | 0 | 5 | 0 | 19 | 0 | 85 | 0 | 509 |
| 4 | 1 | 2 | 6 | 16 | 59 | 265 | 1.544 | 10.778 | 88.168 |
| 5 | 1 | 0 | 3 | 0 | 60 | 0 | 7.848 | 0 | 3.459.383 |
| 6 | 0 | 1 | 1 | 4 | 21 | 266 | 7.849 | 367.860 | 21.609.300 |
| 7 | 0 | 0 | 1 | 0 | 5 | 0 | 1.547 | 0 | 21.609.301 |
| 8 | 0 | 0 | 0 | 1 | 1 | 6 | 94 | 10.786 | 3.459.386 |
| 9 | 0 | 0 | 0 | 0 | 1 | 0 | 9 | 0 | 88.193 |
| 10 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 10 | 540 |
| 11 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 13 |
| 12 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |

Appendix C. Collection of σmax-Graphs with N ≤ 14






References
- Broom, M.; Rychtar, J. Game–Theoretical Models in Biology; Chapman and Hall/CRC: Boca Raton, FL, USA, 2013. [Google Scholar]
- Iyer, S.; Killingback, T. Evolution of cooperation in social dilemmas on complex networks. PLoS Comput. Biol. 2016, 12, e1004779. [Google Scholar] [CrossRef]
- Newton, J. Evolutionary game theory: A renaissance. Games 2018, 9, 31. [Google Scholar] [CrossRef]
- Nowak, M.A. Evolutionary Dynamics: Exploring the Equations of Life; Harvard University Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Wu, B.; Traulsen, A.; Gokhale, C.S. Dynamic properties of evolutionary multi–player games in finite populations. Games 2013, 4, 182–199. [Google Scholar] [CrossRef]
- Allen, B.; Lippner, G.; Chen, Y.T.; Fotouhi, B.; Momeni, N.; Yau, S.T.; Nowak, M.A. Evolutionary dynamics on any population structure. Nature 2017, 544, 227–230. [Google Scholar] [CrossRef]
- Möller, M.; Hindersin, L.; Traulsen, A. Exploring and mapping the universe of evolutionary graphs identifies structural properties affecting fixation probability and time. Commun. Biol. 2019, 2, 137. [Google Scholar] [CrossRef]
- Pavlogiannis, A.; Tkadlec, J.; Chatterjee, K.; Nowak, M.A. Construction of arbitrarily strong amplifiers of natural selection using evolutionary graph theory. Commun. Biol. 2018, 1, 71. [Google Scholar] [CrossRef]
- Tkadlec, J.; Pavlogiannis, A.; Chatterjee, K.; Nowak, M.A. Population structure determines the tradeoff between fixation probability and fixation time. Commun. Biol. 2019, 2, 138. [Google Scholar] [CrossRef]
- Chen, Y.T. Sharp benefit–to–cost rules for the evolution of cooperation on regular graphs. Ann. Appl. Probab. 2013, 3, 637–664. [Google Scholar] [CrossRef]
- Richter, H. Dynamic landscape models of coevolutionary games. BioSystems 2017, 153–154, 26–44. [Google Scholar] [CrossRef]
- Chen, Y.T.; McAvoy, A.; Nowak, M.A. Fixation probabilities for any configuration of two strategies on regular graphs. Sci. Rep. 2016, 6, 39181. [Google Scholar] [CrossRef]
- Richter, H. Properties of network structures, structure coefficients, and benefit–to–cost ratios. BioSystems 2019, 180, 88–100. [Google Scholar] [CrossRef]
- Richter, H. Fixation properties of multiple cooperator configurations on regular graphs. Theory Biosci. 2019, 138, 261–275. [Google Scholar] [CrossRef]
- Tarnita, C.E.; Ohtsuki, H.; Antal, T.; Fu, F.; Nowak, M.A. Strategy selection in structured populations. J. Theor. Biol. 2009, 259, 570–581. [Google Scholar] [CrossRef]
- Ohtsuki, H.; Hauert, C.; Lieberman, E.; Nowak, M.A. A simple rule for the evolution of cooperation on graphs and social networks. Nature 2006, 441, 502–505. [Google Scholar] [CrossRef]
- Giscard, P.L.; Kriege, N.; Wilson, R.C. A general purpose algorithm for counting simple cycles and simple paths of any length. Algorithmica 2019, 81, 2716–2737. [Google Scholar] [CrossRef]
- Hauert, C. Fundamental clusters in spatial 2×2 games. Proc. R. Soc. B 2001, 268, 761–769. [Google Scholar] [CrossRef]
- Hauert, C.; Doebeli, M. Spatial structure often inhibits the evolution of cooperation in the snowdrift game. Nature 2004, 428, 643–646. [Google Scholar] [CrossRef]
- Langer, P.; Nowak, M.A.; Hauert, C. Spatial invasion of cooperation. J. Theor. Biol. 2008, 250, 634–641. [Google Scholar] [CrossRef]
- Page, K.M.; Nowak, M.A.; Sigmund, K. The spatial ultimatum game. Proc. R. Soc. B 2000, 267, 2177–2182. [Google Scholar] [CrossRef]
- Krzywinski, M. Schemaball: A new spin on database visualization. SysAdmin Mag. 2004, 13, 23–28. [Google Scholar]
- Wang, Z.; Kokubo, S.; Jusup, M.; Tanimoto, J. Universal scaling for the dilemma strength in evolutionary games. Phys. Life Rev. 2015, 14, 1–30. [Google Scholar] [CrossRef]
- Richter, H. Relationships between dilemma strength and fixation properties in coevolutionary games. In Advances in Natural Computation, Fuzzy Systems and Knowledge Discovery. ICNC-FSKD 2019; Liu, Y., Wang, L., Zhao, L., Yu, Z., Eds.; Advances in Intelligent Systems and Computing; Springer Nature: Cham, Switzerland, 2020; Volume 1074, pp. 252–259. [Google Scholar]
- Chang, J.M.; Hsu, C.C.; Wang, Y.L.; Ho, T.Y. Finding the set of all hinge vertices for strongly chordal graphs in linear time. Inf. Sci. 1997, 99, 173–182. [Google Scholar] [CrossRef]
- Ho, T.Y.; Wang, Y.L.; Juan, M.T. A linear time algorithm for finding all hinge vertices of a permutation graph. Inf. Process. Lett. 1996, 59, 103–107. [Google Scholar] [CrossRef]
- Fu, F.; Wang, L.; Nowak, M.A.; Hauert, C. Evolutionary dynamics on graphs: Efficient method for weak selection. Phys. Rev. E 2009, 79, 046707. [Google Scholar] [CrossRef]
- Mullon, C.; Lehmann, L. The robustness of the weak selection approximation for the evolution of altruism against strong selection. J. Evol. Biol. 2014, 27, 2272–2282. [Google Scholar] [CrossRef]
- Ohtsuki, H.; Nowak, M.A. The replicator equation on graphs. J. Theor. Biol. 2006, 243, 86–97. [Google Scholar] [CrossRef]
- Wu, B.; Altrock, P.M.; Wang, L.; Traulsen, A. Universality of weak selection. Phys. Rev. E 2010, 82, 046106. [Google Scholar] [CrossRef]
- Wu, B.; Bauer, B.; Galla, T.; Traulsen, A. Fitness–based models and pairwise comparison models of evolutionary games are typically different—Even in unstructured populations. New J. Phys. 2010, 17, 023043. [Google Scholar] [CrossRef]
- Xiao, Y.; Wu, B. Close spatial arrangement of mutants favors and disfavors fixation. PLoS Comput. Biol. 2019, 15, e1007212. [Google Scholar]
- Hindersin, L.; Möller, M.; Traulsen, A.; Bauer, B. Exact numerical calculation of fixation probability and time on graphs. BioSystems 2016, 150, 87–91. [Google Scholar] [CrossRef]
- Ibsen-Jensen, R.; Chatterjee, K.; Nowak, M.A. Computational complexity of ecological and evolutionary spatial dynamics. Proc. Natl. Acad. Sci. USA 2015, 112, 15636–15641. [Google Scholar] [CrossRef] [PubMed]
- Voorhees, B. Birth–death fixation probabilities for structured populations. Proc. R. Soc. A 2013, 469, 20120248. [Google Scholar] [CrossRef]
- McAvoy, A.; Hauert, C. Structural symmetry in evolutionary games. J. R. Soc. Interface 2015, 12, 20150420. [Google Scholar] [CrossRef] [PubMed]
- Meringer, M. Fast generation of regular graphs and construction of cages. J. Graph Theory 1999, 30, 137–146. [Google Scholar] [CrossRef]
- Lieberman, E.; Hauert, C.; Nowak, M.A. Evolutionary dynamics on graphs. Nature 2005, 433, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuki, H.; Pacheco, J.M.; Nowak, M.A. Evolutionary graph theory: Breaking the symmetry between interaction and replacement. J. Theor. Biol. 2007, 246, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.; Nowak, M.A. Games on graphs. EMS Surv. Math. Sci. 2014, 1, 113–151. [Google Scholar] [CrossRef]
- Pattni, K.; Broom, M.; Silvers, L.; Rychtar, J. Evolutionary graph theory revisited: When is an evolutionary process equivalent to the Moran process? Proc. R. Soc. A 2015, 471, 20150334. [Google Scholar] [CrossRef]
- Bondy, J.A.; Murty, U.S.R. Graph Theory; Springer: Berlin, Germany, 2008. [Google Scholar]
- Available online: https://github.com/HendrikRichterLeipzig/StructureCoefficientsRegularGraphs (accessed on 14 February 2020).






| 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
|---|---|---|---|---|---|---|---|---|---|
| 3 | 1 | 0 | 1 | 0 | 1 | 0 | 4 | 0 | 10 |
| 4 | 0 | 2 | 1 | 1 | 1 | 1 | 2 | 10 | 14 |
| 5 | 0 | 0 | 2 | 0 | 1 | 0 | 1 | 0 | 1 |
| 6 | 0 | 0 | 0 | 3 | 2 | 5 | 1 | 2 | 1 |
| 7 | 0 | 0 | 0 | 0 | 2 | 0 | 4 | 0 | 1 |
| 8 | 0 | 0 | 0 | 0 | 0 | 5 | 6 | 49 | 4 |
| 9 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 14 |
| 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 14 |
| 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richter, H. Evolution of Cooperation for Multiple Mutant Configurations on All Regular Graphs with N ≤ 14 Players. Games 2020, 11, 12. https://doi.org/10.3390/g11010012
Richter H. Evolution of Cooperation for Multiple Mutant Configurations on All Regular Graphs with N ≤ 14 Players. Games. 2020; 11(1):12. https://doi.org/10.3390/g11010012
Chicago/Turabian StyleRichter, Hendrik. 2020. "Evolution of Cooperation for Multiple Mutant Configurations on All Regular Graphs with N ≤ 14 Players" Games 11, no. 1: 12. https://doi.org/10.3390/g11010012
APA StyleRichter, H. (2020). Evolution of Cooperation for Multiple Mutant Configurations on All Regular Graphs with N ≤ 14 Players. Games, 11(1), 12. https://doi.org/10.3390/g11010012