
Wnt Drug Discovery: Weaving Through the Screens, Patents and Clinical Trials

Abstract
:1. Introduction
2. Methods
2.1. USPTO
2.2. ClinicalTrials.gov
2.3. Wnt Screens
3. Results
3.1. USPTO
3.1.1. Overall
3.1.2. Enhancers
3.1.3. Inhibitors
3.2. Clinical Trials
3.3. Wnt Screens
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fuchs, E. The tortoise and the hair: Slow-cycling cells in the stem cell race. Cell 2009, 137, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Haegebarth, A.; Clevers, H. Wnt signaling, lgr5, and stem cells in the intestine and skin. Am. J. Pathol. 2009, 174, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Wodarz, A.; Nusse, R. Mechanisms of Wnt signaling in development. Annu. Rev. Cell Dev. Biol. 1998, 14, 59–88. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/β-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Freese, J.L.; Pino, D.; Pleasure, S.J. Wnt signaling in development and disease. Neurobiol. Dis. 2010, 38, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Logan, C.Y.; Nusse, R. The Wnt signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol. 2004, 20, 781–810. [Google Scholar] [CrossRef] [PubMed]
- Angers, S.; Moon, R.T. Proximal events in Wnt signal transduction. Nat. Rev. Mol. Cell Biol. 2009, 10, 468–477. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, E.M.; Vermeulen, L.; Richel, D.; Medema, J.P. Targeting Wnt signaling in colon cancer stem cells. Clin. Cancer Res. 2010, 17, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Duchartre, Y.; Kim, Y.M.; Kahn, M. The Wnt signaling pathway in cancer. Crit. Rev. Oncol. Hematol. 2016, 99, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Le, P.N.; McDermott, J.D.; Jimeno, A. Targeting the Wnt pathway in human cancers: Therapeutic targeting with a focus on OMP-54F28. Pharmacol. Ther. 2015, 146, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Polakis, P. Drugging Wnt signalling in cancer. EMBO J. 2012, 31, 2737–2746. [Google Scholar] [CrossRef] [PubMed]
- Emami, K.H.; Nguyen, C.; Ma, H.; Kim, D.H.; Jeong, K.W.; Eguchi, M.; Moon, R.T.; Teo, J.L.; Kim, H.Y.; Moon, S.H.; et al. A small molecule inhibitor of β-catenin/CREB-binding protein transcription. Proc. Natl. Acad. Sci. USA 2004, 101, 12682–12687. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pan, S.; Hsieh, M.H.; Ng, N.; Sun, F.; Wang, T.; Kasibhatla, S.; Schuller, A.G.; Li, A.G.; Cheng, D.; et al. Targeting Wnt-driven cancer through the inhibition of porcupine by LGK974. Proc. Natl. Acad. Sci. USA 2013, 110, 20224–20229. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Nguyen, C.; Lee, K.S.; Kahn, M. Differential roles for the coactivators cbp and p300 on TCF/β-catenin-mediated survivin gene expression. Oncogene 2005, 24, 3619–3631. [Google Scholar] [CrossRef] [PubMed]
- De Vrieze, E.; Zethof, J.; Schulte-Merker, S.; Flik, G.; Metz, J.R. Identification of novel osteogenic compounds by an ex-vivo sp7:Luciferase zebrafish scale assay. Bone 2015, 74, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shan, J.; Chang, W.; Kim, I.; Bao, J.; Lee, H.J.; Zhang, X.; Samuel, V.T.; Shulman, G.I.; Liu, D.; et al. Chemical and genetic evidence for the involvement of Wnt antagonist Dickkopf2 in regulation of glucose metabolism. Proc. Natl. Acad. Sci. USA 2012, 109, 11402–11407. [Google Scholar] [CrossRef] [PubMed]
- Bertoldo, D.; Khan, M.M.; Dessen, P.; Held, W.; Huelsken, J.; Heinis, C. Phage selection of peptide macrocycles against β-catenin to interfere with Wnt signaling. Chem. Med. Chem. 2016, 11, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Anton, R.; Chatterjee, S.S.; Simundza, J.; Cowin, P.; Dasgupta, R. A systematic screen for micro-RNAs regulating the canonical Wnt pathway. PLoS ONE 2011, 6, e26257. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Su, Y.; Wesslowski, J.; Hagemann, A.I.; Ramialison, M.; Wittbrodt, J.; Scholpp, S.; Davidson, G. Tyrosine phosphorylation of LRP6 by Src and Fer inhibits Wnt/β-catenin signalling. EMBO Rep 2014, 15, 1254–1267. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Gwak, J.; Cho, M.; Ryu, M.J.; Lee, J.H.; Kim, S.K.; Kim, Y.H.; Lee, G.W.; Yun, M.Y.; Cuong, N.M.; et al. Murrayafoline a attenuates the Wnt/β-catenin pathway by promoting the degradation of intracellular β-catenin proteins. Biochem. Biophys. Res. Commun. 2010, 391, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Chuang, K.A.; Lieu, C.H.; Tsai, W.J.; Wu, M.H.; Chen, Y.C.; Liao, J.F.; Wang, C.C.; Kuo, Y.C. Evaluation of anti-Wnt/β-catenin signaling agents by PGL4-top transfected stable cells with a luciferase reporter system. Braz. J. Med. Biol. Res. 2010, 43, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Coombs, G.S.; Schmitt, A.A.; Canning, C.A.; Alok, A.; Low, I.C.; Banerjee, N.; Kaur, S.; Utomo, V.; Jones, C.M.; Pervaiz, S.; et al. Modulation of Wnt/β-catenin signaling and proliferation by a ferrous iron chelator with therapeutic efficacy in genetically engineered mouse models of cancer. Oncogene 2012, 31, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Coombs, G.S.; Yu, J.; Canning, C.A.; Veltri, C.A.; Covey, T.M.; Cheong, J.K.; Utomo, V.; Banerjee, N.; Zhang, Z.H.; Jadulco, R.C.; et al. Wls-dependent secretion of Wnt3a requires Ser209 acylation and vacuolar acidification. J. Cell Sci. 2010, 123, 3357–3367. [Google Scholar] [CrossRef] [PubMed]
- Dale, T.; Clarke, P.A.; Esdar, C.; Waalboer, D.; Adeniji-Popoola, O.; Ortiz-Ruiz, M.J.; Mallinger, A.; Samant, R.S.; Czodrowski, P.; Musil, D.; et al. A selective chemical probe for exploring the role of CDK8 and CDK19 in human disease. Nat. Chem. Bio. 2015, 11, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Duraiswamy, A.J.; Lee, M.A.; Madan, B.; Ang, S.H.; Tan, E.S.; Cheong, W.W.; Ke, Z.; Pendharkar, V.; Ding, L.J.; Chew, Y.S.; et al. Discovery and optimization of a porcupine inhibitor. J. Med. Chem. 2015, 58, 5889–5899. [Google Scholar] [CrossRef] [PubMed]
- Ewan, K.; Pajak, B.; Stubbs, M.; Todd, H.; Barbeau, O.; Quevedo, C.; Botfield, H.; Young, R.; Ruddle, R.; Samuel, L.; et al. A useful approach to identify novel small-molecule inhibitors of Wnt-dependent transcription. Cancer Res. 2010, 70, 5963–5973. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Bourhis, E.; Chiu, C.; Stawicki, S.; DeAlmeida, V.I.; Liu, B.Y.; Phamluong, K.; Cao, T.C.; Carano, R.A.; Ernst, J.A.; et al. Wnt isoform-specific interactions with coreceptor specify inhibition or potentiation of signaling by LRP6 antibodies. PLoS ONE 2010, 5, e12682. [Google Scholar] [CrossRef] [PubMed]
- Gurney, A.; Axelrod, F.; Bond, C.J.; Cain, J.; Chartier, C.; Donigan, L.; Fischer, M.; Chaudhari, A.; Ji, M.; Kapoun, A.M.; et al. Wnt pathway inhibition via the targeting of frizzled receptors results in decreased growth and tumorigenicity of human tumors. Proc. Natl. Acad. Sci. USA 2012, 109, 11717–11722. [Google Scholar] [CrossRef] [PubMed]
- Gwak, J.; Lee, J.H.; Chung, Y.H.; Song, G.Y.; Oh, S. Small molecule-based promotion of PKCα-mediated β-catenin degradation suppresses the proliferation of CRT-positive cancer cells. PLoS ONE 2012, 7, e46697. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Mishina, Y.M.; Liu, S.; Cheung, A.; Stegmeier, F.; Michaud, G.A.; Charlat, O.; Wiellette, E.; Zhang, Y.; Wiessner, S.; et al. Tankyrase inhibition stabilizes axin and antagonizes Wnt signalling. Nature 2009, 461, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Liu, J.X.; Endo, T.; Zhou, H.; Yao, S.; Willert, K.; Schmidt-Wolf, I.G.; Kipps, T.J.; Carson, D.A. Ethacrynic acid exhibits selective toxicity to chronic lymphocytic leukemia cells by inhibition of the Wnt/β-catenin pathway. PLoS ONE 2009, 4, e8294. [Google Scholar] [CrossRef] [PubMed]
- Madan, B.; Walker, M.P.; Young, R.; Quick, L.; Orgel, K.A.; Ryan, M.; Gupta, P.; Henrich, I.C.; Ferrer, M.; Marine, S.; et al. Usp6 oncogene promotes Wnt signaling by deubiquitylating frizzleds. Proc. Natl. Acad. Sci. USA 2016, 113, E2945–E2954. [Google Scholar] [CrossRef] [PubMed]
- Mallinger, A.; Crumpler, S.; Pichowicz, M.; Waalboer, D.; Stubbs, M.; Adeniji-Popoola, O.; Wood, B.; Smith, E.; Thai, C.; Henley, A.T.; et al. Discovery of potent, orally bioavailable, small-molecule inhibitors of Wnt signaling from a cell-based pathway screen. J. Med. Chem. 2015, 58, 1717–1735. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, M.W.; Coombs, G.S.; Banerjee, N.; Bugni, T.S.; Cannon, K.M.; Harper, M.K.; Veltri, C.A.; Virshup, D.M.; Ireland, C.M. Psammaplin A as a general activator of cell-based signaling assays via HDAC inhibition and studies on some bromotyrosine derivatives. Bioorg. Med. Chem. 2009, 17, 2189–2198. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Toume, K.; Arai, M.A.; Koyano, T.; Kowithayakorn, T.; Ishibashi, M. 2-methoxy-1,4-naphthoquinone isolated from impatiens balsamina in a screening program for activity to inhibit Wnt signaling. J. Nat. Med. 2011, 65, 234–236. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Dothager, R.S.; Marasa, J.; Lewis, C.L.; Piwnica-Worms, D. Vascular endothelial growth factor receptor-1 is synthetic lethal to aberrant β -catenin activation in colon cancer. Clin. Cancer Res. 2009, 15, 7529–7537. [Google Scholar] [CrossRef] [PubMed]
- Narwal, M.; Koivunen, J.; Haikarainen, T.; Obaji, E.; Legala, O.E.; Venkannagari, H.; Joensuu, P.; Pihlajaniemi, T.; Lehtio, L. Discovery of tankyrase inhibiting flavones with increased potency and isoenzyme selectivity. J. Med. Chem. 2013, 56, 7880–7889. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, K.; Toume, K.; Arai, M.A.; Koyano, T.; Kowithayakorn, T.; Mizoguchi, T.; Itoh, M.; Ishibashi, M. 9-hydroxycanthin-6-one, a β-carboline alkaloid from eurycoma longifolia, is the first wnt signal inhibitor through activation of glycogen synthase kinase 3β without depending on casein kinase 1α. J. Nat. Prod. 2015, 78, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Toume, K.; Arai, M.A.; Koyano, T.; Kowithayakorn, T.; Ishibashi, M. Β-sitosterol and flavonoids isolated from bauhinia malabarica found during screening for Wnt signaling inhibitory activity. J. Nat. Med. 2014, 68, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Gwak, J.; Cho, M.; Song, T.; Won, J.; Kim, D.E.; Shin, J.G.; Oh, S. Hexachlorophene inhibits Wnt/β-catenin pathway by promoting siah-mediated β-catenin degradation. Mol. Pharmacol. 2006, 70, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, J.H.; Kim, J.E.; Song, G.Y.; Zhou, W.; Goh, S.H.; Na, M.; Oh, S. Cytotoxic activity of aeroplysinin-1 against colon cancer cells by promoting β-catenin degradation. Food. Chem. Toxicol. 2016, 93, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, A.; Ho, S.Y.; Wang, W.; Alam, J.; Jeyaraj, D.A.; Ang, S.H.; Tan, E.S.; Lin, G.R.; Cheong, V.W.; Ke, Z.; et al. Pharmacophore model for wnt/porcupine inhibitors and its use in drug design. J. Chem. Inf. Model. 2015, 55, 1435–1448. [Google Scholar] [CrossRef] [PubMed]
- Toume, K.; Kamiya, K.; Arai, M.A.; Mori, N.; Sadhu, S.K.; Ahmed, F.; Ishibashi, M. Xylogranin b: A potent Wnt signal inhibitory limonoid from xylocarpus granatum. Org. Lett. 2013, 15, 6106–6109. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, M.; Cha, P.H.; Min do, S.; Choi, K.Y. Indirubin-3′-oxime reverses bone loss in ovariectomized and hindlimb-unloaded mice via activation of the Wnt/β-catenin signaling. J. Bone Miner. Res. 2014, 29, 1196–1205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kong, L.M.; Zhan, R.; Ye, Z.N.; Pu, J.X.; Sun, H.D.; Li, Y. Two natural ent-kauranoids as novel Wnt signaling inhibitors. Nat. Prod. Bioprospect. 2014, 4, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Major, M.B.; Takanashi, S.; Camp, N.D.; Nishiya, N.; Peters, E.C.; Ginsberg, M.H.; Jian, X.; Randazzo, P.A.; Schultz, P.G.; et al. Small-molecule synergist of the wnt/β-catenin signaling pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 7444–7448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sviripa, V.; Kril, L.M.; Chen, X.; Yu, T.; Shi, J.; Rychahou, P.; Evers, B.M.; Watt, D.S.; Liu, C. Fluorinated N,N-dialkylaminostilbenes for Wnt pathway inhibition and colon cancer repression. J. Med. Chem. 2011, 54, 1288–1297. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ohtsuki, T.; Koyano, T.; Kowithayakorn, T.; Ishibashi, M. New Wnt/β-catenin signaling inhibitors isolated from eleutherine palmifolia. Chem. Asian. J. 2009, 4, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Christova, T.; Perusini, S.; Alizadeh, S.; Bao, R.Y.; Miller, B.W.; Hurren, R.; Jitkova, Y.; Gronda, M.; Isaac, M.; et al. Wnt inhibitor screen reveals iron dependence of β-catenin signaling in cancers. Cancer Res. 2011, 71, 7628–7639. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, C.; Lan, W.; Huang, C.; Lin, M.; Wang, Z.; Liang, W.; Iwamoto, A.; Yang, X.; Liu, H. Gliotoxin inhibits proliferation and induces apoptosis in colorectal cancer cells. Mar. Drugs 2015, 13, 6259–6273. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.Y.; Deng, X.; Byun, S.; Lee, C.; Lee, S.J.; Suh, H.; Zhang, J.; Kang, Q.; Zhang, T.; Westover, K.D.; et al. Direct targeting of β-catenin by a small molecule stimulates proteasomal degradation and suppresses oncogenic Wnt/β-catenin signaling. Cell. Rep. 2016, 16, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Lan, L.; Appelman, C.; Smith, A.R.; Yu, J.; Larsen, S.; Marquez, R.T.; Liu, H.; Wu, X.; Gao, P.; Roy, A.; et al. Natural product (−)-gossypol inhibits colon cancer cell growth by targeting rna-binding protein musashi-1. Mol. Oncol. 2015, 9, 1406–1420. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Lim, T.G.; Chen, H.; Jung, S.K.; Lee, H.J.; Lee, M.H.; Kim, D.J.; Shin, A.; Lee, K.W.; Bode, A.M.; et al. Esculetin suppresses proliferation of human colon cancer cells by directly targeting β-catenin. Cancer Prev. Res. 2013, 6, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Ashihara, E.; Strovel, J.W.; Nakagawa, Y.; Kuroda, J.; Nagao, R.; Tanaka, R.; Yokota, A.; Takeuchi, M.; Hayashi, Y.; et al. Av-65, a novel Wnt/β-catenin signal inhibitor, successfully suppresses progression of multiple myeloma in a mouse model. Blood Cancer J. 2011. [Google Scholar] [CrossRef] [PubMed]
- Shan, B.E.; Wang, M.X.; Li, R.Q. Quercetin inhibit human SW480 colon cancer growth in association with inhibition of cyclin D1 and survivin expression through Wnt/β-catenin signaling pathway. Cancer Invest. 2009, 27, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Canal, F.; Anthony, E.; Lescure, A.; Del Nery, E.; Camonis, J.; Perez, F.; Ragazzon, B.; Perret, C. A kinome siRNA screen identifies HGS as a potential target for liver cancers with oncogenic mutations in CTNNB1. BMC Cancer 2015. [Google Scholar] [CrossRef] [PubMed]
- Conrad, W.; Major, M.B.; Cleary, M.A.; Ferrer, M.; Roberts, B.; Marine, S.; Chung, N.; Arthur, W.T.; Moon, R.T.; Berndt, J.D.; et al. FAM129B is a novel regulator of Wnt/β-catenin signal transduction in melanoma cells. F1000Research 2013. [Google Scholar] [CrossRef]
- De Robertis, A.; Valensin, S.; Rossi, M.; Tunici, P.; Verani, M.; De Rosa, A.; Giordano, C.; Varrone, M.; Nencini, A.; Pratelli, C.; et al. Identification and characterization of a small-molecule inhibitor of Wnt signaling in glioblastoma cells. Mol. Cancer Ther. 2013, 12, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Gonsalves, F.C.; Klein, K.; Carson, B.B.; Katz, S.; Ekas, L.A.; Evans, S.; Nagourney, R.; Cardozo, T.; Brown, A.M.; DasGupta, R. An RNAi-based chemical genetic screen identifies three small-molecule inhibitors of the Wnt/wingless signaling pathway. Proc. Natl. Acad. Sci. USA 2011, 108, 5954–5963. [Google Scholar] [CrossRef] [PubMed]
- Gurrola-Diaz, C.M.; Garcia-Lopez, P.M.; Gulewicz, K.; Pilarski, R.; Dihlmann, S. Inhibitory mechanisms of two Uncaria tomentosa extracts affecting the Wnt-signaling pathway. Phytomedicine 2011, 18, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Handeli, S.; Simon, J.A. A small-molecule inhibitor of TCF/β-catenin signaling down-regulates ppargamma and ppardelta activities. Mol. Cancer Ther. 2008, 7, 521–529. [Google Scholar] [CrossRef] [PubMed]
- James, R.G.; Biechele, T.L.; Conrad, W.H.; Camp, N.D.; Fass, D.M.; Major, M.B.; Sommer, K.; Yi, X.; Roberts, B.S.; Cleary, M.A.; et al. Bruton’s tyrosine kinase revealed as a negative regulator of Wnt-β-catenin signaling. Sci. Signal 2009. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Park, Y.I.; Dong, M.S.; Gong, Y.D. Identification of 2,3,6-trisubstituted quinoxaline derivatives as a Wnt2/β-catenin pathway inhibitor in non-small-cell lung cancer cell lines. Bioorg. Med. Chem. Lett. 2010, 20, 5900–5904. [Google Scholar] [CrossRef] [PubMed]
- Mazur, M.; Bujak, A.; Matloka, M.; Janowska, S.; Gunerka, P.; Bojarski, L.; Stanczak, A.; Klejman, A.; Bednarek, A.; Lamparska-Przybysz, M.; et al. Cell-based assay for low- and high-scale screening of the Wnt/β-catenin signaling modulators. Anal. Biochem. 2015, 475, 56–67. [Google Scholar] [CrossRef]
- Okada-Iwasaki, R.; Takahashi, Y.; Watanabe, Y.; Ishida, H.; Saito, J.I.; Nakai, R.; Asai, A. The discovery and characterization of K-756, a novel Wnt/β-catenin pathway inhibitor targeting tankyrase. Mol. Cancer Ther. 2016, 15, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.N.; Cheng, C.; Theriault, K.M.; Sheridan, S.D.; Tsai, L.H.; Haggarty, S.J. A high-throughput screen for Wnt/β-catenin signaling pathway modulators in human IPSC-derived neural progenitors. J. Biomol. Screen 2012, 17, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- James, R.G.; Davidson, K.C.; Bosch, K.A.; Biechele, T.L.; Robin, N.C.; Taylor, R.J.; Major, M.B.; Camp, N.D.; Fowler, K.; Martins, T.J.; et al. WIKI4, a novel inhibitor of tankyrase and Wnt/ss-catenin signaling. PLoS ONE 2012, 7, e50457. [Google Scholar] [CrossRef] [PubMed]
- Verkaar, F.; van der Doelen, A.A.; Smits, J.F.; Blankesteijn, W.M.; Zaman, G.J. Inhibition of Wnt/β-catenin signaling by p38 MAP kinase inhibitors is explained by cross-reactivity with casein kinase Iδ/ɛ. Chem. Biol. 2011, 18, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, J.; Lu, J.; Bond, M.C.; Ren, X.R.; Lyerly, H.K.; Barak, L.S.; Chen, W. The anti-helminthic niclosamide inhibits Wnt/Frizzled1 signaling. Biochemistry 2009, 48, 10267–10274. [Google Scholar] [CrossRef] [PubMed]
- Borchert, K.M.; Galvin, R.J.; Frolik, C.A.; Hale, L.V.; Halladay, D.L.; Gonyier, R.J.; Trask, O.J.; Nickischer, D.R.; Houck, K.A. High-content screening assay for activators of the Wnt/Fzd pathway in primary human cells. Assay. Drug Dev. Technol. 2005, 3, 133–141. [Google Scholar] [CrossRef] [PubMed]
- De la Roche, M.; Rutherford, T.J.; Gupta, D.; Veprintsev, D.B.; Saxty, B.; Freund, S.M.; Bienz, M. An intrinsically labile α-helix abutting the BCL9-binding site of β-catenin is required for its inhibition by carnosic acid. Nat. Commun. 2012. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Zhu, Q.; Neuenschwander, M.; Specker, E.; Wulf-Goldenberg, A.; Weis, W.I.; von Kries, J.P.; Birchmeier, W. A small-molecule antagonist of the β-catenin/tcf4 interaction blocks the self-renewal of cancer stem cells and suppresses tumorigenesis. Cancer Res. 2016, 76, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Catrow, J.L.; Ji, H. High-throughput selectivity assays for small-molecule inhibitors of β-catenin/T-cell factor protein-protein interactions. ACS Med. Chem. Lett. 2013, 4, 306–311. [Google Scholar] [CrossRef] [PubMed]
- An, W.F.; Germain, A.R.; Bishop, J.A.; Nag, P.P.; Metkar, S.; Ketterman, J.; Walk, M.; Weiwer, M.; Liu, X.; Patnaik, D.; et al. Discovery of potent and highly selective inhibitors of GSK3B. In Probe Reports From the NIH Molecular Libraries Program; National Center for Biotechnology Information: Bethesda, MD, USA, 2010. [Google Scholar]
- Trosset, J.Y.; Dalvit, C.; Knapp, S.; Fasolini, M.; Veronesi, M.; Mantegani, S.; Gianellini, L.M.; Catana, C.; Sundstrom, M.; Stouten, P.F.; et al. Inhibition of protein-protein interactions: The discovery of druglike β-catenin inhibitors by combining virtual and biophysical screening. Proteins 2006, 64, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Grandy, D.; Shan, J.; Zhang, X.; Rao, S.; Akunuru, S.; Li, H.; Zhang, Y.; Alpatov, I.; Zhang, X.A.; Lang, R.A.; et al. Discovery and characterization of a small molecule inhibitor of the PDZ domain of dishevelled. J. Biol. Chem. 2009, 284, 16256–16263. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.; Shi, D.L.; Wang, J.; Zheng, J. Identification of a specific inhibitor of the dishevelled PDZ domain. Biochemistry 2005, 44, 15495–15503. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.; Zheng, J.J. Virtual ligand screening combined with nmr to identify DVL PDZ domain inhibitors targeting the wnt signaling. Methods Mol. Biol. 2012, 928, 17–28. [Google Scholar] [PubMed]
- Koval, A.V.; Vlasov, P.; Shichkova, P.; Khunderyakova, S.; Markov, Y.; Panchenko, J.; Volodina, A.; Kondrashov, F.A.; Katanaev, V.L. Anti-leprosy drug clofazimine inhibits growth of triple-negative breast cancer cells via inhibition of canonical Wnt signaling. Biochem. Pharmacol. 2014, 87, 571–78. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Bao, J.; Miller, A.; Zhang, C.; Wu, J.; Baday, Y.C.; Guibao, C.; Li, L.; Wu, D.; Zheng, J.J. Structure-based discovery of novel small molecule Wnt signaling inhibitors by targeting the cysteine-rich domain of frizzled. J. Biol. Chem. 2015, 290, 30596–30606. [Google Scholar] [CrossRef] [PubMed]
- Hammad, M.A.; Azam, S.S. Structural dynamics and inhibitor searching for Wnt-4 protein using comparative computational studies. Drug Des. Devel. Ther. 2015, 9, 2449–2461. [Google Scholar] [PubMed]
- Thorne, C.A.; Hanson, A.J.; Schneider, J.; Tahinci, E.; Orton, D.; Cselenyi, C.S.; Jernigan, K.K.; Meyers, K.C.; Hang, B.I.; Waterson, A.G.; et al. Small-molecule inhibition of wnt signaling through activation of casein kinase 1α. Nat. Chem. Biol. 2010, 6, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.K.; Danzmann, K.; Charles, S.; Blake, K.; Olivares, A.; Bamikole, S.; Olson, M.; van Raay, T.J. Targeting the Wnt pathway in zebrafish as a screening method to identify novel therapeutic compounds. Exp. Biol. Med. 2014, 239, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Lin, S. Chemical screening with zebrafish embryos. Methods Mol. Biol. 2011, 716, 193–205. [Google Scholar] [PubMed]
- Hao, J.; Ao, A.; Zhou, L.; Murphy, C.K.; Frist, A.Y.; Keel, J.J.; Thorne, C.A.; Kim, K.; Lee, E.; Hong, C.C. Selective small molecule targeting β-catenin function discovered by in vivo chemical genetic screen. Cell Rep. 2013, 4, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Nishiya, N.; Oku, Y.; Kumagai, Y.; Sato, Y.; Yamaguchi, E.; Sasaki, A.; Shoji, M.; Ohnishi, Y.; Okamoto, H.; Uehara, Y. A zebrafish chemical suppressor screening identifies small molecule inhibitors of the Wnt/β-catenin pathway. Chem. Biol. 2014, 21, 530–540. [Google Scholar] [CrossRef] [PubMed]
- De Groot, R.E.; Ganji, R.S.; Bernatik, O.; Lloyd-Lewis, B.; Seipel, K.; Sedova, K.; Zdrahal, Z.; Dhople, V.M.; Dale, T.C.; Korswagen, H.C.; et al. Huwe1-mediated ubiquitylation of dishevelled defines a negative feedback loop in the Wnt signaling pathway. Sci. Signal 2014. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Curtin, J.C.; Lorenzi, M.V. Drug discovery approaches to target Wnt signaling in cancer stem cells. Oncotarget 2012, 1, 563–577. [Google Scholar] [CrossRef]
- Garber, K. Drugging the Wnt pathway: Problems and progress. J. Natl. Cancer Inst. 2009, 101, 548–550. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/β-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Pattabiraman, D.R.; Weinberg, R.A. Tackling the cancer stem cells—What challenges do they pose? Nat. Rev. Drug Discov. 2014, 13, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Rosenbluh, J.; Wang, X.; Hahn, W.C. Genomic insights into Wnt/β-catenin signaling. Trends Pharmacol. Sci. 2014, 35, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Sawyers, C. Targeted cancer therapy. Nature 2004, 432, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hao, J. Development of anticancer agents targeting the Wnt/β-catenin signaling. Am. J. Cancer Res. 2015, 5, 2344–2360. [Google Scholar] [PubMed]
- Zimmerman, Z.F.; Moon, R.T.; Chien, A.J. Targeting Wnt pathways in disease. Cold Spring Harb. Perspect. Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Network, T.C.G.A. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar]
- Regad, T. Targeting RTK signaling pathways in cancer. Cancers 2015, 7, 1758–1784. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Watkins, D.N.; Jair, K.W.; Schuebel, K.E.; Markowitz, S.D.; Chen, W.D.; Pretlow, T.P.; Yang, B.; Akiyama, Y.; Van Engeland, M.; et al. Epigenetic inactivation of SFRP genes allows constitutive Wnt signaling in colorectal cancer. Nat. Genet. 2004, 36, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Mazieres, J.; He, B.; You, L.; Xu, Z.; Lee, A.Y.; Mikami, I.; Reguart, N.; Rosell, R.; McCormick, F.; Jablons, D.M. Wnt inhibitory factor-1 is silenced by promoter hypermethylation in human lung cancer. Cancer Res. 2004, 64, 4717–4720. [Google Scholar] [CrossRef] [PubMed]
- Surana, R.; Sikka, S.; Cai, W.; Shin, E.M.; Warrier, S.R.; Tan, H.J.; Arfuso, F.; Fox, S.A.; Dharmarajan, A.M.; Kumar, A.P. Secreted frizzled related proteins: Implications in cancers. Biochim. Biophys. Acta. 2014, 1845, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Wissmann, C.; Wild, P.J.; Kaiser, S.; Roepcke, S.; Stoehr, R.; Woenckhaus, M.; Kristiansen, G.; Hsieh, J.C.; Hofstaedter, F.; Hartmann, A.; et al. WIF1, a component of the Wnt pathway, is down-regulated in prostate, breast, lung, and bladder cancer. J. Pathol. 2003, 201, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Korinek, V.; Barker, N.; Morin, P.J.; van Wichen, D.; de Weger, R.; Kinzler, K.W.; Vogelstein, B.; Clevers, H. Constitutive transcriptional activation by a β-catenin-TCF complex in APC−/− colon carcinoma. Science 1997, 275, 1784–1787. [Google Scholar] [CrossRef] [PubMed]
- Veeman, M.T.; Slusarski, D.C.; Kaykas, A.; Louie, S.H.; Moon, R.T. Zebrafish prickle, a modulator of noncanonical Wnt/fz signaling, regulates gastrulation movements. Curr. Biol. 2003, 13, 680–685. [Google Scholar] [CrossRef]
- Albuquerque, C.; Baltazar, C.; Filipe, B.; Penha, F.; Pereira, T.; Smits, R.; Cravo, M.; Lage, P.; Fidalgo, P.; Claro, I.; et al. Colorectal cancers show distinct mutation spectra in members of the canonical wnt signaling pathway according to their anatomical location and type of genetic instability. Genes Chromosomes Cancer 2010, 49, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Christie, M.; Jorissen, R.N.; Mouradov, D.; Sakthianandeswaren, A.; Li, S.; Day, F.; Tsui, C.; Lipton, L.; Desai, J.; Jones, I.T.; et al. Different APC genotypes in proximal and distal sporadic colorectal cancers suggest distinct Wnt/β-catenin signalling thresholds for tumourigenesis. Oncogene 2013, 32, 4675–4682. [Google Scholar] [CrossRef] [PubMed]
- Leedham, S.J.; Rodenas-Cuadrado, P.; Howarth, K.; Lewis, A.; Mallappa, S.; Segditsas, S.; Davis, H.; Jeffery, R.; Rodriguez-Justo, M.; Keshav, S.; et al. A basal gradient of wnt and stem-cell number influences regional tumour distribution in human and mouse intestinal tracts. Gut 2013, 62, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, H.; Rashid, S. Molecular docking studies of flavonoids for their inhibition pattern against β-catenin and pharmacophore model generation from experimentally known flavonoids to fabricate more potent inhibitors for Wnt signaling pathway. Pharmacogn. Mag. 2014, 10, S264–S271. [Google Scholar] [PubMed]
- Koval, A.; Katanaev, V.L. Platforms for high-throughput screening of Wnt/Frizzled antagonists. Drug Discov. Today 2012, 17, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Bol, D.; Ebner, R. Gene expression profiling in the discovery, optimization and development of novel drugs: One universal screening platform. Pharmacogenomics 2006, 7, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Attayek, P.J.; Ahmad, A.A.; Wang, Y.; Williamson, I.; Sims, C.E.; Magness, S.T.; Allbritton, N.L. In vitro polarization of colonoids to create an intestinal stem cell compartment. PLoS ONE 2016, 11, e0153795. [Google Scholar] [CrossRef] [PubMed]
- Drost, J.; Karthaus, W.R.; Gao, D.; Driehuis, E.; Sawyers, C.L.; Chen, Y.; Clevers, H. Organoid culture systems for prostate epithelial and cancer tissue. Nat. Protoc. 2016, 11, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ahmad, A.A.; Shah, P.K.; Sims, C.E.; Magness, S.T.; Allbritton, N.L. Capture and 3D culture of colonic crypts and colonoids in a microarray platform. Lab Chip 2013, 13, 4625–4634. [Google Scholar] [CrossRef] [PubMed]
- Farin, H.F.; Jordens, I.; Mosa, M.H.; Basak, O.; Korving, J.; Tauriello, D.V.; de Punder, K.; Angers, S.; Peters, P.J.; Maurice, M.M.; et al. Visualization of a short-range wnt gradient in the intestinal stem-cell niche. Nature 2016, 530, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Van de Wetering, M.; Francies, H.E.; Francis, J.M.; Bounova, G.; Iorio, F.; Pronk, A.; van Houdt, W.; van Gorp, J.; Taylor-Weiner, A.; Kester, L.; et al. Prospective derivation of a living organoid biobank of colorectal cancer patients. Cell 2015, 161, 933–945. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Search Term | “(ABST/cancer AND ABST/Wnt)” or “(ABST/cancer AND ABST/-catenin)” | Wnt Inhibitor |
|---|---|---|
| Patents | 94 | 101 |
| Patent Application | 203 | 276 |
| Total | 297 | 377 |
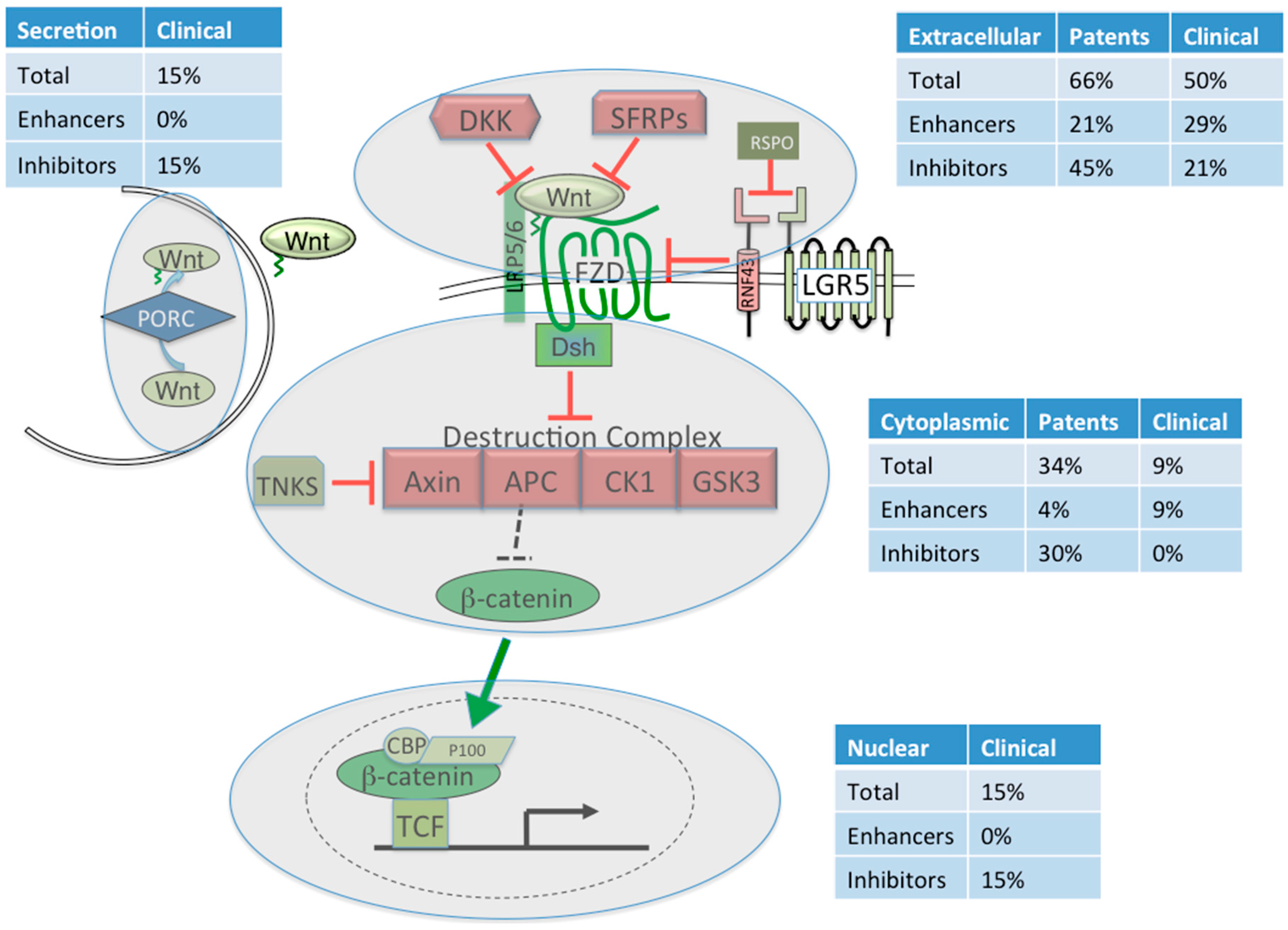
| Cellular Space | Subject of the Patent | Total | Effect on Wnt signaling |
|---|---|---|---|
| Extracellular | N = 68 (66%) | ||
| DKK as a target | 12 | Enhance | |
| DKK as a therapeutic | 13 | Inhibit | |
| Wnt ligand as a target | 11 | Inhibit | |
| Wnt ligand as a therapeutic | 7 | Enhance | |
| Wnt4a Patent #6165751 | 1 | Unknown 1 | |
| Frizzled/Soluble Frizzled | 13 | Inhibit | |
| LRP5/6 | 7 | Inhibit | |
| RSPO as a target | 1 | Inhibit | |
| RSPO as a therapeutic | 1 | Enhance | |
| TIKI1/2 as a target | 1 | Enhance | |
| SOST as a target | 1 | Enhance | |
| Intracellular | N = 35 (34%) | ||
| β-catenin as a target | 21 | Inhibit | |
| β-catenin as a therapeutic | 2 | Enhance | |
| β-catenin Patent PA #20050171005 | 1 | Inhibit and Enhance 2 | |
| Axin1/2 as a target | 6 | Inhibit | |
| Dishevelled as a target | 1 | Inhibit | |
| Casein Kinase 1 as a target | 2 | Inhibit | |
| GSK3 as a target | 2 | Enhance | |
| Total | 103 |
| Method | Number of Patents | Percentage of Patents |
|---|---|---|
| Antibodies | 25 | 25% |
| RNA mediated | 4 | 4% |
| Polypeptides/proteins | 53 | 51% |
| Chemicals | 21 | 20% |
| Total | 103 | 100% |
| Target | Number of Clinical Trials | Clinical Trial from Table S2 |
|---|---|---|
| Porcupine | 5 | 1, 2, 3, 13, 15 |
| Wnt Ligands | 1 | 23 |
| Frizzled Receptor | 4 | 6, 9, 12, 19 |
| LRP5/6 | 2 | 24, 25 |
| CBP/β-catenin | 5 | 4, 8, 10, 16, 18 |
| Epigenetic | 1 | 21 |
| Unspecified | 1 | 11 |
| Total | 19 |
| Target | Number of Clincal Trials | Clinical Trial from Table S2 |
|---|---|---|
| Wnt Ligands | 1 | 5 |
| Dkk | 9 | 22, 26, 27, 28, 29, 31, 32, 33, 34 |
| GSK3 | 3 | 7, 14, 17 |
| Epigenetic | 1 | 30 |
| Unspecified | 1 | 20 |
| Total | 15 |
| Condition | Number of Clinical Trials | Clinical Trial from Table S2 |
|---|---|---|
| Osteoporosis | 2 | 5, 31 |
| Osteopenia | 1 | 31, |
| Alopecia | 2 | 7, 20 |
| Male Pattern Baldness | 1 | 7, |
| Alzheimer’s disease | 1 | 14 |
| Leukemia | 1 | 17 |
| Multiple Myeloma | 6 | 22, 27, 28, 29, 33, 34 |
| Bone Disease * | 1 | 27 |
| Renal Insufficiency * | 1 | 28 |
| Cholangiocarcinoma | 1 | 26 |
| Gastro-esophageal Cancers | 1 | 32 |
| Gall Bladder Cancer | 1 | 26 |
| Bile Duct Cancer | 1 | 26 |
| Total | 20 |
| Condition | Number of Clinical Trails | Clinical Trial from Table S2 |
|---|---|---|
| Colorectal Carcinoma | 7 | 1, 3, 4, 6, 12, 21, 30 |
| Squamous Cell Carcinoma | 2 | 2, 32 |
| Head and Neck | 1 | 2, |
| Pancreatic Cancer | 2 | 3, 8, |
| Breast Cancer | 2 | 6, 9, |
| Myeloid Leukemia | 2 | 10, 16 |
| Basal Cell Carcinoma | 1 | 11, |
| Synovial Sarcoma | 1 | 19, |
| Non-Small Cell Lung Cancer | 1 | 22 |
| Solid Tumors | 5 | 13, 15, 18, 22, 23 |
| Prostate Cancer | 4 | 6, 9, 24, 25 |
| Total | 28 |
| Discovery Platform | Number of Assays | Assay | Reference |
|---|---|---|---|
| Cell Based | |||
| HEK293 | 31 | TCF/Lef reporter | [18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48] |
| HEK293 | 1 | β-catenin-luciferase stability | [49] |
| HCT116 | 5 | TCF/Lef reporter | [50,51,52,53,54] |
| SW480 | 2 | TCF/Lef reporter | [55] USPTO Patent# 8987298 1 |
| TM3 | 2 | TCF/Lef reporter | [13] USPTO PA# 20110237573 2 |
| Others 3 | 12 | TCF/Lef reporter | [56,57,58,59,60,61,62,63,64,65,66,67] |
| U2OS | 1 | Nuclear β-catenin | [68] |
| U2OS | 1 | Frizzled-1 GFP endocytosis | [69] |
| Preosteoblasts | 1 | Nuclear β-catenin | [70] |
| Substrate based | |||
| Elisa-based | 3 | Inhibitors of β-catenin-substrate interaction 4 | [71,72,73] |
| Elisa-based | 1 | GST-GSK3 with ADP-Glo | [74] |
| Biotinylated β-cat ARM repeats | 1 | Phage Display | [17] |
| In Silico Based | |||
| β-catenin-TCF 3D model | 1 | Structure based screen | [53] |
| β-catenin-TCF 3D model | 1 | NMR screening | [75] |
| Dvl PDZ 3D model | 3 | Structure based screen | [76,77,78] |
| DKK-LRP6 3D model | 1 | Structure based screen | [16] |
| FZD8-Wnt8 3D model | 2 | Structure based screen | [79,80] |
| Wnt3a 3D model | 1 | Structure based screen | [81] |
| Model Based | |||
| Xenopus Egg Extract | 1 | Axin-renilla luciferase: β-catenin-Firefly luciferase ratio | [82] |
| Zebrafish | 4 | Phenotypic screen | [83,84,85,86] |
| Zebrafish Ex vivo scales | 1 | SP7: Luciferase | [15] |
| C. elegans | 1 | β-catenin dependent QL.D cell migration phenotype | [87] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, B.; Green, B.A.; Farr, J.M.; Lopes, F.C.M.; Van Raay, T.J. Wnt Drug Discovery: Weaving Through the Screens, Patents and Clinical Trials. Cancers 2016, 8, 82. https://doi.org/10.3390/cancers8090082
Lu B, Green BA, Farr JM, Lopes FCM, Van Raay TJ. Wnt Drug Discovery: Weaving Through the Screens, Patents and Clinical Trials. Cancers. 2016; 8(9):82. https://doi.org/10.3390/cancers8090082
Chicago/Turabian StyleLu, Benjamin, Brooke A. Green, Jacqueline M. Farr, Flávia C.M. Lopes, and Terence J. Van Raay. 2016. "Wnt Drug Discovery: Weaving Through the Screens, Patents and Clinical Trials" Cancers 8, no. 9: 82. https://doi.org/10.3390/cancers8090082
APA StyleLu, B., Green, B. A., Farr, J. M., Lopes, F. C. M., & Van Raay, T. J. (2016). Wnt Drug Discovery: Weaving Through the Screens, Patents and Clinical Trials. Cancers, 8(9), 82. https://doi.org/10.3390/cancers8090082