Bacterial Extracellular Vesicles in Oncology: Molecular Mechanisms and Future Clinical Applications


Simple Summary
Abstract

1. Introduction
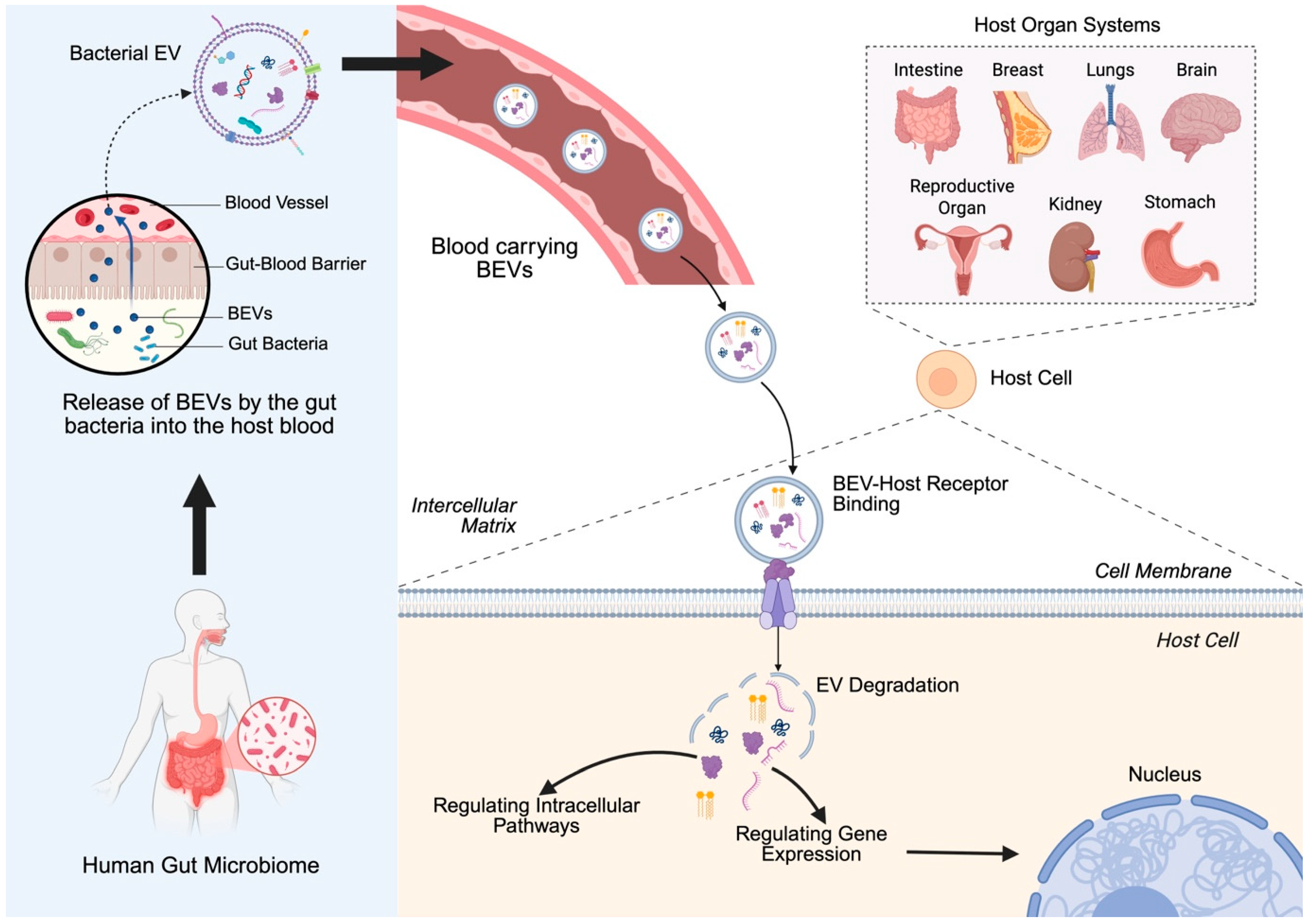
2. Bacterial Extracellular Vesicles (BEVs)
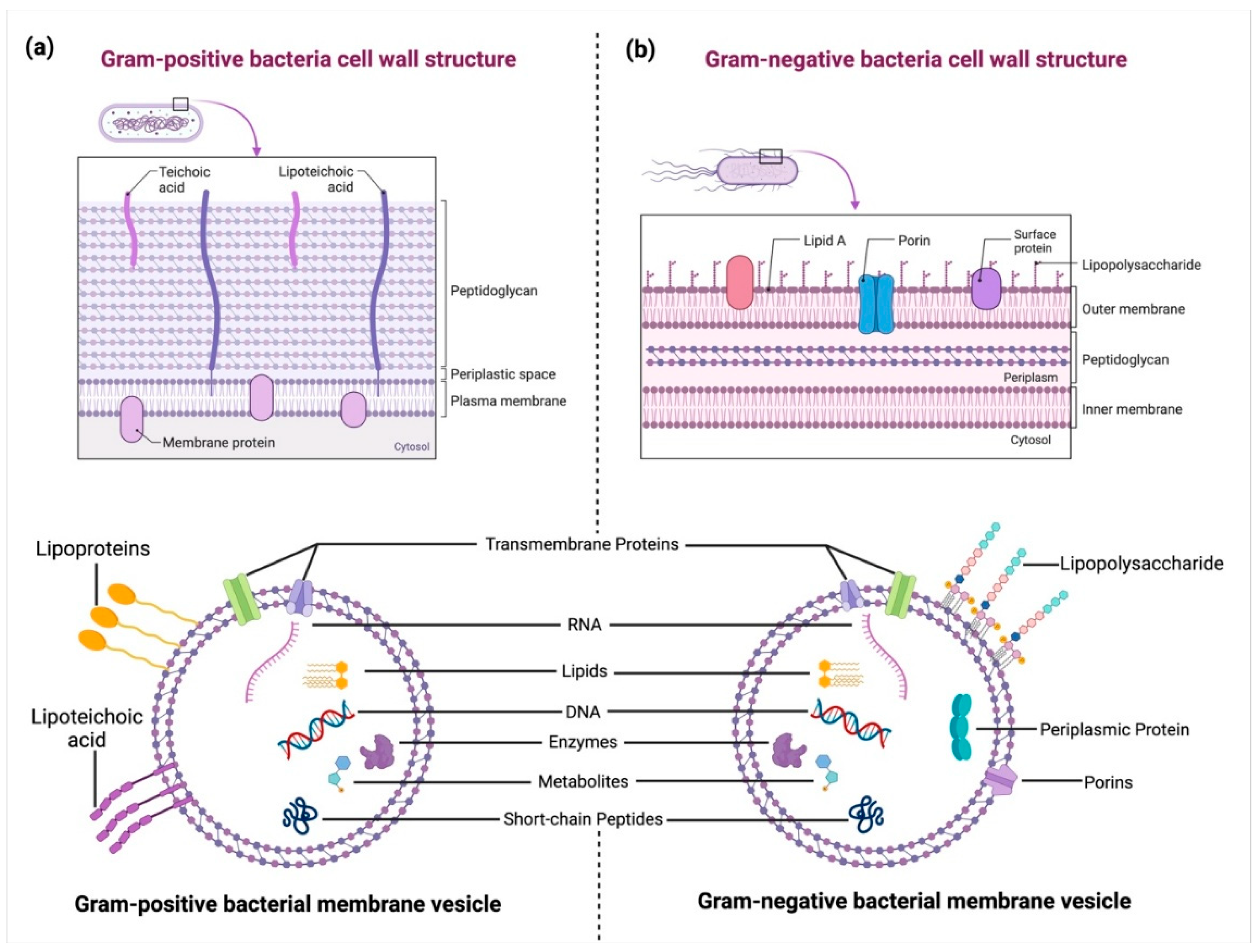
2.1. Biology of Bacterial Extracellular Vesicles
2.2. Molecular Composition of BEVs
2.3. Mechanisms of BEV Formation
2.4. Isolation and Characterization of BEVs
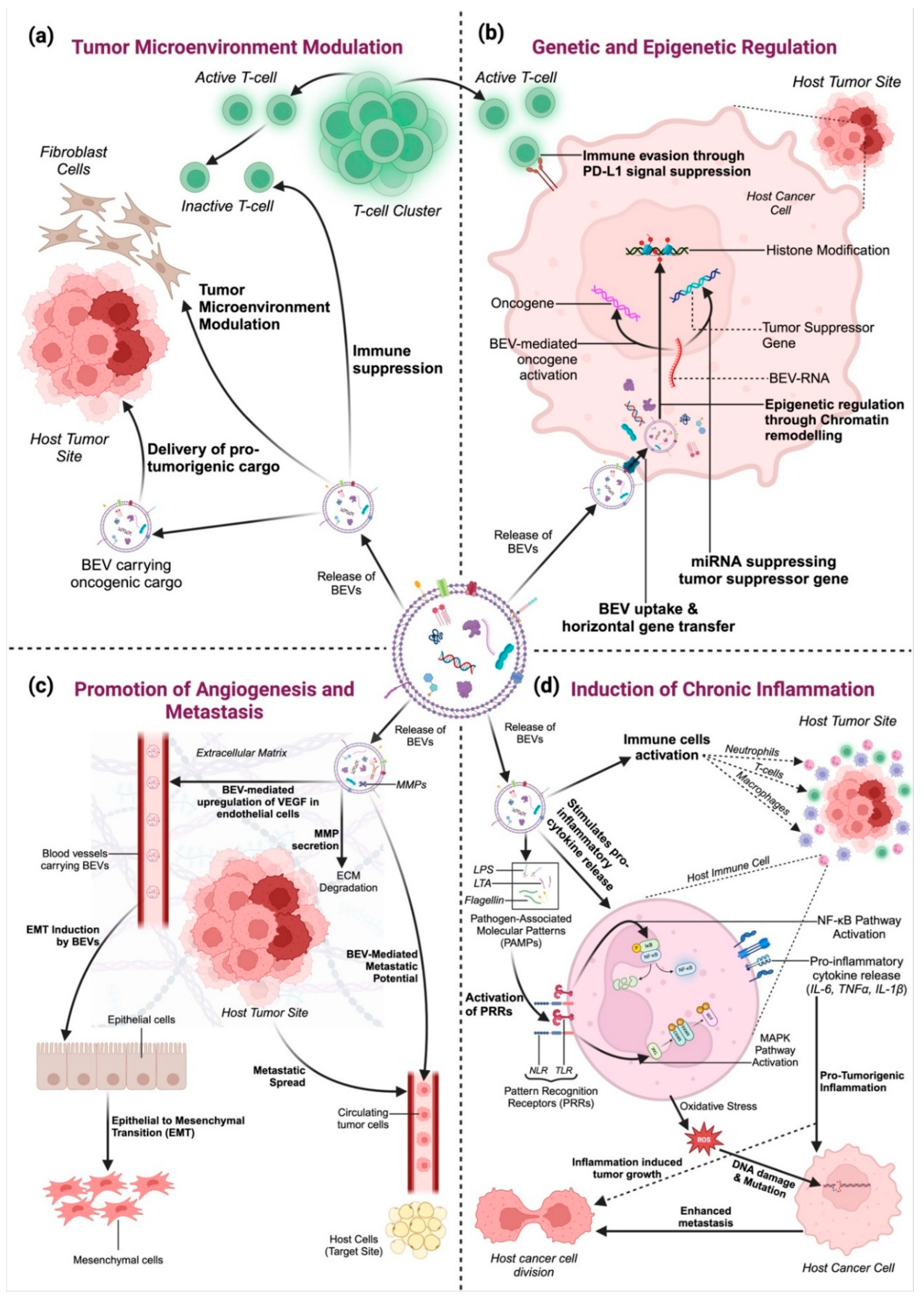
3. Role of BEVs in Cancer
3.1. Tumor Microenvironment Modulation
3.2. Genetic and Epigenetic Regulation
3.3. Promotion of Angiogenesis and Metastasis
3.4. Induction of Chronic Inflammation
4. BEVs Across Human Cancer Types
4.1. Colorectal Cancer (CRC)
4.2. Gastric Cancer (GC)
4.3. Breast Cancer
4.4. Lung Cancer
4.5. Brain Cancer
4.6. Renal and Bladder Cancers
4.7. Ovarian Cancer
5. Role of BEVs in Tumor Management
5.1. BEVs as Diagnostic Biomarkers
5.2. BEVs as Prognostic Indicators
5.3. BEVs as Therapeutic Agents
5.3.1. Drug Delivery
5.3.2. Cancer Vaccines
5.3.3. Immune Checkpoint Modulation
5.3.4. Activation of Innate and Adaptive Immunity
6. Challenges and Future Directions
6.1. Challenges in BEV Research
6.1.1. Standardization of Isolation and Characterization Techniques
6.1.2. Understanding BEV Heterogeneity
6.1.3. Biases in Study Design
6.1.4. Clinical Translation
6.1.5. Mechanistic Insights
6.2. Future Directions
6.2.1. BEVs as Precision Medicine Tools
6.2.2. Engineering BEVs for Therapeutic Applications
6.2.3. Emerging Innovations in BEV Engineering
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
List of Abbreviations
References
- Liang, X.; Dai, N.; Sheng, K.; Lu, H.; Wang, J.; Chen, L.; Wang, Y. Gut bacterial extracellular vesicles: Important players in regulating intestinal microenvironment. Gut Microbes 2022, 14, 2134689. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-W.; Um, J.-H.; Cho, J.-H.; Lee, H.-J. Tiny RNAs and their voyage via extracellular vesicles: Secretion of bacterial small RNA and eukaryotic microRNA. Exp. Biol. Med. 2017, 242, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, N.; Zhou, H.; Zhu, Y. The role of microbial metabolites in endocrine tumorigenesis: From the mechanistic insights to potential therapeutic biomarkers. Biomed. Pharmacother. 2024, 172, 116218. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.Y.; Park, H.T.; Dinh, N.T.H.; Choi, S.J.; Lee, J.; Kim, J.H.; Lee, S.-W.; Gho, Y.S. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat. Commun. 2017, 8, 626. [Google Scholar] [CrossRef]
- Li, D.; Zhu, L.; Wang, Y.; Zhou, X.; Li, Y. Bacterial outer membrane vesicles in cancer: Biogenesis, pathogenesis, and clinical application. Biomed. Pharmacother. 2023, 165, 115120. [Google Scholar] [CrossRef]
- Liu, C.; Yazdani, N.; Moran, C.S.; Salomon, C.; Seneviratne, C.J.; Ivanovski, S.; Han, P. Unveiling clinical applications of bacterial extracellular vesicles as natural nanomaterials in disease diagnosis and therapeutics. Acta Biomater. 2024, 180, 18–45. [Google Scholar] [CrossRef]
- Chronopoulos, A.; Kalluri, R. Emerging role of bacterial extracellular vesicles in cancer. Oncogene 2020, 39, 6951–6960. [Google Scholar] [CrossRef]
- Preet, R.; Islam, M.A.; Shim, J.; Rajendran, G.; Mitra, A.; Vishwakarma, V.; Kutz, C.; Choudhury, S.; Pathak, H.; Dai, Q. Gut commensal Bifidobacterium-derived extracellular vesicles modulate the therapeutic effects of anti-PD-1 in lung cancer. Nat. Commun. 2025, 16, 3500. [Google Scholar] [CrossRef]
- Hendrix, A.; De Wever, O. Systemically circulating bacterial extracellular vesicles: Origin, fate, and function. Trends Microbiol. 2022, 30, 213–216. [Google Scholar] [CrossRef]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef]
- Sangiorgio, G.; Nicitra, E.; Bivona, D.; Bonomo, C.; Bonacci, P.; Santagati, M.; Musso, N.; Bongiorno, D.; Stefani, S. Interactions of Gram-Positive Bacterial Membrane Vesicles and Hosts: Updates and Future Directions. Int. J. Mol. Sci. 2024, 25, 2904. [Google Scholar] [CrossRef] [PubMed]
- Peregrino, E.S.; Castañeda-Casimiro, J.; Vázquez-Flores, L.; Estrada-Parra, S.; Wong-Baeza, C.; Serafín-López, J.; Wong-Baeza, I. The Role of Bacterial Extracellular Vesicles in the Immune Response to Pathogens, and Therapeutic Opportunities. Int. J. Mol. Sci. 2024, 25, 6210. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Sun, F.; Zhao, B.; Kong, F.; Li, Z.; Kong, X. Bacteria derived extracellular vesicles in the pathogenesis and treatment of gastrointestinal tumours. Front. Oncol. 2023, 12, 1103446. [Google Scholar] [CrossRef] [PubMed]
- Grande, R.; Di Marcantonio, M.C.; Robuffo, I.; Pompilio, A.; Celia, C.; Di Marzio, L.; Paolino, D.; Codagnone, M.; Muraro, R.; Stoodley, P. Helicobacter pylori ATCC 43629/NCTC 11639 outer membrane vesicles (OMVs) from biofilm and planktonic phase associated with extracellular DNA (eDNA). Front. Microbiol. 2015, 6, 1369. [Google Scholar] [CrossRef]
- Kim, B.J.; McDonagh, M.A.; Deng, L.; Gastfriend, B.D.; Schubert-Unkmeir, A.; Doran, K.S.; Shusta, E.V. Streptococcus agalactiae disrupts P-glycoprotein function in brain endothelial cells. Fluids Barriers CNS 2019, 16, 26. [Google Scholar] [CrossRef]
- Le, L.H.M.; Steele, J.R.; Ying, L.; Schittenhelm, R.B.; Ferrero, R.L. A new isolation method for bacterial extracellular vesicles providing greater purity and improved proteomic detection of vesicle proteins. J. Extracell. Biol. 2023, 2, e84. [Google Scholar] [CrossRef]
- Guo, S.; Chen, J.; Chen, F.; Zeng, Q.; Liu, W.-L.; Zhang, G. Exosomes derived from Fusobacterium nucleatum-infected colorectal cancer cells facilitate tumour metastasis by selectively carrying miR-1246/92b-3p/27a-3p and CXCL16. Gut 2021, 70, 1507–1519. [Google Scholar] [CrossRef]
- Xiu, L.; Wu, Y.; Lin, G.; Zhang, Y.; Huang, L. Bacterial membrane vesicles: Orchestrators of interkingdom interactions in microbial communities for environmental adaptation and pathogenic dynamics. Front. Immunol. 2024, 15, 1371317. [Google Scholar] [CrossRef]
- Prince, A.; Tiwari, A.; Mandal, T.; Koiri, D.; Meher, G.; Sinha, D.K.; Saleem, M. Lipid specificity of the fusion of bacterial extracellular vesicles with the host membrane. J. Phys. Chem. B 2024, 128, 8116–8130. [Google Scholar] [CrossRef]
- Parris, B.A.; O’Farrell, H.E.; Fong, K.M.; Yang, I.A. Chronic obstructive pulmonary disease (COPD) and lung cancer: Common pathways for pathogenesis. J. Thorac. Dis. 2019, 11 (Suppl. 17), S2155. [Google Scholar] [CrossRef]
- Ma, B.; Barathan, M.; Ng, M.H.; Law, J.X. Oxidative Stress, Gut Microbiota, and Extracellular Vesicles: Interconnected Pathways and Therapeutic Potentials. Int. J. Mol. Sci. 2025, 26, 3148. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Giv, N.; Basas, A.; Hicks, C.; El-Omar, E.; El-Assaad, F.; Hosseini-Beheshti, E. Bacterial extracellular vesicles and their novel therapeutic applications in health and cancer. Front. Cell. Infect. Microbiol. 2022, 12, 962216. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, X.; Xiang, X.; Hao, C.; Ma, D. Roles of bacterial extracellular vesicles in systemic diseases. Front. Microbiol. 2023, 14, 1258860. [Google Scholar] [CrossRef] [PubMed]
- Tran, F.; Boedicker, J.Q. Genetic cargo and bacterial species set the rate of vesicle-mediated horizontal gene transfer. Sci. Rep. 2017, 7, 8813. [Google Scholar] [CrossRef] [PubMed]
- Werner Lass, S.; Smith, B.E.; Camphire, S.; Eutsey, R.A.; Prentice, J.A.; Yerneni, S.S.; Arun, A.; Bridges, A.A.; Rosch, J.W.; Conway, J.F. Pneumococcal extracellular vesicles mediate horizontal gene transfer via the transformation machinery. mSphere 2024, 9, e00727-24. [Google Scholar] [CrossRef]
- Verbunt, J.; Jocken, J.; Blaak, E.; Savelkoul, P.; Stassen, F. Gut-bacteria derived membrane vesicles and host metabolic health: A narrative review. Gut Microbes 2024, 16, 2359515. [Google Scholar] [CrossRef]
- Karaman, I.; Pathak, A.; Bayik, D.; Watson, D.C. Harnessing Bacterial Extracellular Vesicle Immune Effects for Cancer Therapy. Pathog. Immun. 2024, 9, 56. [Google Scholar] [CrossRef]
- Gao, X.; Yang, Y.; Liu, X.; Wang, Y.; Yang, Y.; Boireau, P.; Liu, M.; Bai, X. Extracellular vesicles derived from Trichinella spiralis prevent colitis by inhibiting M1 macrophage polarization. Acta Trop. 2021, 213, 105761. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Feng, Y.; Li, L.; Kong, F.; Gao, J.; Kong, X. Engineered bacterial outer membrane vesicles: A versatile bacteria-based weapon against gastrointestinal tumors. Theranostics 2024, 14, 761. [Google Scholar] [CrossRef]
- Briaud, P.; Carroll, R.K. Extracellular vesicle biogenesis and functions in gram-positive bacteria. Infect. Immun. 2020, 88, 10-1128. [Google Scholar] [CrossRef]
- Mozaheb, N.; Mingeot-Leclercq, M.-P. Membrane vesicle production as a bacterial defense against stress. Front. Microbiol. 2020, 11, 600221. [Google Scholar] [CrossRef] [PubMed]
- Wen, M.; Wang, J.; Ou, Z.; Nie, G.; Chen, Y.; Li, M.; Wu, Z.; Xiong, S.; Zhou, H.; Yang, Z. Bacterial extracellular vesicles: A position paper by the microbial vesicles task force of the Chinese society for extracellular vesicles. Interdiscip. Med. 2023, 1, e20230017. [Google Scholar] [CrossRef]
- Xue, Y.; Huang, X.; Ou, Z.; Wu, Y.; Li, Q.; Huang, X.; Wen, M.; Yang, Y.; Bo, S.; Zheng, L. Isolation and purification of bacterial extracellular vesicles from human feces using density gradient centrifugation. J. Vis. Exp. 2023, 199, e65574. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Arneth, B. Tumor microenvironment. Medicina 2019, 56, 15. [Google Scholar] [CrossRef]
- Elinav, E.; Nowarski, R.; Thaiss, C.A.; Hu, B.; Jin, C.; Flavell, R.A. Inflammation-induced cancer: Crosstalk between tumours, immune cells and microorganisms. Nat. Rev. Cancer 2013, 13, 759–771. [Google Scholar] [CrossRef]
- An, J.; Kim, J.B.; Yang, E.Y.; Kim, H.O.; Lee, W.-H.; Yang, J.; Kwon, H.; Paik, N.S.; Lim, W.; Kim, Y.-K. Bacterial extracellular vesicles affect endocrine therapy in MCF7 cells. Medicine 2021, 100, e25835. [Google Scholar] [CrossRef]
- Meganathan, V.; Moyana, R.; Natarajan, K.; Kujur, W.; Kusampudi, S.; Mulik, S.; Boggaram, V. Bacterial extracellular vesicles isolated from organic dust induce neutrophilic inflammation in the lung. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2020, 319, L893–L907. [Google Scholar] [CrossRef]
- Malla, R.R.; Kiran, P. Tumor microenvironment pathways: Cross regulation in breast cancer metastasis. Genes Dis. 2022, 9, 310–324. [Google Scholar] [CrossRef]
- Gorda, B.; Toelzer, C.; Aulicino, F.; Berger, I. The MultiBac BEVS: Basics, applications, performance and recent developments. Methods Enzymol. 2021, 660, 129–154. [Google Scholar]
- Tang, W.; Chen, Z.; Zhang, W.; Cheng, Y.; Zhang, B.; Wu, F.; Wang, Q.; Wang, S.; Rong, D.; Reiter, F. The mechanisms of sorafenib resistance in hepatocellular carcinoma: Theoretical basis and therapeutic aspects. Signal Transduct. Target. Ther. 2020, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Rudraprasad, D.; Nirmal, J.; Mishra, D.K.; Joseph, J. RNA-Sequencing Reveals the Modulation of the NLRP3 Inflammasome by miR-223-3p in Extracellular Vesicles in Bacterial Endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2025, 66, 53. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.; Mottawea, W.; Yeo, J.; Hammami, R. Gut microbiota extracellular vesicles as signaling molecules mediating host-microbiota communications. Int. J. Mol. Sci. 2021, 22, 13166. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Sun, Y.; Huang, Y.; Lian, J.; Wu, S.; Luo, D.; Gong, H. Fusobacterium nucleatum-derived small extracellular vesicles facilitate tumor growth and metastasis via TLR4 in breast cancer. BMC Cancer 2023, 23, 473. [Google Scholar] [CrossRef]
- Margutti, P.; D’Ambrosio, A.; Zamboni, S. Microbiota-Derived Extracellular Vesicle as Emerging Actors in Host Interactions. Int. J. Mol. Sci. 2024, 25, 8722. [Google Scholar] [CrossRef]
- Yu, S.; Zhao, Z.; Hao, P.; Qiu, Y.; Zhao, M.; Zhou, G.; Zhang, C.; Kang, J.; Li, P. Biological functions and cross-kingdom host gene regulation of small RNAs in Lactobacillus plantarum-derived extracellular vesicles. Front. Microbiol. 2022, 13, 944361. [Google Scholar] [CrossRef]
- Chaussé, A.-M.; Roche, S.M.; Moroldo, M.; Hennequet-Antier, C.; Holbert, S.; Kempf, F.; Barilleau, E.; Trotereau, J.; Velge, P. Epithelial cell invasion by Salmonella typhimurium induces modulation of genes controlled by aryl hydrocarbon receptor signaling and involved in extracellular matrix biogenesis. Virulence 2023, 14, 2158663. [Google Scholar] [CrossRef]
- Doré, E.; Boilard, E. Bacterial extracellular vesicles and their interplay with the immune system. Pharmacol. Ther. 2023, 247, 108443. [Google Scholar] [CrossRef]
- Vdovikova, S.; Gilfillan, S.; Wang, S.; Dongre, M.; Wai, S.N.; Hurtado, A. Modulation of gene transcription and epigenetics of colon carcinoma cells by bacterial membrane vesicles. Sci. Rep. 2018, 8, 7434. [Google Scholar] [CrossRef]
- Meng, R.; Zeng, M.; Ji, Y.; Huang, X.; Xu, M. The potential role of gut microbiota outer membrane vesicles in colorectal cancer. Front. Microbiol. 2023, 14, 1270158. [Google Scholar] [CrossRef]
- Choi, H.-I.; Choi, J.-P.; Seo, J.; Kim, B.J.; Rho, M.; Han, J.K.; Kim, J.G. Helicobacter pylori-derived extracellular vesicles increased in the gastric juices of gastric adenocarcinoma patients and induced inflammation mainly via specific targeting of gastric epithelial cells. Exp. Mol. Med. 2017, 49, e330. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.D.; Akbar, R.; Oliverio, A.; Thapa, K.; Wang, X.; Fan, G.-C. Bacterial Extracellular Vesicles in the Regulation of Inflammatory Response and Host-Microbe Interactions. Shock 2024, 61, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Johnston, E.L.; Heras, B.; Kufer, T.A.; Kaparakis-Liaskos, M. Detection of bacterial membrane vesicles by NOD-like receptors. Int. J. Mol. Sci. 2021, 22, 1005. [Google Scholar] [CrossRef] [PubMed]
- Kaparakis-Liaskos, M.; Ferrero, R.L. Immune modulation by bacterial outer membrane vesicles. Nat. Rev. Immunol. 2015, 15, 375–387. [Google Scholar] [CrossRef]
- Johnston, E.L.; Kufer, T.A.; Kaparakis-Liaskos, M. Immunodetection and pathogenesis mediated by bacterial membrane vesicles. In Bacterial Membrane Vesicles: Biogenesis, Functions and Applications; Springer: Berlin/Heidelberg, Germany, 2020; pp. 159–188. [Google Scholar]
- Amatya, S.B.; Salmi, S.; Kainulainen, V.; Karihtala, P.; Reunanen, J. Bacterial extracellular vesicles in gastrointestinal tract cancer: An unexplored territory. Cancers 2021, 13, 5450. [Google Scholar] [CrossRef]
- Zheng, X.; Gong, T.; Luo, W.; Hu, B.; Gao, J.; Li, Y.; Liu, R.; Xie, N.; Yang, W.; Xu, X. Fusobacterium nucleatum extracellular vesicles are enriched in colorectal cancer and facilitate bacterial adhesion. Sci. Adv. 2024, 10, eado0016. [Google Scholar] [CrossRef]
- Meng, X.; Ma, G.; Zhang, X.; Yin, H.; Miao, Y.; He, F. Extracellular vesicles from Fusobacterium nucleatum: Roles in the malignant phenotypes of gastric cancer. Cell Cycle 2024, 23, 294–307. [Google Scholar] [CrossRef]
- Gao, L.; Jiang, F. Streptococcus pneumoniae infection is associated with Matrix Metalloproteinase-9 in Lung Cancer Progression and Brain Metastases. medRxiv 2025. [Google Scholar] [CrossRef]
- Kaisanlahti, A.; Salmi, S.; Kumpula, S.; Amatya, S.B.; Turunen, J.; Tejesvi, M.; Byts, N.; Tapiainen, T.; Reunanen, J. Bacterial extracellular vesicles–brain invaders? A systematic review. Front. Mol. Neurosci. 2023, 16, 1227655. [Google Scholar] [CrossRef]
- Uemura, T.; Kawashima, A.; Jingushi, K.; Motooka, D.; Saito, T.; Nesrine, S.; Oka, T.; Okuda, Y.; Yamamoto, A.; Yamamichi, G. Bacteria-derived DNA in serum extracellular vesicles are biomarkers for renal cell carcinoma. Heliyon 2023, 9, e19800. [Google Scholar] [CrossRef]
- Kim, S.I.; Kang, N.; Leem, S.; Yang, J.; Jo, H.; Lee, M.; Kim, H.S.; Dhanasekaran, D.N.; Kim, Y.-K.; Park, T. Metagenomic analysis of serum microbe-derived extracellular vesicles and diagnostic models to differentiate ovarian cancer and benign ovarian tumor. Cancers 2020, 12, 1309. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.L.; Jin, J.X.; Zhu, J.; Ruan, H.B.; Huang, J.Q. Extracellular vesicles of Bifidobacterium longum reverse the acquired carboplatin resistance in ovarian cancer cells via p53 phosphorylation on Ser15. Kaohsiung J. Med. Sci. 2024, 40, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, A.; Sharma, D.; Kumar, A.; Singh, S.; Kumar, P. Microbiome and development of ovarian cancer. Endocr. Metab. Immune Disord.-Drug Targets (Former. Curr. Drug Targets-Immune Endocr. Metab. Disord.) 2022, 22, 1073–1090. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Xu, P. Global colorectal cancer burden in 2020 and projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef]
- Bostanghadiri, N.; Razavi, S.; Shariati, A.; Talebi, M.; Mirkalantari, S.; Emami Razavi, A.; Darban-Sarokhalil, D. Exploring the interplay between Fusobacterium nucleatum with the expression of microRNA, and inflammatory mediators in colorectal cancer. Front. Microbiol. 2023, 14, 1302719. [Google Scholar] [CrossRef]
- Wang, X.; Lin, S.; Wang, L.; Cao, Z.; Zhang, M.; Zhang, Y.; Liu, R.; Liu, J. Versatility of bacterial outer membrane vesicles in regulating intestinal homeostasis. Sci. Adv. 2023, 9, eade5079. [Google Scholar] [CrossRef]
- Galen, J.E.; Pham, T. Engineering New Immunotherapies Against Cancer Using Bacterial Outer Membrane Vesicles and Supported by Preclinical Data. Med. Res. Arch. 2022, 10. [Google Scholar] [CrossRef]
- Kim, D.J.; Yang, J.; Seo, H.; Lee, W.H.; Ho Lee, D.; Kym, S.; Park, Y.S.; Kim, J.G.; Jang, I.-J.; Kim, Y.-K. Colorectal cancer diagnostic model utilizing metagenomic and metabolomic data of stool microbial extracellular vesicles. Sci. Rep. 2020, 10, 2860. [Google Scholar] [CrossRef]
- Shirani, M.; Pakzad, R.; Haddadi, M.H.; Akrami, S.; Asadi, A.; Kazemian, H.; Moradi, M.; Kaviar, V.H.; Zomorodi, A.R.; Khoshnood, S. The global prevalence of gastric cancer in Helicobacter pylori-infected individuals: A systematic review and meta-analysis. BMC Infect. Dis. 2023, 23, 543. [Google Scholar] [CrossRef]
- Park, J.-Y.; Kang, C.-S.; Seo, H.-C.; Shin, J.-C.; Kym, S.-M.; Park, Y.-S.; Shin, T.-S.; Kim, J.-G.; Kim, Y.-K. Bacteria-derived extracellular vesicles in urine as a novel biomarker for gastric cancer: Integration of liquid biopsy and metagenome analysis. Cancers 2021, 13, 4687. [Google Scholar] [CrossRef]
- Łukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanisławek, A. Breast cancer—Epidemiology, risk factors, classification, prognostic markers, and current treatment strategies—An updated review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wan, S.; Zhou, H.; Du, J.; Li, Y.; Zhu, H.; Weng, L.; Ding, X.; Wang, L. Programmed Nanocloak of Commensal Bacteria-Derived Nanovesicles Amplify Strong Immunoreactivity Against Tumor Growth and Metastatic Progression. ACS Nano 2024, 18, 9613–9626. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.L.; Freire-de-Lima, C.G.; Nosanchuk, J.D.; Casadevall, A.; Rodrigues, M.L.; Nimrichter, L. Extracellular vesicles from Cryptococcus neoformans modulate macrophage functions. Infect. Immun. 2010, 78, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, J.Y.; Moon, S.; Lee, H.; Lee, S.; Kim, J.Y.; Kim, M.W.; Kim, S.I. Tumor-derived EV miRNA signatures surpass total EV miRNA in supplementing mammography for precision breast cancer diagnosis. Theranostics 2024, 14, 6587. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Guo, T.; Zhang, S.; Yang, M.; Cheng, J.; Wang, J.; Kang, J.; Ma, W.; Nian, Y.; Sun, Z. Bacteria-derived outer membrane vesicles engineered with over-expressed pre-miRNA as delivery nanocarriers for cancer therapy. Nanomed. Nanotechnol. Biol. Med. 2022, 45, 102585. [Google Scholar] [CrossRef]
- Liang, A.; Korani, L.; Yeung, C.L.S.; Tey, S.K.; Yam, J.W.P. The emerging role of bacterial extracellular vesicles in human cancers. J. Extracell. Vesicles 2024, 13, e12521. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, Z.; Xia, F.; Wen, Z.; Chen, R.; Zhu, D.; Wang, M.; Zhuge, X.; Dai, J. Reductions in bacterial viability stimulate the production of extra-intestinal pathogenic Escherichia coli (ExPEC) cytoplasm-carrying extracellular vesicles (EVs). PLoS Pathog. 2022, 18, e1010908. [Google Scholar] [CrossRef]
- Sanaei, M.-J.; Razi, S.; Pourbagheri-Sigaroodi, A.; Bashash, D. The PI3K/Akt/mTOR pathway in lung cancer; oncogenic alterations, therapeutic opportunities, challenges, and a glance at the application of nanoparticles. Transl. Oncol. 2022, 18, 101364. [Google Scholar] [CrossRef]
- Xiao, T.; Ma, Y.; Zhang, Z.; Zhang, Y.; Zhao, Y.; Zhou, X.; Wang, X.; Ge, K.; Guo, J.; Zhang, J. Tailoring therapeutics via a systematic beneficial elements comparison between photosynthetic bacteria-derived OMVs and extruded nanovesicles. Bioact. Mater. 2024, 36, 48–61. [Google Scholar] [CrossRef]
- Li, S.; Yue, H.; Wang, S.; Li, X.; Wang, X.; Guo, P.; Ma, G.; Wei, W. Advances of bacteria-based delivery systems for modulating tumor microenvironment. Adv. Drug Deliv. Rev. 2022, 188, 114444. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, D.; Li, S.; Ren, J.; Huang, W.; Liu, D.; Wang, W. Bronchoalveolar lavage fluid assessment facilitates precision medicine for lung cancer. Cancer Biol. Med. 2023, 21, 230. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Li, Y.; Yin, X.; Fan, Y.; Liu, J.; Wang, Y.; Liu, X.; Bai, G.; Li, K.; Shi, Y. Utilizing Engineered Bacteria as “Cell Factories” In Vivo for Intracellular RNA-Loaded Outer Membrane Vesicles’ Self-Assembly in Tumor Treatment. ACS Nano 2024, 18, 35296–35309. [Google Scholar] [CrossRef] [PubMed]
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Elashiry, M.; Carroll, A.; Yuan, J.; Liu, Y.; Hamrick, M.; Cutler, C.W.; Wang, Q.; Elsayed, R. Oral microbially-induced small extracellular vesicles cross the blood–brain barrier. Int. J. Mol. Sci. 2024, 25, 4509. [Google Scholar] [CrossRef]
- Grandi, A.; Tomasi, M.; Zanella, I.; Ganfini, L.; Caproni, E.; Fantappiè, L.; Irene, C.; Frattini, L.; Isaac, S.J.; König, E. Synergistic protective activity of tumor-specific epitopes engineered in bacterial outer membrane vesicles. Front. Oncol. 2017, 7, 253. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, H.; Wang, S.; Du, Y.; Wei, B.; Wu, Q.; Wang, H. Inhibitors of bacterial extracellular vesicles. Front. Microbiol. 2022, 13, 835058. [Google Scholar] [CrossRef]
- Chen, Q.; Zheng, Y.; Chen, X.; Xing, Y.; Zhang, J.; Yan, X.; Zhang, Q.; Wu, D.; Chen, Z. Bacteria Synergized with PD-1 Blockade Enhance Positive Feedback Loop of Cancer Cells-M1 Macrophages-T Cells in Glioma. Adv. Sci. 2024, 11, 2308124. [Google Scholar] [CrossRef]
- Sun, M.; Ma, J.; Zhang, G.; Song, M.; Lv, R.; Liang, J.; Shi, Y.; Zhao, L. Brain Targeting Bacterial Extracellular Vesicles Enhance Ischemic Stroke Therapy via Efficient ROS Elimination and Suppression of Immune Infiltration. ACS Nano 2025, 19, 15491–15508. [Google Scholar] [CrossRef]
- Gakis, G. The role of inflammation in bladder cancer. Inflamm. Cancer 2014, 816, 183–196. [Google Scholar]
- Blenkiron, C.; Simonov, D.; Muthukaruppan, A.; Tsai, P.; Dauros, P.; Green, S.; Hong, J.; Print, C.G.; Swift, S.; Phillips, A.R. Uropathogenic Escherichia coli releases extracellular vesicles that are associated with RNA. PLoS ONE 2016, 11, e0160440. [Google Scholar] [CrossRef]
- Wang, Z.; Jiang, Z.; Zhang, Y.; Wang, C.; Liu, Z.; Jia, Z.; Bhushan, S.; Yang, J.; Zhang, Z. Exosomes derived from bladder epithelial cells infected with uropathogenic Escherichia coli increase the severity of urinary tract infections (UTIs) by impairing macrophage function. PLoS Pathog. 2024, 20, e1011926. [Google Scholar] [CrossRef] [PubMed]
- Dauros-Singorenko, P.; Hong, J.; Swift, S.; Phillips, A.; Blenkiron, C. Effect of the extracellular vesicle RNA cargo from uropathogenic Escherichia coli on bladder cells. Front. Mol. Biosci. 2020, 7, 580913. [Google Scholar] [CrossRef] [PubMed]
- Jingushi, K.; Kawashima, A.; Tanikawa, S.; Saito, T.; Yamamoto, A.; Uemura, T.; Sassi, N.; Ishizuya, Y.; Yamamoto, Y.; Kato, T. Cutibacterium acnes-derived extracellular vesicles promote tumor growth in renal cell carcinoma. Cancer Sci. 2024, 115, 2578–2587. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, S.; Calin, G.A. Gut microbiota: A new player in regulating immune-and chemo-therapy efficacy. Cancer Drug Resist. 2020, 3, 356. [Google Scholar] [CrossRef]
- Ijaz, M.; Hasan, I.; Chaudhry, T.H.; Huang, R.; Zhang, L.; Hu, Z.; Tan, Q.; Guo, B. Bacterial derivatives mediated drug delivery in cancer therapy: A new generation strategy. J. Nanobiotechnol. 2024, 22, 510. [Google Scholar] [CrossRef]
- Matulonis, U.A.; Sood, A.K.; Fallowfield, L.; Howitt, B.E.; Sehouli, J.; Karlan, B.Y. Ovarian cancer. Nat. Rev. Dis. Primers 2016, 2, 16061. [Google Scholar] [CrossRef]
- Asano-Inami, E.; Yokoi, A.; Yoshida, K.; Taki, K.; Kitagawa, M.; Suzuki, K.; Uekusa, R.; Nagao, Y.; Yoshikawa, N.; Niimi, K. Proteomic profiling of bacterial extracellular vesicles for exploring ovarian cancer biomarkers. bioRxiv 2025. [Google Scholar] [CrossRef]
- Bamberger, E.; Perrett, C. Angiogenesis in epithelian ovarian cancer. Mol. Pathol. 2002, 55, 348. [Google Scholar] [CrossRef]
- Behzadi, E.; Hosseini, H.M.; Fooladi, A.A.I. The inhibitory impacts of Lactobacillus rhamnosus GG-derived extracellular vesicles on the growth of hepatic cancer cells. Microb. Pathog. 2017, 110, 1–6. [Google Scholar] [CrossRef]
- Zeng, Y.; Yin, Y.; Zhou, X. Insights into Microbiota–Host Crosstalk in the Intestinal Diseases Mediated by Extracellular Vesicles and Their Encapsulated MicroRNAs. Int. J. Mol. Sci. 2024, 25, 13001. [Google Scholar] [CrossRef]
- Yokoi, A.; Asano-Inami, E.; Yoshida, K.; Nagao, Y.; Kitagawa, M.; Kajiyama, H. Extracellular vesicles derived from vaginal bacteria initiate the progression of early phase ovarian carcinoma at fallopian tubes. Cancer Res. 2025, 85, 6517. [Google Scholar] [CrossRef]
- Yang, J.; Kim, E.K.; McDowell, A.; Kim, Y.-K. Microbe-derived extracellular vesicles as a smart drug delivery system. Transl. Clin. Pharmacol. 2018, 26, 103. [Google Scholar] [CrossRef] [PubMed]
- Bleich, R.M.; Arthur, J.C. Revealing a microbial carcinogen. Science 2019, 363, 689–690. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.R.; Jiang, Y.; Villalta, P.W.; Stornetta, A.; Boudreau, P.D.; Carrá, A.; Brennan, C.A.; Chun, E.; Ngo, L.; Samson, L.D. The human gut bacterial genotoxin colibactin alkylates DNA. Science 2019, 363, eaar7785. [Google Scholar] [CrossRef]
- Lai, H.; Guo, Y.; Tian, L.; Wu, L.; Li, X.; Yang, Z.; Chen, S.; Ren, Y.; He, S.; He, W. Protein panel of serum-derived small extracellular vesicles for the screening and diagnosis of epithelial ovarian cancer. Cancers 2022, 14, 3719. [Google Scholar] [CrossRef]
- Neettiyath, A.; Chung, K.; Liu, W.; Lee, L.P. Nanoplasmonic sensors for extracellular vesicles and bacterial membrane vesicles. Nano Converg. 2024, 11, 23. [Google Scholar] [CrossRef]
- Qin, Y.-F.; Lu, X.-Y.; Shi, Z.; Huang, Q.-S.; Wang, X.; Ren, B.; Cui, L. Deep learning-enabled Raman spectroscopic identification of pathogen-derived extracellular vesicles and the biogenesis process. Anal. Chem. 2022, 94, 12416–12426. [Google Scholar] [CrossRef]
- Su, K.-Y.; Koh Kok, J.-Y.; Chua, Y.-W.; Ong, S.-D.; Ser, H.L.; Pusparajah, P.; San Saw, P.; Goh, B.H.; Lee, W.-L. Bacterial extracellular vesicles in biofluids as potential diagnostic biomarkers. Expert Rev. Mol. Diagn. 2022, 22, 1057–1062. [Google Scholar] [CrossRef]
- Ghazi, B.; Harmak, Z.; Rghioui, M.; Kone, A.-S.; El Ghanmi, A.; Badou, A. Decoding the secret of extracellular vesicles in the immune tumor microenvironment of the glioblastoma: On the border of kingdoms. Front. Immunol. 2024, 15, 1423232. [Google Scholar] [CrossRef]
- Kuerban, K.; Gao, X.; Zhang, H.; Liu, J.; Dong, M.; Wu, L.; Ye, R.; Feng, M.; Ye, L. Doxorubicin-loaded bacterial outer-membrane vesicles exert enhanced anti-tumor efficacy in non-small-cell lung cancer. Acta Pharm. Sin. B 2020, 10, 1534–1548. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, Y.; Liu, S. KRAS mutation promotes the colonization of Fusobacterium nucleatum in colorectal cancer by down-regulating SERTAD4. J. Cell. Mol. Med. 2024, 28, e70182. [Google Scholar] [CrossRef] [PubMed]
- Di Filippo, L.D.; Duarte, J.L.; Luiz, M.T.; de Araújo, J.T.C.; Chorilli, M. Drug delivery nanosystems in glioblastoma multiforme treatment: Current state of the art. Curr. Neuropharmacol. 2021, 19, 787–812. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Zhao, R.; Li, Y.; Qi, Y.; Wang, Y.; Zhang, Y.; Qin, H.; Qin, Y.; Chen, L.; Li, C. Bioengineered bacteria-derived outer membrane vesicles as a versatile antigen display platform for tumor vaccination via Plug-and-Display technology. Nat. Commun. 2021, 12, 2041. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Sonar, S.; Kalele, K.; Subramaniyan, V. Bacterial surface derived extracellular vesicles: A ground-breaking approach in cancer therapy. Clin. Transl. Discov. 2024, 4, e351. [Google Scholar] [CrossRef]
- Mishra, S.; Amatya, S.B.; Salmi, S.; Koivukangas, V.; Karihtala, P.; Reunanen, J. Microbiota and extracellular vesicles in anti-PD-1/PD-L1 therapy. Cancers 2022, 14, 5121. [Google Scholar] [CrossRef]
- Hui, W.W.; Emerson, L.E.; Clapp, B.; Sheppe, A.E.; Sharma, J.; del Castillo, J.; Ou, M.; Maegawa, G.H.B.; Hoffman, C.; Larkin, I.J.; et al. Antigen-encapsulating host extracellular vesicles derived from Salmonella-infected cells stimulate pathogen-specific Th1-type responses in vivo. PLoS Pathog. 2021, 17, e1009465. [Google Scholar] [CrossRef]
- Hong, J.; Dauros-Singorenko, P.; Whitcombe, A.; Payne, L.; Blenkiron, C.; Phillips, A.; Swift, S. Analysis of the Escherichia coli extracellular vesicle proteome identifies markers of purity and culture conditions. J. Extracell. Vesicles 2019, 8, 1632099. [Google Scholar] [CrossRef]
- Melo, J.; Pinto, V.; Fernandes, T.; Malheiro, A.R.; Osório, H.; Figueiredo, C.; Leite, M. Isolation method and characterization of outer membranes vesicles of Helicobacter pylori grown in a chemically defined medium. Front. Microbiol. 2021, 12, 654193. [Google Scholar] [CrossRef]
- Du, X.; Ley, R.; Buck, A.H. MicroRNAs and extracellular vesicles in the gut: New host modulators of the microbiome? Microlife 2021, 2, uqab010. [Google Scholar] [CrossRef]
- Zhao, M.; Cheng, X.; Shao, P.; Dong, Y.; Wu, Y.; Xiao, L.; Cui, Z.; Sun, X.; Gao, C.; Chen, J. Bacterial protoplast-derived nanovesicles carrying CRISPR-Cas9 tools re-educate tumor-associated macrophages for enhanced cancer immunotherapy. Nat. Commun. 2024, 15, 950. [Google Scholar] [CrossRef]
- Liu, H.; Geng, Z.; Su, J. Engineered mammalian and bacterial extracellular vesicles as promising nanocarriers for targeted therapy. Extracell. Vesicles Circ. Nucleic Acids 2022, 3, 63–86. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Shi, M.; Liu, X.; Jin, C.; Xing, X.; Qiu, L.; Tan, W. Aptamer-functionalized exosomes: Elucidating the cellular uptake mechanism and the potential for cancer-targeted chemotherapy. Anal. Chem. 2019, 91, 2425–2430. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, H.; Han, Y.; Hu, Y.; Geng, Z.; Su, J. Bacterial extracellular vesicles-based therapeutic strategies for bone and soft tissue tumors therapy. Theranostics 2022, 12, 6576. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.Y.; Liu, S.; Xing, B. Bacteria extracellular vesicle as nanopharmaceuticals for versatile biomedical potential. Nano Converg. 2024, 11, 28. [Google Scholar] [CrossRef]
- Suri, K.; D’Souza, A.; Huang, D.; Bhavsar, A.; Amiji, M. Bacterial extracellular vesicle applications in cancer immunotherapy. Bioact. Mater. 2023, 22, 551–566. [Google Scholar] [CrossRef]
- Peng, L.-H.; Wang, M.-Z.; Chu, Y.; Zhang, L.; Niu, J.; Shao, H.-T.; Yuan, T.-J.; Jiang, Z.-H.; Gao, J.-Q.; Ning, X.-H. Engineering bacterial outer membrane vesicles as transdermal nanoplatforms for photo-TRAIL–programmed therapy against melanoma. Sci. Adv. 2020, 6, eaba2735. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Cargo | Function in Cancer | Relevant Cancers | References |
|---|---|---|---|
| Proteins | Regulate tumor progression, immune modulation, and EMT | Breast, Lung, Ovarian, Colorectal, Gastric, and Brain | [8,12,13,14,15,16,17] |
| Lipids | Mediate cell signaling and influence membrane dynamics | Lung, Ovarian, Breast, and Gastric | [15,18,19,20,21] |
| RNA | Alter gene expression and regulate immune evasion | Ovarian, Breast, Lung, Brain, and Colorectal | [12,16,20,22,23] |
| DNA | Promote horizontal gene transfer and affect tumor genetics | Colorectal and Gastric | [7,14,15,24,25] |
| Metabolites | Modulate metabolism and influence immune suppression | Ovarian, Lung, and Gastric | [14,20,26,27,28] |
| Enzymes | Carries degradative enzymes such as proteases and lipases | Breast, Colorectal, Gastric, and Lung | [5,15,28,29,30] |
| Cancer Type | Tumor Progression | Metastasis | Therapy Resistance | Key Bacterial Sources of BEVs | References |
|---|---|---|---|---|---|
| Colorectal Cancer | Promotes DNA damage and modulates immune responses | Enhances EMT and increases cell motility | BEVs transport drug efflux pumps and regulatory RNAs | Fusobacterium nucleatum and Escherichia coli | [13,56,57] |
| Gastric Cancer | Delivers virulence factors and induces oncogenic pathways | Remodels extracellular matrix and promotes angiogenesis | BEVs inhibit apoptosis and enhance drug efflux | Helicobacter pylori | [22,58] |
| Breast Cancer | Alters gene expression and promotes immune evasion | Facilitates EMT and tumor invasion | Modulates redox balance and activates survival pathways | Staphylococcus aureus and Fusobacterium nucleatum | [27,37,44] |
| Lung Cancer | Facilitates chronic inflammation and promotes oxidative stress | Forms pre-metastatic niches and increases migration | BEVs carry survival signals and downregulate immune responses | Pseudomonas aeruginosa and Streptococcus pneumoniae | [27,38,59] |
| Brain Cancer | Crosses the blood–brain barrier and activates oncogenic pathways | Induces angiogenesis and enhances invasiveness | Alters DNA repair mechanisms and promotes radioresistance | Escherichia coli and Streptococcus pneumoniae | [59,60] |
| Renal and Bladder Cancer | Triggers chronic inflammation and epithelial–mesenchymal transition (EMT) | Prepares metastatic niches and facilitates migration | Upregulates survival pathways and interferes with drug action | Escherichia coli (UPEC) | [22,61] |
| Ovarian Cancer | Induces inflammation, immune evasion, and promotes metastasis | Promotes peritoneal dissemination | Enhances chemoresistance and suppresses immune function | Lactobacillus spp. and Streptococcus spp. | [62,63,64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhanu, P.; Godwin, A.K.; Umar, S.; Mahoney, D.E. Bacterial Extracellular Vesicles in Oncology: Molecular Mechanisms and Future Clinical Applications. Cancers 2025, 17, 1774. https://doi.org/10.3390/cancers17111774
Bhanu P, Godwin AK, Umar S, Mahoney DE. Bacterial Extracellular Vesicles in Oncology: Molecular Mechanisms and Future Clinical Applications. Cancers. 2025; 17(11):1774. https://doi.org/10.3390/cancers17111774
Chicago/Turabian StyleBhanu, Piyush, Andrew K. Godwin, Shahid Umar, and Diane E. Mahoney. 2025. "Bacterial Extracellular Vesicles in Oncology: Molecular Mechanisms and Future Clinical Applications" Cancers 17, no. 11: 1774. https://doi.org/10.3390/cancers17111774
APA StyleBhanu, P., Godwin, A. K., Umar, S., & Mahoney, D. E. (2025). Bacterial Extracellular Vesicles in Oncology: Molecular Mechanisms and Future Clinical Applications. Cancers, 17(11), 1774. https://doi.org/10.3390/cancers17111774