
Hypoxia-Inducible Factor-Dependent and Independent Mechanisms Underlying Chemoresistance of Hypoxic Cancer Cells

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Brief Introduction
2. Hypoxic Microenvironment in Malignant Solid Tumors and Its Association with Therapy Resistance
3. HIF-Mediated Mechanisms behind Chemotherapy Resistance of Cancer Cells under Hypoxia
3.1. The Molecular Mechanisms behind the Regulation of HIFs’ Activity
3.2. HIF-Mediated Mechanisms behind Chemotherapy Resistance
3.3. HIF-Related Hypoxia-Responsive Non-Coding RNAs and Chemotherapy Resistance
3.4. HIF, EMT, and Chemotherapy Resistance
4. HIF-Independent Mechanisms behind Chemotherapy Resistance of Cancer Cells under Hypoxia
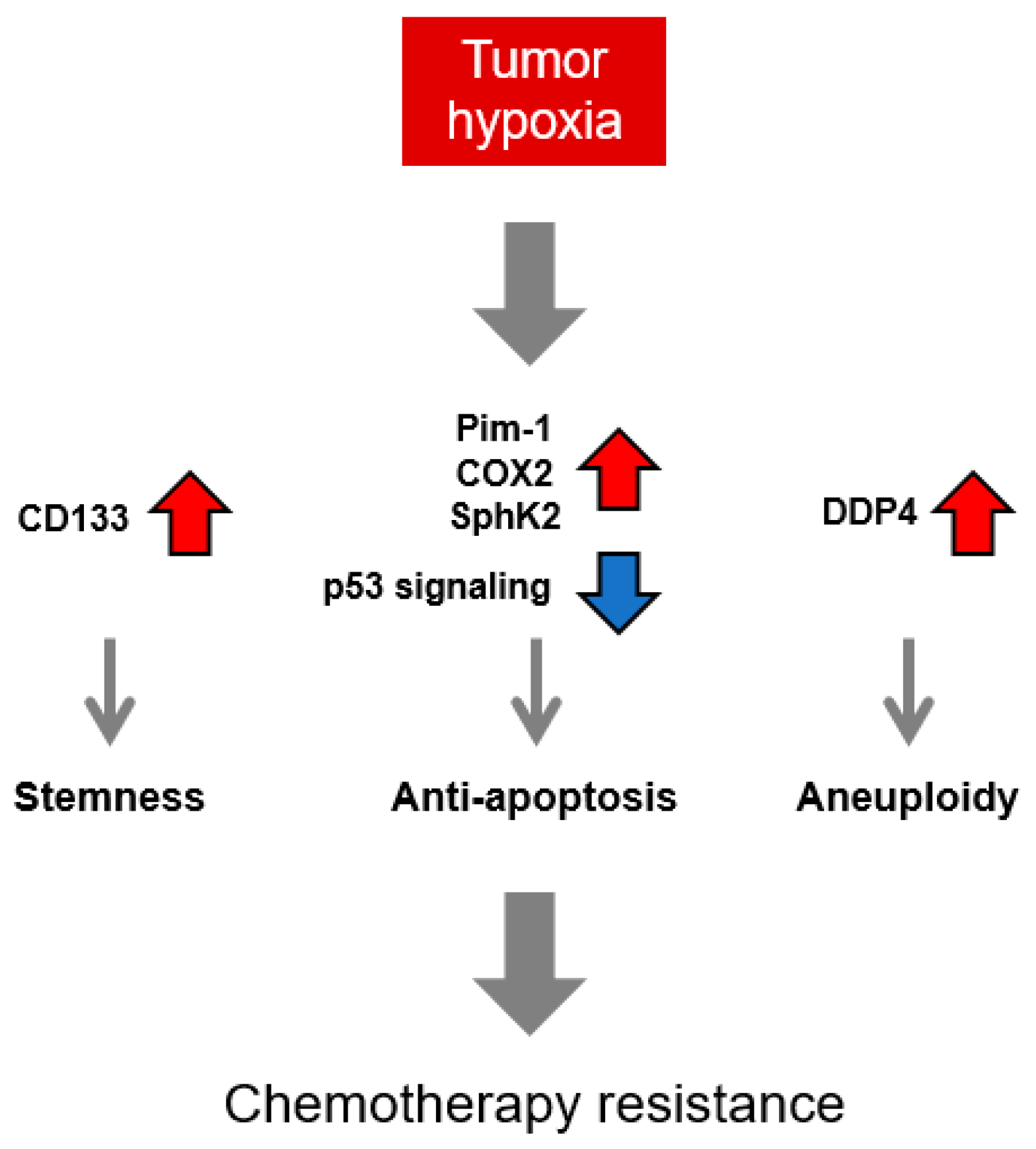
4.1. HIF-Independent Mechanisms behind Chemotherapy Resistance
4.2. Hypoxia-Associated Proteotoxicity, UPR, and Chemotherapy Resistance
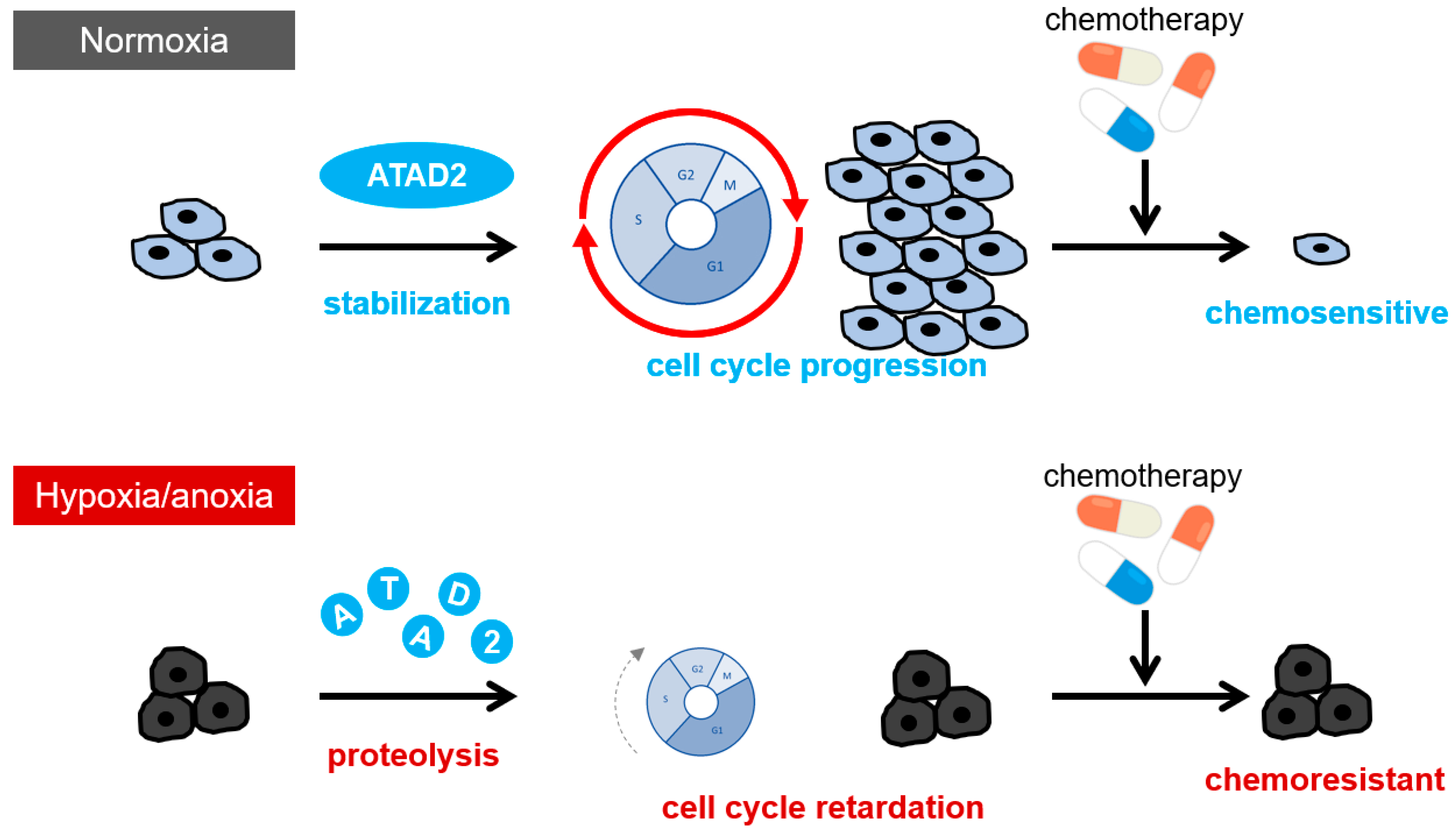
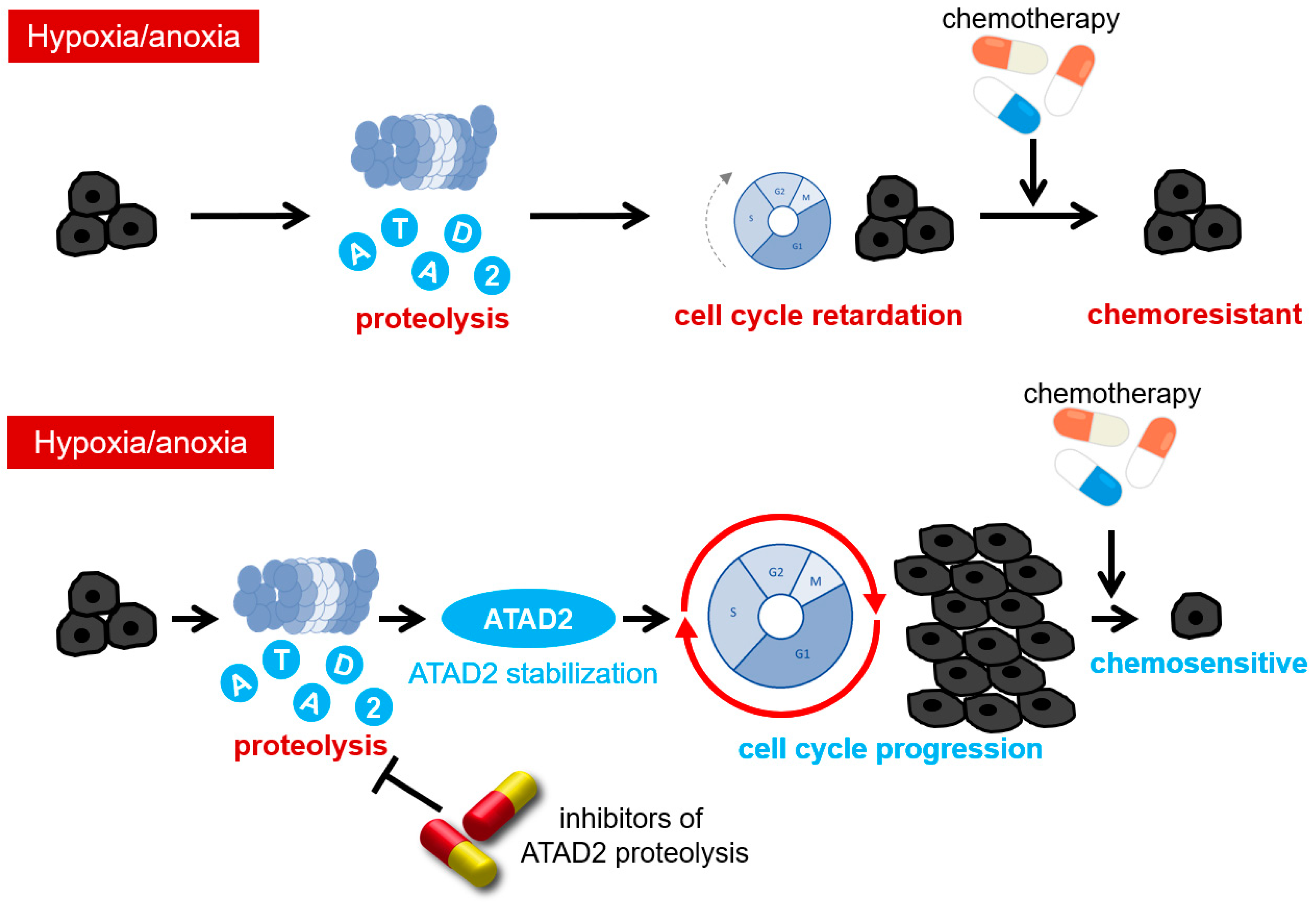
5. The Mechanisms of Therapy Resistance Acquisition through Hypoxia-Dependent Epigenetic Regulations: The Role of a Histone Acetyl Reader Protein, ATAD2
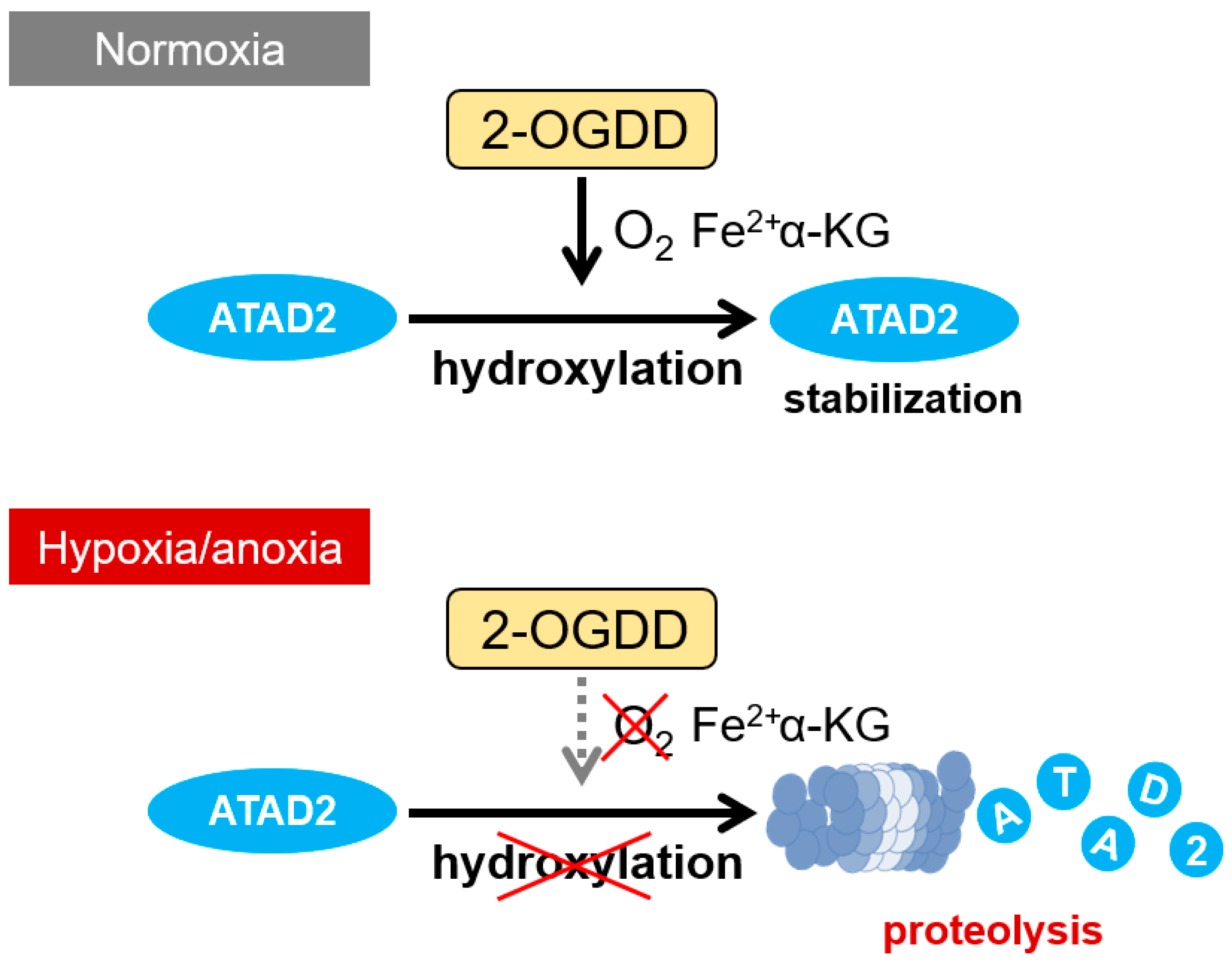
6. Oxygen-Dependent Regulatory Mechanism of ATAD2 Expression
7. Summary and Perspectives

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Target/Effect | Resistance | Cancer Type, Model | Ref. | |
|---|---|---|---|---|---|
| HIF downstream | LINC03000-201 (lncMat2B, ENST00000486913.3) | ↓ ROS production ↓ DNA damage | Cisplatin | Breast cancer, in vitro | [223] |
| PVT1 | miR-140-3p/ATG5 ↓ autophagy | Cisplatin | Lung cancer, in vitro and in vivo | [60] | |
| lncRNA-CBSLR | YTHDF2/CBS ↓ ferroptosis | Cisplatin | Gastric cancer, in vivo | [224] | |
| ANRIL | miR-328/ABCG2, MDR1 | Cisplatin | Retinoblastoma, in vitro | [225] | |
| LUCAT1 | Interaction with PTBP1 ↓ DNA damage | 5-Fluorouracil, Camptothecin, Doxorubicin and Oxaliplatin | Colorectal cancer, in vitro, in vivo, and patient cohorts | [226] | |
| NLUCAT1 (HIF-2α-dependent) | ↓ ROS production | Cisplatin | Lung adenocarcinoma, in vitro | [227] | |
| HIF upstream | PVT1 | miR-194-5p/HIF1A ↑ proliferation | Cisplatin | Oral SCC, in vitro | [228] |
| HIF1A-AS1 | Interaction with YB1 ↑ HIF-1α (positive feedback) ↑ glycolysis | Gemcitabine | Pancreatic cancer, in vitro, in vivo, and patient cohorts | [229] | |
| HIF1A-AS2 | ↑ HIF-1α ↑ autophagy | Doxorubicin | Small cell lung cancer, in vitro | [230] | |
| NORAD | miR-495-3p/HIF-1α ↑ vasculogenic mimicry | 5-Fluorouracil | Colorectal cancer, in vitro | [231] | |
| Potentially HIF-dependent | HOTAIR | miR-1277-5p/ZEB1 ↑ EMT | Oxaliplatin | Colorectal cancer, in vitro and in vivo | [232] |
| lncRNA-EMS | miR-758-3p/WTAP | Cisplatin | Esophageal cancer, in vitro and in vivo | [233] |
Author Contributions
Funding
Conflicts of Interest
References
- Yeom, C.J.; Goto, Y.; Zhu, Y.; Hiraoka, M.; Harada, H. Microenvironments and cellular characteristics in the micro tumor cords of malignant solid tumors. Int. J. Mol. Sci. 2012, 13, 13949–13965. [Google Scholar] [CrossRef]
- Walsh, J.C.; Lebedev, A.; Aten, E.; Madsen, K.; Marciano, L.; Kolb, H.C. The clinical importance of assessing tumor hypoxia: Relationship of tumor hypoxia to prognosis and therapeutic opportunities. Antioxid. Redox Signal. 2014, 21, 1516–1554. [Google Scholar] [CrossRef]
- Osinsky, S.; Zavelevich, M.; Vaupel, P. Tumor hypoxia and malignant progression. Exp. Oncol. 2009, 31, 80–86. [Google Scholar]
- Haitani, T.; Kobayashi, M.; Koyasu, S.; Akamatsu, S.; Suwa, T.; Onodera, Y.; Nam, J.-M.; Nguyen, P.T.L.; Menju, T.; Date, H.; et al. Proteolysis of a histone acetyl reader, atad2, induces chemoresistance of cancer cells under severe hypoxia by inhibiting cell cycle progression in s phase. Cancer Lett. 2022, 528, 76–84. [Google Scholar] [CrossRef]
- Shirai, Y.; Chow, C.C.T.; Kambe, G.; Suwa, T.; Kobayashi, M.; Takahashi, I.; Harada, H.; Nam, J.-M. An overview of the recent development of anticancer agents targeting the hif-1 transcription factor. Cancers 2021, 13, 2813. [Google Scholar] [CrossRef]
- Yoshimura, M.; Itasaka, S.; Harada, H.; Hiraoka, M. Microenvironment and radiation therapy. Biomed. Res. Int. 2013, 2013, 685308. [Google Scholar] [CrossRef]
- Mellor, H.R.; Callaghan, R. Resistance to chemotherapy in cancer: A complex and integrated cellular response. Pharmacology 2008, 81, 275–300. [Google Scholar] [CrossRef]
- Yeldag, G.; Rice, A.; Del Río Hernández, A. Chemoresistance and the self-maintaining tumor microenvironment. Cancers 2018, 10, 471. [Google Scholar] [CrossRef]
- Weniger, M.; Honselmann, K.C.; Liss, A.S. The extracellular matrix and pancreatic cancer: A complex relationship. Cancers 2018, 10, 316. [Google Scholar] [CrossRef] [PubMed]
- Masamune, A.; Kikuta, K.; Watanabe, T.; Satoh, K.; Hirota, M.; Shimosegawa, T. Hypoxia stimulates pancreatic stellate cells to induce fibrosis and angiogenesis in pancreatic cancer. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G709–G717. [Google Scholar] [CrossRef] [PubMed]
- Goda, N.; Ryan, H.E.; Khadivi, B.; McNulty, W.; Rickert, R.C.; Johnson, R.S. Hypoxia-inducible factor 1alpha is essential for cell cycle arrest during hypoxia. Mol. Cell. Biol. 2003, 23, 359–369. [Google Scholar] [CrossRef]
- Harada, H. How can we overcome tumor hypoxia in radiation therapy? J. Radiat. Res. 2011, 52, 545–556. [Google Scholar] [CrossRef]
- Kizaka-Kondoh, S.; Inoue, M.; Harada, H.; Hiraoka, M. Tumor hypoxia: A target for selective cancer therapy. Cancer Sci. 2003, 94, 1021–1028. [Google Scholar] [CrossRef]
- Kizaka-Kondoh, S.; Tanaka, S.; Harada, H.; Hiraoka, M. The hif-1-active microenvironment: An environmental target for cancer therapy. Adv. Drug Deliv. Rev. 2009, 61, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Wilson, W.R. Exploiting tumour hypoxia in cancer treatment. Nat. Rev. Cancer 2004, 4, 437–447. [Google Scholar] [CrossRef]
- Begg, K.; Tavassoli, M. Inside the hypoxic tumour: Reprogramming of the ddr and radioresistance. Cell Death Discov. 2020, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Semenza, G.L. Purification and characterization of hypoxia-inducible factor 1. J. Biol. Chem. 1995, 270, 1230–1237. [Google Scholar] [CrossRef]
- Hirota, K.; Semenza, G.L. Regulation of hypoxia-inducible factor 1 by prolyl and asparaginyl hydroxylases. Biochem. Biophys. Res. Commun. 2005, 338, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Epstein, A.C.; Gleadle, J.M.; McNeill, L.A.; Hewitson, K.S.; O’Rourke, J.; Mole, D.R.; Mukherji, M.; Metzen, E.; Wilson, M.I.; Dhanda, A.; et al. C. Elegans egl-9 and mammalian homologs define a family of dioxygenases that regulate hif by prolyl hydroxylation. Cell 2001, 107, 43–54. [Google Scholar] [CrossRef]
- Ivan, M.; Kondo, K.; Yang, H.; Kim, W.; Valiando, J.; Ohh, M.; Salic, A.; Asara, J.M.; Lane, W.S.; Kaelin, W.G., Jr. Hifalpha targeted for vhl-mediated destruction by proline hydroxylation: Implications for o2 sensing. Science 2001, 292, 464–468. [Google Scholar] [CrossRef]
- Jaakkola, P.; Mole, D.R.; Tian, Y.-M.; Wilson, M.I.; Gielbert, J.; Gaskell, S.J.; von Kriegsheim, A.; Hebestreit, H.F.; Mukherji, M.; Schofield, C.J.; et al. Targeting of hif-alpha to the von hippel-lindau ubiquitylation complex by o2-regulated prolyl hydroxylation. Science 2001, 292, 468–472. [Google Scholar] [CrossRef]
- Rosenberg, N.; Gervais, P. Evaluation of the sequelae of occupational asthma. Rev. Mal. Respir. 1989, 6, 35–38. [Google Scholar]
- Kaelin, W.G., Jr. The von hippel-lindau tumour suppressor protein: O2 sensing and cancer. Nat. Rev. Cancer 2008, 8, 865–873. [Google Scholar] [CrossRef]
- Tanimoto, K.; Makino, Y.; Pereira, T.; Poellinger, L. Mechanism of regulation of the hypoxia-inducible factor-1 alpha by the von hippel-lindau tumor suppressor protein. EMBO J. 2000, 19, 4298–4309. [Google Scholar] [CrossRef]
- Ohh, M.; Park, C.W.; Ivan, M.; Hoffman, M.A.; Kim, T.-Y.; Huang, L.E.; Pavletich, N.; Chau, V.; Kaelin, W.G. Ubiquitination of hypoxia-inducible factor requires direct binding to the beta-domain of the von hippel-lindau protein. Nat. Cell Biol. 2000, 2, 423–427. [Google Scholar] [CrossRef]
- Maxwell, P.H.; Wiesener, M.S.; Chang, G.-W.; Clifford, S.C.; Vaux, E.C.; Cockman, M.E.; Wykoff, C.C.; Pugh, C.W.; Maher, E.R.; Ratcliffe, P.J. The tumour suppressor protein vhl targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 1999, 399, 271–275. [Google Scholar] [CrossRef]
- Arany, Z.; Huang, L.E.; Eckner, R.; Bhattacharya, S.; Jiang, C.; Goldberg, M.A.; Bunn, H.F.; Livingston, D.M. An essential role for p300/cbp in the cellular response to hypoxia. Proc. Natl. Acad. Sci. USA 1996, 93, 12969–12973. [Google Scholar] [CrossRef]
- Lando, D.; Peet, D.J.; Whelan, D.A.; Gorman, J.J.; Whitelaw, M.L. Asparagine hydroxylation of the hif transactivation domain a hypoxic switch. Science 2002, 295, 858–861. [Google Scholar] [CrossRef]
- Mahon, P.C.; Hirota, K.; Semenza, G.L. Fih-1: A novel protein that interacts with hif-1alpha and vhl to mediate repression of hif-1 transcriptional activity. Genes Dev. 2001, 15, 2675–2686. [Google Scholar] [CrossRef]
- Koyasu, S.; Kobayashi, M.; Goto, Y.; Hiraoka, M.; Harada, H. Regulatory mechanisms of hypoxia-inducible factor 1 activity: Two decades of knowledge. Cancer Sci. 2018, 109, 560–571. [Google Scholar] [CrossRef]
- Yeom, C.J.; Zeng, L.; Goto, Y.; Morinibu, A.; Zhu, Y.; Shinomiya, K.; Kobayashi, M.; Itasaka, S.; Yoshimura, M.; Hur, C.-G.; et al. Ly6e: A conductor of malignant tumor growth through modulation of the pten/pi3k/akt/hif-1 axis. Oncotarget 2016, 7, 65837–65848. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Morinibu, A.; Kobayashi, M.; Zhu, Y.; Wang, X.; Goto, Y.; Yeom, C.J.; Zhao, T.; Hirota, K.; Shinomiya, K.; et al. Aberrant idh3alpha expression promotes malignant tumor growth by inducing hif-1-mediated metabolic reprogramming and angiogenesis. Oncogene 2015, 34, 4758–4766. [Google Scholar] [CrossRef]
- Harada, H.; Kizaka-Kondoh, S.; Li, G.; Itasaka, S.; Shibuya, K.; Inoue, M.; Hiraoka, M. Significance of hif-1-active cells in angiogenesis and radioresistance. Oncogene 2007, 26, 7508–7516. [Google Scholar] [CrossRef]
- Liu, Y.; Cox, S.R.; Morita, T.; Kourembanas, S. Hypoxia regulates vascular endothelial growth factor gene expression in endothelial cells. Identification of a 5′ enhancer. Circ. Res. 1995, 77, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Zeng, L.; Yeom, C.J.; Zhu, Y.; Morinibu, A.; Shinomiya, K.; Kobayashi, M.; Hirota, K.; Itasaka, S.; Yoshimura, M.; et al. Uchl1 provides diagnostic and antimetastatic strategies due to its deubiquitinating effect on hif-1alpha. Nat. Commun. 2015, 6, 6153. [Google Scholar] [CrossRef]
- Semenza, G.L. Molecular mechanisms mediating metastasis of hypoxic breast cancer cells. Trends Mol. Med. 2012, 18, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; De Marzo, A.M.; Laughner, E.; Lim, M.; Hilton, D.A.; Zagzag, D.; Buechler, P.; Isaacs, W.B.; Semenza, G.L.; Simons, J.W. Overexpression of hypoxia-inducible factor 1alpha in common human cancers and their metastases. Cancer Res. 1999, 59, 5830–5835. [Google Scholar]
- Semenza, G.L. Mitochondrial autophagy: Life and breath of the cell. Autophagy 2008, 4, 534–536. [Google Scholar] [CrossRef] [PubMed]
- Comerford, K.M.; Wallace, T.J.; Karhausen, J.; Louis, N.A.; Montalto, M.C.; Colgan, S.P. Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance (mdr1) gene. Cancer Res. 2002, 62, 3387–3394. [Google Scholar]
- He, M.; Wu, H.; Jiang, Q.; Liu, Y.; Han, L.; Yan, Y.; Wei, B.; Liu, F.; Deng, X.; Chen, H.; et al. Hypoxia-inducible factor-2alpha directly promotes bcrp expression and mediates the resistance of ovarian cancer stem cells to adriamycin. Mol. Oncol. 2019, 13, 403–421. [Google Scholar] [CrossRef]
- Pinzón-Daza, M.L.; Cuellar-Saenz, Y.; Nualart, F.; Ondo-Mendez, A.; Del Riesgo, L.; Castillo-Rivera, F.; Garzón, R. Oxidative stress promotes doxorubicin-induced pgp and bcrp expression in colon cancer cells under hypoxic conditions. J. Cell. Biochem. 2017, 118, 1868–1878. [Google Scholar] [CrossRef]
- Wang, H.; Wu, X.; Hudkins, K.; Mikheev, A.; Zhang, H.; Gupta, A.; Unadkat, J.D.; Mao, Q. Expression of the breast cancer resistance protein (bcrp1/abcg2) in tissues from pregnant mice: Effects of pregnancy and correlations with nuclear receptors. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E1295–E1304. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.-Y.; Chun, Y.-S.; Kim, T.-Y.; Kim, H.-L.; Kim, M.-S.; Park, J.-W. Hypoxia-inducible factor 1alpha-mediated resistance to phenolic anticancer. Chemotherapy 2004, 50, 119–126. [Google Scholar] [CrossRef]
- Chen, J.; Ding, Z.; Peng, Y.; Pan, F.; Li, J.; Zou, L.; Zhang, Y.; Liang, H. Hif-1alpha inhibition reverses multidrug resistance in colon cancer cells via downregulation of mdr1/p-glycoprotein. PLoS ONE 2014, 9, e98882. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-Y.; Fu, Z.-X.; Wang, X.-H. Effect of hypoxia-inducible factor 1-alpha on survivin in colorectal cancer. Mol. Med. Rep. 2010, 3, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Dong, X.; Lin, L.; Jiang, X.; Wei, Z.; Zhai, B.; Sun, B.; Zhang, Q.; Wang, X.; Jiang, H.; et al. Up-regulation of survivin by akt and hypoxia-inducible factor 1alpha contributes to cisplatin resistance in gastric cancer. FEBS J. 2014, 281, 115–128. [Google Scholar] [CrossRef]
- Li, H.; Sun, X.; Li, J.; Liu, W.; Pan, G.; Mao, A.; Liu, J.; Zhang, Q.; Rao, L.; Xie, X.; et al. Hypoxia induces docetaxel resistance in triple-negative breast cancer via the hif-1alpha/mir-494/survivin signaling pathway. Neoplasia 2022, 32, 100821. [Google Scholar] [CrossRef] [PubMed]
- Zichittella, C.; Barreca, M.M.; Cordaro, A.; Corrado, C.; Alessandro, R.; Conigliaro, A. Mir-675-5p supports hypoxia-induced drug resistance in colorectal cancer cells. BMC Cancer 2022, 22, 567. [Google Scholar] [CrossRef]
- Feng, L.; Shen, F.; Zhou, J.; Li, Y.; Jiang, R.; Chen, Y. Hypoxia-induced up-regulation of mir-27a promotes paclitaxel resistance in ovarian cancer. Biosci. Rep. 2020, 40, BSR20192457. [Google Scholar] [CrossRef]
- Xu, K.; Zhan, Y.; Yuan, Z.; Qiu, Y.; Wang, H.; Fan, G.; Wang, J.; Li, W.; Cao, Y.; Shen, X.; et al. Hypoxia induces drug resistance in colorectal cancer through the hif-1alpha/mir-338-5p/il-6 feedback loop. Mol. Ther. 2019, 27, 1810–1824. [Google Scholar] [CrossRef]
- Lu, H.; Samanta, D.; Xiang, L.; Zhang, H.; Hu, H.; Chen, I.; Bullen, J.W.; Semenza, G.L. Chemotherapy triggers hif-1-dependent glutathione synthesis and copper chelation that induces the breast cancer stem cell phenotype. Proc. Natl. Acad. Sci. USA 2015, 112, E4600–E4609. [Google Scholar] [CrossRef] [PubMed]
- Roncuzzi, L.; Pancotti, F.; Baldini, N. Involvement of hif-1alpha activation in the doxorubicin resistance of human osteosarcoma cells. Oncol. Rep. 2014, 32, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Ding, J.; Sun, W.; Wu, K.; Ning, B.; Gong, W.; He, G.; Huang, S.; Ding, X.; Yin, P.; et al. Suppression of cyclin d1 by hypoxia-inducible factor-1 via direct mechanism inhibits the proliferation and 5-fluorouracil-induced apoptosis of a549 cells. Cancer Res. 2010, 70, 2010–2019. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.; Paré, G.C.; Frederiksen, L.J.; Semenza, G.L.; Graham, C.H. Hypoxia-induced resistance to anticancer drugs is associated with decreased senescence and requires hypoxia-inducible factor-1 activity. Mol. Cancer Ther. 2008, 7, 1961–1973. [Google Scholar] [CrossRef]
- Johnson, A.B.; Denko, N.; Barton, M.C. Hypoxia induces a novel signature of chromatin modifications and global repression of transcription. Mutat. Res. 2008, 640, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-F.; Luo, D.; Li, X.M.; Li, Z.-Q.; Yu, X.; Zhu, H.-W. Pvt1 knockdown inhibits autophagy and improves gemcitabine sensitivity by regulating the mir-143/hif-1alpha/vmp1 axis in pancreatic cancer. Pancreas 2021, 50, 227–234. [Google Scholar] [CrossRef]
- Wu, H.-M.; Jiang, Z.-F.; Ding, P.-S.; Shao, L.-J.; Liu, R.-Y. Hypoxia-induced autophagy mediates cisplatin resistance in lung cancer cells. Sci. Rep. 2015, 5, 12291. [Google Scholar] [CrossRef]
- Liang, C.; Dong, Z.; Cai, X.; Shen, J.; Xu, Y.; Zhang, M.; Li, H.; Yu, W.; Chen, W. Hypoxia induces sorafenib resistance mediated by autophagy via activating foxo3a in hepatocellular carcinoma. Cell Death Dis. 2020, 11, 1017. [Google Scholar] [CrossRef]
- Yang, X.; Yin, H.; Zhang, Y.; Li, X.; Tong, H.; Zeng, Y.; Wang, Q.; He, W. Hypoxia-induced autophagy promotes gemcitabine resistance in human bladder cancer cells through hypoxia-inducible factor 1alpha activation. Int. J. Oncol. 2018, 53, 215–224. [Google Scholar] [CrossRef]
- Wang, J.; Dong, Z.; Sheng, Z.; Cai, Y. Hypoxia-induced pvt1 promotes lung cancer chemoresistance to cisplatin by autophagy via pvt1/mir-140-3p/atg5 axis. Cell Death Discov. 2022, 8, 104. [Google Scholar] [CrossRef]
- Huang, S.; Qi, P.; Zhang, T.; Li, F.; He, X. The hif-1alpha/mir-224-3p/atg5 axis affects cell mobility and chemosensitivity by regulating hypoxia-induced protective autophagy in glioblastoma and astrocytoma. Oncol. Rep. 2019, 41, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Weng, L.; Jia, Y.; Liu, B.; Wu, S.; Xue, L.; Yin, X.; Mao, A.; Wang, Z.; Shang, M. Ptbp3 promotes malignancy and hypoxia-induced chemoresistance in pancreatic cancer cells by atg12 up-regulation. J. Cell. Mol. Med. 2020, 24, 2917–2930. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Dominguez, R.; Perez-Medina, M.; Lopez-Gonzalez, J.S.; Galicia-Velasco, M.; Aguilar-Cazares, D. The double-edge sword of autophagy in cancer: From tumor suppression to pro-tumor activity. Front. Oncol. 2020, 10, 578418. [Google Scholar] [CrossRef]
- Sannigrahi, M.; Singh, V.; Sharma, R.; Panda, N.; Khullar, M. Role of autophagy in head and neck cancer and therapeutic resistance. Oral Dis. 2015, 21, 283–291. [Google Scholar] [CrossRef]
- Sullivan, R.; Graham, C.H. Hypoxia prevents etoposide-induced DNA damage in cancer cells through a mechanism involving hypoxia-inducible factor 1. Mol. Cancer Ther. 2009, 8, 1702–1713. [Google Scholar] [CrossRef]
- Yan, Y.; He, M.; Zhao, L.; Wu, H.; Zhao, Y.; Han, L.; Wei, B.; Ye, D.; Lv, X.; Wang, Y.; et al. A novel hif-2alpha targeted inhibitor suppresses hypoxia-induced breast cancer stemness via sod2-mtros-pdi/gpr78-upr(er) axis. Cell Death Differ. 2022, 29, 1769–1789. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, F.; Han, L.; Zhao, L.; Chen, J.; I Olopade, O.; He, M.; Wei, M. Hif-2alpha promotes conversion to a stem cell phenotype and induces chemoresistance in breast cancer cells by activating wnt and notch pathways. J. Exp. Clin. Cancer Res. 2018, 37, 256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Tao, X.; Ji, B.; Gong, J. Hypoxia-driven m2-polarized macrophages facilitate cancer aggressiveness and temozolomide resistance in glioblastoma. Oxidative Med. Cell. Longev. 2022, 2022, 1614336. [Google Scholar] [CrossRef]
- Zheng, H.; Yu, S.; Zhu, C.; Guo, T.; Liu, F.; Xu, Y. Hif1alpha promotes tumor chemoresistance via recruiting gdf15-producing tams in colorectal cancer. Exp. Cell Res. 2021, 398, 112394. [Google Scholar] [CrossRef]
- Yu, S.; Li, Q.; Yu, Y.; Cui, Y.; Li, W.; Liu, T.; Liu, F. Activated hif1alpha of tumor cells promotes chemoresistance development via recruiting gdf15-producing tumor-associated macrophages in gastric cancer. Cancer Immunol. Immunother. 2020, 69, 1973–1987. [Google Scholar] [CrossRef]
- Eptaminitaki, G.C.; Stellas, D.; Bonavida, B.; Baritaki, S. Long non-coding rnas (lncrnas) signaling in cancer chemoresistance: From prediction to druggability. Drug Resist. Updat. 2022, 65, 100866. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-Y.; Zhang, Q.; Guo, J.; Zhang, P.; Liu, H.; Tian, Z.-B.; Zhang, C.-P.; Li, X.-Y. The role of circular rnas in the drug resistance of cancers. Front. Oncol. 2021, 11, 790589. [Google Scholar] [CrossRef] [PubMed]
- Papatsirou, M.; I Artemaki, P.; Scorilas, A.; Kontos, C.K. The role of circular rnas in therapy resistance of patients with solid tumors. Pers. Med. 2020, 17, 469–490. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Tan, H.-Y.; Chan, Y.-T.; Jiang, H.; Wang, N.; Wang, D. The functional role of long noncoding rna in resistance to anticancer treatment. Ther. Adv. Med. Oncol. 2020, 12, 1758835920927850. [Google Scholar] [CrossRef]
- Cui, C.; Yang, J.; Li, X.; Liu, D.; Fu, L.; Wang, X. Functions and mechanisms of circular rnas in cancer radiotherapy and chemotherapy resistance. Mol. Cancer 2020, 19, 58. [Google Scholar] [CrossRef] [PubMed]
- Pucci, P.; Rescigno, P.; Sumanasuriya, S.; de Bono, J.; Crea, F. Hypoxia and noncoding rnas in taxane resistance. Trends Pharmacol. Sci. 2018, 39, 695–709. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, A.; Bakhshaei Shahrebabaki, P.; Fouladi, H.; Mansoori Derakhshan, S. The impact of micrornas on the resistance of breast cancer subtypes to chemotherapy. Pathol. Res. Pract. 2023, 249, 154702. [Google Scholar] [CrossRef] [PubMed]
- Funamizu, N.; Honjo, M.; Tamura, K.; Sakamoto, K.; Ogawa, K.; Takada, Y. Micrornas associated with gemcitabine resistance via emt, tme, and drug metabolism in pancreatic cancer. Cancers 2023, 15, 1230. [Google Scholar] [CrossRef]
- Doghish, A.S.; Ismail, A.; Elrebehy, M.A.; Elbadry, A.M.; Mahmoud, H.H.; Farouk, S.M.; Abu Serea, G.A.; Elghany, R.A.A.; El-Halwany, K.K.; Alsawah, A.O.; et al. A study of mirnas as cornerstone in lung cancer pathogenesis and therapeutic resistance: A focus on signaling pathways interplay. Pathol. Res. Pract. 2022, 237, 154053. [Google Scholar] [CrossRef]
- Karami Fath, M.; Azargoonjahromi, A.; Kiani, A.; Jalalifar, F.; Osati, P.; Akbari Oryani, M.; Shakeri, F.; Nasirzadeh, F.; Khalesi, B.; Nabi-Afjadi, M.; et al. The role of epigenetic modifications in drug resistance and treatment of breast cancer. Cell. Mol. Biol. Lett. 2022, 27, 52. [Google Scholar] [CrossRef]
- Zeng, Z.; Chen, Y.; Geng, X.; Zhang, Y.; Wen, X.; Yan, Q.; Wang, T.; Ling, C.; Xu, Y.; Duan, J.; et al. Ncrnas: Multi-angle participation in the regulation of glioma chemotherapy resistance (review). Int. J. Oncol. 2022, 60, 76. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Hashemi, F.; Moghadam, E.R.; Owrang, M.; Hashemi, F.; Makvandi, P.; Goharrizi, M.A.S.B.; Najafi, M.; et al. Lung cancer cells and their sensitivity/resistance to cisplatin chemotherapy: Role of micrornas and upstream mediators. Cell. Signal. 2021, 78, 109871. [Google Scholar] [CrossRef]
- Lin, Z.; Lu, S.; Xie, X.; Yi, X.; Huang, H. Noncoding rnas in drug-resistant pancreatic cancer: A review. Biomed. Pharmacother. 2020, 131, 110768. [Google Scholar] [CrossRef]
- Yete, S.; Saranath, D. Micrornas in oral cancer: Biomarkers with clinical potential. Oral Oncol. 2020, 110, 105002. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Hashemi, F.; Hashemi, F.; Samarghandian, S.; Najafi, M. Micrornas in cancer therapy: Their involvement in oxaliplatin sensitivity/resistance of cancer cells with a focus on colorectal cancer. Life Sci. 2020, 256, 117973. [Google Scholar] [CrossRef]
- Si, W.; Shen, J.; Zheng, H.; Fan, W. The role and mechanisms of action of micrornas in cancer drug resistance. Clin. Epigenetics 2019, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Cano, I.; Pattanayak, B.; Adam-Artigues, A.; Lameirinhas, A.; Torres-Ruiz, S.; Tormo, E.; Cervera, R.; Eroles, P. Micrornas as a clue to overcome breast cancer treatment resistance. Cancer Metastasis Rev. 2022, 41, 77–105. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Li, S.; Zhu, S.; Yi, M.; Luo, S.; Wu, K. Mirna-mediated emt and cscs in cancer chemoresistance. Exp. Hematol. Oncol. 2021, 10, 12. [Google Scholar] [CrossRef]
- Pan, G.; Liu, Y.; Shang, L.; Zhou, F.; Yang, S. Emt-associated micrornas and their roles in cancer stemness and drug resistance. Cancer Commun. 2021, 41, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Liang, W.; Liu, J.; Zhang, L.; Wei, J.; Yang, J.; Zhang, Y.; Huang, Z. Autophagy-mediating micrornas in cancer chemoresistance. Cell Biol. Toxicol. 2020, 36, 517–536. [Google Scholar] [CrossRef]
- Zeng, X.; Wang, H.-Y.; Bai, S.-Y.; Pu, K.; Wang, Y.-P.; Zhou, Y.-N. The roles of micrornas in multidrug-resistance mechanisms in gastric cancer. Curr. Mol. Med. 2020, 20, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, C.; Zeng, J.; Zhao, Z.; Hu, Q. Microrna-210-3p is transcriptionally upregulated by hypoxia induction and thus promoting emt and chemoresistance in glioma cells. PLoS ONE 2021, 16, e0253522. [Google Scholar] [CrossRef] [PubMed]
- Jawad, S.F.; Altalbawy, F.M.A.; Hussein, R.M.; Fadhil, A.A.; Jawad, M.A.; Zabibah, R.S.; Taraki, T.Y.; Mohan, C.D.; Rangappa, K.S. The strict regulation of hif-1alpha by non-coding rnas: New insight towards proliferation, metastasis, and therapeutic resistance strategies. Cancer Metastasis Rev. 2024, 43, 5–27. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Sheng, L.; Qu, W.; Xue, X.; Chen, H.; Zheng, G.; Chen, W. Mir-194-5p enhances the sensitivity of nonsmall-cell lung cancer to doxorubicin through targeted inhibition of hypoxia-inducible factor-1. World J. Surg. Oncol. 2021, 19, 174. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Xia, X.; Wang, X.; Zhang, K.; Cao, J.; Jiang, T.; Zhao, Q.; Qiu, Z. Mir-301a plays a pivotal role in hypoxia-induced gemcitabine resistance in pancreatic cancer. Exp. Cell Res. 2018, 369, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Jasim, S.A.; Majeed, A.A.; Uinarni, H.; Alshuhri, M.; Alzahrani, A.A.; Ibrahim, A.A.; Alawadi, A.; Abed Al-Abadi, N.K.; Mustafa, Y.F.; Ahmed, B.A. Long non-coding rna (lncrna) pvt1 in drug resistance of cancers: Focus on pathological mechanisms. Pathol. Res. Pract. 2024, 254, 155119. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, M.; Deng, Y. Long noncoding rnas in taxane resistance of breast cancer. Int. J. Mol. Sci. 2023, 24, 12253. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Zhang, G.; Wang, X.; Wang, Y.; Wang, K. Functions and mechanisms of lncrna malat1 in cancer chemotherapy resistance. Biomark. Res. 2023, 11, 23. [Google Scholar] [CrossRef]
- Zhu, C.; Wang, X.; Wang, Y.; Wang, K. Functions and underlying mechanisms of lncrna hotair in cancer chemotherapy resistance. Cell Death Discov. 2022, 8, 383. [Google Scholar] [CrossRef]
- Yao, W.; Li, S.; Liu, R.; Jiang, M.; Gao, L.; Lu, Y.; Liang, X.; Zhang, H. Long non-coding rna pvt1: A promising chemotherapy and radiotherapy sensitizer. Front. Oncol. 2022, 12, 959208. [Google Scholar] [CrossRef]
- Ashrafizaveh, S.; Ashrafizadeh, M.; Zarrabi, A.; Husmandi, K.; Zabolian, A.; Shahinozzaman; Aref, A.R.; Hamblin, M.R.; Nabavi, N.; Crea, F.; et al. Long non-coding rnas in the doxorubicin resistance of cancer cells. Cancer Lett. 2021, 508, 104–114. [Google Scholar] [CrossRef]
- Taheri, M.; Shoorei, H.; Anamag, F.T.; Ghafouri-Fard, S.; Dinger, M.E. Lncrnas and mirnas participate in determination of sensitivity of cancer cells to cisplatin. Exp. Mol. Pathol. 2021, 123, 104602. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Guan, Z.; He, K.; Qian, J.; Cao, J.; Teng, L. Lncrna uca1 in anti-cancer drug resistance. Oncotarget 2017, 8, 64638–64650. [Google Scholar] [CrossRef]
- Jiao, B.; Liu, S.; Zhao, H.; Zhuang, Y.; Ma, S.; Lin, C.; Hu, J.; Liu, X. Hypoxia-responsive circrnas: A novel but important participant in non-coding rnas ushered toward tumor hypoxia. Cell Death Dis. 2022, 13, 666. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.; Cao, G.; Hua, J.; Shan, G.; Lin, W. Emerging roles of circular rnas in gastric cancer metastasis and drug resistance. J. Exp. Clin. Cancer Res. 2022, 41, 218. [Google Scholar] [CrossRef]
- Mu, Q.; Lv, Y.; Luo, C.; Liu, X.; Huang, C.; Xiu, Y.; Tang, L. Research progress on the functions and mechanism of circrna in cisplatin resistance in tumors. Front. Pharmacol. 2021, 12, 709324. [Google Scholar] [CrossRef]
- Ameli-Mojarad, M.; Ameli-Mojarad, M.; Hadizadeh, M.; Young, C.; Babini, H.; Nazemalhosseini-Mojarad, E.; Bonab, M.A. The effective function of circular rna in colorectal cancer. Cancer Cell Int. 2021, 21, 496. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Li, M.; Wu, J.; Qin, C.; Tao, Y.; He, H. Circular rna circnrip1 sponges microrna-138-5p to maintain hypoxia-induced resistance to 5-fluorouracil through hif-1alpha-dependent glucose metabolism in gastric carcinoma. Cancer Manag. Res. 2020, 12, 2789–2802. [Google Scholar] [CrossRef]
- Xu, M.; Liu, X.; Zhou, X.; Qin, Y.; Yang, L.; Wen, S.; Qiu, Y.; Chen, S.; Tang, R.; Guo, Y.; et al. Hypoxia-induced circstt3a enhances serine synthesis and promotes h3k4me3 modification to facilitate breast cancer stem cell formation. Pharmacol. Res. 2023, 197, 106964. [Google Scholar] [CrossRef]
- Zeng, Z.; Zhao, Y.; Chen, Q.; Zhu, S.; Niu, Y.; Ye, Z.; Hu, P.; Chen, D.; Xu, P.; Chen, J.; et al. Hypoxic exosomal hif-1alpha-stabilizing circznf91 promotes chemoresistance of normoxic pancreatic cancer cells via enhancing glycolysis. Oncogene 2021, 40, 5505–5517. [Google Scholar] [CrossRef]
- Deng, K.; Zou, F.; Xu, J.; Xu, D.; Luo, Z. Cancer-associated fibroblasts promote stemness maintenance and gemcitabine resistance via hif-1alpha/mir-21 axis under hypoxic conditions in pancreatic cancer. Mol. Carcinog. 2024, 63, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Tam, S.Y.; Wu, V.W.C.; Law, H.K.W. Hypoxia-induced epithelial-mesenchymal transition in cancers: Hif-1alpha and beyond. Front. Oncol. 2020, 10, 486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lou, Y.; Zhang, J.; Fu, Q.; Wei, T.; Sun, X.; Chen, Q.; Yang, J.; Bai, X.; Liang, T. Hypoxia-inducible factor-2alpha promotes tumor progression and has crosstalk with wnt/beta-catenin signaling in pancreatic cancer. Mol. Cancer 2017, 16, 119. [Google Scholar] [CrossRef] [PubMed]
- Kohnoh, T.; Hashimoto, N.; Ando, A.; Sakamoto, K.; Miyazaki, S.; Aoyama, D.; Kusunose, M.; Kimura, M.; Omote, N.; Imaizumi, K.; et al. Hypoxia-induced modulation of pten activity and emt phenotypes in lung cancers. Cancer Cell Int. 2016, 16, 33. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Tang, Y.-L.; Liang, X.-H. Emt: A new vision of hypoxia promoting cancer progression. Cancer Biol. Ther. 2011, 11, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, S.; Schuhwerk, H.; Brabletz, T.; Stemmler, M.P. Dynamic emt: A multi-tool for tumor progression. EMBO J. 2021, 40, e108647. [Google Scholar] [CrossRef] [PubMed]
- Gaponova, A.V.; Rodin, S.; Mazina, A.A.; Volchkov, P.V. Epithelial-mesenchymal transition: Role in cancer progression and the perspectives of antitumor treatment. Acta Naturae 2020, 12, 4–23. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Tamma, R.; Annese, T. Epithelial-mesenchymal transition in cancer: A historical overview. Transl. Oncol. 2020, 13, 100773. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Arani, H.Z.; Orouei, S.; Fallah, S.; Ghorbani, A.; Khaledabadi, M.; Kakavand, A.; Tavakolpournegari, A.; Saebfar, H.; Heidari, H.; et al. Emt mechanism in breast cancer metastasis and drug resistance: Revisiting molecular interactions and biological functions. Biomed. Pharmacother. 2022, 155, 113774. [Google Scholar] [CrossRef]
- Palamaris, K.; Felekouras, E.; Sakellariou, S. Epithelial to mesenchymal transition: Key regulator of pancreatic ductal adenocarcinoma progression and chemoresistance. Cancers 2021, 13, 5532. [Google Scholar] [CrossRef]
- Sha, J.; Bai, Y.; Ngo, H.X.; Okui, T.; Kanno, T. Overview of evidence-based chemotherapy for oral cancer: Focus on drug resistance related to the epithelial-mesenchymal transition. Biomolecules 2021, 11, 893. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Mirzaei, S.; Hashemi, F.; Zarrabi, A.; Zabolian, A.; Saleki, H.; Sharifzadeh, S.O.; Soleymani, L.; Daneshi, S.; Hushmandi, K.; et al. New insight towards development of paclitaxel and docetaxel resistance in cancer cells: Emt as a novel molecular mechanism and therapeutic possibilities. Biomed. Pharmacother. 2021, 141, 111824. [Google Scholar] [CrossRef] [PubMed]
- De Las Rivas, J.; Brozovic, A.; Izraely, S.; Casas-Pais, A.; Witz, I.P.; Figueroa, A. Cancer drug resistance induced by emt: Novel therapeutic strategies. Arch. Toxicol. 2021, 95, 2279–2297. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Kalantari, M.; Mohammadinejad, R.; Javaheri, T.; Sethi, G. Association of the epithelial-mesenchymal transition (emt) with cisplatin resistance. Int. J. Mol. Sci. 2020, 21, 4002. [Google Scholar] [CrossRef] [PubMed]
- Dudás, J.; Ladányi, A.; Ingruber, J.; Steinbichler, T.B.; Riechelmann, H. Epithelial to mesenchymal transition: A mechanism that fuels cancer radio/chemoresistance. Cells 2020, 9, 428. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Carstens, J.L.; Kim, J.; Scheible, M.; Kaye, J.; Sugimoto, H.; Wu, C.-C.; LeBleu, V.S.; Kalluri, R. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature 2015, 527, 525–530. [Google Scholar] [CrossRef]
- Fischer, K.R.; Durrans, A.; Lee, S.; Sheng, J.; Li, F.; Wong, S.T.C.; Choi, H.; El Rayes, T.; Ryu, S.; Troeger, J.; et al. Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature 2015, 527, 472–476. [Google Scholar] [CrossRef]
- Debaugnies, M.; Rodríguez-Acebes, S.; Blondeau, J.; Parent, M.-A.; Zocco, M.; Song, Y.; de Maertelaer, V.; Moers, V.; Latil, M.; Dubois, C.; et al. Rhoj controls emt-associated resistance to chemotherapy. Nature 2023, 616, 168–175. [Google Scholar] [CrossRef]
- Petrella, G.; Corsi, F.; Ciufolini, G.; Germini, S.; Capradossi, F.; Pelliccia, A.; Torino, F.; Ghibelli, L.; Cicero, D.O. Metabolic reprogramming of castration-resistant prostate cancer cells as a response to chemotherapy. Metabolites 2022, 13, 65. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, Z.; Zhang, Z.; Zhao, C.; Li, J.; Jiang, J.; Huang, B.; Qin, Y. Puerarin inhibits emt induced by oxaliplatin via targeting carbonic anhydrase xii. Front. Pharmacol. 2022, 13, 969422. [Google Scholar] [CrossRef]
- Boulding, T.; McCuaig, R.D.; Tan, A.; Hardy, K.; Wu, F.; Dunn, J.; Kalimutho, M.; Sutton, C.R.; Forwood, J.K.; Bert, A.G.; et al. Lsd1 activation promotes inducible emt programs and modulates the tumour microenvironment in breast cancer. Sci. Rep. 2018, 8, 73. [Google Scholar] [CrossRef]
- Xie, S.; Fan, S.; Zhang, S.; Chen, W.; Li, Q.; Pan, G.; Zhang, H.; Wang, W.; Weng, B.; Zhang, Z.; et al. Sox8 regulates cancer stem-like properties and cisplatin-induced emt in tongue squamous cell carcinoma by acting on the wnt/beta-catenin pathway. Int. J. Cancer 2018, 142, 1252–1265. [Google Scholar] [CrossRef]
- Zheng, H.C. The molecular mechanisms of chemoresistance in cancers. Oncotarget 2017, 8, 59950–59964. [Google Scholar] [CrossRef]
- Li, P.; Zhang, X.; Wang, H.; Wang, L.; Liu, T.; Du, L.; Yang, Y.; Wang, C. Malat1 is associated with poor response to oxaliplatin-based chemotherapy in colorectal cancer patients and promotes chemoresistance through ezh2. Mol. Cancer Ther. 2017, 16, 739–751. [Google Scholar] [CrossRef]
- Li, Q.-Q.; Chen, Z.-Q.; Cao, X.-X.; Xu, J.-W.; Chen, Y.-Y.; Wang, W.-J.; Chen, Q.; Tang, F.; Liu, X.-P.; Xu, Z.-D. Involvement of nf-kappab/mir-448 regulatory feedback loop in chemotherapy-induced epithelial-mesenchymal transition of breast cancer cells. Cell Death Differ. 2011, 18, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gao, L.; Zhan, N.; Xu, P.; Yang, J.; Yuan, F.; Xu, Y.; Cai, Q.; Geng, R.; Chen, Q. Hypoxia induced ferritin light chain (ftl) promoted epithelia mesenchymal transition and chemoresistance of glioma. J. Exp. Clin. Cancer Res. 2020, 39, 137. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Zhou, S.; Yuan, W.; Cen, F.; Yan, Q. Mechanism of mir-210 involved in epithelial-mesenchymal transition of pancreatic cancer cells under hypoxia. J. Recept. Signal Transduct. Res. 2019, 39, 399–406. [Google Scholar] [CrossRef]
- Okumura, Y.; Noda, T.; Eguchi, H.; Sakamoto, T.; Iwagami, Y.; Yamada, D.; Asaoka, T.; Wada, H.; Kawamoto, K.; Gotoh, K.; et al. Hypoxia-induced plod2 is a key regulator in epithelial-mesenchymal transition and chemoresistance in biliary tract cancer. Ann. Surg. Oncol. 2018, 25, 3728–3737. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-E.; Zhuang, Y.-W.; Zhou, J.-Y.; Liu, S.-L.; Wang, R.-P.; Shu, P. Cinnamaldehyde enhances apoptotic effect of oxaliplatin and reverses epithelial-mesenchymal transition and stemnness in hypoxic colorectal cancer cells. Exp. Cell Res. 2019, 383, 111500. [Google Scholar] [CrossRef]
- Zhou, Q.-Y.; Tu, C.-Y.; Shao, C.-X.; Wang, W.-K.; Zhu, J.-D.; Cai, Y.; Mao, J.-Y.; Chen, W. Gc7 blocks epithelial-mesenchymal transition and reverses hypoxia-induced chemotherapy resistance in hepatocellular carcinoma cells. Am. J. Transl. Res. 2017, 9, 2608–2617. [Google Scholar]
- Adamski, J.; Price, A.; Dive, C.; Makin, G. Hypoxia-induced cytotoxic drug resistance in osteosarcoma is independent of hif-1alpha. PLoS ONE 2013, 8, e65304. [Google Scholar] [CrossRef] [PubMed]
- Cosse, J.-P.; Ronvaux, M.; Ninane, N.; Raes, M.J.; Michiels, C. Hypoxia-induced decrease in p53 protein level and increase in c-jun DNA binding activity results in cancer cell resistance to etoposide. Neoplasia 2009, 11, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Piret, J.-P.; Cosse, J.-P.; Ninane, N.; Raes, M.; Michiels, C. Hypoxia protects hepg2 cells against etoposide-induced apoptosis via a hif-1-independent pathway. Exp. Cell Res. 2006, 312, 2908–2920. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Kobayashi, M.; Darmanin, S.; Qiao, Y.; Gully, C.; Zhao, R.; Yeung, S.C.; Lee, M.H. Pim-1 plays a pivotal role in hypoxia-induced chemoresistance. Oncogene 2009, 28, 2581–2592. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, S.E.; Weigert, A.; Zhou, J.; Brüne, B. Hypoxia enhances sphingosine kinase 2 activity and provokes sphingosine-1-phosphate-mediated chemoresistance in a549 lung cancer cells. Mol. Cancer Res. 2009, 7, 393–401. [Google Scholar] [CrossRef]
- Schnitzer, S.E.; Schmid, T.; Zhou, J.; Brune, B. Hypoxia and hif-1alpha protect a549 cells from drug-induced apoptosis. Cell Death Differ. 2006, 13, 1611–1613. [Google Scholar] [CrossRef]
- Chen, X.; Liu, M.; Meng, F.; Sun, B.; Jin, X.; Jia, C. The long noncoding rna hif1a-as2 facilitates cisplatin resistance in bladder cancer. J. Cell Biochem. 2019, 120, 243–252. [Google Scholar] [CrossRef]
- Su, Y.; Yang, W.; Jiang, N.; Shi, J.; Chen, L.; Zhong, G.; Bi, J.; Dong, W.; Wang, Q.; Wang, C.; et al. Hypoxia-elevated circelp3 contributes to bladder cancer progression and cisplatin resistance. Int. J. Biol. Sci. 2019, 15, 441–452. [Google Scholar] [CrossRef]
- Huang, H.; Peng, J.; Yi, S.; Ding, C.; Ji, W.; Huang, Q.; Zeng, S. Circular rna circube2d2 functions as an oncogenic factor in hepatocellular carcinoma sorafenib resistance and glycolysis. Am. J. Transl. Res. 2021, 13, 6076–6086. [Google Scholar]
- Wang, H.; Min, J.; Xu, C.; Liu, Y.; Yu, Z.; Gong, A.; Xu, M. Hypoxia-elicited exosomes promote the chemoresistance of pancreatic cancer cells by transferring lncror via hippo signaling. J. Cancer 2023, 14, 1075–1087. [Google Scholar] [CrossRef]
- Tao, S.; Wang, J.; Li, F.; Shi, B.; Ren, Q.; Zhuang, Y.; Qian, X. Extracellular vesicles released by hypoxia-induced tumor-associated fibroblasts impart chemoresistance to breast cancer cells via long noncoding rna h19 delivery. FASEB J. 2024, 38, e23165. [Google Scholar] [CrossRef] [PubMed]
- Musah-Eroje, A.; Watson, S. A novel 3d in vitro model of glioblastoma reveals resistance to temozolomide which was potentiated by hypoxia. J. Neurooncol 2019, 142, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.M.; Bandopadhyay, G.; Coyle, B.; Grabowska, A. A hif-independent, cd133-mediated mechanism of cisplatin resistance in glioblastoma cells. Cell. Oncol. 2018, 41, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Zhang, Y.; Fu, X.; Xue, J.; Guo, W.; Meng, M.; Zhou, Z.; Mo, X.; Lu, Y. A tumor hypoxic niche protects human colon cancer stem cells from chemotherapy. J. Cancer Res. Clin. Oncol. 2013, 139, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Saigusa, S.; Tanaka, K.; Toiyama, Y.; Yokoe, T.; Okugawa, Y.; Koike, Y.; Fujikawa, H.; Inoue, Y.; Miki, C.; Kusunoki, M. Clinical significance of cd133 and hypoxia inducible factor-1alpha gene expression in rectal cancer after preoperative chemoradiotherapy. Clin. Oncol. 2011, 23, 323–332. [Google Scholar] [CrossRef]
- Lu, C.; Mahajan, A.; Hong, S.-H.; Galli, S.; Zhu, S.; Tilan, J.U.; Abualsaud, N.; Adnani, M.; Chung, S.; Elmansy, N.; et al. Hypoxia-activated neuropeptide y/y5 receptor/rhoa pathway triggers chromosomal instability and bone metastasis in ewing sarcoma. Nat. Commun. 2022, 13, 2323. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, L.R.; Bilandzic, M.; Wilson, A.L.; Chen, Y.; Gorrell, M.D.; Oehler, M.K.; Plebanski, M.; Stephens, A.N. Hypoxia regulates dpp4 expression, proteolytic inactivation, and shedding from ovarian cancer cells. Int. J. Mol. Sci. 2020, 21, 8110. [Google Scholar] [CrossRef] [PubMed]
- Tilan, J.U.; Lu, C.; Galli, S.; Izycka-Swieszewska, E.; Earnest, J.P.; Shabbir, A.; Everhart, L.M.; Wang, S.; Martin, S.; Horton, M.; et al. Hypoxia shifts activity of neuropeptide y in ewing sarcoma from growth-inhibitory to growth-promoting effects. Oncotarget 2013, 4, 2487–2501. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.-Y.; Zhuang, L.-H.; Hu, Y.; Zhou, Y.-L.; Lin, W.-K.; Wang, D.-D.; Wan, Z.-Q.; Chang, L.-L.; Chen, Y.; Ying, M.-D.; et al. Inactivation of hypoxia-induced yap by statins overcomes hypoxic resistance tosorafenib in hepatocellular carcinoma cells. Sci. Rep. 2016, 6, 30483. [Google Scholar] [CrossRef]
- Lang, L. Fda approves sorafenib for patients with inoperable liver cancer. Gastroenterology 2008, 134, 379. [Google Scholar] [CrossRef]
- Xia, X.; Wang, Q.; Ye, T.; Liu, Y.; Liu, D.; Song, S.; Zheng, C. Nrf2/abcb1-mediated efflux and parp1-mediated dampening of DNA damage contribute to doxorubicin resistance in chronic hypoxic hepg2 cells. Fundam. Clin. Pharmacol. 2020, 34, 41–50. [Google Scholar] [CrossRef]
- Syu, J.-P.; Chi, J.-T.; Kung, H.-N. Nrf2 is the key to chemotherapy resistance in mcf7 breast cancer cells under hypoxia. Oncotarget 2016, 7, 14659–14672. [Google Scholar] [CrossRef]
- Li, T.; Fu, X.; Wang, J.; Shang, W.; Wang, X.; Zhang, L.; Li, J. Mechanism of nurp1 in temozolomide resistance in hypoxia-treated glioma cells via the kdm3a/tfeb axis. Oncol. Res. 2023, 31, 345–359. [Google Scholar] [CrossRef]
- Koritzinsky, M.; Levitin, F.; van den Beucken, T.; Rumantir, R.A.; Harding, N.J.; Chu, K.C.; Boutros, P.C.; Braakman, I.; Wouters, B.G. Two phases of disulfide bond formation have differing requirements for oxygen. J. Cell Biol. 2013, 203, 615–627. [Google Scholar] [CrossRef]
- Bartoszewska, S.; Collawn, J.F.; Bartoszewski, R. The role of the hypoxia-related unfolded protein response (upr) in the tumor microenvironment. Cancers 2022, 14, 4870. [Google Scholar] [CrossRef]
- Chipurupalli, S.; Kannan, E.; Tergaonkar, V.; D’andrea, R.; Robinson, N. Hypoxia induced er stress response as an adaptive mechanism in cancer. Int. J. Mol. Sci. 2019, 20, 749. [Google Scholar] [CrossRef]
- Avril, T.; Vauléon, E.; Chevet, E. Endoplasmic reticulum stress signaling and chemotherapy resistance in solid cancers. Oncogenesis 2017, 6, e373. [Google Scholar] [CrossRef]
- Benedetti, R.; Romeo, M.A.; Arena, A.; Montani, M.S.G.; Di Renzo, L.; D’orazi, G.; Cirone, M. Atf6 prevents DNA damage and cell death in colon cancer cells undergoing er stress. Cell Death Discov. 2022, 8, 295. [Google Scholar] [CrossRef]
- Cho, J.; Min, H.-Y.; Pei, H.; Wei, X.; Sim, J.Y.; Park, S.-H.; Hwang, S.J.; Lee, H.-J.; Hong, S.; Shin, Y.K.; et al. The atf6-egf pathway mediates the awakening of slow-cycling chemoresistant cells and tumor recurrence by stimulating tumor angiogenesis. Cancers 2020, 12, 1772. [Google Scholar] [CrossRef] [PubMed]
- Akman, M.; Belisario, D.C.; Salaroglio, I.C.; Kopecka, J.; Donadelli, M.; De Smaele, E.; Riganti, C. Hypoxia, endoplasmic reticulum stress and chemoresistance: Dangerous liaisons. J. Exp. Clin. Cancer Res. 2021, 40, 28. [Google Scholar] [CrossRef] [PubMed]
- Pi, L.; Li, X.; Song, Q.; Shen, Y.; Lu, X.; DI, B. Knockdown of glucose-regulated protein 78 abrogates chemoresistance of hypopharyngeal carcinoma cells to cisplatin induced by unfolded protein in response to severe hypoxia. Oncol. Lett. 2014, 7, 685–692. [Google Scholar] [CrossRef]
- Lee, D.; Sun, S.; Ho, A.S.; Kiang, K.M.; Zhang, X.Q.; Xu, F.F.; Leung, G.K. Hyperoxia resensitizes chemoresistant glioblastoma cells to temozolomide through unfolded protein response. Anticancer. Res. 2014, 34, 2957–2966. [Google Scholar]
- Bouznad, N.; Rokavec, M.; Öner, M.G.; Hermeking, H. Mir-34a and ire1a/xbp-1(s) form a double-negative feedback loop to regulate hypoxia-induced emt, metastasis, chemo-resistance and autophagy. Cancers 2023, 15, 1143. [Google Scholar] [CrossRef]
- Moszyńska, A.; Collawn, J.F.; Bartoszewski, R. Ire1 endoribonuclease activity modulates hypoxic hif-1alpha signaling in human endothelial cells. Biomolecules 2020, 10, 895. [Google Scholar] [CrossRef]
- Lee, J.H.; Yoon, Y.M.; Lee, S.H. Hypoxic preconditioning promotes the bioactivities of mesenchymal stem cells via the hif-1alpha-grp78-akt axis. Int. J. Mol. Sci. 2017, 18, 1320. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.; Newton, I.P.; Zhang, L.; Ji, P.; Li, Z. Grp78 is implicated in the modulation of tumor aerobic glycolysis by promoting autophagic degradation of ikkbeta. Cell. Signal. 2015, 27, 1237–1245. [Google Scholar] [CrossRef]
- Chen, X.; Iliopoulos, D.; Zhang, Q.; Tang, Q.; Greenblatt, M.B.; Hatziapostolou, M.; Lim, E.; Tam, W.L.; Ni, M.; Chen, Y.; et al. Xbp1 promotes triple-negative breast cancer by controlling the hif1alpha pathway. Nature 2014, 508, 103–107. [Google Scholar] [CrossRef]
- Miharada, K.; Karlsson, G.; Rehn, M.; Rörby, E.; Siva, K.; Cammenga, J.; Karlsson, S. Hematopoietic stem cells are regulated by cripto, as an intermediary of hif-1alpha in the hypoxic bone marrow niche. Ann. N. Y. Acad. Sci. 2012, 1266, 55–62. [Google Scholar] [CrossRef]
- Kang, M.J.; Jung, S.M.; Kim, M.J.; Bae, J.H.; Kim, H.B.; Kim, J.Y.; Park, S.J.; Song, H.S.; Kim, D.W.; Kang, C.D.; et al. DNA-dependent protein kinase is involved in heat shock protein-mediated accumulation of hypoxia-inducible factor-1alpha in hypoxic preconditioned hepg2 cells. FEBS J. 2008, 275, 5969–5981. [Google Scholar] [CrossRef] [PubMed]
- Mivechi, N.F.; Koong, A.C.; Giaccia, A.J.; Hahn, G.M. Analysis of hsf-1 phosphorylation in a549 cells treated with a variety of stresses. Int. J. Hyperth. 1994, 10, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Cyran, A.M.; Zhitkovich, A. Heat shock proteins and hsf1 in cancer. Front. Oncol. 2022, 12, 860320. [Google Scholar] [CrossRef]
- Landriscina, M.; Maddalena, F.; Laudiero, G.; Esposito, F. Adaptation to oxidative stress, chemoresistance, and cell survival. Antioxid. Redox Signal 2009, 11, 2701–2716. [Google Scholar] [CrossRef]
- Vydra, N.; Toma, A.; Glowala-Kosinska, M.; Gogler-Piglowska, A.; Widlak, W. Overexpression of heat shock transcription factor 1 enhances the resistance of melanoma cells to doxorubicin and paclitaxel. BMC Cancer 2013, 13, 504. [Google Scholar] [CrossRef]
- Vilaboa, N.E.; Galán, A.; Troyano, A.; de Blas, E.; Aller, P. Regulation of multidrug resistance 1 (mdr1)/p-glycoprotein gene expression and activity by heat-shock transcription factor 1 (hsf1). J. Biol. Chem. 2000, 275, 24970–24976. [Google Scholar] [CrossRef]
- Yoo, H.J.; Im, C.-N.; Youn, D.-Y.; Yun, H.H.; Lee, J.-H. Bis is induced by oxidative stress via activation of hsf1. Korean J. Physiol. Pharmacol. 2014, 18, 403–409. [Google Scholar] [CrossRef]
- Shanker, M.; Willcutts, D.; Roth, J.A.; Ramesh, R. Drug resistance in lung cancer. Lung Cancer 2010, 1, 23–36. [Google Scholar]
- Banerjee Mustafi, S.; Chakraborty, P.K.; Dey, R.S.; Raha, S. Heat stress upregulates chaperone heat shock protein 70 and antioxidant manganese superoxide dismutase through reactive oxygen species (ros), p38mapk, and akt. Cell Stress Chaperones 2009, 14, 579–589. [Google Scholar] [CrossRef]
- Yoo, H.Y.; Chang, M.S.; Rho, H.M. The activation of the rat copper/zinc superoxide dismutase gene by hydrogen peroxide through the hydrogen peroxide-responsive element and by paraquat and heat shock through the same heat shock element. J. Biol. Chem. 1999, 274, 23887–23892. [Google Scholar] [CrossRef]
- Egger, G.; Liang, G.; Aparicio, A.; Jones, P.A. Epigenetics in human disease and prospects for epigenetic therapy. Nature 2004, 429, 457–463. [Google Scholar] [CrossRef]
- Marmorstein, R.; Zhou, M.-M. Writers and readers of histone acetylation: Structure, mechanism, and inhibition. Cold Spring Harb. Perspect. Biol. 2014, 6, a018762. [Google Scholar] [CrossRef]
- Kim, M.S.; Kwon, H.J.; Lee, Y.M.; Baek, J.H.; Jang, J.-E.; Lee, S.-W.; Moon, E.-J.; Kim, H.-S.; Lee, S.-K.; Chung, H.Y.; et al. Histone deacetylases induce angiogenesis by negative regulation of tumor suppressor genes. Nat. Med. 2001, 7, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Pluemsampant, S.; Safronova, O.S.; Nakahama, K.; Morita, I. Protein kinase ck2 is a key activator of histone deacetylase in hypoxia-associated tumors. Int. J. Cancer 2008, 122, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Kirmes, I.; Szczurek, A.; Prakash, K.; Charapitsa, I.; Heiser, C.; Musheev, M.; Schock, F.; Fornalczyk, K.; Ma, D.; Birk, U.; et al. A transient ischemic environment induces reversible compaction of chromatin. Genome Biol. 2015, 16, 246. [Google Scholar] [CrossRef] [PubMed]
- Leszczynska, K.B.; Dzwigonska, M.; Estephan, H.; Moehlenbrink, J.; Bowler, E.; Giaccia, A.J.; Mieczkowski, J.; Kaminska, B.; Hammond, E.M. Hypoxia-mediated regulation of ddx5 through decreased chromatin accessibility and post-translational targeting restricts r-loop accumulation. Mol. Oncol. 2023, 17, 1173–1191. [Google Scholar] [CrossRef] [PubMed]
- Gujral, P.; Mahajan, V.; Lissaman, A.C.; Ponnampalam, A.P. Histone acetylation and the role of histone deacetylases in normal cyclic endometrium. Reprod. Biol. Endocrinol. 2020, 18, 84. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-Q.; Pan, B.-Z.; Huang, J.-Y.; Zhang, K.; Cui, S.-Y.; De, W.; Wang, R.; Chen, L.-B. Hdac 1/4-mediated silencing of microrna-200b promotes chemoresistance in human lung adenocarcinoma cells. Oncotarget 2014, 5, 3333–3349. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Langdon, S.P.; Tse, M.; Mullen, P.; Um, I.H.; Faratian, D.; Harrison, D.J. The role of hdac2 in chromatin remodelling and response to chemotherapy in ovarian cancer. Oncotarget 2016, 7, 4695–4711. [Google Scholar] [CrossRef] [PubMed]
- Taddei, A.; Roche, D.; Bickmore, W.A.; Almouzni, G. The effects of histone deacetylase inhibitors on heterochromatin: Implications for anticancer therapy? EMBO Rep. 2005, 6, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, S.A.; Sahu, K.K.; Sengupta, S.; Partap, S.; Karpoormath, R.; Kumar, B.; Kumar, D. Recent advancement of hdac inhibitors against breast cancer. Med. Oncol. 2023, 40, 201. [Google Scholar] [CrossRef]
- Falkenberg, K.J.; Johnstone, R.W. Histone deacetylases and their inhibitors in cancer, neurological diseases and immune disorders. Nat. Rev. Drug Discov. 2014, 13, 673–691. [Google Scholar] [CrossRef]
- Hu, Z.; Wei, F.; Su, Y.; Wang, Y.; Shen, Y.; Fang, Y.; Ding, J.; Chen, Y. Histone deacetylase inhibitors promote breast cancer metastasis by elevating nedd9 expression. Signal Transduct. Target. Ther. 2023, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Quinn, D.I.; Tsao-Wei, D.D.; Twardowski, P.; Aparicio, A.M.; Frankel, P.; Chatta, G.; Wright, J.J.; Groshen, S.G.; Khoo, S.; Lenz, H.-J.; et al. Phase ii study of the histone deacetylase inhibitor vorinostat (suberoylanilide hydroxamic acid; saha) in recurrent or metastatic transitional cell carcinoma of the urothelium—An nci-ctep sponsored: California cancer consortium trial, nci 6879. Investig. New Drugs 2021, 39, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.; Filippakopoulos, P. Functions of bromodomain-containing proteins and their roles in homeostasis and cancer. Nat. Rev. Mol. Cell Biol. 2017, 18, 246–262. [Google Scholar] [CrossRef]
- Filippakopoulos, P.; Picaud, S.; Mangos, M.; Keates, T.; Lambert, J.-P.; Barsyte-Lovejoy, D.; Felletar, I.; Volkmer, R.; Müller, S.; Pawson, T.; et al. Histone recognition and large-scale structural analysis of the human bromodomain family. Cell 2012, 149, 214–231. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Zhao, Y.; Wang, X.; Xie, X.; Liu, M.; Zhang, K.; Wang, L.; Bai, D.; Foster, L.J.; Shu, R.; et al. Targeting bromodomain-containing proteins: Research advances of drug discovery. Mol. Biomed. 2023, 4, 13. [Google Scholar] [CrossRef]
- Zaware, N.; Zhou, M.-M. Bromodomain biology and drug discovery. Nat. Struct. Mol. Biol. 2019, 26, 870–879. [Google Scholar] [CrossRef]
- Muller, S.; Filippakopoulos, P.; Knapp, S. Bromodomains as therapeutic targets. Expert. Rev. Mol. Med. 2011, 13, e29. [Google Scholar] [CrossRef] [PubMed]
- Gajjela, B.K.; Zhou, M.-M. Bromodomain inhibitors and therapeutic applications. Curr. Opin. Chem. Biol. 2023, 75, 102323. [Google Scholar] [CrossRef] [PubMed]
- Lazarchuk, P.; Hernandez-Villanueva, J.; Pavlova, M.N.; Federation, A.; MacCoss, M.; Sidorova, J.M. Mutual balance of histone deacetylases 1 and 2 and the acetyl reader atad2 regulates the level of acetylation of histone h4 on nascent chromatin of human cells. Mol. Cell. Biol. 2020, 40, e00421-19. [Google Scholar] [CrossRef]
- Koo, S.J.; Fernández-Montalván, A.E.; Badock, V.; Ott, C.J.; Holton, S.J.; von Ahsen, O.; Toedling, J.; Vittori, S.; Bradner, J.E.; Gorjánácz, M. Atad2 is an epigenetic reader of newly synthesized histone marks during DNA replication. Oncotarget 2016, 7, 70323–70335. [Google Scholar] [CrossRef]
- Revenko, A.S.; Kalashnikova, E.V.; Gemo, A.T.; Zou, J.X.; Chen, H.-W. Chromatin loading of e2f-mll complex by cancer-associated coregulator ancca via reading a specific histone mark. Mol. Cell. Biol. 2010, 30, 5260–5272. [Google Scholar] [CrossRef]
- Ciró, M.; Prosperini, E.; Quarto, M.; Grazini, U.; Walfridsson, J.; McBlane, F.; Nucifero, P.; Pacchiana, G.; Capra, M.; Christensen, J.; et al. Atad2 is a novel cofactor for myc, overexpressed and amplified in aggressive tumors. Cancer Res. 2009, 69, 8491–8498. [Google Scholar] [CrossRef]
- Losman, J.-A.; Koivunen, P.; Kaelin, W.G., Jr. 2-oxoglutarate-dependent dioxygenases in cancer. Nat. Rev. Cancer 2020, 20, 710–726. [Google Scholar] [CrossRef]
- Kao, T.-W.; Bai, G.-H.; Wang, T.-L.; Shih, I.-M.; Chuang, C.-M.; Lo, C.-L.; Tsai, M.-C.; Chiu, L.-Y.; Lin, C.-C.; Shen, Y.-A. Novel cancer treatment paradigm targeting hypoxia-induced factor in conjunction with current therapies to overcome resistance. J. Exp. Clin. Cancer Res. 2023, 42, 171. [Google Scholar] [CrossRef]
- Meehan, R.; Kummar, S.; Do, K.; Coyne, G.O.; Juwara, L.; Zlott, J.; Rubinstein, L.; Doroshow, J.H.; Chen, A.P. A phase i study of ganetespib and ziv-aflibercept in patients with advanced carcinomas and sarcomas. Oncologist 2018, 23, 1269-e125. [Google Scholar] [CrossRef]
- Kim, A.; Lu, Y.; Okuno, S.H.; Reinke, D.; Maertens, O.; Perentesis, J.; Basu, M.; Wolters, P.L.; De Raedt, T.; Chawla, S.; et al. Targeting refractory sarcomas and malignant peripheral nerve sheath tumors in a phase i/ii study of sirolimus in combination with ganetespib (sarc023). Sarcoma 2020, 2020, 5784876. [Google Scholar] [CrossRef]
- Ramalingam, S.; Goss, G.; Rosell, R.; Schmid-Bindert, G.; Zaric, B.; Andric, Z.; Bondarenko, I.; Komov, D.; Ceric, T.; Khuri, F.; et al. A randomized phase ii study of ganetespib, a heat shock protein 90 inhibitor, in combination with docetaxel in second-line therapy of advanced non-small cell lung cancer (galaxy-1). Ann. Oncol. 2015, 26, 1741–1748. [Google Scholar] [CrossRef]
- Fallah, J.; Brave, M.H.; Weinstock, C.; Mehta, G.U.; Bradford, D.; Gittleman, H.; Bloomquist, E.W.; Charlab, R.; Hamed, S.S.; Miller, C.P.; et al. Fda approval summary: Belzutifan for von hippel-lindau disease-associated tumors. Clin. Cancer Res. 2022, 28, 4843–4848. [Google Scholar] [CrossRef]
- Strowd, R.; Ellingson, B.; Raymond, C.; Yao, J.; Wen, P.Y.; Ahluwalia, M.; Piotrowski, A.; Desai, A.; Clarke, J.L.; Lieberman, F.S.; et al. Activity of a first-in-class oral hif2-alpha inhibitor, pt2385, in patients with first recurrence of glioblastoma. J. Neuro-Oncol. 2023, 165, 101–112. [Google Scholar] [CrossRef]
- Xu, P.; Wu, Q.; Yu, J.; Rao, Y.; Kou, Z.; Fang, G.; Shi, X.; Liu, W.; Han, H. A systematic way to infer the regulation relations of mirnas on target genes and critical mirnas in cancers. Front. Genet. 2020, 11, 278. [Google Scholar] [CrossRef]
- He, Z.; Zhu, Q. Circular rnas: Emerging roles and new insights in human cancers. Biomed. Pharmacother. 2023, 165, 115217. [Google Scholar] [CrossRef]
- Brenner, A.J.; Floyd, J.; Fichtel, L.; Michalek, J.; Kanakia, K.P.; Huang, S.; Reardon, D.; Wen, P.Y.; Lee, E.Q. Phase 2 trial of hypoxia activated evofosfamide (th302) for treatment of recurrent bevacizumab-refractory glioblastoma. Sci. Rep. 2021, 11, 2306. [Google Scholar] [CrossRef]
- García-Venzor, A.; Mandujano-Tinoco, E.A.; Ruiz-Silvestre, A.; Sánchez, J.M.; Lizarraga, F.; Zampedri, C.; Melendez-Zajgla, J.; Maldonado, V. Lncmat2b regulated by severe hypoxia induces cisplatin resistance by increasing DNA damage repair and tumor-initiating population in breast cancer cells. Carcinogenesis 2020, 41, 1485–1497. [Google Scholar] [CrossRef]
- Yang, H.; Hu, Y.; Weng, M.; Liu, X.; Wan, P.; Hu, Y.; Ma, M.; Zhang, Y.; Xia, H.; Lv, K. Hypoxia inducible lncrna-cbslr modulates ferroptosis through m6a-ythdf2-dependent modulation of cbs in gastric cancer. J. Adv. Res. 2022, 37, 91–106. [Google Scholar] [CrossRef]
- Yin, X.; Liao, Y.; Xiong, W.; Zhang, Y.; Zhou, Y.; Yang, Y. Hypoxia-induced lncrna anril promotes cisplatin resistance in retinoblastoma cells through regulating abcg2 expression. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1049–1057. [Google Scholar] [CrossRef]
- Huan, L.; Guo, T.; Wu, Y.; Xu, L.; Huang, S.; Xu, Y.; Liang, L.; He, X. Hypoxia induced lucat1/ptbp1 axis modulates cancer cell viability and chemotherapy response. Mol. Cancer 2020, 19, 11. [Google Scholar] [CrossRef]
- Moreno Leon, L.; Gautier, M.; Allan, R.; Ilié, M.; Nottet, N.; Pons, N.; Paquet, A.; Lebrigand, K.; Truchi, M.; Fassy, J.; et al. The nuclear hypoxia-regulated nlucat1 long non-coding rna contributes to an aggressive phenotype in lung adenocarcinoma through regulation of oxidative stress. Oncogene 2019, 38, 7146–7165. [Google Scholar] [CrossRef]
- Wang, F.; Ji, X.; Wang, J.; Ma, X.; Yang, Y.; Zuo, J.; Cui, J. Lncrna pvt1 enhances proliferation and cisplatin resistance via regulating mir-194-5p/hif1a axis in oral squamous cell carcinoma. Onco Targets Ther. 2020, 13, 243–252. [Google Scholar] [CrossRef]
- Xu, F.; Huang, M.; Chen, Q.; Niu, Y.; Hu, Y.; Hu, P.; Chen, D.; He, C.; Huang, K.; Zeng, Z.; et al. Lncrna hif1a-as1 promotes gemcitabine resistance of pancreatic cancer by enhancing glycolysis through modulating the akt/yb1/hif1alpha pathway. Cancer Res. 2021, 81, 5678–5691. [Google Scholar] [CrossRef]
- Güçlü, E.; Güneş, C.E.; Kurar, E.; Vural, H. Knockdown of lncrna hif1a-as2 increases drug sensitivity of sclc cells in association with autophagy. Med. Oncol. 2021, 38, 113. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, H.; Zhang, Y.; Xiao, X.; Chu, F. Induction of lncrna norad accounts for hypoxia-induced chemoresistance and vasculogenic mimicry in colorectal cancer by sponging the mir-495-3p/ hypoxia-inducible factor-1alpha (hif-1alpha). Bioengineered 2022, 13, 950–962. [Google Scholar] [CrossRef]
- Weng, X.; Liu, H.; Ruan, J.; Du, M.; Wang, L.; Mao, J.; Cai, Y.; Lu, X.; Chen, W.; Huang, Y.; et al. Hotair/mir-1277-5p/zeb1 axis mediates hypoxia-induced oxaliplatin resistance via regulating epithelial-mesenchymal transition in colorectal cancer. Cell Death Discov. 2022, 8, 310. [Google Scholar] [CrossRef]
- Zhu, Z.-J.; Pang, Y.; Jin, G.; Zhang, H.-Y.; Wang, W.-H.; Liu, J.-W.; Tuo, G.-X.; Wu, P.; Yang, Y.; Wang, Z.-Q.; et al. Hypoxia induces chemoresistance of esophageal cancer cells to cisplatin through regulating the lncrna-ems/mir-758-3p/wtap axis. Aging 2021, 13, 17155–17176. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, P.W.T.; Koseki, L.R.; Haitani, T.; Harada, H.; Kobayashi, M. Hypoxia-Inducible Factor-Dependent and Independent Mechanisms Underlying Chemoresistance of Hypoxic Cancer Cells. Cancers 2024, 16, 1729. https://doi.org/10.3390/cancers16091729
Lee PWT, Koseki LR, Haitani T, Harada H, Kobayashi M. Hypoxia-Inducible Factor-Dependent and Independent Mechanisms Underlying Chemoresistance of Hypoxic Cancer Cells. Cancers. 2024; 16(9):1729. https://doi.org/10.3390/cancers16091729
Chicago/Turabian StyleLee, Peter Wai Tik, Lina Rochelle Koseki, Takao Haitani, Hiroshi Harada, and Minoru Kobayashi. 2024. "Hypoxia-Inducible Factor-Dependent and Independent Mechanisms Underlying Chemoresistance of Hypoxic Cancer Cells" Cancers 16, no. 9: 1729. https://doi.org/10.3390/cancers16091729
APA StyleLee, P. W. T., Koseki, L. R., Haitani, T., Harada, H., & Kobayashi, M. (2024). Hypoxia-Inducible Factor-Dependent and Independent Mechanisms Underlying Chemoresistance of Hypoxic Cancer Cells. Cancers, 16(9), 1729. https://doi.org/10.3390/cancers16091729