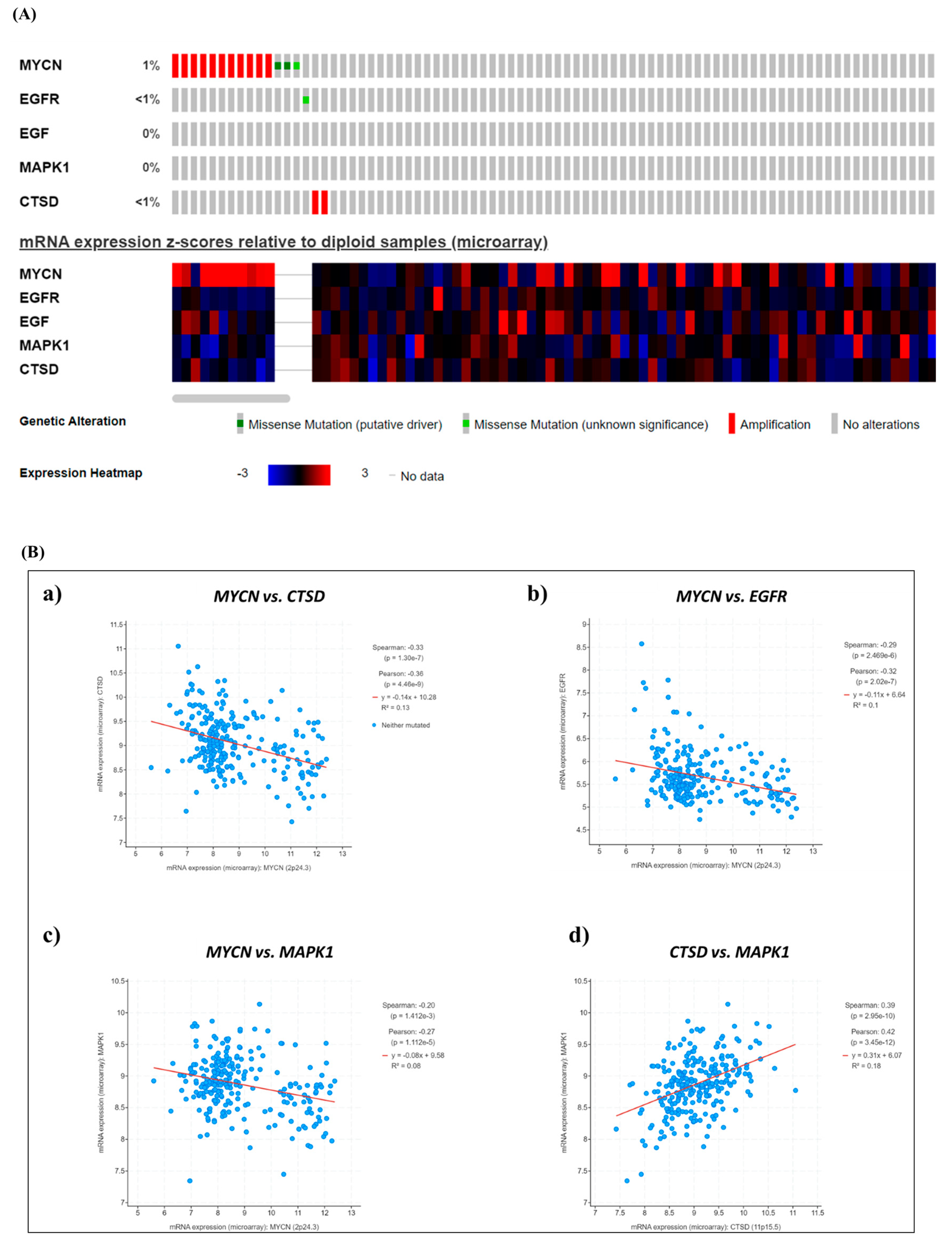
Figure 1.
Neuroblastoma patients bearing a tumor with MYCN amplification or high MYCN mRNA levels show a poor prognosis. (A) Oncoprint of somatic mutations shows the genetic alterations and mRNA expression levels of MYCN, EGFR, EGF, MAPK1 and CTSD in pediatric NB patients (TARGET, 2018). (B) Scatter plots representing the correlations between MYCN–CTSD (a), MYCN–EGFR (b), MYCN–MAPK1 (c) and CTSD–MAPK1 (d) mRNA expression.
Figure 1.
Neuroblastoma patients bearing a tumor with MYCN amplification or high MYCN mRNA levels show a poor prognosis. (A) Oncoprint of somatic mutations shows the genetic alterations and mRNA expression levels of MYCN, EGFR, EGF, MAPK1 and CTSD in pediatric NB patients (TARGET, 2018). (B) Scatter plots representing the correlations between MYCN–CTSD (a), MYCN–EGFR (b), MYCN–MAPK1 (c) and CTSD–MAPK1 (d) mRNA expression.
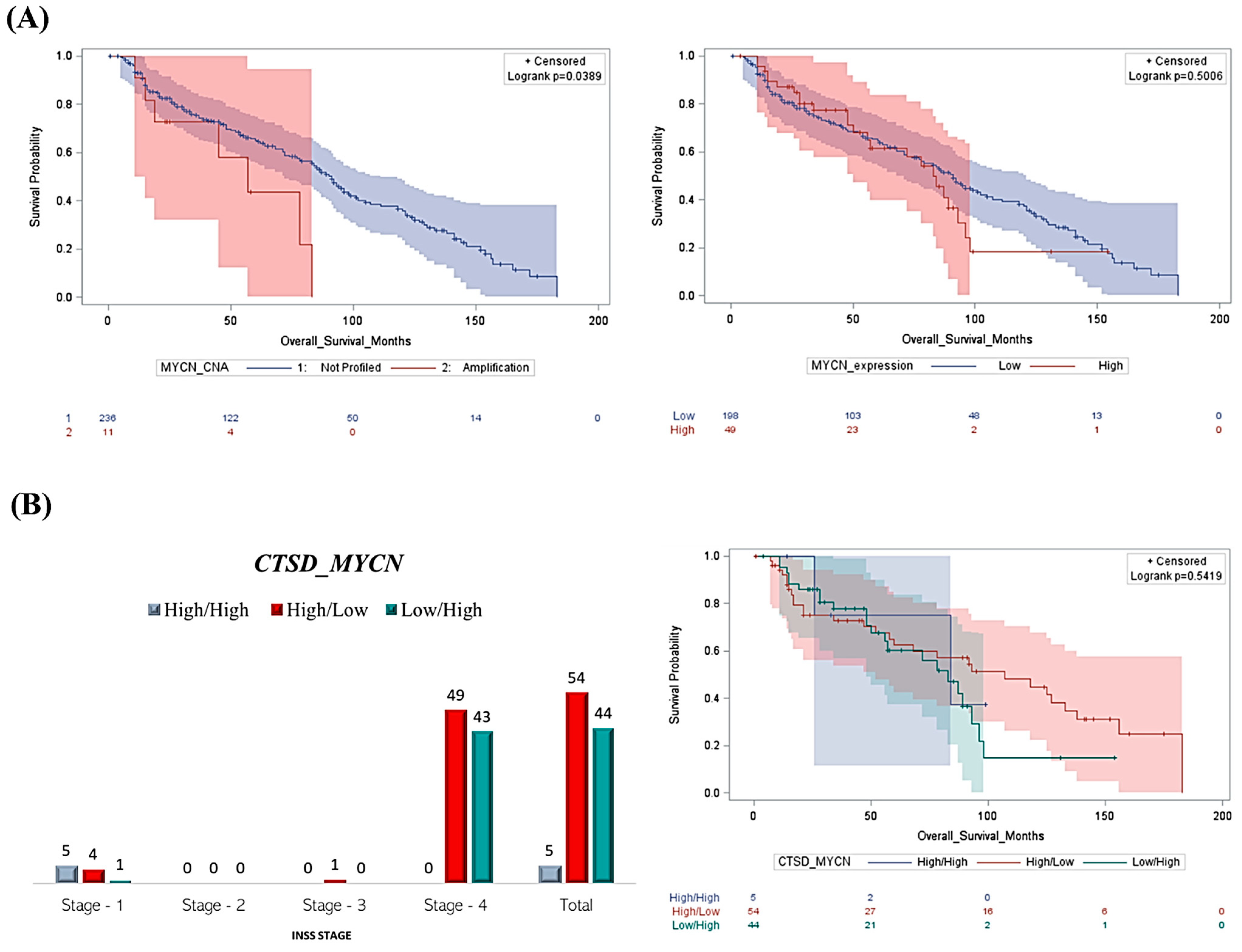
Figure 2.
Patients with high CTSD and low MYCN expression show better clinical outcomes despite the advanced stage. (A) The Kaplan–Meier curves reporting the overall survival rate were obtained based on MYCN copy number alterations and MYCN mRNA expression (high versus low). (B) Graph reporting the INSS staging of NB patients based on CTSD and MYCN expression and Kaplan–Meier plot representing the overall survival of NB patients according to a differential expression of CTSD and MYCN (High/High, High/Low, and Low/High groups, respectively).
Figure 2.
Patients with high CTSD and low MYCN expression show better clinical outcomes despite the advanced stage. (A) The Kaplan–Meier curves reporting the overall survival rate were obtained based on MYCN copy number alterations and MYCN mRNA expression (high versus low). (B) Graph reporting the INSS staging of NB patients based on CTSD and MYCN expression and Kaplan–Meier plot representing the overall survival of NB patients according to a differential expression of CTSD and MYCN (High/High, High/Low, and Low/High groups, respectively).
Figure 3.
EGF differentially modulates cathepsin D levels and cell proliferation in MYCN-amplified and MYCN-not-amplified neuroblastoma cell lines. IMR-32, LAN-5, SK-N-BE(2) and SH-SY5Y cells were cultured for 72 h, medium was renewed, and EGF was re-added every day. Samples were collected after 72 h and processed for Western blot analysis of CD. (A) Western blotting of CD expression in NB cell homogenates. The membrane was probed with GAPDH as loading control. The blot is representative of three independent experiments. Densitometry of the bands is reported in the histogram. One-way ANOVA test was performed. Significance was considered as follow: ** p < 0.01. (B) Cells were seeded, allowed to adhere for 24 h and then counted before (time 0) and at the end of the treatment (72 h) with PstA, EGF or both. Doubling time (Dt) was calculated as described in the methods. Two-way ANOVA test was performed. Significance was considered as follow: **** p < 0.0001; *** p < 0.001; * p < 0.05. “Co” refers to untreated cells (control).
Figure 3.
EGF differentially modulates cathepsin D levels and cell proliferation in MYCN-amplified and MYCN-not-amplified neuroblastoma cell lines. IMR-32, LAN-5, SK-N-BE(2) and SH-SY5Y cells were cultured for 72 h, medium was renewed, and EGF was re-added every day. Samples were collected after 72 h and processed for Western blot analysis of CD. (A) Western blotting of CD expression in NB cell homogenates. The membrane was probed with GAPDH as loading control. The blot is representative of three independent experiments. Densitometry of the bands is reported in the histogram. One-way ANOVA test was performed. Significance was considered as follow: ** p < 0.01. (B) Cells were seeded, allowed to adhere for 24 h and then counted before (time 0) and at the end of the treatment (72 h) with PstA, EGF or both. Doubling time (Dt) was calculated as described in the methods. Two-way ANOVA test was performed. Significance was considered as follow: **** p < 0.0001; *** p < 0.001; * p < 0.05. “Co” refers to untreated cells (control).
![Cancers 16 01343 g003]()
Figure 4.
Cathepsin D expression differs in 2D and 3D SH-SY5Y cell cultures. SH-SY5Y cells were seeded both in adherent and non-adherent Petri dishes, at 4000 cells/cm2 and 1,000,000 cells/Petri, respectively, and allowed to grow for 48 h. Medium was refreshed every 48 h, supplemented with 100 μM PstA where indicated. (A) Cell growth was monitored at the phase-contrast microscope and images were acquired at the time point indicated (time 0 h, 2-, 5- and 7-days). Scale bar = 100 μm; magnification = 20×. Viable SH-SY5Y cells grown in adherent condition were counted, and data from triplicate for each experimental condition are shown in the graph. One-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; *** p < 0.001. (B) The spheroid’s growth was monitored at the phase-contrast microscope, and images were acquired at the time point indicated (time 0 h, 2-, 5- and 7-days). Scale bar = 100 μm; magnification = 20×. The quantification of 3D spheroid’s size was performed with ImageJ software (v.1.48). The area is indicated as arbitrary unit (A.U.). Data represent the average ± S.D. calculated for at least 5 to 10 spheroids for each condition in three separate experiments. t-test was performed. Significance was considered as follows: ** p < 0.01. (C) Western blotting analysis of CD expression in 2D and 3D cell homogenates. The membrane was probed with β-tubulin as loading control. The blot is representative of three independent experiments. Densitometry of the bands is reported in the histogram. Two-way ANOVA test was performed. Significance was considered as follow: ** p < 0.01. “Co” refers to untreated cells (control).
Figure 4.
Cathepsin D expression differs in 2D and 3D SH-SY5Y cell cultures. SH-SY5Y cells were seeded both in adherent and non-adherent Petri dishes, at 4000 cells/cm2 and 1,000,000 cells/Petri, respectively, and allowed to grow for 48 h. Medium was refreshed every 48 h, supplemented with 100 μM PstA where indicated. (A) Cell growth was monitored at the phase-contrast microscope and images were acquired at the time point indicated (time 0 h, 2-, 5- and 7-days). Scale bar = 100 μm; magnification = 20×. Viable SH-SY5Y cells grown in adherent condition were counted, and data from triplicate for each experimental condition are shown in the graph. One-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; *** p < 0.001. (B) The spheroid’s growth was monitored at the phase-contrast microscope, and images were acquired at the time point indicated (time 0 h, 2-, 5- and 7-days). Scale bar = 100 μm; magnification = 20×. The quantification of 3D spheroid’s size was performed with ImageJ software (v.1.48). The area is indicated as arbitrary unit (A.U.). Data represent the average ± S.D. calculated for at least 5 to 10 spheroids for each condition in three separate experiments. t-test was performed. Significance was considered as follows: ** p < 0.01. (C) Western blotting analysis of CD expression in 2D and 3D cell homogenates. The membrane was probed with β-tubulin as loading control. The blot is representative of three independent experiments. Densitometry of the bands is reported in the histogram. Two-way ANOVA test was performed. Significance was considered as follow: ** p < 0.01. “Co” refers to untreated cells (control).
![Cancers 16 01343 g004]()
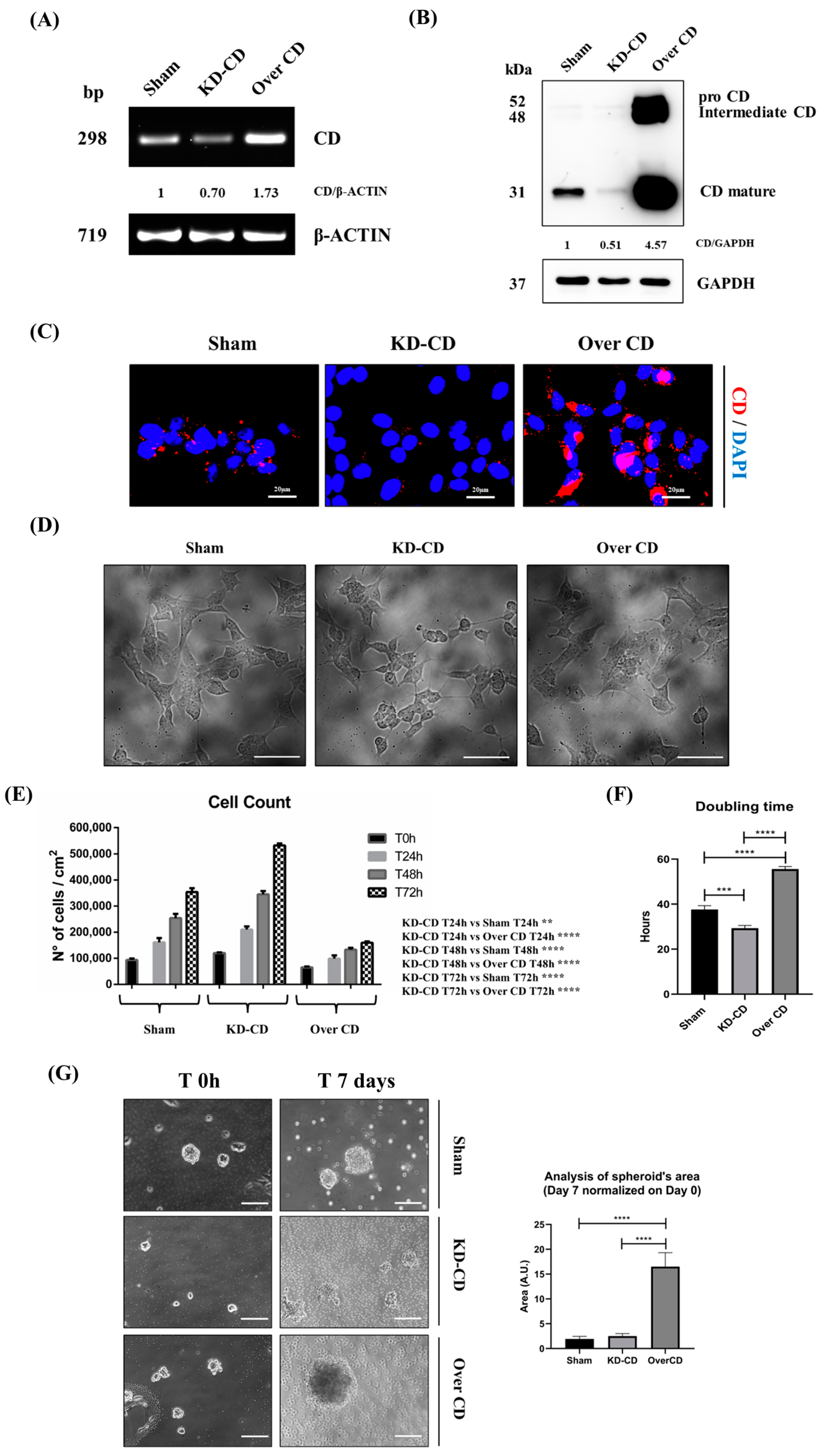
Figure 5.
SH-SY5Y Sham, KD-CD and Over CD clones show different growth rates. (A) Agarose gel electrophoresis of the products of the RT-PCR for CD and β-actin. Densitometric analysis of CD/actin ratio is shown. (B) Western blotting showing the expression of CD in the three SH-SY5Y clones. The blot was probed with GAPDH as loading control. Densitometric analysis of CD/GAPDH ratio is shown. (C) Immunofluorescence performed on SH-SY5Y clones. Cells were seeded on sterile coverslips and then fixed and stained for CD (red); nuclei were marked with the UV fluorescent dye DAPI. Scale bar = 20 µm; magnification = 63×. (D) Phase contrast images of the three SH-SY5Y clones. (E) Graph representing cell count. Cell counting was performed in triplicate for each experimental condition. Two-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; ** p < 0.01. (F) Graph reporting the doubling time of the three SH-SY5Y clones. One-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; *** p < 0.001. (G) 3D cultures of neuroblastoma clones. SH-SY5Y Sham, KD-CD and Over CD cells were seeded on non-adherent Petri dishes and maintained in culture for 7 days. New fresh medium was replaced every 48 h. The 3D spheroid’s growth was monitored at the phase-contrast microscope, and pictures were acquired. Scale bar = 100 μm; magnification = 20×. The quantification of 3D spheroid’s size was performed with ImageJ software (v.1.48). The area was indicated as arbitrary unit (A.U.). Data represent the average ± S.D, calculated for at least 5 to 10 spheroids for each condition in three separate experiments. One-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001.
Figure 5.
SH-SY5Y Sham, KD-CD and Over CD clones show different growth rates. (A) Agarose gel electrophoresis of the products of the RT-PCR for CD and β-actin. Densitometric analysis of CD/actin ratio is shown. (B) Western blotting showing the expression of CD in the three SH-SY5Y clones. The blot was probed with GAPDH as loading control. Densitometric analysis of CD/GAPDH ratio is shown. (C) Immunofluorescence performed on SH-SY5Y clones. Cells were seeded on sterile coverslips and then fixed and stained for CD (red); nuclei were marked with the UV fluorescent dye DAPI. Scale bar = 20 µm; magnification = 63×. (D) Phase contrast images of the three SH-SY5Y clones. (E) Graph representing cell count. Cell counting was performed in triplicate for each experimental condition. Two-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; ** p < 0.01. (F) Graph reporting the doubling time of the three SH-SY5Y clones. One-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; *** p < 0.001. (G) 3D cultures of neuroblastoma clones. SH-SY5Y Sham, KD-CD and Over CD cells were seeded on non-adherent Petri dishes and maintained in culture for 7 days. New fresh medium was replaced every 48 h. The 3D spheroid’s growth was monitored at the phase-contrast microscope, and pictures were acquired. Scale bar = 100 μm; magnification = 20×. The quantification of 3D spheroid’s size was performed with ImageJ software (v.1.48). The area was indicated as arbitrary unit (A.U.). Data represent the average ± S.D, calculated for at least 5 to 10 spheroids for each condition in three separate experiments. One-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001.
![Cancers 16 01343 g005]()
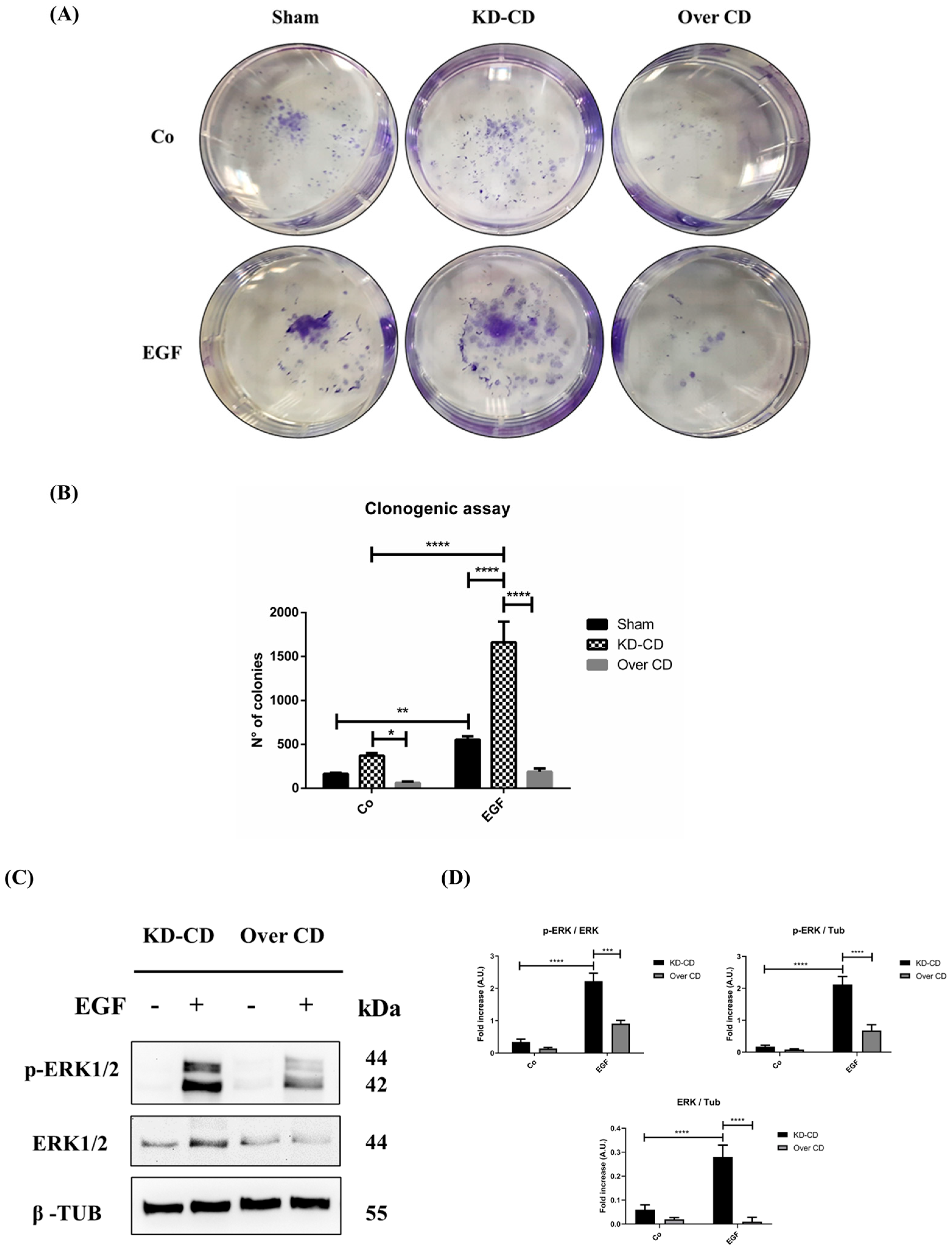
Figure 6.
EGF stimulation of neuroblastoma growth depends on cellular level of cathepsin D. (A) Clonogenic assay performed on SH-SY5Y transgenic clones upon stimulation with 20 ng/mL EGF. Colonies were stained as described in the Methods section. (B) Cell growth and number of colonies were estimated through photometric measurements and CellCounter Software and are shown in the graph. Data ± S.D. are representative of three independent replicates. (C) Western blotting showing the phosphorylation of ERK 1/2 (p-ERK 1/2) in KD-CD and Over CD cells treated with EGF. Cells were cultured for 72 h; medium was renewed and EGF re-added every day. Samples were processed for Western blot analysis of p-ERK and total ERK. Membranes were probed with β-tubulin as loading control. All blots are representative of three independent experiments. (D) Densitometry of the p-ERK/ERK, p-ERK/Tubulin and ERK/Tubulin ratios are reported in the histograms. Two-way ANOVA test was performed in all the statistical analysis reported in the figure. Significance was considered as follows: **** p < 0.0001; *** p < 0.001; ** p < 0.01; * p < 0.05. “Co” refers to untreated cells (control).
Figure 6.
EGF stimulation of neuroblastoma growth depends on cellular level of cathepsin D. (A) Clonogenic assay performed on SH-SY5Y transgenic clones upon stimulation with 20 ng/mL EGF. Colonies were stained as described in the Methods section. (B) Cell growth and number of colonies were estimated through photometric measurements and CellCounter Software and are shown in the graph. Data ± S.D. are representative of three independent replicates. (C) Western blotting showing the phosphorylation of ERK 1/2 (p-ERK 1/2) in KD-CD and Over CD cells treated with EGF. Cells were cultured for 72 h; medium was renewed and EGF re-added every day. Samples were processed for Western blot analysis of p-ERK and total ERK. Membranes were probed with β-tubulin as loading control. All blots are representative of three independent experiments. (D) Densitometry of the p-ERK/ERK, p-ERK/Tubulin and ERK/Tubulin ratios are reported in the histograms. Two-way ANOVA test was performed in all the statistical analysis reported in the figure. Significance was considered as follows: **** p < 0.0001; *** p < 0.001; ** p < 0.01; * p < 0.05. “Co” refers to untreated cells (control).
![Cancers 16 01343 g006]()
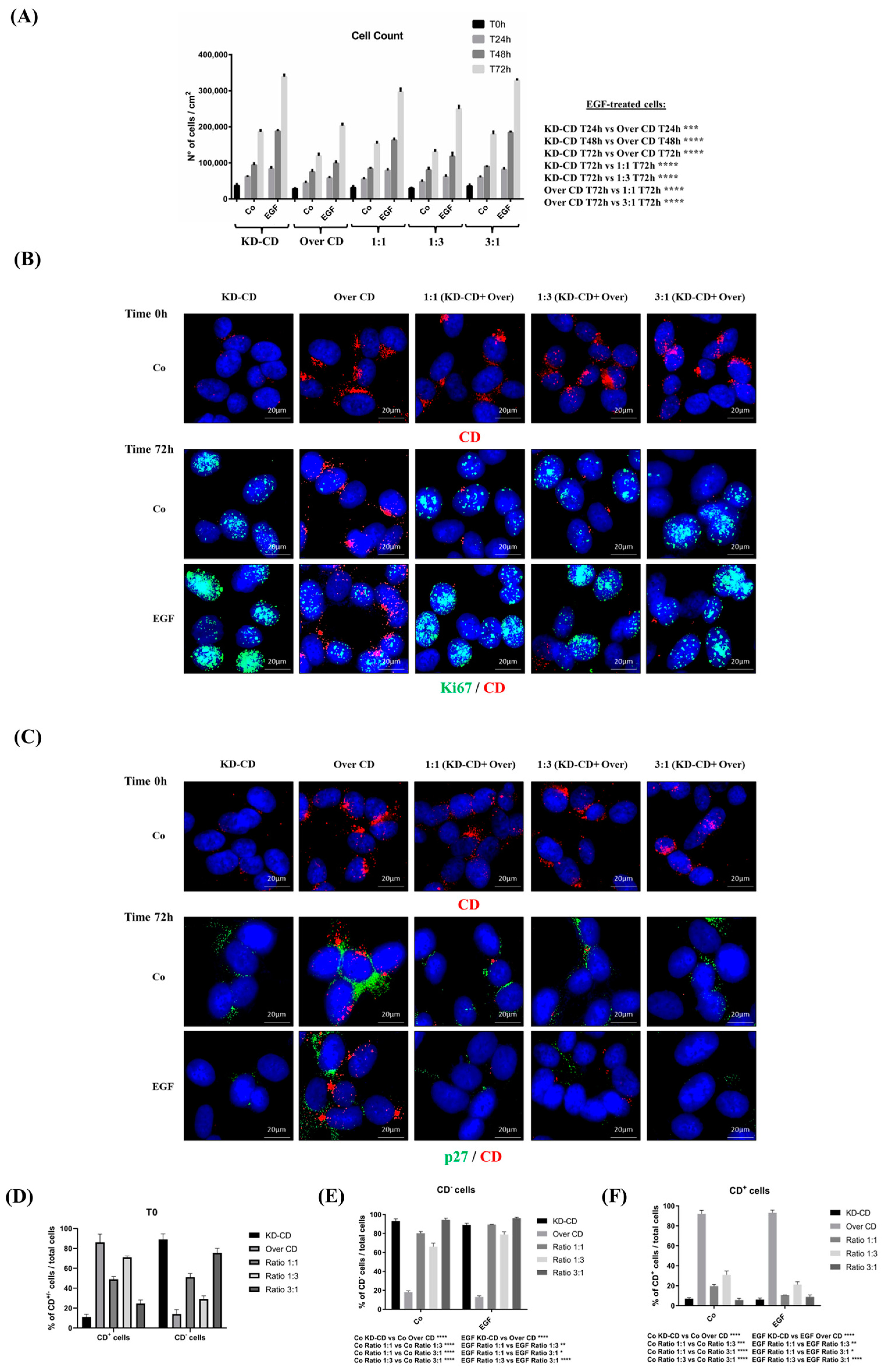
Figure 7.
CD knockdown clone expresses high level of nuclear Ki-67 proliferation marker and reduced p27 cell cycle inhibitor. (A) Cell counting of viable cells of KD-CD or Over CD or a mix of both at the ratio indicated, in the absence or the presence of EGF. Medium was renewed and EGF added every 24 h. The cells were cultivated for 24, 48 and 72 h. (B,C) At the beginning of the experiment (time 0), cells were fixed and stained for CD to confirm the composition of co-cultures. Immunofluorescence double staining of CD and Ki-67 or p27 was performed on SH-SY5Y KD-CD, Over CD and mixed cultures at different ratios: 50% KD-CD + 50% Over CD cells (1:1), 25% KD-CD + 75% Over CD (1:3) and 75% KD-CD + 25% Over CD cells (3:1). Fresh medium was replaced every day, and EGF was added as indicated. After 72 h of treatment, cells were fixed and stained for Ki-67 (green)/CD (red) (B) and p27 (green)/CD (red) (C). Scale bar = 20 µm; magnification = 63×. Representative images of three independent experiments are shown. (D–F) Percentages of CD negative and CD positive cells relative to the total cell population, calculated in random fields, are shown in the graphs. Two-way ANOVA test was performed in all the statistical analysis reported in the graphs. Significance was considered as follows: **** p < 0.0001; *** p < 0.001; ** p < 0.01; * p < 0.05. “Co” refers to untreated cells (control).
Figure 7.
CD knockdown clone expresses high level of nuclear Ki-67 proliferation marker and reduced p27 cell cycle inhibitor. (A) Cell counting of viable cells of KD-CD or Over CD or a mix of both at the ratio indicated, in the absence or the presence of EGF. Medium was renewed and EGF added every 24 h. The cells were cultivated for 24, 48 and 72 h. (B,C) At the beginning of the experiment (time 0), cells were fixed and stained for CD to confirm the composition of co-cultures. Immunofluorescence double staining of CD and Ki-67 or p27 was performed on SH-SY5Y KD-CD, Over CD and mixed cultures at different ratios: 50% KD-CD + 50% Over CD cells (1:1), 25% KD-CD + 75% Over CD (1:3) and 75% KD-CD + 25% Over CD cells (3:1). Fresh medium was replaced every day, and EGF was added as indicated. After 72 h of treatment, cells were fixed and stained for Ki-67 (green)/CD (red) (B) and p27 (green)/CD (red) (C). Scale bar = 20 µm; magnification = 63×. Representative images of three independent experiments are shown. (D–F) Percentages of CD negative and CD positive cells relative to the total cell population, calculated in random fields, are shown in the graphs. Two-way ANOVA test was performed in all the statistical analysis reported in the graphs. Significance was considered as follows: **** p < 0.0001; *** p < 0.001; ** p < 0.01; * p < 0.05. “Co” refers to untreated cells (control).
![Cancers 16 01343 g007]()
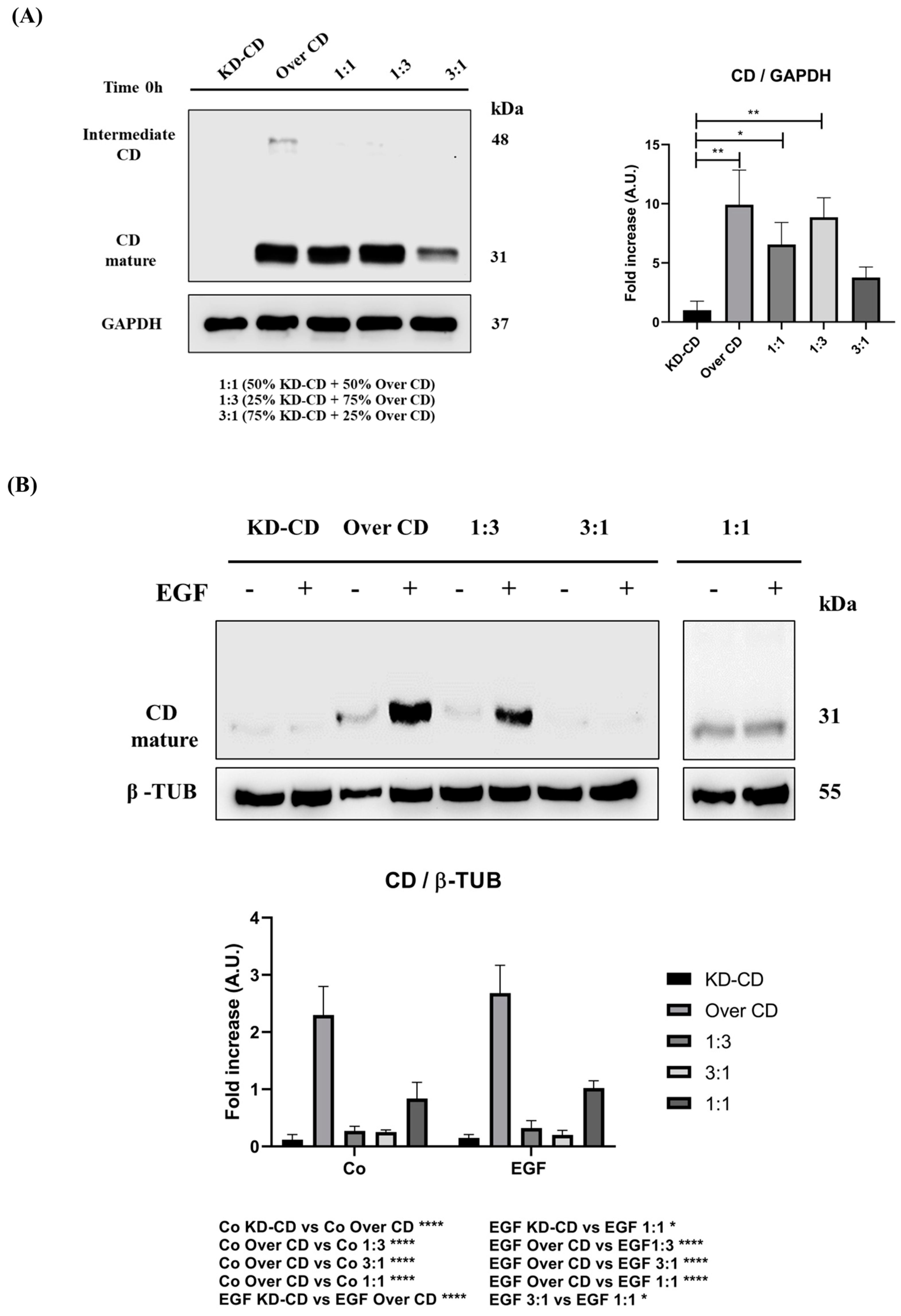
Figure 8.
Analysis of cathepsin D content in SH-SY5Y mixed clones. Western blotting showing the expression of cathepsin D in pure cultures (KD-CD and Over CD) and in mixed co-cultures: 50% KD-CD + 50% Over CD cells (1:1), 25% KD-CD + 75% Over CD (1:3) and 75% KD-CD + 25% Over CD cells (3:1). (A) Samples were collected at time 0h, before EGF treatment, and processed for Western blot analysis of CD expression. One-way ANOVA test was performed. (B,C) Cells were cultured for 72 h, medium was renewed, and EGF re-added every day. Samples were collected after 72 h and processed for Western blot analysis of CD. Membranes were probed with β-actin as loading control. All blots are representative of three independent experiments. Densitometry of the bands is reported in the histograms. Two-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; ** p < 0.01; * p < 0.05. “Co” refers to untreated cells (control).
Figure 8.
Analysis of cathepsin D content in SH-SY5Y mixed clones. Western blotting showing the expression of cathepsin D in pure cultures (KD-CD and Over CD) and in mixed co-cultures: 50% KD-CD + 50% Over CD cells (1:1), 25% KD-CD + 75% Over CD (1:3) and 75% KD-CD + 25% Over CD cells (3:1). (A) Samples were collected at time 0h, before EGF treatment, and processed for Western blot analysis of CD expression. One-way ANOVA test was performed. (B,C) Cells were cultured for 72 h, medium was renewed, and EGF re-added every day. Samples were collected after 72 h and processed for Western blot analysis of CD. Membranes were probed with β-actin as loading control. All blots are representative of three independent experiments. Densitometry of the bands is reported in the histograms. Two-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; ** p < 0.01; * p < 0.05. “Co” refers to untreated cells (control).
Figure 9.
Monitoring of spheroid’s formation in pure and mixed clones co-cultured in the absence or presence of EGF. SH-SY5Y KD-CD, Over CD and co-cultures (50% KD-CD + 50% Over CD cells (1:1), 25% KD-CD + 75% Over CD (1:3) and 75% KD-CD + 25% Over CD cells (3:1)) were plated on non-adherent Petri dishes and allowed to grow for 48 h to allow spheroid formation. Cells were cultured for 7 days after the first treatment. At time 0h, cells were incubated with EGF and re-treated in fresh medium at days 2 and 5 until the endpoint of 7 days. (A) The 3D spheroid’s growth was monitored at the phase-contrast microscope and images were acquired at different time points (time 0 h, 2-, 5- and 7-days). Scale bar = 100 μm; magnification = 20×. (B) Quantification of spheroid’s size was obtained through ImageJ software (v.1.48). The area is indicated as arbitrary unit (A.U.). Data represent the average ± S.D., calculated for at least 5 to 10 spheroids for each condition in three separate experiments. Graphs show the statistically significant differences in spheroid area detected at time day 7 (endpoint) normalized at time 0 h. One-way ANOVA test was performed. Significance was considered as follows: * p < 0.05. (C) Graphs representing the increasing growth of spheroids in control and EGF-treated conditions. The area is indicated as arbitrary unit (A.U.).
Figure 9.
Monitoring of spheroid’s formation in pure and mixed clones co-cultured in the absence or presence of EGF. SH-SY5Y KD-CD, Over CD and co-cultures (50% KD-CD + 50% Over CD cells (1:1), 25% KD-CD + 75% Over CD (1:3) and 75% KD-CD + 25% Over CD cells (3:1)) were plated on non-adherent Petri dishes and allowed to grow for 48 h to allow spheroid formation. Cells were cultured for 7 days after the first treatment. At time 0h, cells were incubated with EGF and re-treated in fresh medium at days 2 and 5 until the endpoint of 7 days. (A) The 3D spheroid’s growth was monitored at the phase-contrast microscope and images were acquired at different time points (time 0 h, 2-, 5- and 7-days). Scale bar = 100 μm; magnification = 20×. (B) Quantification of spheroid’s size was obtained through ImageJ software (v.1.48). The area is indicated as arbitrary unit (A.U.). Data represent the average ± S.D., calculated for at least 5 to 10 spheroids for each condition in three separate experiments. Graphs show the statistically significant differences in spheroid area detected at time day 7 (endpoint) normalized at time 0 h. One-way ANOVA test was performed. Significance was considered as follows: * p < 0.05. (C) Graphs representing the increasing growth of spheroids in control and EGF-treated conditions. The area is indicated as arbitrary unit (A.U.).
![Cancers 16 01343 g009]()
Figure 10.
SH-SY5Y Over CD shows a greater ability to grow in suspension compared to KD-CD cells. SH-SY5Y 3D spheroids of pure clones or clones mixed at the indicated ratios were cultured for 7 days in the absence or presence of EGF. Cell homogenates were assayed by Western blotting for CD at time 0 (A) and after 7 days (B). The filters were stripped and re-probed for GAPDH (A) or β-tubulin (B) as loading control. Densitometry of the bands is reported in the histograms. One-way ANOVA and two-way ANOVA tests were performed in (A) and (B), respectively. Significance was considered as follows: **** p < 0.0001; ** p < 0.01; * p < 0.05. “Co” refers to untreated cells (control).
Figure 10.
SH-SY5Y Over CD shows a greater ability to grow in suspension compared to KD-CD cells. SH-SY5Y 3D spheroids of pure clones or clones mixed at the indicated ratios were cultured for 7 days in the absence or presence of EGF. Cell homogenates were assayed by Western blotting for CD at time 0 (A) and after 7 days (B). The filters were stripped and re-probed for GAPDH (A) or β-tubulin (B) as loading control. Densitometry of the bands is reported in the histograms. One-way ANOVA and two-way ANOVA tests were performed in (A) and (B), respectively. Significance was considered as follows: **** p < 0.0001; ** p < 0.01; * p < 0.05. “Co” refers to untreated cells (control).
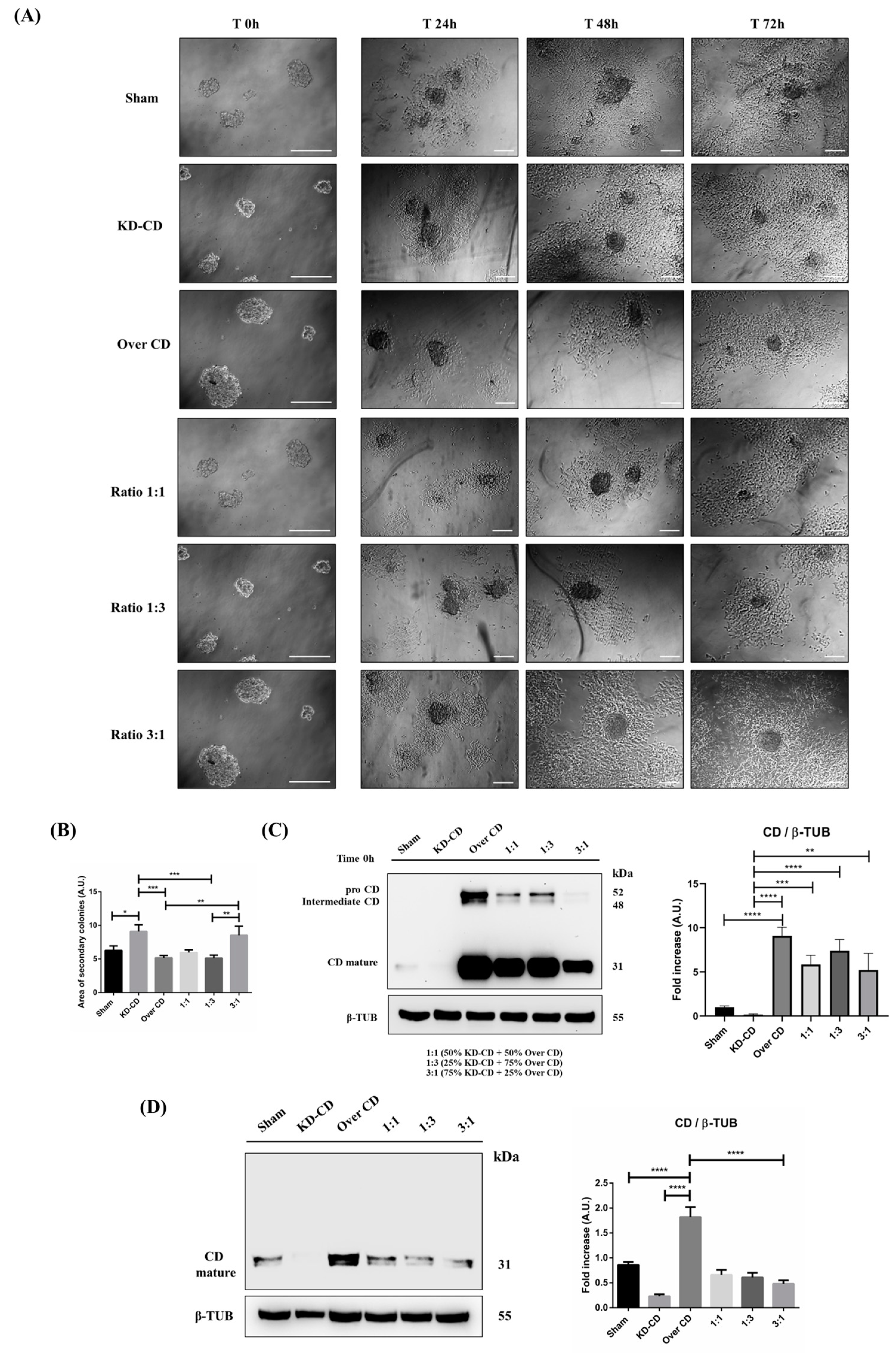
Figure 11.
CD knocked-down SH-SY5Y cells rescue the ability to grow in the adherent condition. SH-SY5Y Sham, KD-CD, Over CD and mixed co-cultures, 50% KD-CD + 50% Over CD cells (1:1), 25% KD-CD + 75% Over CD (1:3) and 75% KD-CD + 25% Over CD cells (3:1) were seeded in non-adherent Petri dishes and allowed to grow for 72 h to allow spheroid formation (500,000 cells/Petri). On the third day (indicated in Figure as Time 0 h), neurospheres were collected, resuspended in fresh medium, plated in adherent Petri dishes and maintained in culture for a further 72 h. New fresh medium was replaced every day. Cell homogenates were processed for Western blot analysis. (A) Images were acquired at the phase-contrast microscope every day to monitor cell attachment and growth. Scale bar = 100 μm; magnification = 20× (T0 h), 5× (T24 h, 48 h, 72 h). (B) Graph representing the area of secondary colonies calculated in different representative fields of three separate experiments (at the endpoint of 72 h). One-way ANOVA test was performed. Significance was considered as follows: *** p < 0.001; ** p < 0.01; * p < 0.05. (C,D) Western blot analysis of CD in cell homogenates collected at time 0 (C) and after 72 h (D). The membranes were stripped and re-probed for β-tubulin as loading control. The blots are representative of three independent experiments. Densitometry of the bands is reported in the histograms. One-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; *** p < 0.001; ** p < 0.01.
Figure 11.
CD knocked-down SH-SY5Y cells rescue the ability to grow in the adherent condition. SH-SY5Y Sham, KD-CD, Over CD and mixed co-cultures, 50% KD-CD + 50% Over CD cells (1:1), 25% KD-CD + 75% Over CD (1:3) and 75% KD-CD + 25% Over CD cells (3:1) were seeded in non-adherent Petri dishes and allowed to grow for 72 h to allow spheroid formation (500,000 cells/Petri). On the third day (indicated in Figure as Time 0 h), neurospheres were collected, resuspended in fresh medium, plated in adherent Petri dishes and maintained in culture for a further 72 h. New fresh medium was replaced every day. Cell homogenates were processed for Western blot analysis. (A) Images were acquired at the phase-contrast microscope every day to monitor cell attachment and growth. Scale bar = 100 μm; magnification = 20× (T0 h), 5× (T24 h, 48 h, 72 h). (B) Graph representing the area of secondary colonies calculated in different representative fields of three separate experiments (at the endpoint of 72 h). One-way ANOVA test was performed. Significance was considered as follows: *** p < 0.001; ** p < 0.01; * p < 0.05. (C,D) Western blot analysis of CD in cell homogenates collected at time 0 (C) and after 72 h (D). The membranes were stripped and re-probed for β-tubulin as loading control. The blots are representative of three independent experiments. Densitometry of the bands is reported in the histograms. One-way ANOVA test was performed. Significance was considered as follows: **** p < 0.0001; *** p < 0.001; ** p < 0.01.
![Cancers 16 01343 g011]()
Figure 12.
IMR-32 cells expressing low CD levels show a greater ability to grow in the adherent condition. IMR-32, LAN-5 and SK-N-BE(2) cells were seeded in non-adherent Petri dishes and allowed to grow for 72 h to allow spheroid formation (500,000 cells/Petri). On the third day (indicated in Figure as Time 0 h), neurospheres were collected, resuspended in fresh medium, plated in adherent Petri dishes and maintained in culture for a further 72 h. New fresh medium was replaced every day. Cell homogenates were processed for Western blot analysis. (A) Images were acquired at the phase-contrast microscope every day to monitor cell attachment and growth. Scale bar = 100 μm; magnification = 20× (T0 h), 5× (T24 h, 48 h, 72 h). (B) Graph representing the area of secondary colonies calculated in different representative fields of three separate experiments (at the endpoint of 72 h). One-way ANOVA test was performed. Significance was considered as follows: *** p < 0.001. (C,D) Western blot analysis of CD expression in cell homogenates collected at time 0 (C) and after 72 h (D). The membranes were re-probed for β-tubulin as loading control. The blots are representative of three independent experiments. Densitometry of the bands is reported in the histograms. One-way ANOVA test was performed. Significance was considered as follows: ** p < 0.01; * p < 0.05.
Figure 12.
IMR-32 cells expressing low CD levels show a greater ability to grow in the adherent condition. IMR-32, LAN-5 and SK-N-BE(2) cells were seeded in non-adherent Petri dishes and allowed to grow for 72 h to allow spheroid formation (500,000 cells/Petri). On the third day (indicated in Figure as Time 0 h), neurospheres were collected, resuspended in fresh medium, plated in adherent Petri dishes and maintained in culture for a further 72 h. New fresh medium was replaced every day. Cell homogenates were processed for Western blot analysis. (A) Images were acquired at the phase-contrast microscope every day to monitor cell attachment and growth. Scale bar = 100 μm; magnification = 20× (T0 h), 5× (T24 h, 48 h, 72 h). (B) Graph representing the area of secondary colonies calculated in different representative fields of three separate experiments (at the endpoint of 72 h). One-way ANOVA test was performed. Significance was considered as follows: *** p < 0.001. (C,D) Western blot analysis of CD expression in cell homogenates collected at time 0 (C) and after 72 h (D). The membranes were re-probed for β-tubulin as loading control. The blots are representative of three independent experiments. Densitometry of the bands is reported in the histograms. One-way ANOVA test was performed. Significance was considered as follows: ** p < 0.01; * p < 0.05.
![Cancers 16 01343 g012]()
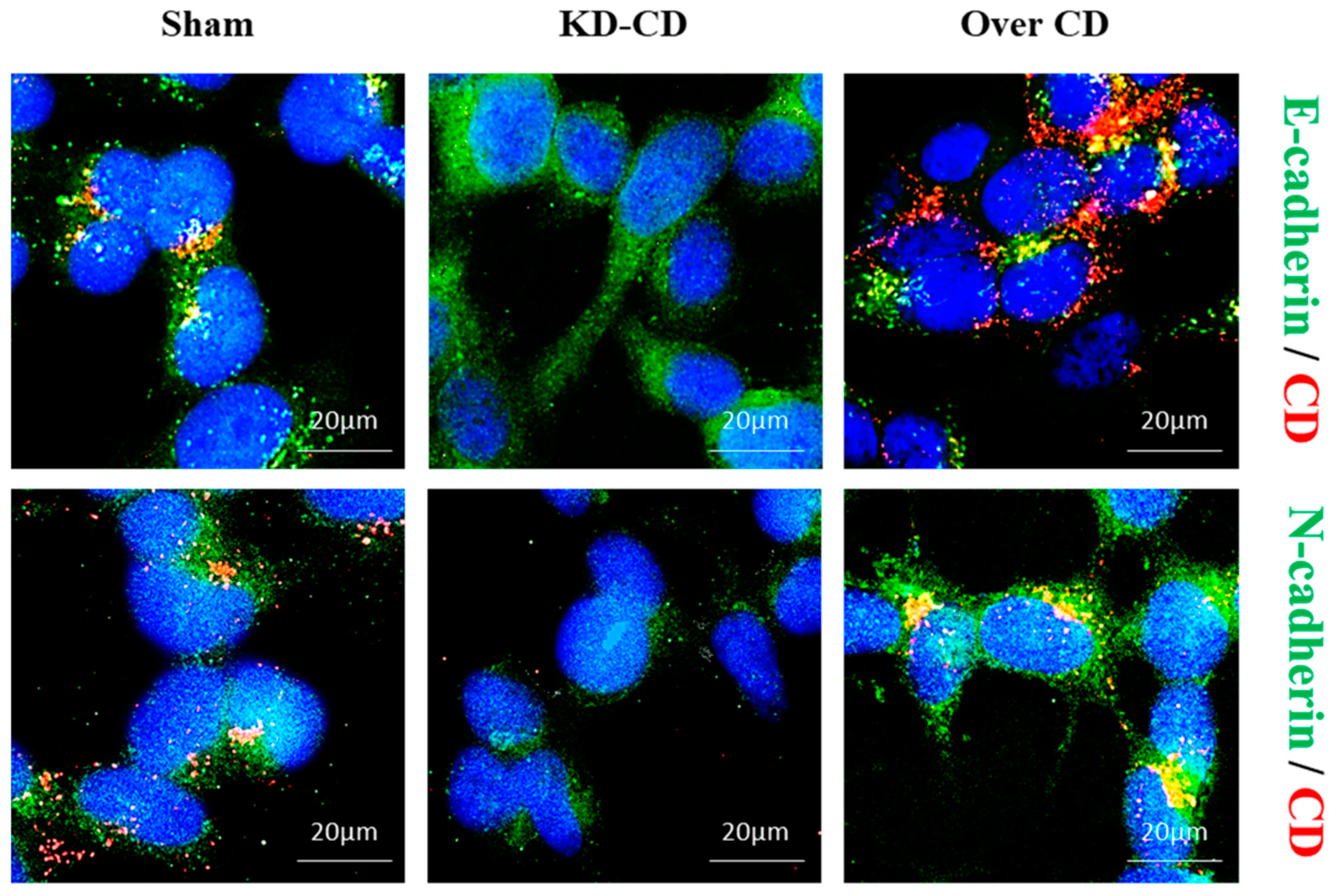
Figure 13.
Low expression of Cathepsin D promotes the acquisition of an epithelial phenotype.Sham, KD-CD and Over CD clones were seeded in 3D cultures and allowed to grow for 4 days. Later, 3D spheroids were transferred on to poly-lysine-coated coverslips and allowed to grow for a further 2 days. At the end of the experiment, coverslips were fixed and stained for immunofluorescence double staining of CD (red)–E-cadherin (green) or CD (red)–N-cadherin (green). Nuclei were stained with DAPI. Scale bar = 20 µm; magnification = 63×. Representative images of three independent experiments are shown.
Figure 13.
Low expression of Cathepsin D promotes the acquisition of an epithelial phenotype.Sham, KD-CD and Over CD clones were seeded in 3D cultures and allowed to grow for 4 days. Later, 3D spheroids were transferred on to poly-lysine-coated coverslips and allowed to grow for a further 2 days. At the end of the experiment, coverslips were fixed and stained for immunofluorescence double staining of CD (red)–E-cadherin (green) or CD (red)–N-cadherin (green). Nuclei were stained with DAPI. Scale bar = 20 µm; magnification = 63×. Representative images of three independent experiments are shown.
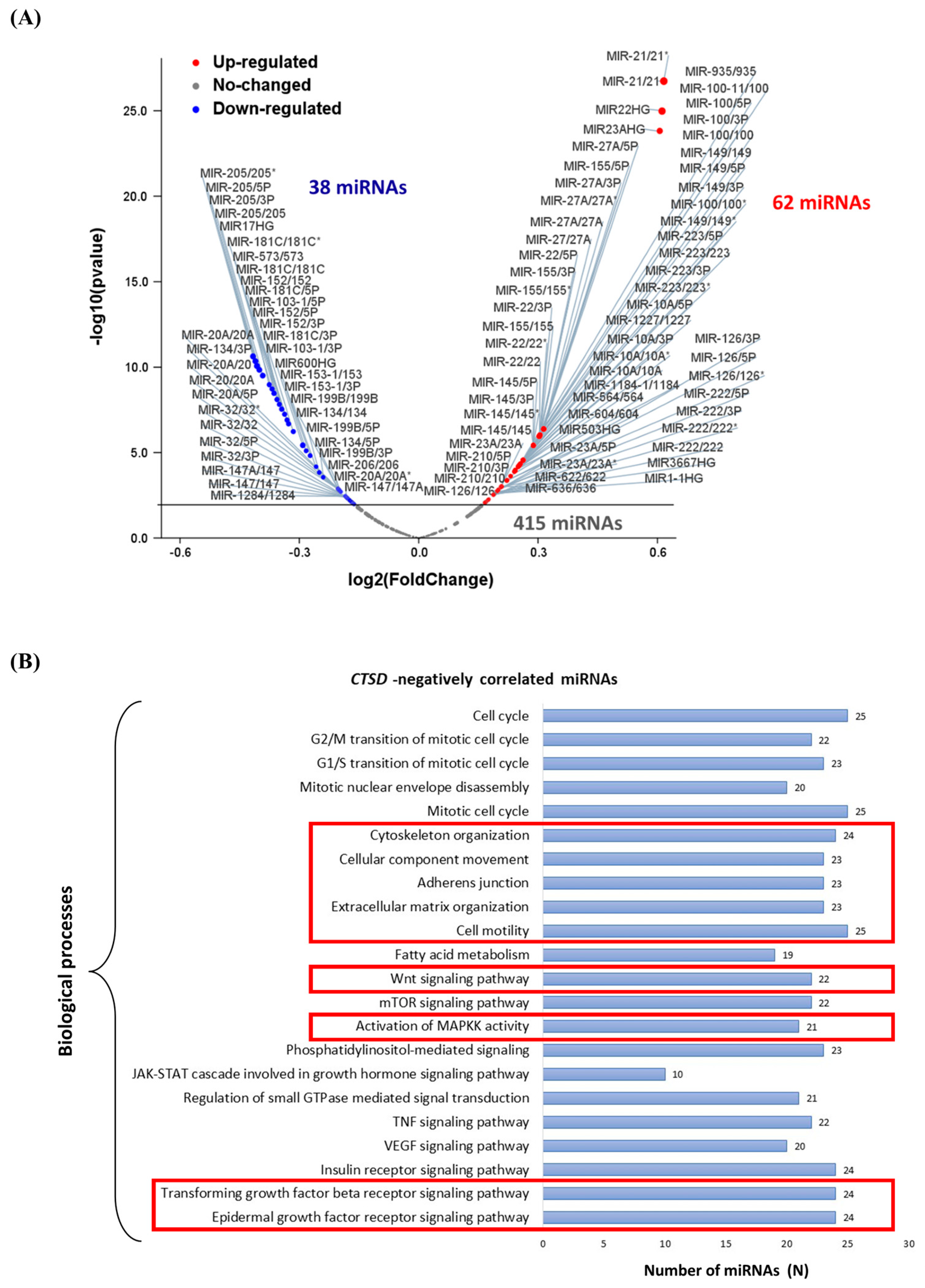
Figure 14.
CTSD negatively correlated microRNA-modulated biological processes and pathways. (A) Volcano plot displaying the differentially expressed miRNAs. Red dots represent CTSD positively correlated miRNAs, while blue dots represent CTSD negatively correlated miRNAs. (B) Graph reporting the CTSD negatively correlated miRNAs biological processes.
Figure 14.
CTSD negatively correlated microRNA-modulated biological processes and pathways. (A) Volcano plot displaying the differentially expressed miRNAs. Red dots represent CTSD positively correlated miRNAs, while blue dots represent CTSD negatively correlated miRNAs. (B) Graph reporting the CTSD negatively correlated miRNAs biological processes.
Figure 15.
The cartoon summarizes in graphical form the findings here reported.
Figure 15.
The cartoon summarizes in graphical form the findings here reported.



 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}