

Pathogenesis of Warthin’s Tumor: Neoplastic or Non-Neoplastic?


{kind=link}
{kind=link}
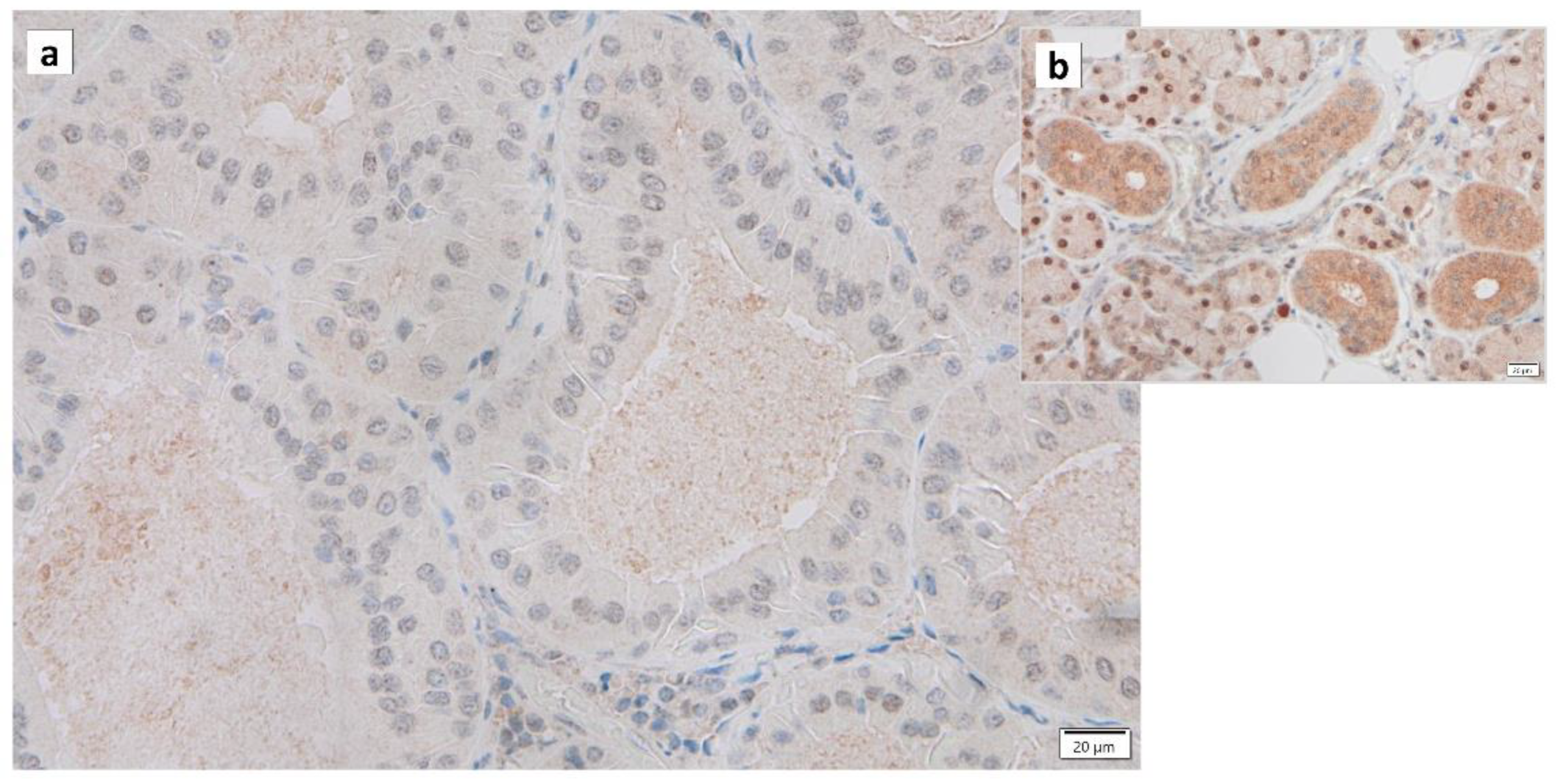{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
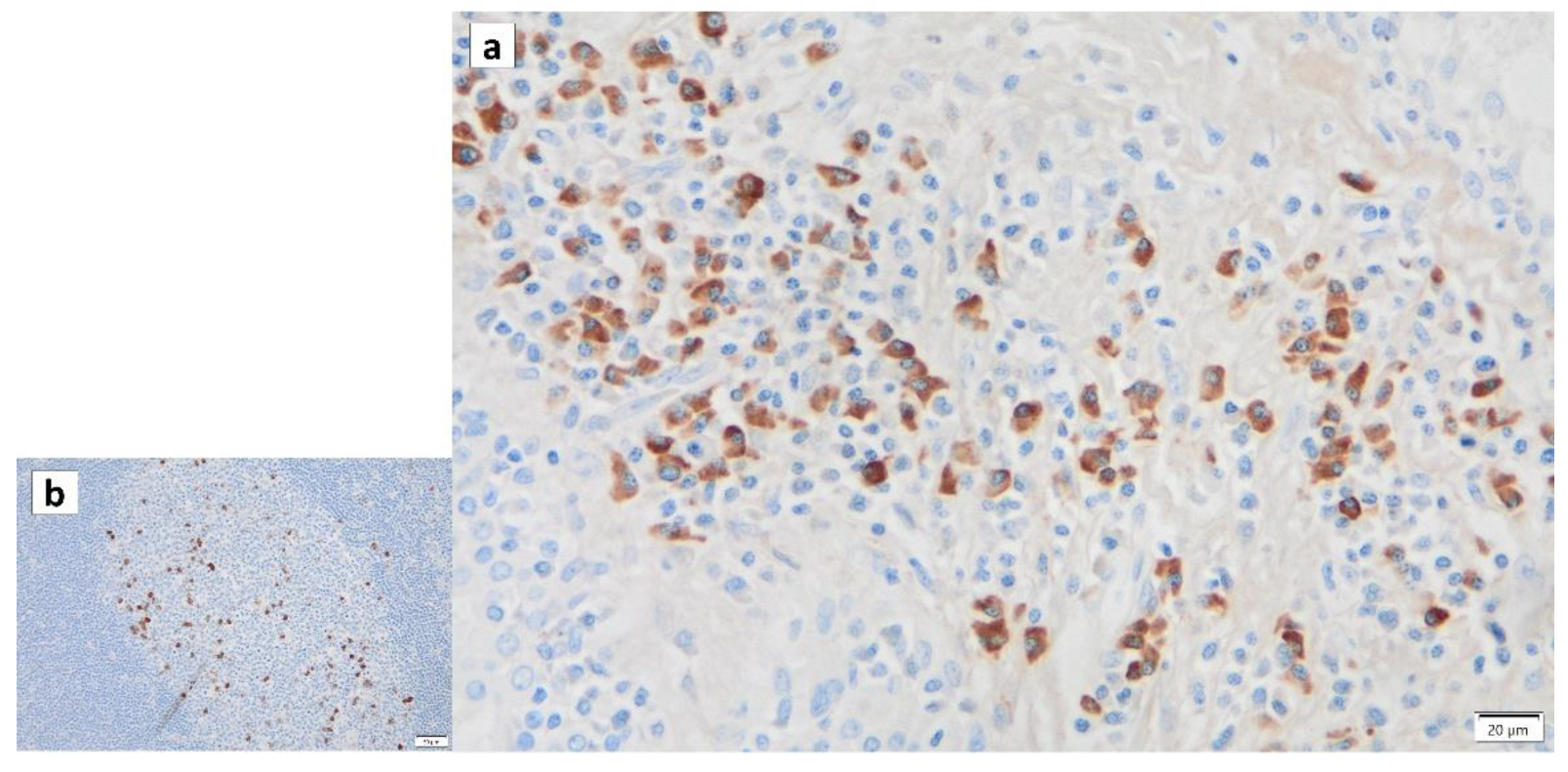{kind=link}
{kind=link}
{kind=link}
{kind=link}
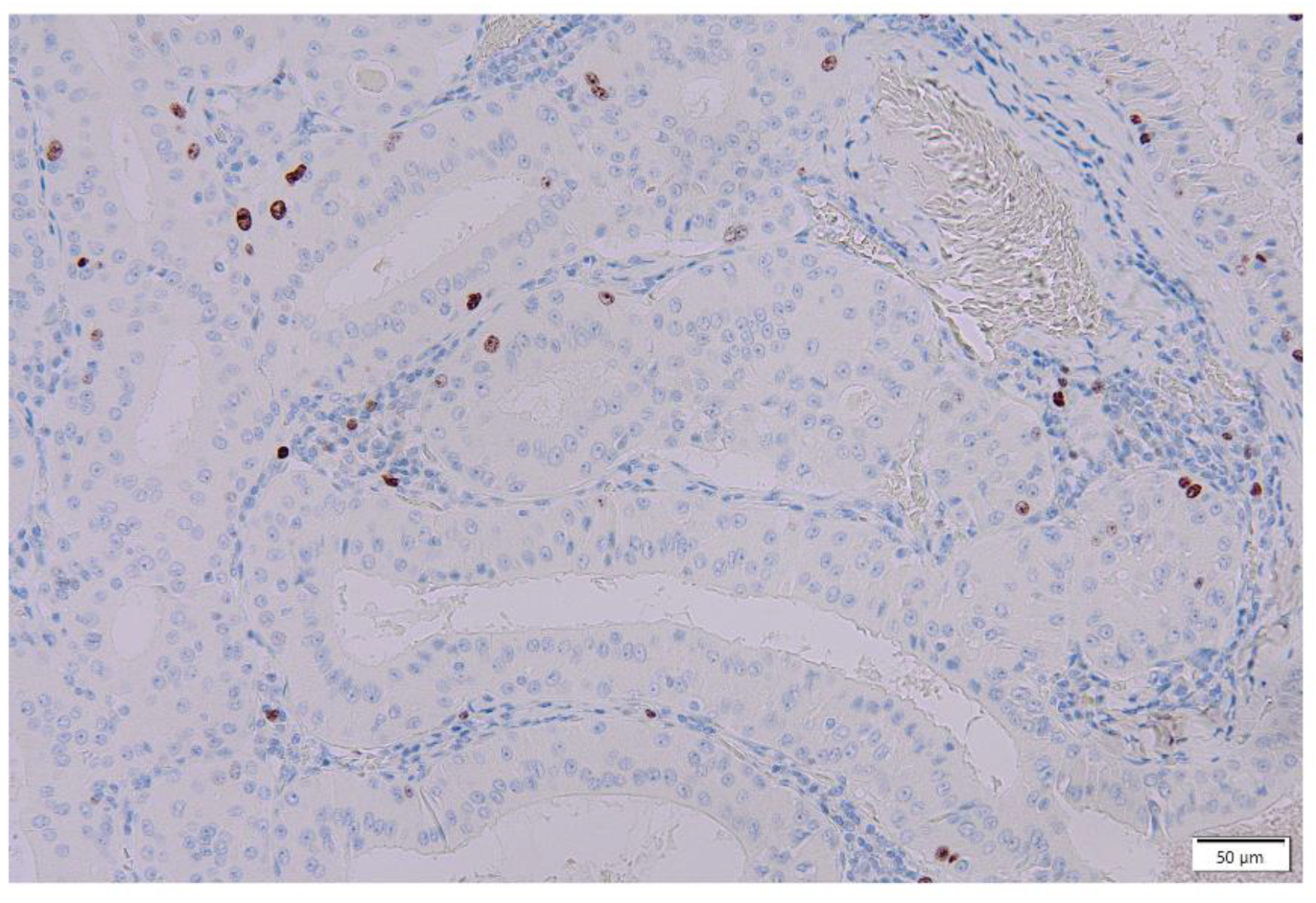{kind=link}
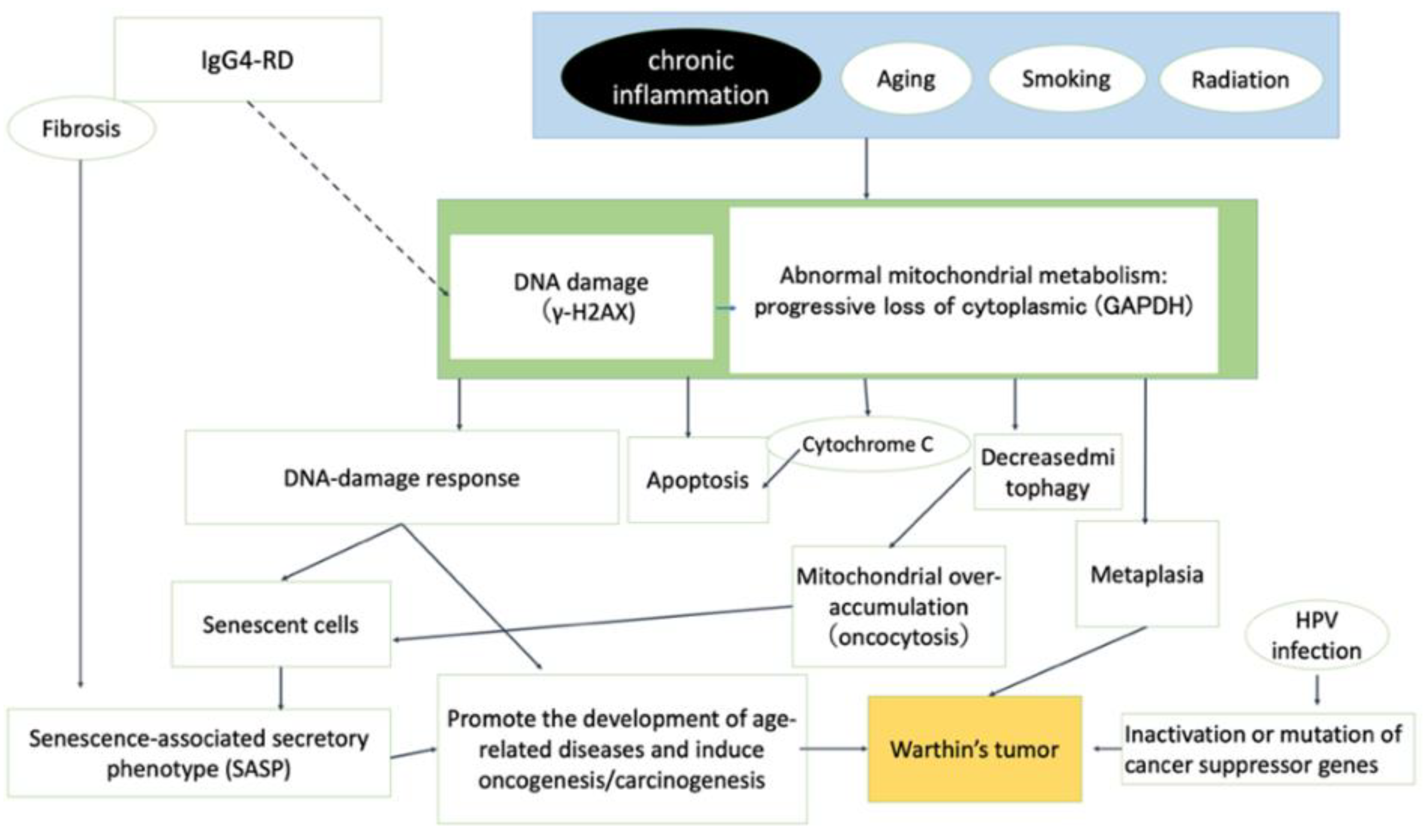{kind=link}
Simple Summary
Abstract
1. Introduction
2. Anatomy/Histology of the Salivary Glands
3. Histopathology of Warthin’s Tumor
4. Cytology of Warthin’s Tumor
5. Risk Factors for Warthin’s Tumor
6. Association between SASP and Warthin’s Tumor
7. Mitochondrial Dysfunction and Warthin’s Tumor
8. IgG4 and Warthin’s Tumor
9. The Role of GAPDH in Warthin’s Tumor Cells
10. The Role of p16 and p53 in Warthin’s Tumor
11. HPV Infection and Warthin’s Tumor
12. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Teymoortash, A.; Krasnewicz, Y.; Werner, J.A. Clinical features of cystadenolymphoma (Warthin’s tumor) of the parotid gland: A retrospective comparative study of 96 cases. Oral Oncol. 2006, 42, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Kuzenko, Y.V.; Romanuk, A.M.; Dyachenko, O.O.; Hudymenko, O. Pathogenesis of Warthin’s tumors. Interv. Med. Appl. Sci. 2016, 8, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Franzen, A.M.; Kaup Franzen, C.; Guenzel, T.; Lieder, A. Increased incidence of Warthin tumours of the parotid gland: A 42-year evaluation. Eur. Arch. Otorhinolaryngol. 2018, 275, 2593–2598. [Google Scholar] [CrossRef] [PubMed]
- Schwalje, A.T.; Uzelac, A.; Ryan, W.R. Growth rate characteristics of Warthin’s tumours of the parotid gland. Int. J. Oral Maxillofac. Surg. 2015, 44, 1474–1479. [Google Scholar] [CrossRef] [PubMed]
- Teymoortash, A.; Schrader, C.; Shimoda, H.; Kato, S.; Werner, J.A. Evidence of lymphangiogenesis in Warthin’s tumor of the parotid gland. Oral Oncol. 2007, 43, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Karthikeya, P.; Mahima, V.G.; Shalini, K. Papillary cystadenoma lymphomatosum: Case report and review of literature. Indian J. Dent. Res. 2005, 16, 153–158. [Google Scholar]
- Limaiem, F.; Jain, P. Warthin Tumor; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- Ning, C.; Koo, J.S.; Kim, E.K.; Lee, S. Clinical and sonographic characteristics of Warthin-like variant papillary thyroid carcinomas. Med. Ultrason. 2019, 21, 152–157. [Google Scholar] [CrossRef]
- Wu, S.; Liu, G.; Chen, R.; Guan, Y. Role of ultrasound in the assessment of benignity and malignancy of parotid masses. Dentomaxillofac Radiol. 2012, 41, 131–135. [Google Scholar] [CrossRef]
- Grandis, A.; El-Naggar, J.; Chan, J.; Takata, T.S.; Slootweg, P.J. WHO Classification of Head and Neck Tumours; IARC: Lyon, France, 2017; p. 85. [Google Scholar]
- Quer, M.; Hernandez-Prera, J.C.; Silver, C.E.; Casasayas, M.; Simo, R.; Vander Poorten, V.; Guntinas-Lichius, O.; Bradley, P.J.; Tong-Ng, W.; Rodrigo, J.P.; et al. Current Trends and Controversies in the Management of Warthin Tumor of the Parotid Gland. Diagnostics 2021, 11, 1467. [Google Scholar] [CrossRef]
- Orabona, G.D.; Abbate, V.; Piombino, P.; Romano, A.; Schonauer, F.; Iaconetta, G.; Salzano, G.; Farina, F.; Califano, L. Warthin’s tumour: Aetiopathogenesis dilemma, ten years of our experience. J. Craniomaxillofac Surg. 2015, 43, 427–431. [Google Scholar] [CrossRef]
- Daguci, L.; Simionescu, C.; Daguci, C.; Bataiosu, M.; Dragomir, L.P. Clinical and Morphological Aspects of Warthin’s Tumor. Curr. Health Sci. J. 2009, 35, 115–118. [Google Scholar]
- Bishop, J.A.; Thompson, L.D.R.; Paul EWakely, J.; Weinreb, I. Tumors of the Salivary Glands; AFIP Atlas of Tumor & Non-Tumor Pathology, 5th Series, Fascicle 5; American Registry of Pathology: Silver Spring, MD, USA, 2021. [Google Scholar]
- Barnes, L.; Eveson, J.; Reichart, P.; Sidransky, D. World Health Organization Classification of Tumours: Pathology and Genetics of Head and Neck Tumours; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Tandler, B.; Shipkey, F.H. Ultrastructure of Warthin’s Tumor. I. Mitochondria. J. Ultrastruct. Res. 1964, 11, 292–305. [Google Scholar] [CrossRef]
- Hartwick, R.W.; Batsakis, J.G. Non-Warthin’s tumor oncocytic lesions. Ann. Otol. Rhinol. Laryngol. 1990, 99, 674–677. [Google Scholar] [CrossRef]
- Mandic, R.; Agaimy, A.; Pinto-Quintero, D.; Roth, K.; Teymoortash, A.; Schwarzbach, H.; Stoehr, C.G.; Rodepeter, F.R.; Stuck, B.A.; Bette, M. Aberrant Expression of Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) in Warthin Tumors. Cancers 2020, 12, 1112. [Google Scholar] [CrossRef]
- Capone, R.B.; Ha, P.K.; Westra, W.H.; Pilkington, T.M.; Sciubba, J.J.; Koch, W.M.; Cummings, C.W. Oncocytic neoplasms of the parotid gland: A 16-year institutional review. Otolaryngol. Head. Neck Surg. 2002, 126, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Soares, P.; Maximo, V.; Samuels, D.C. Somatic mitochondrial DNA mutations in cancer escape purifying selection and high pathogenicity mutations lead to the oncocytic phenotype: Pathogenicity analysis of reported somatic mtDNA mutations in tumors. BMC Cancer 2012, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Bengoechea, O.; Sanchez, F.; Larrinaga, B.; Martinez-Penuela, J.M. Oncocytic adenocarcinoma arising in Warthin’s tumor. Pathol. Res. Pract. 1989, 185, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Alnoor, F.; Gandhi, J.S.; Stein, M.K.; Gradowski, J.F. Follicular Lymphoma Diagnosed in Warthin Tumor: A Case Report and Review of the Literature. Head. Neck Pathol. 2020, 14, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Gorai, S.; Numata, T.; Kawada, S.; Nakano, M.; Tamaru, J.; Kobayashi, T. Malignant lymphoma arising from heterotopic Warthin’s tumor in the neck: Case report and review of the literature. Tohoku J. Exp. Med. 2007, 212, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Smolka, W.; Markowski, J.; Piotrowska-Seweryn, A.; Palen, P.; Dobrosz, Z.; Dziubdziela, W. Mucoepidermoid carcinoma in Warthin tumor of the parotid gland. Arch. Med. Sci. 2015, 11, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.D.; Simmons, B.H.; el-Naggar, A.; Medeiros, L.J. Mucoepidermoid carcinoma involving Warthin tumor. A report of five cases and review of the literature. Am. J. Clin. Pathol. 2000, 114, 564–570. [Google Scholar] [CrossRef]
- Yu, C.; Song, Z.; Xiao, Z.; Lin, Q.; Dong, X. Mucoepidermoid carcinoma arising in Warthin’s tumor of the parotid gland: Clinicopathological characteristics and immunophenotypes. Sci. Rep. 2016, 6, 30149. [Google Scholar] [CrossRef]
- Yaranal, P.J.; Umashankar, T. Squamous Cell Carcinoma Arising in Warthin’s Tumour: A Case Report. J. Clin. Diagn. Res. 2013, 7, 163–165. [Google Scholar] [CrossRef]
- Banik, S.; Howell, J.S.; Wright, D.H. Non-Hodgkin’s lymphoma arising in adenolymphoma—A report of two cases. J. Pathol. 1985, 146, 167–177. [Google Scholar] [CrossRef]
- Kim, J.E.; Kim, T.G. Squamous cell carcinoma arising from Warthin’s tumor in the parotid gland. BJR Case Rep. 2019, 5, 20190032. [Google Scholar] [CrossRef]
- Mohapatra, M.; Satyanarayana, S. Low grade mucoepidermoid carcinoma in a setting of Warthin’s tumor. Indian J. Pathol. Microbiol. 2012, 55, 392–394. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-C.; Li, C.-F.; Lin, C.-N. Unreported Cytologic Characteristics of Oncocytes in Warthin’s Tumors. Tzu Chi Med. J. 2010, 22, 137–140. [Google Scholar] [CrossRef]
- Koybasioglu, F.F.; Onal, B.; Han, U.; Adabag, A.; Sahpaz, A. Cytomorphological findings in diagnosis of Warthin tumor. Turk. J. Med. Sci. 2020, 50, 148–154. [Google Scholar]
- Ballo, M.S.; Shin, H.J.; Sneige, N. Sources of diagnostic error in the fine-needle aspiration diagnosis of Warthin’s tumor and clues to a correct diagnosis. Diagn. Cytopathol. 1997, 17, 230–234. [Google Scholar] [CrossRef]
- Das, D.K.; Petkar, M.A.; Al-Mane, N.M.; Sheikh, Z.A.; Mallik, M.K.; Anim, J.T. Role of fine needle aspiration cytology in the diagnosis of swellings in the salivary gland regions: A study of 712 cases. Med. Princ. Pract. 2004, 13, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.; Ronen, O. Cytologic diagnosis of parotid gland Warthin tumor: Systematic review and meta-analysis. Head Neck 2022, 44, 2277–2287. [Google Scholar] [CrossRef] [PubMed]
- Zahran, M.; Alsedra, S.; Cope, D.; Youssef, A. The Role of FNAC in the Diagnosis and Management of Warthin Tumour: Analysis of 74 Cases. Int. Arch. Otorhinolaryngol. 2021, 25, e379–e382. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.T.; Sugihara, K.; Kawashima, K.; Yamashita, S. Scanning electron microscopic study of Warthin’s tumor. J. Oral. Pathol. 1987, 16, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, S.; Chen, F.; Aguirre, A. Oncocytoma of the Parotid Gland and its Mimickers: A Comprehensive Review. N. Am. J. Med. Sci. 2010, 3, 171. [Google Scholar] [CrossRef]
- el-Hossary, N.M.; Fathy, L.M. Luminal epithelium of Warthin’s tumours: A scanning electron microscopic study. Egypt Dent. J. 1994, 40, 791–794. [Google Scholar] [PubMed]
- Seifert, G. Primary salivary gland tumors in lymph nodes of the parotid gland. Report of 3 cases and review of the literature. Pathologe 1997, 18, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.M.; Hsu, P.L.; Nayak, R.N. Warthin’s tumor: An immunohistochemical study of its lymphoid stroma. Hum. Pathol. 1981, 12, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.M.; Raine, L. Warthin’s tumor—Epithelial cell differences. Am. J. Clin. Pathol. 1982, 77, 78–82. [Google Scholar] [CrossRef]
- Peter Klussmann, J.; Wittekindt, C.; Florian Preuss, S.; Al Attab, A.; Schroeder, U.; Guntinas-Lichius, O. High risk for bilateral Warthin tumor in heavy smokers—Review of 185 cases. Acta Otolaryngol. 2006, 126, 1213–1217. [Google Scholar] [CrossRef]
- Saku, T.; Hayashi, Y.; Takahara, O.; Matsuura, H.; Tokunaga, M.; Tokuoka, S.; Soda, M.; Mabuchi, K.; Land, C.E. Salivary gland tumors among atomic bomb survivors, 1950–1987. Cancer 1997, 79, 1465–1475. [Google Scholar] [CrossRef]
- Gallo, O.; Bocciolini, C. Warthin’s tumour associated with autoimmune diseases and tobacco use. Acta Otolaryngol. 1997, 117, 623–627. [Google Scholar] [CrossRef]
- Rabia, A.B.; Ebru, L.S.; Tuba, K.; Didar, G.; Gulhan, O.; Cengiz, O. Warthin’s tumor: An unknown pathogenesis: A neoplasm or a reactive hyperplasia? Indian J. Pathol. Microbiol. 2015, 58, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Huhns, M.; Simm, G.; Erbersdobler, A.; Zimpfer, A. HPV Infection, but Not EBV or HHV-8 Infection, Is Associated with Salivary Gland Tumours. Biomed. Res. Int. 2015, 2015, 829349. [Google Scholar] [CrossRef]
- Aga, M.; Kondo, S.; Yamada, K.; Sawada-Kitamura, S.; Yagi-Nakanishi, S.; Endo, K.; Wakisaka, N.; Murono, S.; Kawano, M.; Yoshizaki, T. Warthin’s tumor associated with IgG4-related disease. Auris Nasus Larynx. 2013, 40, 514–517. [Google Scholar] [CrossRef]
- Hayflick, L. The Limited in Vitro Lifetime of Human Diploid Cell Strains. Exp. Cell Res. 1965, 37, 614–636. [Google Scholar] [CrossRef]
- Campisi, J.; d’Adda di Fagagna, F. Cellular senescence: When bad things happen to good cells. Nat. Rev. Mol. Cell Biol. 2007, 8, 729–740. [Google Scholar] [CrossRef]
- Engeland, K. Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ. 2022, 29, 946–960. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.D.; Wagner, N. The Senescence Markers p16INK4A, p14ARF/p19ARF, and p21 in Organ Development and Homeostasis. Cells 2022, 11, 1966. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Kawamoto, S.; Ohtani, N.; Hara, E. Impact of senescence-associated secretory phenotype and its potential as a therapeutic target for senescence-associated diseases. Cancer Sci. 2017, 108, 563–569. [Google Scholar] [CrossRef]
- Birch, J.; Gil, J. Senescence and the SASP: Many therapeutic avenues. Genes Dev. 2020, 34, 1565–1576. [Google Scholar] [CrossRef]
- Terlecki-Zaniewicz, L.; Lammermann, I.; Latreille, J.; Bobbili, M.R.; Pils, V.; Schosserer, M.; Weinmüllner, R.; Dellago, H.; Skalicky, S.; Pum, D.; et al. Small extracellular vesicles and their miRNA cargo are anti-apoptotic members of the senescence-associated secretory phenotype. Aging 2018, 10, 1103–1132. [Google Scholar] [CrossRef] [PubMed]
- Lecot, P.; Alimirah, F.; Desprez, P.Y.; Campisi, J.; Wiley, C. Context-dependent effects of cellular senescence in cancer development. Br. J. Cancer 2016, 114, 1180–1184. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J. Aging, cellular senescence, and cancer. Annu. Rev. Physiol. 2013, 75, 685–705. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Aoki, R.; Terasaki, M. Potential Chemopreventive Effects of Dietary Combination of Phytochemicals against Cancer Development. Pharmaceuticals 2023, 16, 1591. [Google Scholar] [CrossRef]
- Krtolica, A.; Parrinello, S.; Lockett, S.; Desprez, P.Y.; Campisi, J. Senescent fibroblasts promote epithelial cell growth and tumorigenesis: A link between cancer and aging. Proc. Natl. Acad. Sci. USA 2001, 98, 12072–12077. [Google Scholar] [CrossRef]
- Yasuda, T.; Koiwa, M.; Yonemura, A.; Miyake, K.; Kariya, R.; Kubota, S.; Yokomizo-Nakano, T.; Yasuda-Yoshihara, N.; Uchihara, T.; Itoyama, R.; et al. Inflammation-driven senescence-associated secretory phenotype in cancer-associated fibroblasts enhances peritoneal dissemination. Cell Rep. 2021, 34, 108779. [Google Scholar] [CrossRef]
- Ou, H.L.; Hoffmann, R.; Gonzalez-Lopez, C.; Doherty, G.J.; Korkola, J.E.; Munoz-Espin, D. Cellular senescence in cancer: From mechanisms to detection. Mol. Oncol. 2021, 15, 2634–2671. [Google Scholar] [CrossRef]
- Fane, M.; Weeraratna, A.T. How the ageing microenvironment influences tumour progression. Nat. Rev. Cancer 2020, 20, 89–106. [Google Scholar] [CrossRef]
- Kaur, A.; Webster, M.R.; Weeraratna, A.T. In the Wnt-er of life: Wnt signalling in melanoma and ageing. Br. J. Cancer 2016, 115, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Ishikawa, H. Mast cells and inflammation-associated colorectal carcinogenesis. Semin. Immunopathol. 2013, 35, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Bu, L.; Zhu, H.; Racila, E.; Khaja, S.; Hamlar, D.; Li, F. Xanthogranulomatous sialadenitis, an uncommon reactive change is often associated with Warthin’s tumor. Head Neck Pathol. 2020, 14, 525–553. [Google Scholar] [CrossRef] [PubMed]
- Pedroza-Gonzalez, A.; Xu, K.; Wu, T.C.; Aspord, C.; Tindle, S.; Marches, F.; Gallegos, M.; Burton, E.C.; Savino, D.; Hori, T.; et al. Thymic stromal lymphopoietin fosters human breast tumor growth by promoting type 2 inflammation. J. Exp. Med. 2011, 208, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Traub, B.; Shi, J.; Kornmann, M. Possible Roles of Interleukin-4 and -13 and Their Receptors in Gastric and Colon Cancer. Int. J. Mol. Sci. 2021, 22, 727. [Google Scholar] [CrossRef]
- Coussens, L.M.; Zitvogel, L.; Palucka, A.K. Neutralizing tumor-promoting chronic inflammation: A magic bullet? Science 2013, 339, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Liu, V.W.; Zhang, C.; Nagley, P. Mutations in mitochondrial DNA accumulate differentially in three different human tissues during ageing. Nucleic Acids Res. 1998, 26, 1268–1275. [Google Scholar] [CrossRef] [PubMed]
- Wiley, C.D.; Velarde, M.C.; Lecot, P.; Liu, S.; Sarnoski, E.A.; Freund, A.; Shirakawa, K.; Lim, H.W.; Davis, S.S.; Ramanathan, A.; et al. Mitochondrial Dysfunction Induces Senescence with a Distinct Secretory Phenotype. Cell Metab. 2016, 23, 303–314. [Google Scholar] [CrossRef]
- Fakouri, N.B.; Hou, Y.; Demarest, T.G.; Christiansen, L.S.; Okur, M.N.; Mohanty, J.G.; Croteau, D.L.; Bohr, V.A. Toward understanding genomic instability, mitochondrial dysfunction and aging. FEBS J. 2019, 286, 1058–1073. [Google Scholar] [CrossRef]
- Lu, J.; Sharma, L.K.; Bai, Y. Implications of mitochondrial DNA mutations and mitochondrial dysfunction in tumorigenesis. Cell Res. 2009, 19, 802–815. [Google Scholar] [CrossRef]
- Lewis, P.D.; Baxter, P.; Paul Griffiths, A.; Parry, J.M.; Skibinski, D.O. Detection of damage to the mitochondrial genome in the oncocytic cells of Warthin’s tumour. J. Pathol. 2000, 191, 274–281. [Google Scholar] [CrossRef]
- Gao, S.; Hu, J. Mitochondrial Fusion: The Machineries In and Out. Trends Cell Biol. 2021, 31, 62–74. [Google Scholar] [CrossRef]
- Ferreira-da-Silva, A.; Valacca, C.; Rios, E.; Populo, H.; Soares, P.; Sobrinho-Simoes, M.; Scorrano, L.; Máximo, V.; Campello, S. Mitochondrial dynamics protein Drp1 is overexpressed in oncocytic thyroid tumors and regulates cancer cell migration. PLoS ONE 2015, 10, e0122308. [Google Scholar] [CrossRef]
- Nagdas, S.; Kashatus, J.A.; Nascimento, A.; Hussain, S.S.; Trainor, R.E.; Pollock, S.R.; Adair, S.J.; Michaels, A.D.; Sesaki, H.; Stelow, E.B.; et al. Drp1 Promotes KRas-Driven Metabolic Changes to Drive Pancreatic Tumor Growth. Cell Rep. 2019, 28, 1845–1859.e5. [Google Scholar] [CrossRef]
- Tsuyoshi, H.; Orisaka, M.; Fujita, Y.; Asare-Werehene, M.; Tsang, B.K.; Yoshida, Y. Prognostic impact of Dynamin related protein 1 (Drp1) in epithelial ovarian cancer. BMC Cancer 2020, 20, 467. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Chen, E.; Pan, Z. The role of mitochondrial dysfunction in mesenchymal stem cell senescence. Cell Tissue Res. 2020, 382, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Bu, W.; Lv, G.; Xu, L.; Hou, D.; Wang, J.; Liu, X.; Yang, T.; Zhang, X.; Liu, Q.; et al. Disrupted mitochondrial homeostasis coupled with mitotic arrest generates antineoplastic oxidative stress. Oncogene 2022, 41, 427–443. [Google Scholar] [CrossRef]
- Zhu, M.; Min, S.; Mao, X.; Zhou, Y.; Zhang, Y.; Li, W.; Li, L.; Wu, L.; Cong, X.; Yu, G. Interleukin-13 promotes cellular senescence through inducing mitochondrial dysfunction in IgG4-related sialadenitis. Int. J. Oral. Sci. 2022, 14, 29. [Google Scholar] [CrossRef] [PubMed]
- Kamisawa, T.; Zen, Y.; Pillai, S.; Stone, J.H. IgG4-related disease. Lancet 2015, 385, 1460–1471. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.H.; Zen, Y.; Deshpande, V. IgG4-related disease. N. Engl. J. Med. 2012, 366, 539–551. [Google Scholar] [CrossRef]
- Floreani, A.; Okazaki, K.; Uchida, K.; Gershwin, M.E. IgG4-related disease: Changing epidemiology and new thoughts on a multisystem disease. J. Transl. Autoimmun. 2021, 4, 100074. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C.; Schuurman, J. IgG4 breaking the rules. Immunology 2002, 105, 9–19. [Google Scholar] [CrossRef]
- Nirula, A.; Glaser, S.M.; Kalled, S.L.; Taylor, F.R. What is IgG4? A review of the biology of a unique immunoglobulin subtype. Curr. Opin. Rheumatol. 2011, 23, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C.; Stapel, S.O.; Schuurman, J.; Rispens, T. Immunoglobulin G4: An odd antibody. Clin. Exp. Allergy 2009, 39, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Koike, T. IgG4-related disease: Why high IgG4 and fibrosis? Arthritis Res. Ther. 2013, 15, 103. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Aga, M.; Kondo, S.; Yamada, K.; Wakisaka, N.; Yagi-Nakanishi, S.; Tsuji, A.; Endo, K.; Murono, S.; Ito, M.; Muramatsu, M.; et al. Immunoglobulin class switching to IgG4 in Warthin tumor and analysis of serum IgG4 levels and IgG4-positive plasma cells in the tumor. Hum. Pathol. 2014, 45, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Shimo, T.; Yao, M.; Takebe, Y.; Ono, Y.; Obata, K.; Kurio, N.; Ibaragi, S.; Yoshioka, N.; Kishimoto, K.; Yanagi, Y.; et al. A case of adenoid cystic carcinoma associated with IgG4-related disease. Int. J. Surg. Case Rep. 2015, 10, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Tristan, C.; Shahani, N.; Sedlak, T.W.; Sawa, A. The diverse functions of GAPDH: Views from different subcellular compartments. Cell Signal. 2011, 23, 317–323. [Google Scholar] [CrossRef]
- Sirover, M.A. New insights into an old protein: The functional diversity of mammalian glyceraldehyde-3-phosphate dehydrogenase. Biochim. Biophys. Acta 1999, 1432, 159–184. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, R.; Sunaga, K.; Hirano, A.; Saunders, P.; Katsube, N.; Chuang, D.M. Evidence that glyceraldehyde-3-phosphate dehydrogenase is involved in age-induced apoptosis in mature cerebellar neurons in culture. J. Neurochem. 1996, 66, 928–935. [Google Scholar] [CrossRef]
- Dastoor, Z.; Dreyer, J.L. Potential role of nuclear translocation of glyceraldehyde-3-phosphate dehydrogenase in apoptosis and oxidative stress. J. Cell Sci. 2001, 114 Pt 9, 1643–1653. [Google Scholar] [CrossRef]
- Phadke, M.; Krynetskaia, N.; Mishra, A.; Krynetskiy, E. Accelerated cellular senescence phenotype of GAPDH-depleted human lung carcinoma cells. Biochem. Biophys. Res. Commun. 2011, 411, 409–415. [Google Scholar] [CrossRef]
- Phadke, M.S.; Krynetskaia, N.F.; Mishra, A.K.; Krynetskiy, E. Glyceraldehyde 3-phosphate dehydrogenase depletion induces cell cycle arrest and resistance to antimetabolites in human carcinoma cell lines. J. Pharmacol. Exp. Ther. 2009, 331, 77–86. [Google Scholar] [CrossRef]
- Colell, A.; Green, D.R.; Ricci, J.E. Novel roles for GAPDH in cell death and carcinogenesis. Cell Death Differ. 2009, 16, 1573–1581. [Google Scholar] [CrossRef]
- Hwang, S.; Disatnik, M.H.; Mochly-Rosen, D. Impaired GAPDH-induced mitophagy contributes to the pathology of Huntington’s disease. EMBO Mol. Med. 2015, 7, 1307–1326. [Google Scholar] [CrossRef]
- Teymoortash, A.; Bohne, F.; Jonsdottir, T.; Hoch, S.; Eivazi, B.; Roessler, M.; Werner, J.A.; Mandic, R. Human papilloma virus (HPV) is not implicated in the etiology of Warthin’s tumor of the parotid gland. Acta Otolaryngol. 2013, 133, 972–976. [Google Scholar] [CrossRef]
- Wiest, T.; Schwarz, E.; Enders, C.; Flechtenmacher, C.; Bosch, F.X. Involvement of intact HPV16 E6/E7 gene expression in head and neck cancers with unaltered p53 status and perturbed pRb cell cycle control. Oncogene 2002, 21, 1510–1517. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elhamid, E.S.; Elshafei, M.M. Immunohistochemical localization of mdm-2, p27Kip1 and bcl-2 in Warthin’s tumor of the parotid gland. Diagn. Pathol. 2009, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Rivlin, N.; Brosh, R.; Oren, M.; Rotter, V. Mutations in the p53 Tumor Suppressor Gene: Important Milestones at the Various Steps of Tumorigenesis. Genes Cancer 2011, 2, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Fonseca, I.; Roque, L.; Soares, J. Cytogenetic characterisation of Warthin’s tumour. Oral Oncol. 1997, 33, 344–347. [Google Scholar] [CrossRef]
- Bullerdiek, J.; Haubrich, J.; Meyer, K.; Bartnitzke, S. Translocation t(11;19)(q21;p13.1) as the sole chromosome abnormality in a cystadenolymphoma (Warthin’s tumor) of the parotid gland. Cancer Genet. Cytogenet. 1988, 35, 129–132. [Google Scholar] [CrossRef]
- Arida, M.; Barnes, E.L.; Hunt, J.L. Molecular assessment of allelic loss in Warthin tumors. Mod. Pathol. 2005, 18, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Kashima, K.; Daa, T.; Yokoyama, S.; Nakayama, I. Clonal analysis of the epithelial component of Warthin’s tumor. Hum. Pathol. 2000, 31, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Skalova, A.; Kaspirkova, J.; Andrle, P.; Hosticka, L.; Vanecek, T. Human papillomaviruses are not involved in the etiopathogenesis of salivary gland tumors. Cesk Patol. 2013, 49, 72–75. [Google Scholar]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoki, R.; Tanaka, T. Pathogenesis of Warthin’s Tumor: Neoplastic or Non-Neoplastic? Cancers 2024, 16, 912. https://doi.org/10.3390/cancers16050912
Aoki R, Tanaka T. Pathogenesis of Warthin’s Tumor: Neoplastic or Non-Neoplastic? Cancers. 2024; 16(5):912. https://doi.org/10.3390/cancers16050912
Chicago/Turabian StyleAoki, Ryogo, and Takuji Tanaka. 2024. "Pathogenesis of Warthin’s Tumor: Neoplastic or Non-Neoplastic?" Cancers 16, no. 5: 912. https://doi.org/10.3390/cancers16050912
APA StyleAoki, R., & Tanaka, T. (2024). Pathogenesis of Warthin’s Tumor: Neoplastic or Non-Neoplastic? Cancers, 16(5), 912. https://doi.org/10.3390/cancers16050912