Genotoxic and Anti-Migratory Effects of Camptothecin Combined with Celastrol or Resveratrol in Metastatic and Stem-like Cells of Colon Cancer

 , ,
, ,  ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Drug Solutions
2.3. Cell Line and Culture Conditions
2.4. Apoptosis and Necrosis Detection
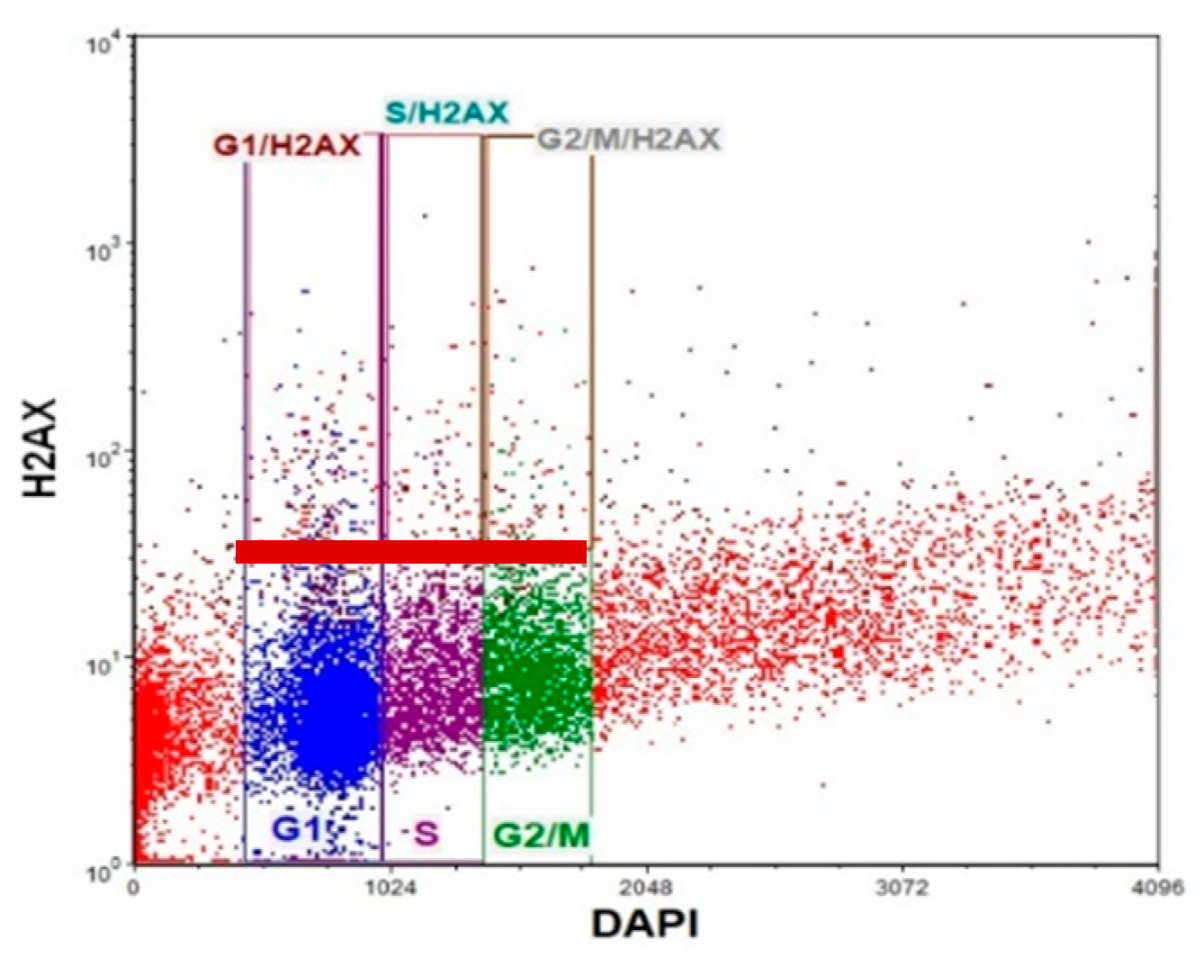
2.5. γH2AX and γH2AX Cell Cycle Detection
2.6. Fast Halo Assay (FHA)
2.7. Scratch Assay
2.8. Statistical Analysis
3. Results
3.1. Evaluation of Genotoxic Effects
3.1.1. Induction of Double-Stranded DNA Breaks (DSBs)
Camptothecin-Induced γH2AX Formation
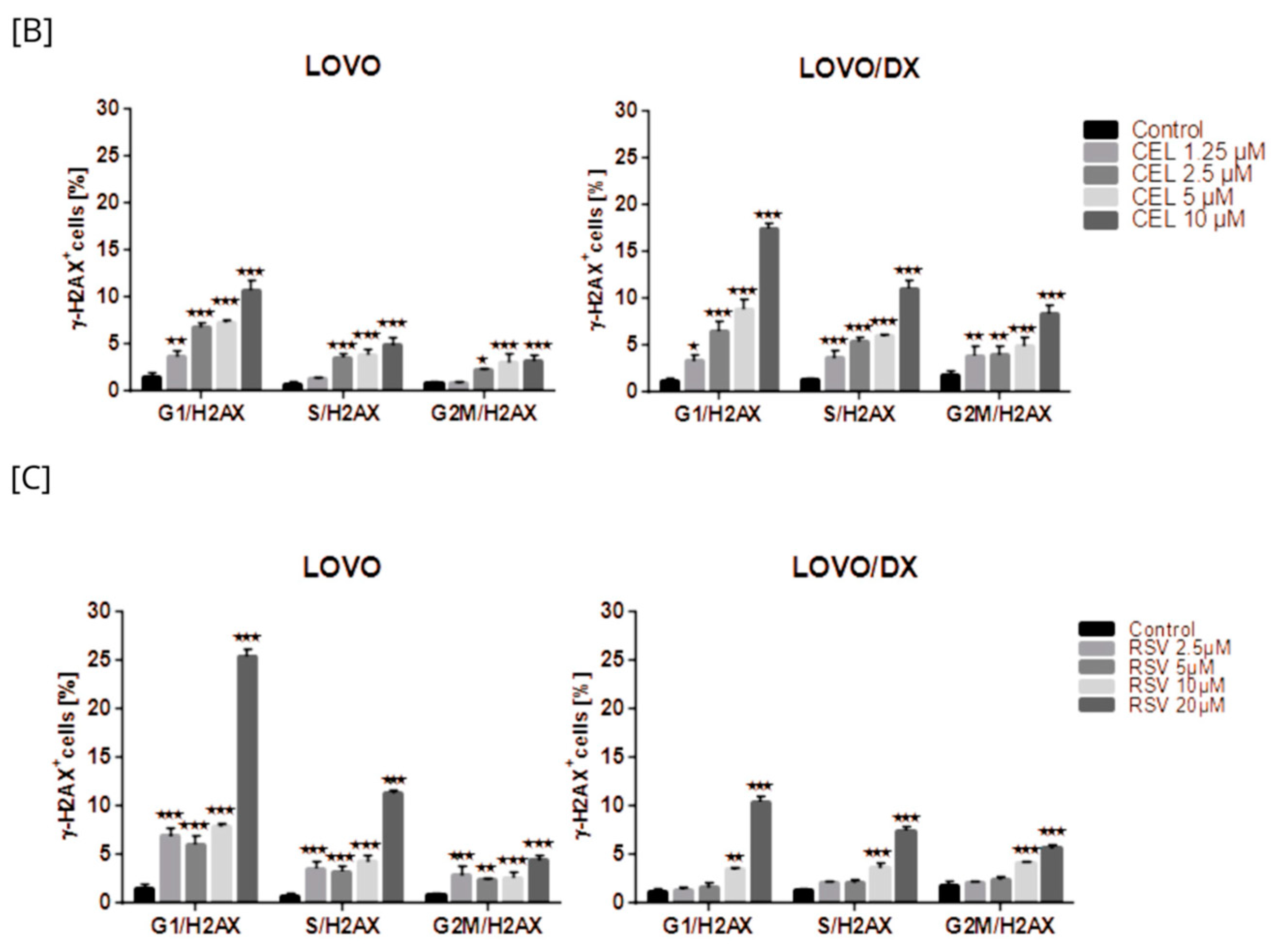
Induction of γH2AX by the Combination of Camptothecin with Celastrol
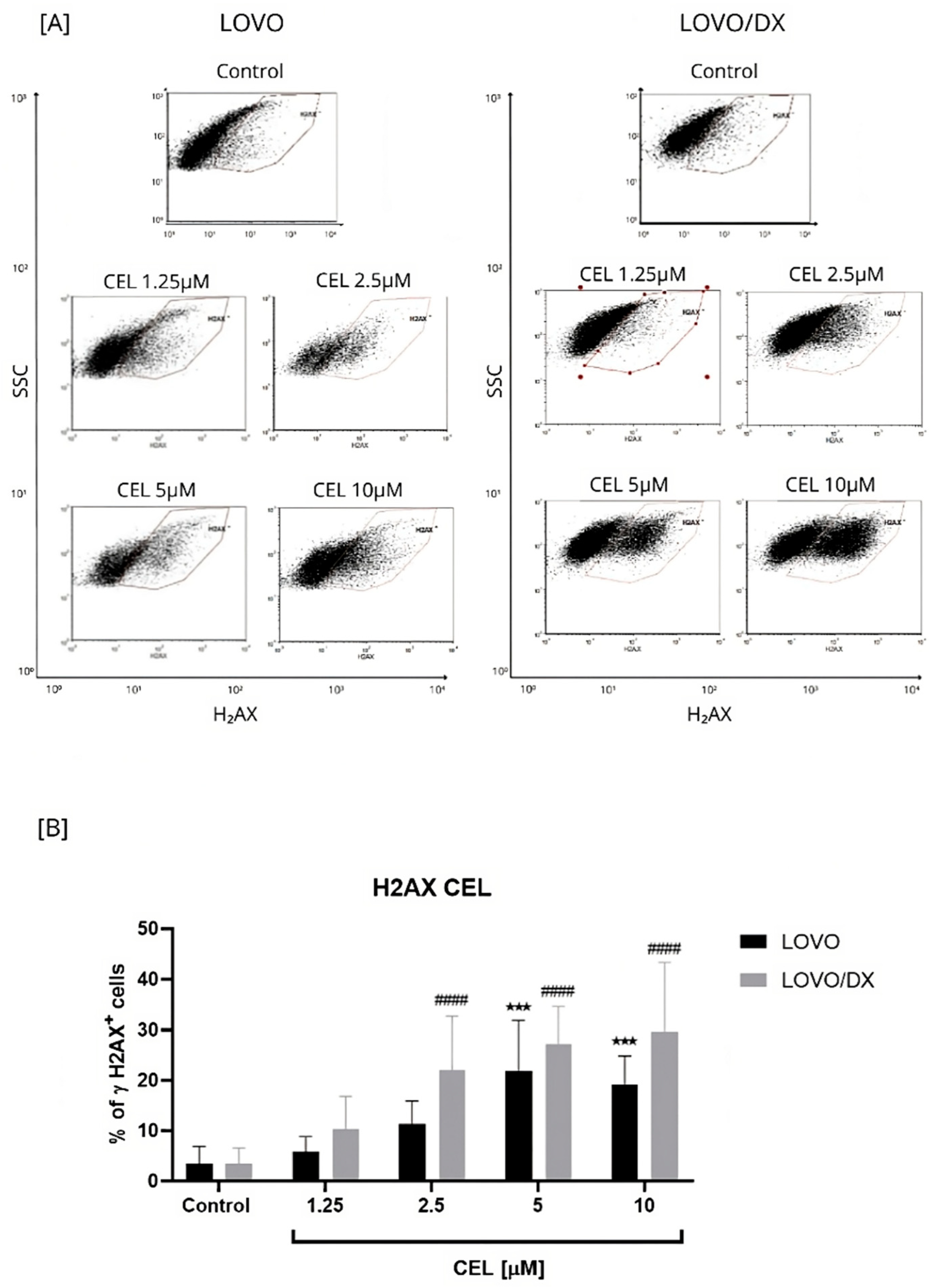

Induction of γH2AX by the Combination of Camptothecin with Resveratrol

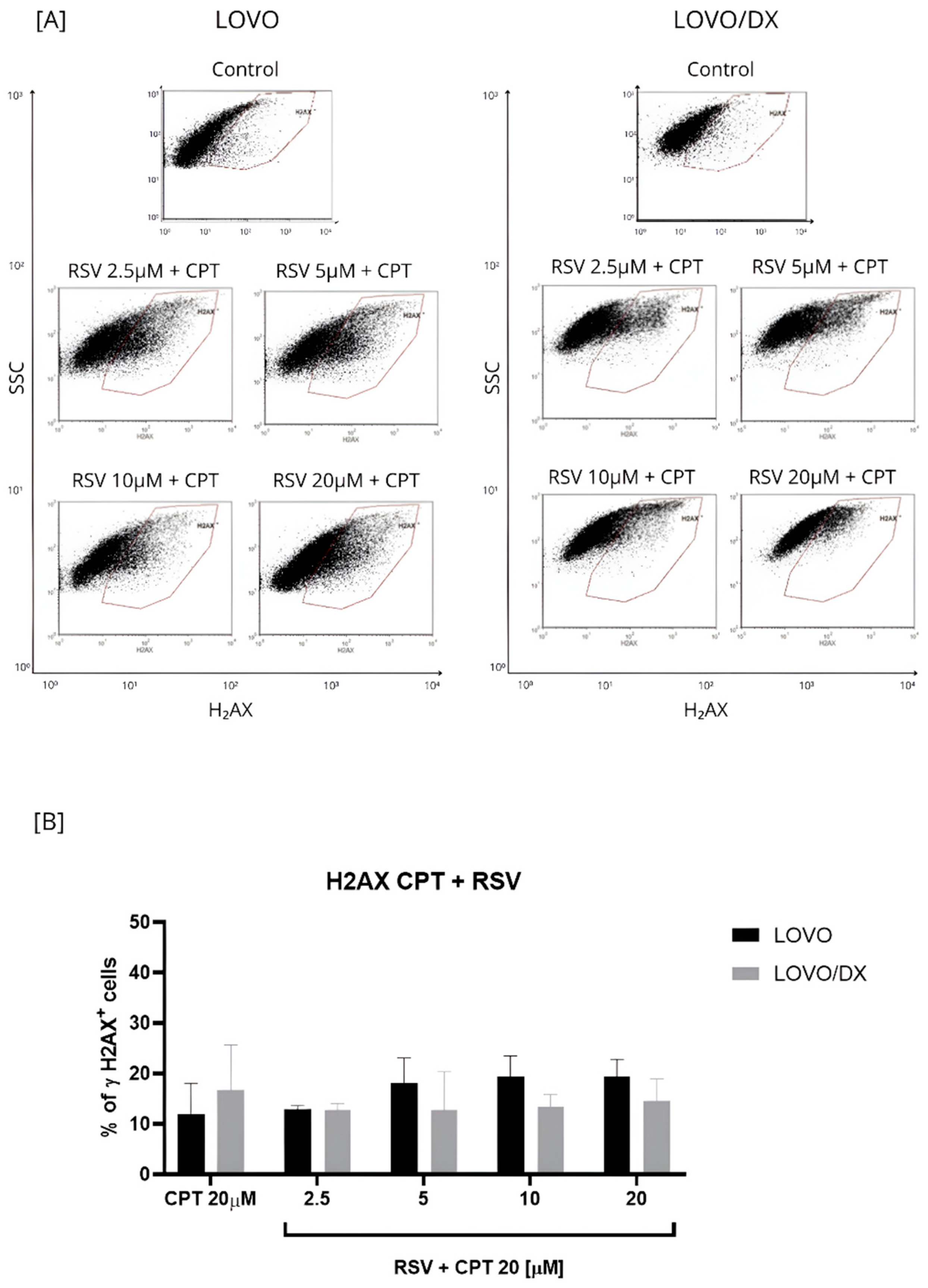
3.1.2. Induction of DNA Fragmentation
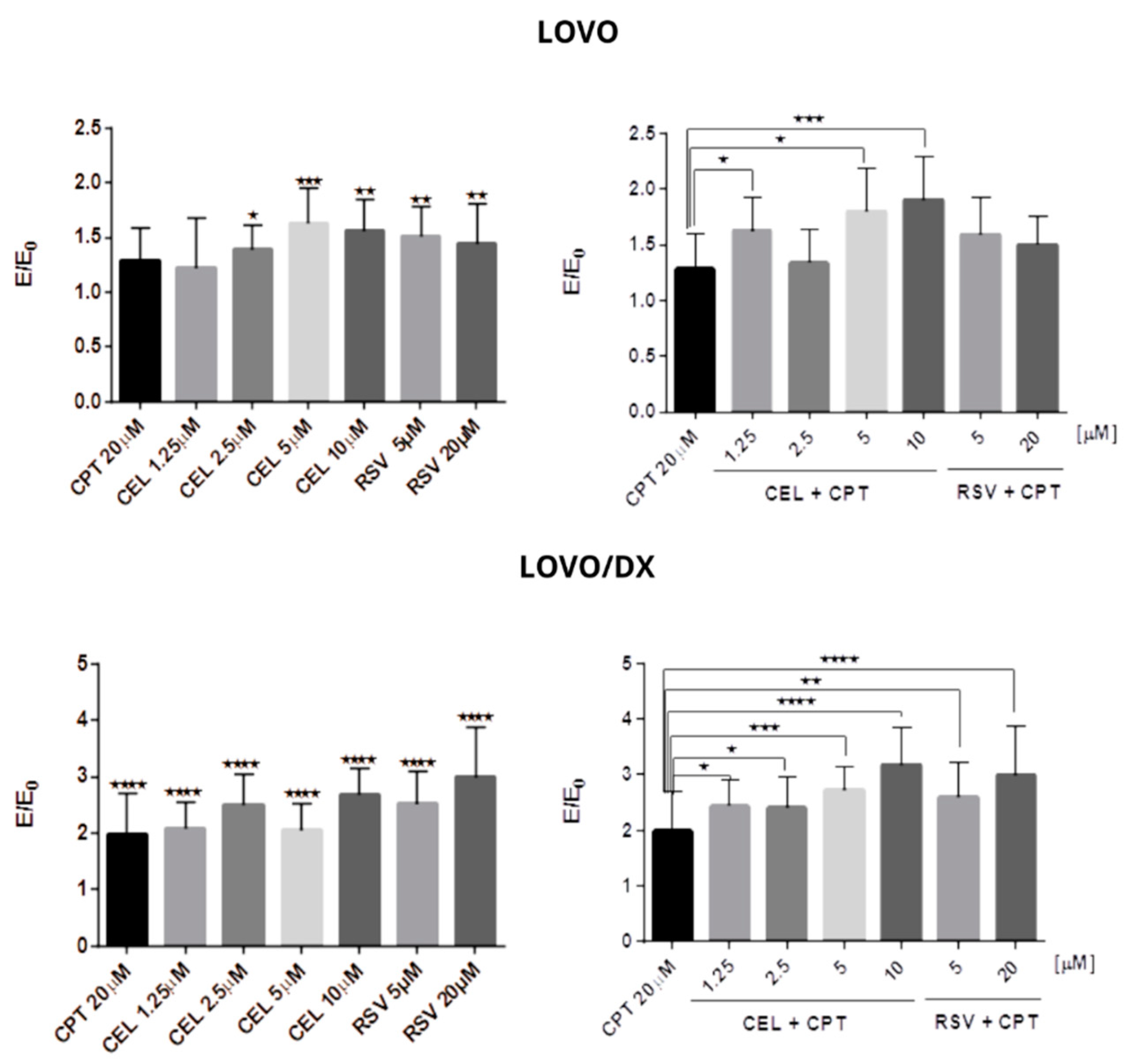
3.1.3. Calculation of the Combination Index (CI)
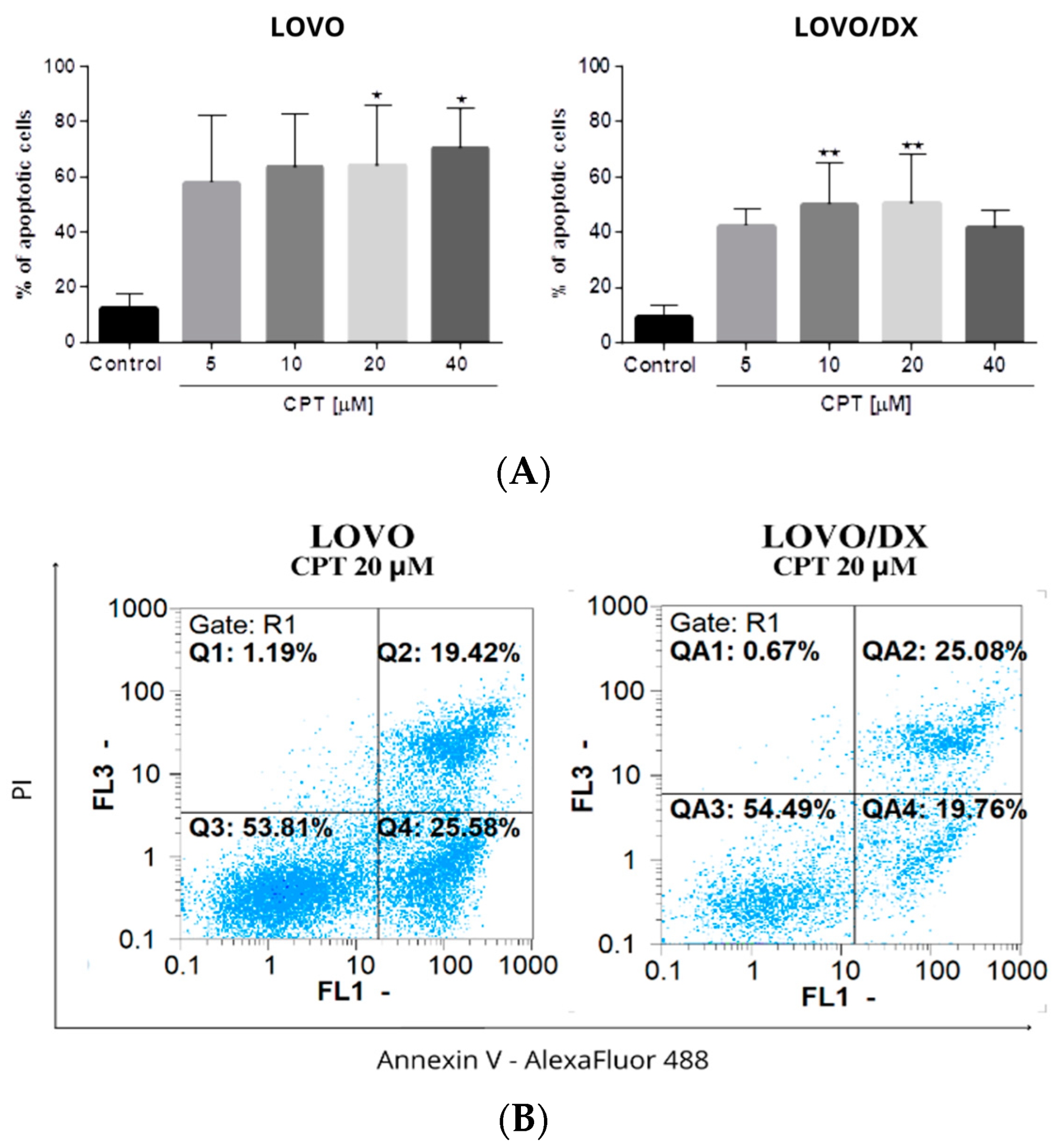
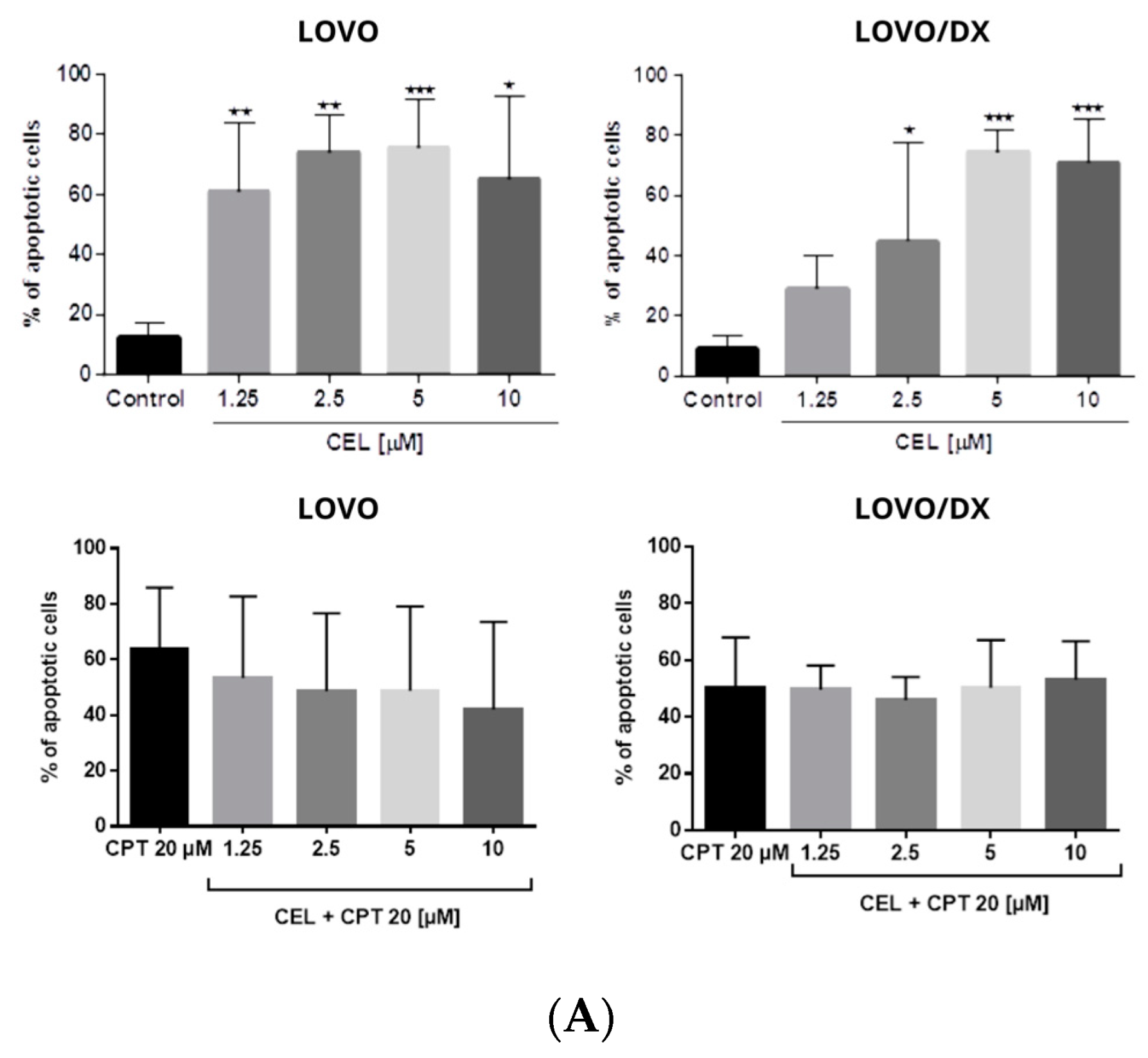
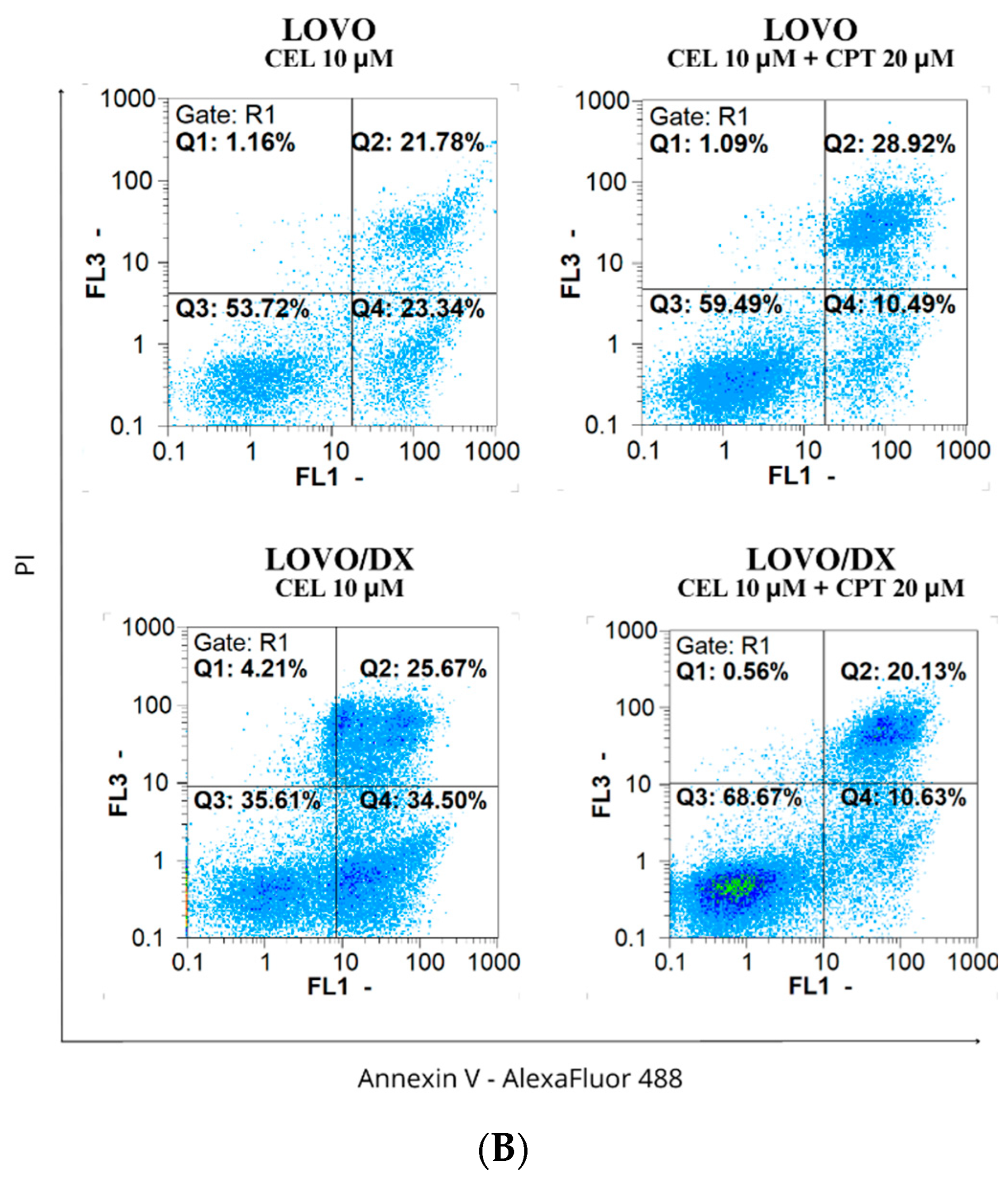
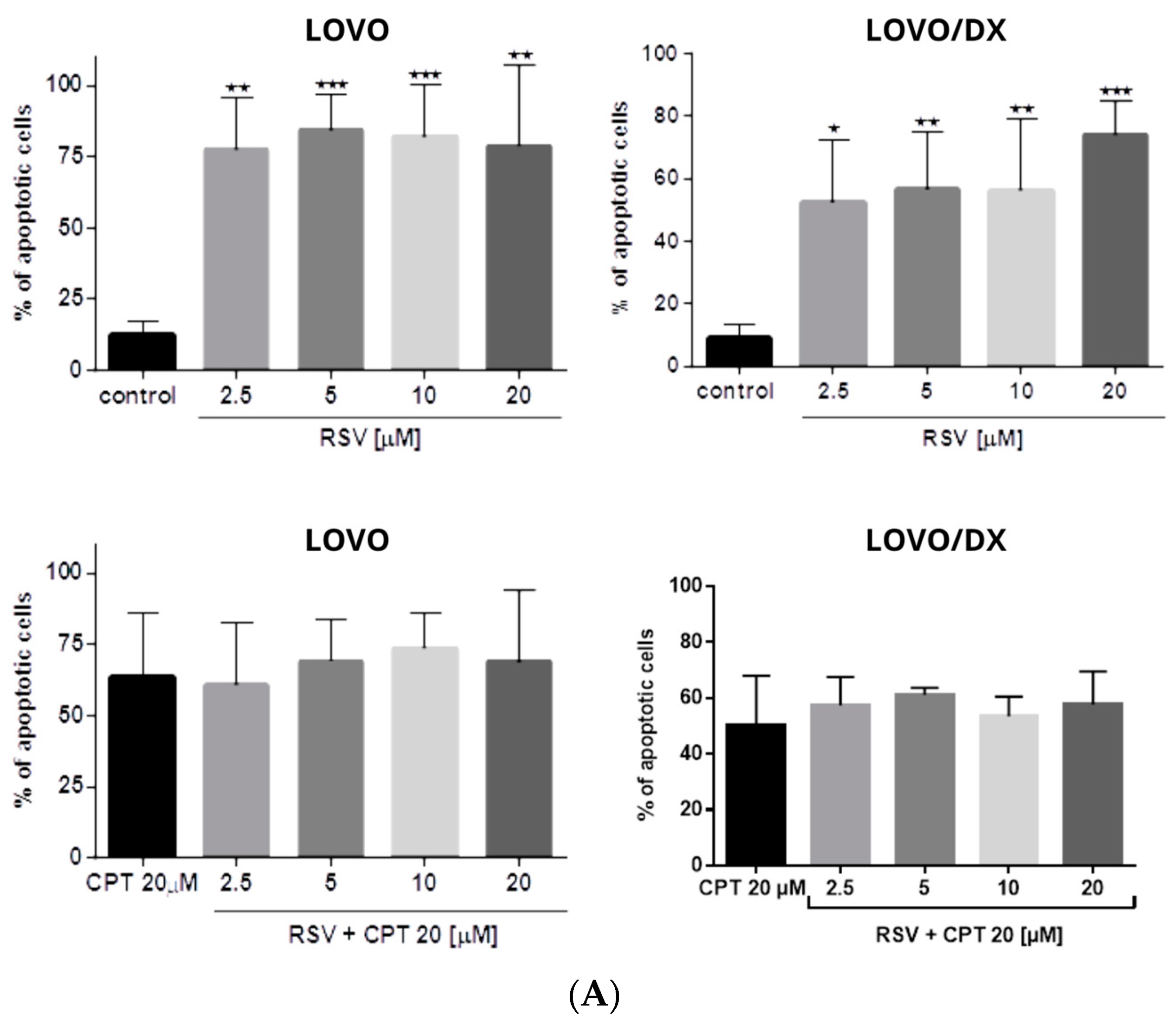
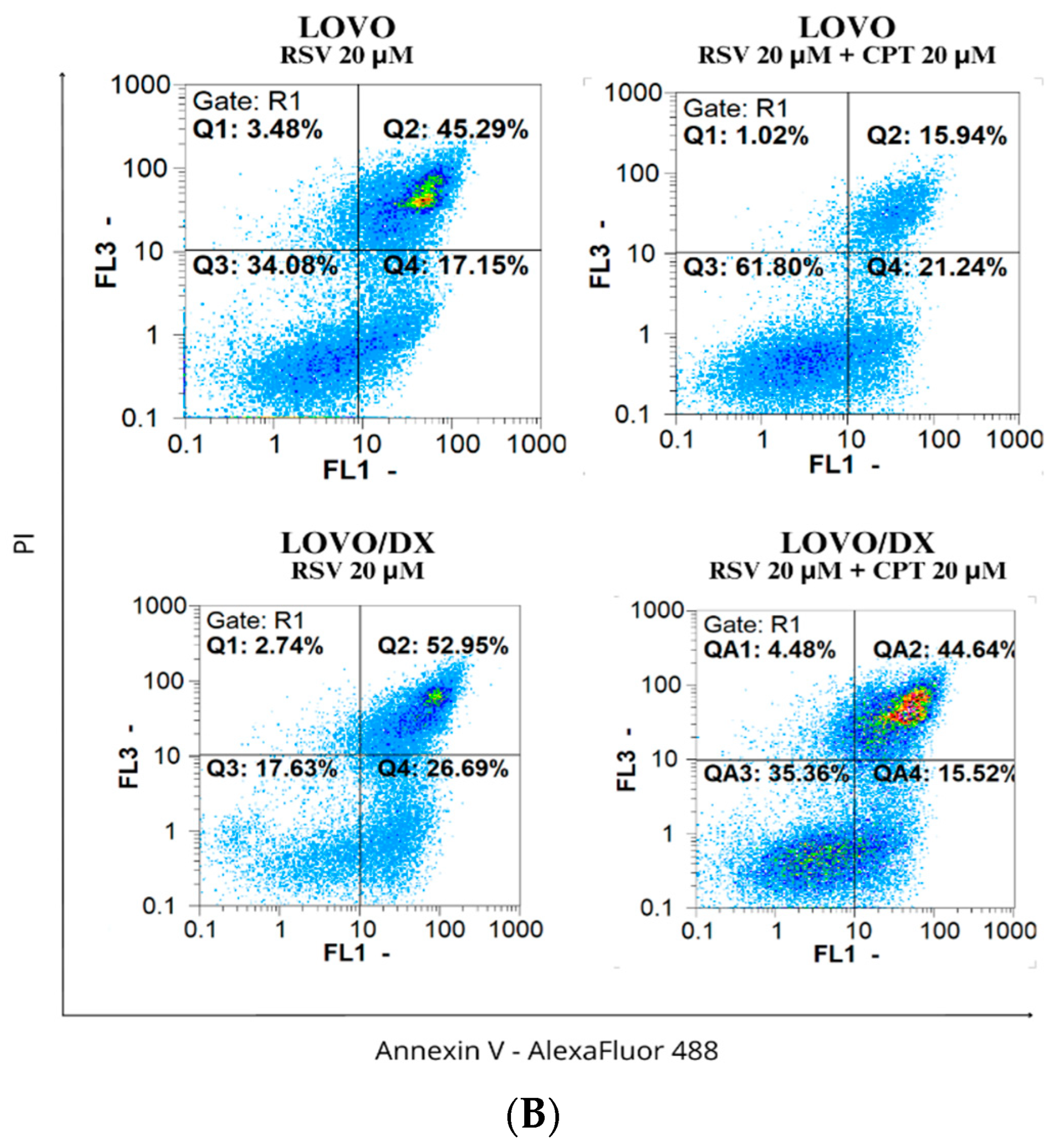
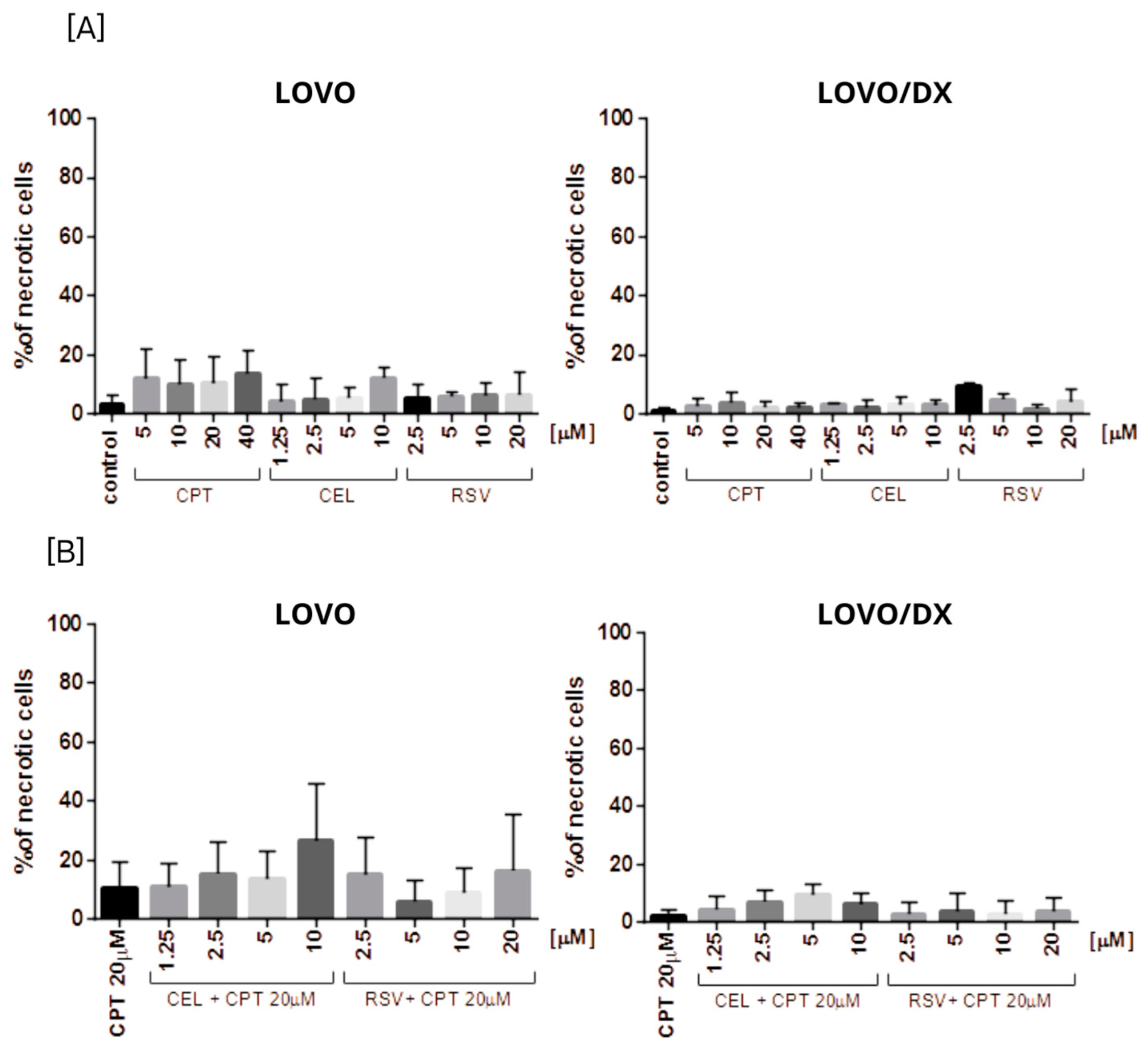
3.2. Effects of Camptothecin, Celastrol, and Resveratrol and Their Combination on Apoptotic and Necrotic Cell Death
3.3. Frequency of γ-H2AX in Different Phases of the Cell Cycle after Cell Exposure to Tested Compounds and Their Combination
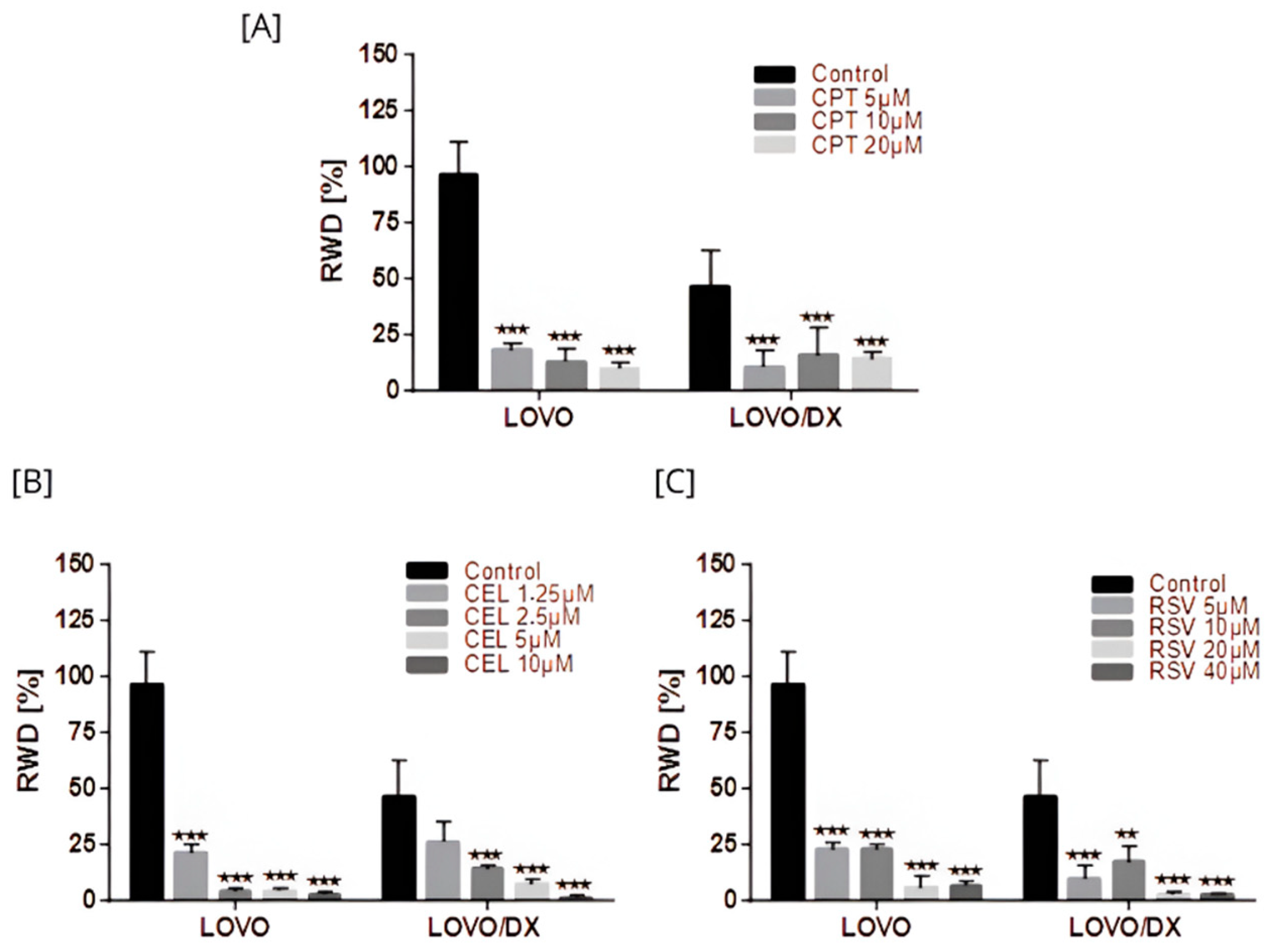
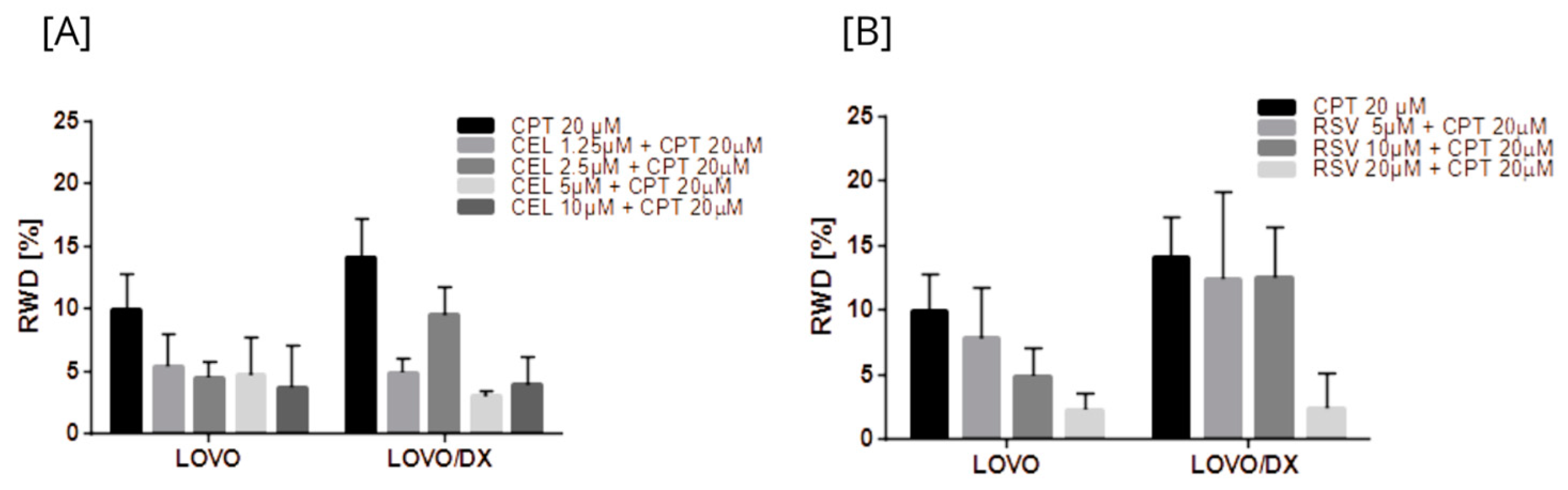
3.4. Effects of Camptothecin, Celastrol, Resveratrol, and Their Combination on Cancer Cells Migration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baidoun, F.; Elshiwy, K.; Elkeraie, Y.; Merjaneh, Z.; Khoudari, G.; Sarmini, M.T.; Gad, M.; Al-Husseini, M.; Saad, A. Colorectal Cancer Epidemiology: Recent Trends and Impact on Outcomes. Curr. Drug Targets 2021, 22, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Xu, P. Global colorectal cancer burden in 2020 and projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Vogel, J.D.; Felder, S.I.; Bhama, A.R.; Hawkins, A.T.; Langenfeld, S.J.; Shaffer, V.O.; Thorsen, A.J.; Weiser, M.R.; Chang, G.J.; Lightner, A.L.; et al. The American Society of Colon and Rectal Surgeons Clinical Practice Guidelines for the Management of Colon Cancer. Dis. Colon Rectum 2021, 65, 148–177. [Google Scholar] [CrossRef]
- Morris, V.K.; Kennedy, E.B.; Baxter, N.N.; Benson, A.B.; Cercek, A.; Cho, M.; Ciombor, K.K.; Cremolini, C.; Davis, A.; Deming, D.A.; et al. Treatment of Metastatic Colorectal Cancer: ASCO Guideline. J. Clin. Oncol. 2023, 41, 678–700. [Google Scholar] [CrossRef]
- Chakedis, J.; Schmidt, C.R. Surgical Treatment of Metastatic Colorectal Cancer. Surg. Oncol. Clin. N. Am. 2018, 27, 377–399. [Google Scholar] [CrossRef]
- Hervieu, C.; Christou, N.; Battu, S.; Mathonnet, M. The Role of Cancer Stem Cells in Colorectal Cancer: From the Basics to Novel Clinical Trials. Cancers 2021, 13, 1092. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Garcia, E.; Grasselli, J.; Argiles, G.; Elez, M.E.; Tabernero, J. Current and advancing treatments for metastatic colorectal cancer. Expert Opin. Biol. Ther. 2015, 16, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.; Li, S.; Zhu, Z.; Liu, Q.; Zhang, R.; Yang, Y.; Li, X. Exploring immunotherapy in colorectal cancer. J. Hematol. Oncol. 2022, 15, 1–28. [Google Scholar] [CrossRef]
- Lin, S.R.; Chang, C.H.; Hsu, C.F.; Tsai, M.J.; Cheng, H.; Leong, M.K.; Sung, P.J.; Chen, J.C.; Weng, C.F. Natural compounds as potential adjuvants to cancer therapy: Preclinical evidence. Br. J. Pharmacol. 2020, 177, 1409–1423. [Google Scholar] [CrossRef]
- Kubczak, M.; Szustka, A.; Rogalińska, M. Molecular Targets of Natural Compounds with Anti-Cancer Properties. Int. J. Mol. Sci. 2021, 22, 13659. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.V.; Duarte, M.; Silva, P.; da Silva, A.B.; Duarte, C.M.M.; Cifuentes, A.; García-Cañas, V.; Bronze, M.R.; Albuquerque, C.; Serra, A.T. Polymethoxylated Flavones Target Cancer Stemness and Improve the Antiproliferative Effect of 5-Fluorouracil in a 3D Cell Model of Colorectal Cancer. Nutrients 2019, 11, 326. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, Y.; Luo, W.; Chen, S.; Lin, F.; Zhang, X.; Fan, S.; Shen, X.; Wang, Y.; Liang, G. Celastrol induces ROS-mediated apoptosis via directly targeting peroxiredoxin-2 in gastric cancer cells. Theranostics 2020, 10, 10290–10308. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an anti-cancer agent: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1428–1447. [Google Scholar] [CrossRef]
- Shi, J.; Li, J.; Xu, Z.; Chen, L.; Luo, R.; Zhang, C.; Gao, F.; Zhang, J.; Fu, C. Celastrol: A Review of Useful Strategies Overcoming its Limitation in Anticancer Application. Front. Pharmacol. 2020, 11, 558741. [Google Scholar] [CrossRef]
- Amin, S.A.; Adhikari, N.; Jha, T.; Gayen, S. A Review on Camptothecin Analogs with Promising Cytotoxic Profile. Anti-Cancer Agents Med. Chem. 2018, 18, 1796–1814. [Google Scholar] [CrossRef]
- Kümler, I.; Brünner, N.; Stenvang, J.; Balslev, E.; Nielsen, D.L. A systematic review on topoisomerase 1 inhibition in the treatment of metastatic breast cancer. Breast Cancer Res. Treat. 2013, 138, 347–358. [Google Scholar] [CrossRef]
- Khaiwa, N.; Maarouf, N.R.; Darwish, M.H.; Alhamad, D.W.; Sebastian, A.; Hamad, M.; Omar, H.A.; Orive, G.; Al-Tel, T.H. Camptothecin’s journey from discovery to WHO Essential Medicine: Fifty years of promise. Eur. J. Med. Chem. 2021, 223, 113639. [Google Scholar] [CrossRef]
- Bailly, C. Irinotecan: 25 years of cancer treatment. Pharmacol. Res. 2019, 148, 104398. [Google Scholar] [CrossRef]
- Xu, Y.; Villalona-Calero, M.A. Irinotecan: Mechanisms of tumor resistance and novel strategies for modulating its activity. Ann. Oncol. 2002, 13, 1841–1851. [Google Scholar] [CrossRef]
- Moreira, H.; Szyjka, A.; Grzesik, J.; Pelc, K.; Żuk, M.; Kulma, A.; Emhemmed, F.; Muller, C.D.; Gąsiorowski, K.; Barg, E. Celastrol and Resveratrol Modulate SIRT Genes Expression and Exert Anticancer Activity in Colon Cancer Cells and Cancer Stem-like Cells. Cancers 2022, 14, 1372. [Google Scholar] [CrossRef]
- Moreira, H.; Szyjka, A.; Gąsiorowski, K. Chemopreventive activity of celastrol in drug-resistant human colon carcinoma cell cultures. Oncotarget 2018, 9, 21211–21223. [Google Scholar] [CrossRef] [PubMed]
- Moreira, H.; Szyjka, A.; Paliszkiewicz, K.; Barg, E. Prooxidative Activity of Celastrol Induces Apoptosis, DNA Damage, and Cell Cycle Arrest in Drug-Resistant Human Colon Cancer Cells. Oxidative Med. Cell. Longev. 2019, 2019, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, Y.; Bindokas, V.P.; Duggan, R.C.; Murley, J.S.; Grdina, D.J. Flow Cytometric Analysis of Phosphorylated Histone H2AX Following Exposure to Ionizing Radiation in Human Microvascular Endothelial Cells. J. Radiat. Res. 2006, 47, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Radajewska, A.; Moreira, H.; Bęben, D.; Siwiela, O.; Szyjka, A.; Gębczak, K.; Nowak, P.; Frąszczak, J.; Emhemmed, F.; Muller, C.D.; et al. Combination of Irinotecan and Melatonin with the Natural Compounds Wogonin and Celastrol for Colon Cancer Treatment. Int. J. Mol. Sci. 2023, 24, 9544. [Google Scholar] [CrossRef]
- Bonner, W.M.; Redon, C.E.; Dickey, J.S.; Nakamura, A.J.; Sedelnikova, O.A.; Solier, S.; Pommier, Y. γH2AX and cancer. Nat. Rev. Cancer 2008, 8, 957–967. [Google Scholar] [CrossRef]
- Podhorecka, M. Gamma H2AX in the recognition of DNA double-strand breaks. Postep. Hig. Med. Dosw. 2009, 63, 92–98. [Google Scholar]
- Ivashkevich, A.; Redon, C.E.; Nakamura, A.J.; Martin, R.F.; Martin, O.A. Use of the γ-H2AX assay to monitor DNA damage and repair in translational cancer research. Cancer Lett. 2011, 327, 123–133. [Google Scholar] [CrossRef]
- Johansson, P.; Fasth, A.; Ek, T.; Hammarsten, O. Validation of a flow cytometry-based detection of γ-H2AX, to measure DNA damage for clinical applications. Cytom. Part B (Clin. Cytom.) 2017, 92B, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Wiatrak, B.; Krzyżak, E.; Szczęśniak-Sięga, B.; Szandruk-Bender, M.; Szeląg, A.; Nowak, B. Effect of tricyclic 1,2-thiazine derivatives in neuroinflammation induced by preincubation with lipopolysaccharide or coculturing with microglia-like cells. Pharmacol. Rep. 2022, 74, 890–908. [Google Scholar] [CrossRef]
- Matthews, H.; Deakin, J.; Rajab, M.; Idris-Usman, M.; Nirmalan, N.J. Investigating antimalarial drug interactions of emetine dihydrochloride hydrate using CalcuSyn-based interactivity calculations. PLoS ONE 2017, 12, e0173303. [Google Scholar] [CrossRef] [PubMed]
- Vinogradov, S.; Wei, X. Cancer stem cells and drug resistance: The potential of nanomedicine. Nanomedicine 2012, 7, 597–615. [Google Scholar] [CrossRef] [PubMed]
- Vladu, A.F.; Ficai, D.; Ene, A.G.; Ficai, A. Combination Therapy Using Polyphenols: An Efficient Way to Improve Antitumoral Activity and Reduce Resistance. Int. J. Mol. Sci. 2022, 23, 10244. [Google Scholar] [CrossRef] [PubMed]
- Stearns, M.E.; Amatangelo, M.D.; Varma, D.; Sell, C.; Goodyear, S.M. Combination therapy with epigallocatechin-3-gallate and doxorubicin in human prostate tumor modeling studies. Am. J. Pathol. 2010, 177, 3169–3179. [Google Scholar] [CrossRef]
- Moracci, L.; Sensi, F.; Biccari, A.; Crotti, S.; Gaio, E.; Benetti, F.; Traldi, P.; Pucciarelli, S.; Agostini, M. An investigation on [5 fluorouracil and epigallocatechin-3-gallate] complex activity on HT-29 cell death and its stability in gastrointestinal fluid. Oncotarget 2022, 13, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, R.; Zhang, X.; Zhang, B.; Yao, Q. Curcumin may reverse 5-fluorouracil resistance on colonic cancer cells by regulating TET1-NKD-Wnt signal pathway to inhibit the EMT progress. Biomed. Pharmacother. 2020, 129, 110381. [Google Scholar] [CrossRef]
- Chung, S.S.; Dutta, P.; Austin, D.; Wang, P.; Awad, A.; Vadgama, J.V. Combination of resveratrol and 5-flurouracil enhanced anti-telomerase activity and apoptosis by inhibiting STAT3 and Akt signaling pathways in human colorectal cancer cells. Oncotarget 2018, 9, 32943–32957. [Google Scholar] [CrossRef]
- Cao, P.; Xia, Y.; He, W.; Zhang, T.; Hong, L.; Zheng, P.; Shen, X.; Liang, G.; Cui, R.; Zou, P. Enhancement of oxaliplatin-induced colon cancer cell apoptosis by alantolactone, a natural product inducer of ROS. Int. J. Biol. Sci. 2019, 15, 1676–1684. [Google Scholar] [CrossRef]
- Wu, W.; Dong, J.; Gou, H.; Geng, R.; Yang, X.; Chen, D.; Xiang, B.; Zhang, Z.; Ren, S.; Chen, L.; et al. EGCG synergizes the therapeutic effect of irinotecan through enhanced DNA damage in human colorectal cancer cells. J. Cell. Mol. Med. 2021, 25, 7913–7921. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, K.; Yan, C.; Yin, Y.; He, S.; Qiu, L.; Li, G. Natural Polyphenols for Treatment of Colorectal Cancer. Molecules 2022, 27, 8810. [Google Scholar] [CrossRef]
- Honari, M.; Shafabakhsh, R.; Reiter, R.J.; Mirzaei, H.; Asemi, Z. Resveratrol is a promising agent for colorectal cancer prevention and treatment: Focus on molecular mechanisms. Cancer Cell Int. 2019, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y. Topoisomerase I inhibitors: Camptothecins and beyond. Nat. Rev. Cancer 2006, 6, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xu, C.; Gao, X.; Yao, Q. Platinum-based drugs for cancer therapy and anti-tumor strategies. Theranostics 2022, 12, 2115–2132. [Google Scholar] [CrossRef] [PubMed]
- Bartek, J. DNA damage response, genetic instability and cancer: From mechanistic insights to personalized treatment. Mol. Oncol. 2011, 5, 303–307. [Google Scholar] [CrossRef]
- Sestili, P.; Calcabrini, C.; Diaz, A.R.; Fimognari, C.; Stocchi, V. The fast-halo assay for the detection of DNA damage. Methods Mol. Biol. 2017, 1644, 75–93. [Google Scholar] [CrossRef]
- Sestili, P. The fast-halo assay for the assessment of DNA damage at the single-cell level. Methods Mol. Biol. 2009, 521, 517–533. [Google Scholar] [CrossRef]
- Roy, A.; Tesauro, C.; Frøhlich, R.; Hede, M.S.; Nielsen, M.J.; Kjeldsen, E.; Bonven, B.; Stougaard, M.; Gromova, I.; Knudsen, B.R. Decreased camptothecin sensitivity of the stem-cell-like fraction of caco2 cells correlates with an altered phosphorylation pattern of topoisomerase I. PLoS ONE 2014, 9, e99628. [Google Scholar] [CrossRef]
- Tesauro, C.; Keller, J.G.; Gromova, I.; Gromov, P.; Frøhlich, R.; Erlandsen, J.U.; Andersen, A.H.; Stougaard, M.; Knudsen, B.R. Different Camptothecin Sensitivities in Subpopulations of Colon Cancer Cells Correlate with Expression of Different Phospho-Isoforms of Topoisomerase I with Different Activities. Cancers 2020, 12, 1240. [Google Scholar] [CrossRef]
- Lalloo, A.K.; Luo, F.R.; Guo, A.; Paranjpe, P.V.; Lee, S.-H.; Vyas, V.; Rubin, E.; Sinko, P.J. Membrane transport of camptothecin: Facilitation by human P-glycoprotein (ABCB1) and multidrug resistance protein 2 (ABCC2). BMC Med. 2004, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Demoulin, B.; Hermant, M.; Castrogiovanni, C.; Staudt, C.; Dumont, P. Resveratrol induces DNA damage in colon cancer cells by poisoning topoisomerase II and activates the ATM kinase to trigger p53-dependent apoptosis. Toxicol. Vitr. 2015, 29, 1156–1165. [Google Scholar] [CrossRef]
- Wang, J. DNA damage and apoptosis. Cell Death Differ. 2001, 8, 1047–1048. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Wei, W.; Dunaway, S.; Darnowski, J.W.; Calabresi, P.; Sedivy, J.; Hendrickson, E.A.; Balan, K.V.; Pantazis, P.; Wyche, J.H. Role of p21 in apoptosis and senescence of human colon cancer cells treated with camptothecin. J. Biol. Chem. 2002, 277, 17154–17160. [Google Scholar] [CrossRef] [PubMed]
- Liskova, V.; Kajsik, M.; Chovancova, B.; Roller, L.; Krizanova, O. Camptothecin, triptolide, and apoptosis inducer kit have differential effects on mitochondria in colorectal carcinoma cells. FEBS Open Bio 2022, 12, 913–924. [Google Scholar] [CrossRef]
- Park, K.; Woubit, A.S.; Fermin, C.D.; Reddy, G.; Habtemariam, T.; Chung, J.W.; Park, M.; Seol, D.-W.; Kim, M. Hypoxia inhibition of camptothecin-induced apoptosis by Bax loss. Biologia 2012, 67, 616–621. [Google Scholar] [CrossRef]
- Shen, S.; Shao, Y.; Li, C. Different types of cell death and their shift in shaping disease. Cell Death Discov. 2023, 9, 284. [Google Scholar] [CrossRef]
- Luo, P.; Liu, D.; Zhang, Q.; Yang, F.; Wong, Y.-K.; Xia, F.; Zhang, J.; Chen, J.; Tian, Y.; Yang, C.; et al. Celastrol induces ferroptosis in activated HSCs to ameliorate hepatic fibrosis via targeting peroxiredoxins and HO-1. Acta Pharm. Sin. B 2021, 12, 2300–2314. [Google Scholar] [CrossRef]
- Li, J.; Fan, Y.; Zhang, Y.; Liu, Y.; Yu, Y.; Ma, M. Resveratrol Induces Autophagy and Apoptosis in Non-Small-Cell Lung Cancer Cells by Activating the NGFR-AMPK-mTOR Pathway. Nutrients 2022, 14, 2413. [Google Scholar] [CrossRef]
- MacPhail, S.H.; Banáth, J.P.; Yu, Y.; Chu, E.; Olive, P.L. Cell Cycle-Dependent Expression of Phosphorylated Histone H2AX: Reduced Expression in Unirradiated but not X-Irradiated G1-Phase Cells. Radiat. Res. 2003, 159, 759–767. [Google Scholar] [CrossRef]
- Ree, A.H.; Stokke, T.; Bratland, A.; Patzke, S.; Nome, R.V.; Folkvord, S.; Meza-Zepeda, L.A.; Flatmark, K.; Fodstad, O.; Andersson, Y. DNA damage responses in cell cycle G2 phase and mitosis-tracking and targeting. Anticancer Res. 2006, 26, 1909–1916. [Google Scholar]
- Clay, D.E.; Fox, D.T. DNA Damage Responses during the Cell Cycle: Insights from Model Organisms and Beyond. Genes 2021, 12, 1882. [Google Scholar] [CrossRef]
- Yoo, J.-M.; Kim, Y.J.; Lee, S.-J.; Kim, S.H. Sequential administration of camptothecin sensitizes human colon cancer HCT116 cells to paclitaxel via p21Cip1/WAF1. Anim. Cells Syst. 2011, 15, 9–17. [Google Scholar] [CrossRef]
- Li, B.; Huang, J.; Guan, J.; Chen, L. Camptothecin inhibits the progression of NPC by regulating TGF-β-induced activation of the PI3K/AKT signaling pathway. Oncol. Lett. 2018, 16, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wu, S.; Li, X. Camptothecin inhibits migration, invasion and clonogenic property of liver cancer cells by modulating microrna expression. Acta Pol. Pharm. -Drug Res. 2020, 77, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Bufu, T.; Di, X.; Yilin, Z.; Gege, L.; Xi, C.; Ling, W. Celastrol inhibits colorectal cancer cell proliferation and migration through suppression of MMP3 and MMP7 by the PI3K/AKT signaling pathway. Anti-Cancer Drugs 2018, 29, 530–538. [Google Scholar] [CrossRef]
- Yuan, L.; Zhou, M.; Huang, D.; Wasan, H.S.; Zhang, K.; Sun, L.; Huang, H.; Ma, S.; Shen, M.; Ruan, S. Resveratrol inhibits the invasion and metastasis of colon cancer through reversal of epithelial mesenchymal transition via the AKT/GSK 3β/Snail signaling pathway. Mol. Med. Rep. 2019, 20, 2783–2795. [Google Scholar] [CrossRef]
- Piet, M.; Paduch, R. Ursolic and oleanolic acids in combination therapy inhibit migration of colon cancer cells through downregulation of the uPA/uPAR-dependent MMPs pathway. Chem.-Biol. Interact. 2022, 368, 110202. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CI (Combination Index) | ||
|---|---|---|
| LOVO | LOVO/DX | |
| CPT 20 µM + CEL 1.25 µM | 1.2 | 0.3 |
| CPT 20 µM + CEL 2.5 µM | 2.8 | 0.4 |
| CPT 20 µM + CEL 5 µM | 3.0 | 0.5 |
| CPT 20 µM + CEL 10 µM | >10 | 0.6 |
| CI (Combination Index) | ||
|---|---|---|
| LOVO | LOVO/DX | |
| CPT 20 µM + RSV 2.5 µM | 8.4 | 0.5 |
| CPT 20 µM + RSV 5 µM | >10 | 0.6 |
| CPT 20 µM + RSV 10 µM | >10 | 0.6 |
| CPT 20 µM + RSV 20 µM | >10 | 1.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, H.; Szyjka, A.; Bęben, D.; Siwiela, O.; Radajewska, A.; Stankiewicz, N.; Grzesiak, M.; Wiatrak, B.; Emhemmed, F.; Muller, C.D.; et al. Genotoxic and Anti-Migratory Effects of Camptothecin Combined with Celastrol or Resveratrol in Metastatic and Stem-like Cells of Colon Cancer. Cancers 2024, 16, 3279. https://doi.org/10.3390/cancers16193279
Moreira H, Szyjka A, Bęben D, Siwiela O, Radajewska A, Stankiewicz N, Grzesiak M, Wiatrak B, Emhemmed F, Muller CD, et al. Genotoxic and Anti-Migratory Effects of Camptothecin Combined with Celastrol or Resveratrol in Metastatic and Stem-like Cells of Colon Cancer. Cancers. 2024; 16(19):3279. https://doi.org/10.3390/cancers16193279
Chicago/Turabian StyleMoreira, Helena, Anna Szyjka, Dorota Bęben, Oliwia Siwiela, Anna Radajewska, Nadia Stankiewicz, Małgorzata Grzesiak, Benita Wiatrak, Fathi Emhemmed, Christian D. Muller, and et al. 2024. "Genotoxic and Anti-Migratory Effects of Camptothecin Combined with Celastrol or Resveratrol in Metastatic and Stem-like Cells of Colon Cancer" Cancers 16, no. 19: 3279. https://doi.org/10.3390/cancers16193279
APA StyleMoreira, H., Szyjka, A., Bęben, D., Siwiela, O., Radajewska, A., Stankiewicz, N., Grzesiak, M., Wiatrak, B., Emhemmed, F., Muller, C. D., & Barg, E. (2024). Genotoxic and Anti-Migratory Effects of Camptothecin Combined with Celastrol or Resveratrol in Metastatic and Stem-like Cells of Colon Cancer. Cancers, 16(19), 3279. https://doi.org/10.3390/cancers16193279