
An Injury-like Signature of the Extracellular Glioma Metabolome

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
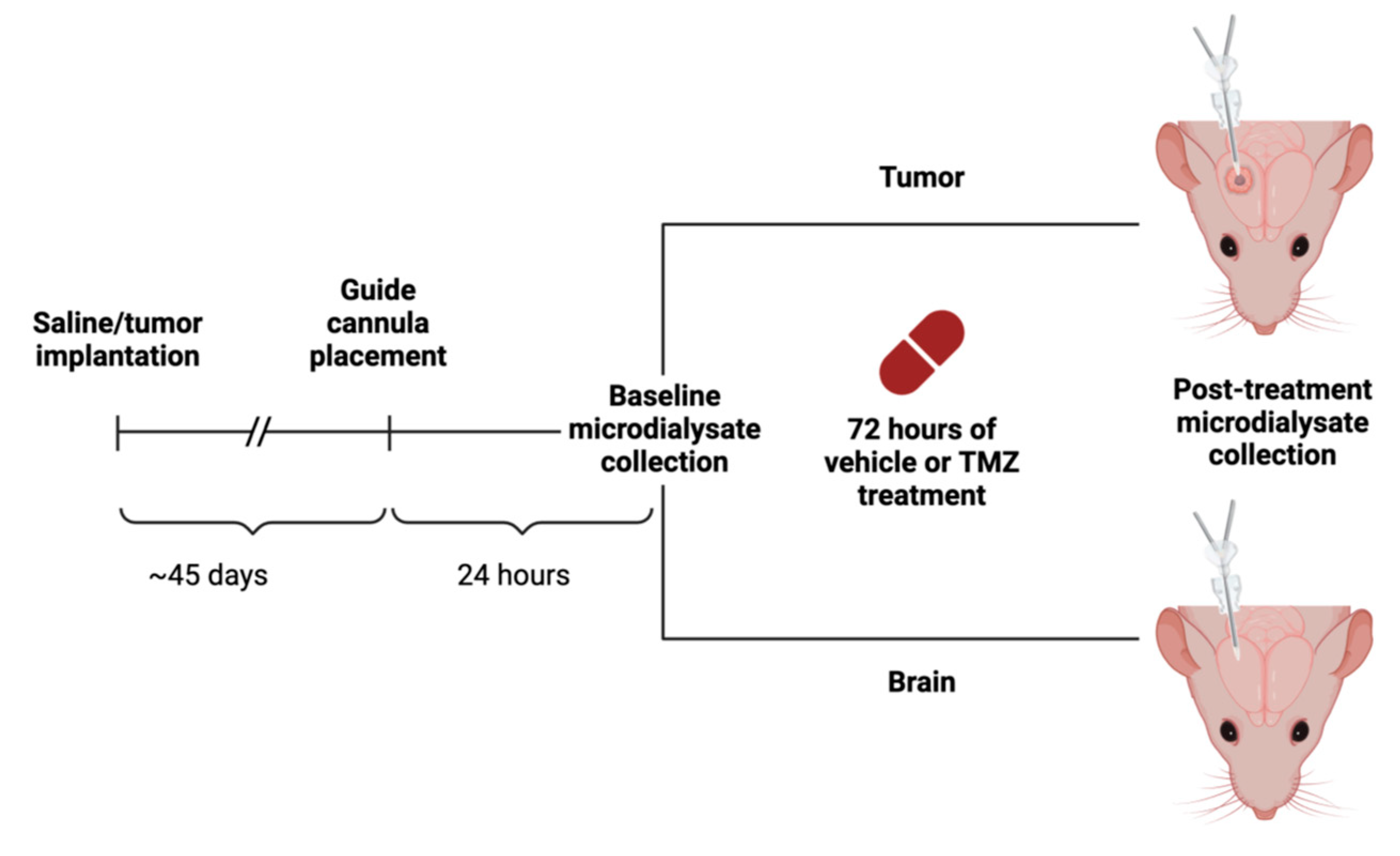
2.1. Ethics Statement
2.2. Intracranial Implantation
2.3. Microdialysis Implantation and Collection
2.4. Untargeted Metabolomic Analysis
2.5. Ranked Metabolite Lists
2.6. Enrichment Analysis
2.7. Statistics
3. Results
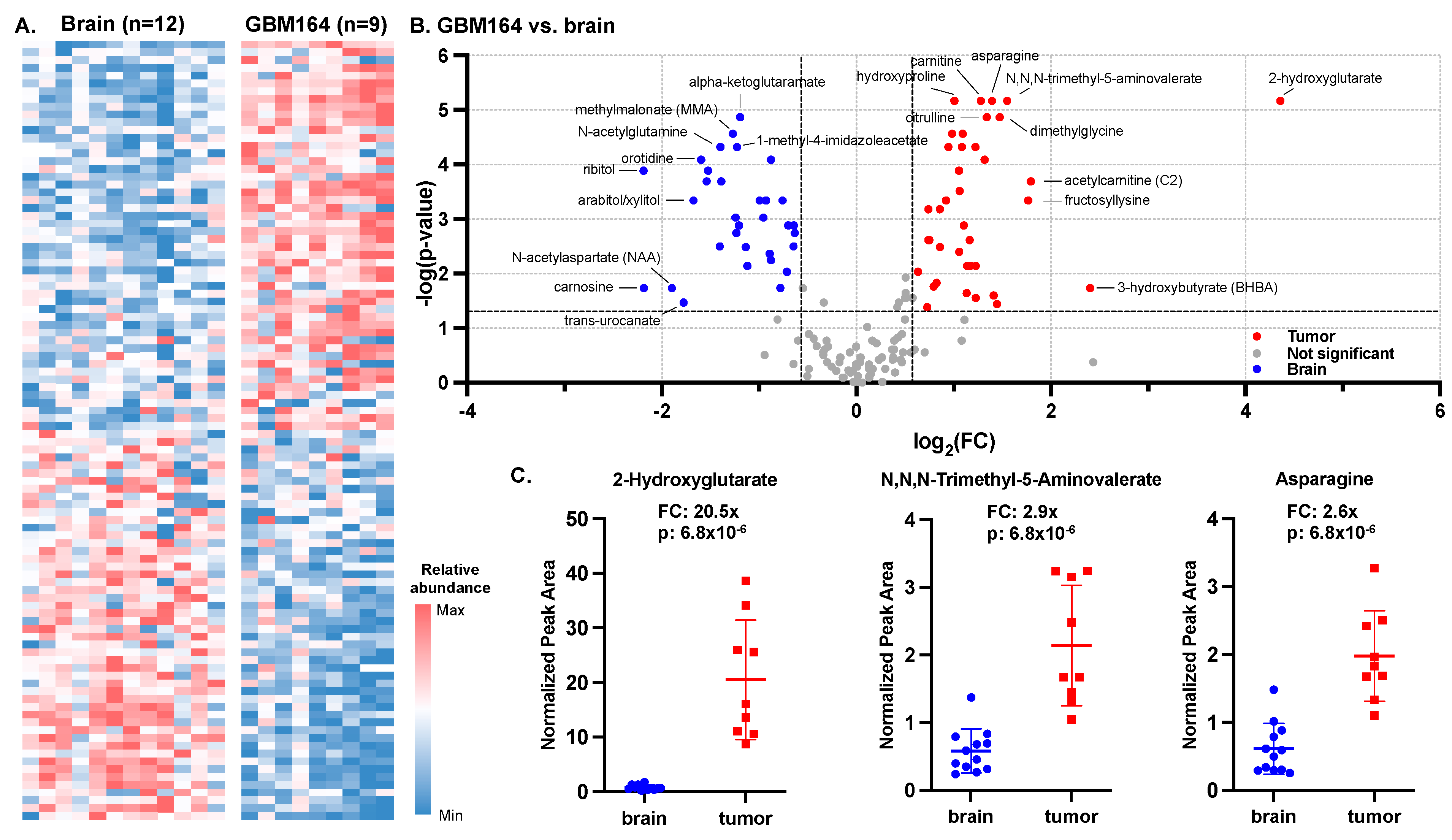
3.1. Metabolic Signature of GBM164, an Orthotopic Model of High-Grade Glioma
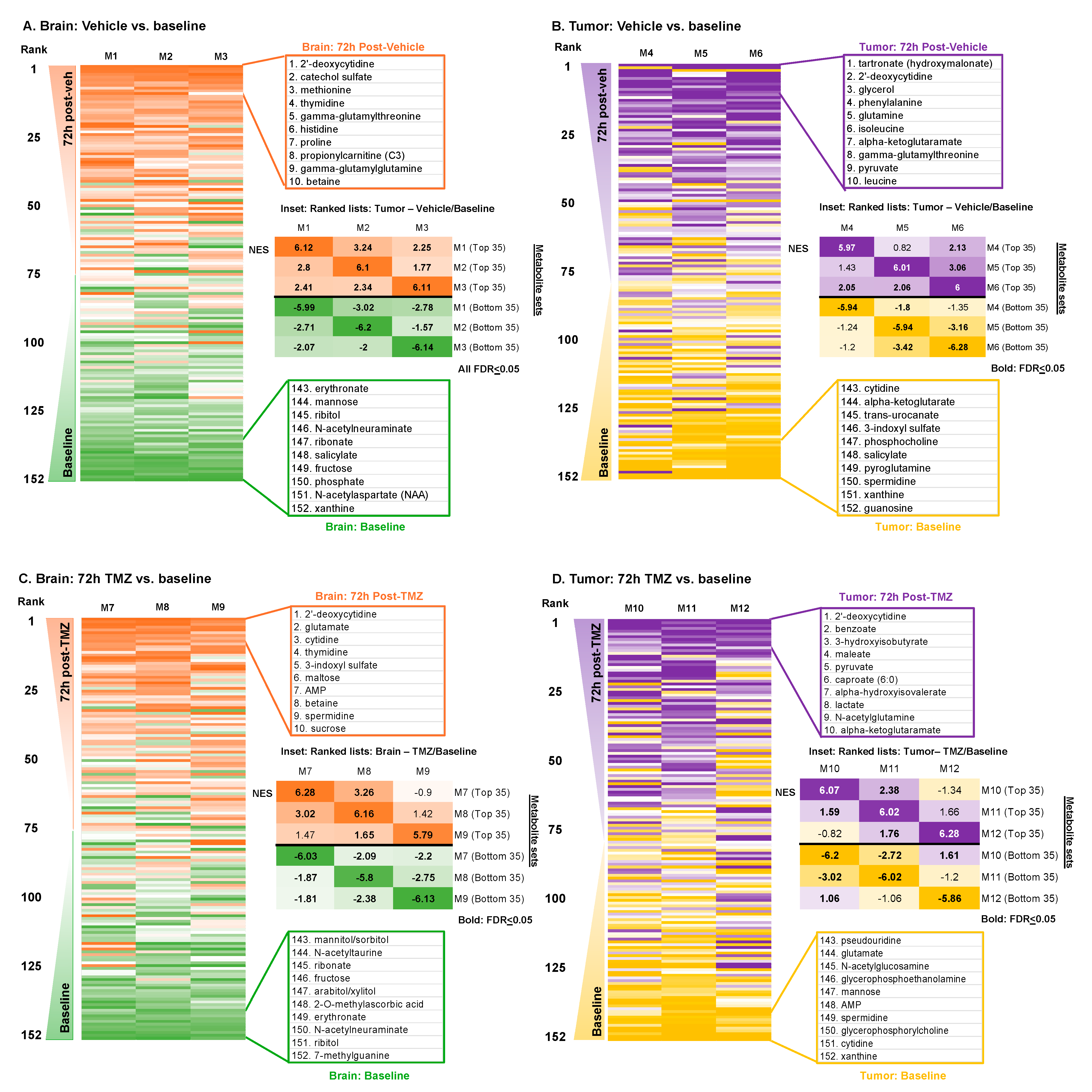
3.2. Metabolic Impacts of Catheter Insertion over Time
3.3. Metabolic Impacts of Catheter Insertion over Time in the Presence of TMZ
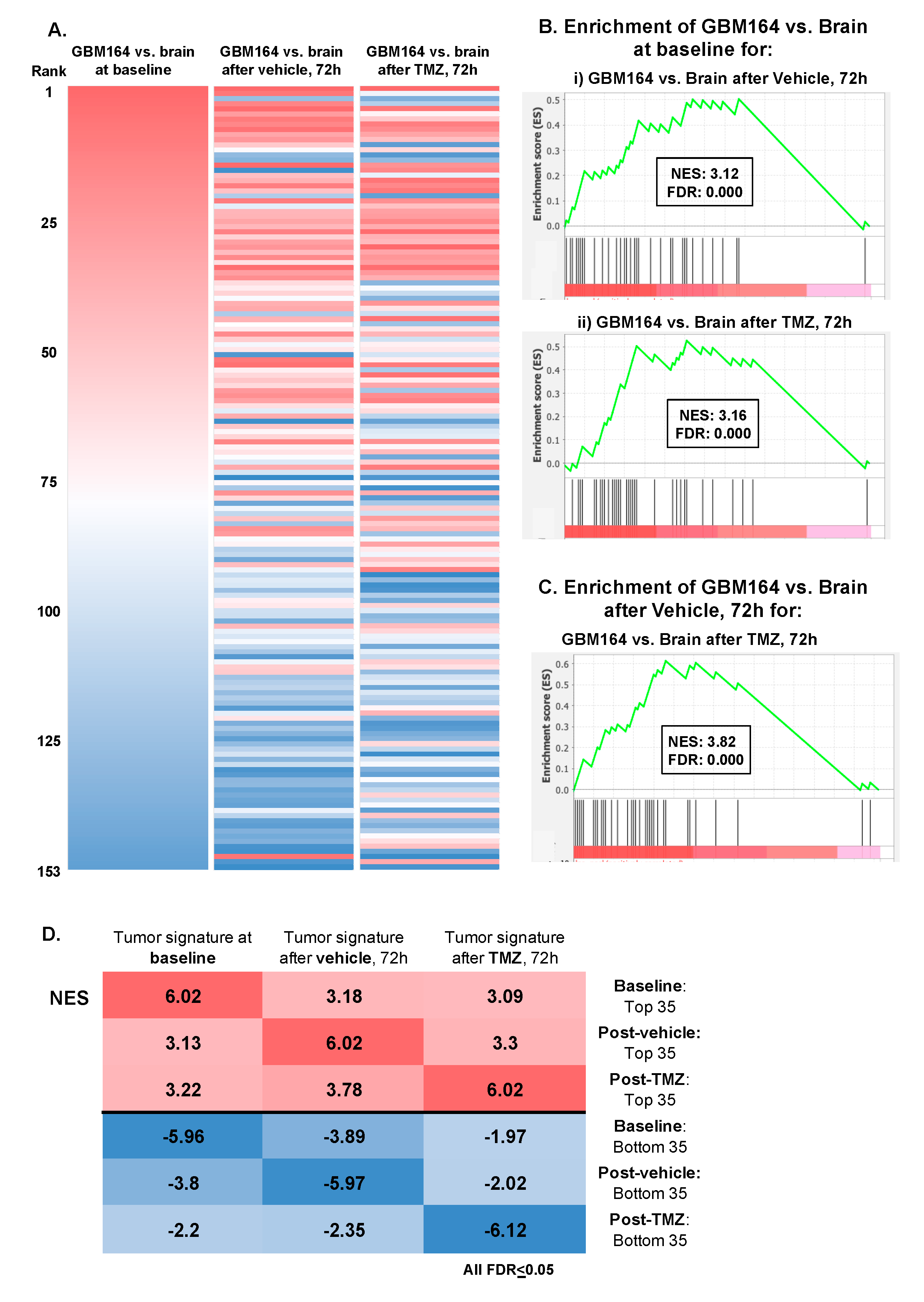
3.4. Impact of TMZ or Catheter Insertion on the Glioma versus Brain Metabolome
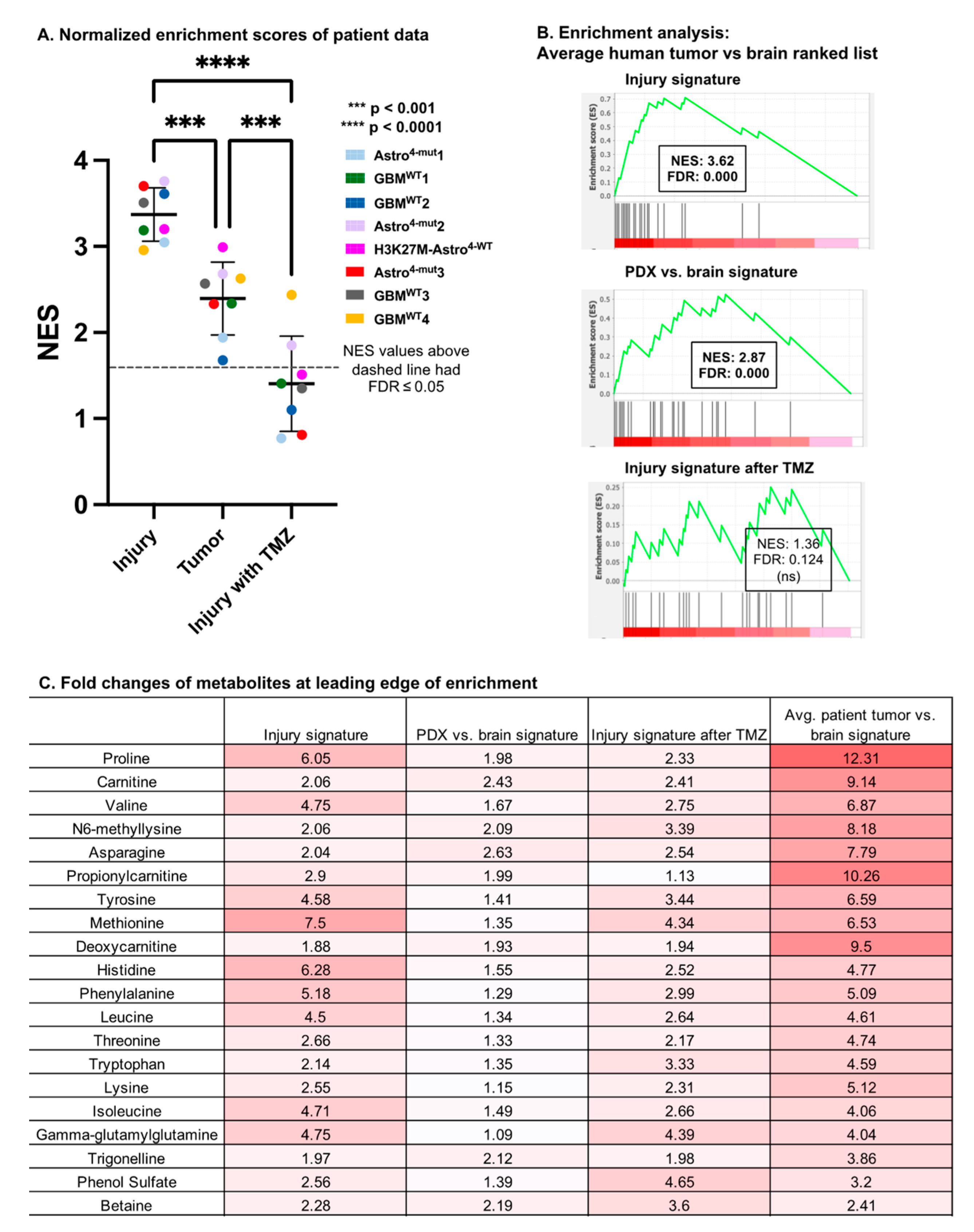
3.5. Comparison with Human Glioma Microdialysate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Actaneuropathologica 2007, 114, 97–109. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, H.; Lee, C.K.; Tateishi, K.; Higuchi, F.; Subramanian, M.; Rafferty, S.; Melamed, L.; Miller, J.J.; Wakimoto, H.; Cahill, D.P. Poly(ADP-ribose) Glycohydrolase Inhibition Sequesters NAD+ to Potentiate the Metabolic Lethality of Alkylating Chemotherapy in IDH-Mutant Tumor Cells. Cancer Discov. 2020, 10, 1672–1689. [Google Scholar] [CrossRef]
- Shi, D.D.; Savani, M.R.; Levitt, M.M.; Wang, A.C.; Endress, J.E.; Bird, C.E.; Buehler, J.; Stopka, S.A.; Regan, M.S.; Lin, Y.-F.; et al. De novo pyrimidine synthesis is a targetable vulnerability in IDH mutant glioma. Cancer Cell 2022, 40, 939–956.e16. [Google Scholar] [CrossRef] [PubMed]
- Björkblom, B.; Jonsson, P.; Tabatabaei, P.; Bergström, P.; Johansson, M.; Asklund, T.; Bergenheim, A.T.; Antti, H. Metabolic response patterns in brain microdialysis fluids and serum during interstitial cisplatin treatment of high-grade glioma. Br. J. Cancer 2019, 122, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Johansen, M.J.; Newman, R.A.; Madden, T. The Use of Microdialysis in Pharmacokinetics and Pharmacodynamics. Pharmacother. J. Hum. Pharmacol. Drug Ther. 1997, 17, 464–481. [Google Scholar] [CrossRef]
- Riviere-Cazaux, C.; Carlstrom, L.P.; Rajani, K.; Munoz-Casabella, A.; Rahman, M.; Gharibi-Loron, A.; Brown, D.A.; Miller, K.J.; White, J.J.; Himes, B.T.; et al. Blood-brain barrier disruption defines the extracellular metabolome of live human high-grade gliomas. Commun. Biol. 2023, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1 and IDH2 Mutations in Gliomas. N. Engl. J. Med. 2009, 360, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Vaubel, R.A.; Tian, S.; Remonde, D.; Schroeder, M.A.; Mladek, A.C.; Kitange, G.J.; Caron, A.; Kollmeyer, T.M.; Grove, R.; Peng, S.; et al. Genomic and Phenotypic Characterization of a Broad Panel of Patient-Derived Xenografts Reflects the Diversity of Glioblastoma. Clin. Cancer Res. 2020, 26, 1094–1104. [Google Scholar] [CrossRef]
- Rajani, K.; Olson, I.; Jacobs, J.J.; Riviere-Cazaux, C.; Burns, K.; Carlstrom, L.; Schroeder, M.; Oh, J.; Howe, C.L.; Rahman, M.; et al. Methods for intratumoral microdialysis probe targeting and validation in murine brain tumor models. J. Neurosci. Methods 2021, 363, 109321. [Google Scholar] [CrossRef]
- Björkblom, B.; Wibom, C.; Eriksson, M.; Bergenheim, A.T.; Sjöberg, R.L.; Jonsson, P.; Brännström, T.; Antti, H.; Sandström, M.; Melin, B. Distinct metabolic hallmarks of WHO classified adult glioma subtypes. Neuro-Oncology 2022, 24, 1454–1468. [Google Scholar] [CrossRef] [PubMed]
- Sarkaria, J.N. Mayo Clinic Brain Tumor Patient-Derived Xenograft National Resource; Mayo Clinic: Rochester, MN, USA, 2022; Available online: https://www.mayo.edu/research/labs/translational-neuro-oncology/mayo-clinic-brain-tumor-patient-derived-xenograft-national-resource/overview (accessed on 1 August 2022).
- Carlson, B.L.; Pokorny, J.L.; Schroeder, M.A.; Sarkaria, J.N. Establishment, Maintenance, and In Vitro and In Vivo Applications of Primary Human Glioblastoma Multiforme (GBM) Xenograft Models for Translational Biology Studies and Drug Discovery. Curr. Protoc. Pharmacol. 2011, 52, 14.16.1–14.16.23. [Google Scholar] [CrossRef]
- Ryals, J.; Lawton, K.; Stevens, D.; Milburn, M. Metabolon, Inc. Pharmacogenomics 2007, 8, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Waitkus, M.S.; Diplas, B.H.; Yan, H. Isocitrate dehydrogenase mutations in gliomas. Neuro-Oncology 2015, 18, 16–26. [Google Scholar] [CrossRef]
- Roberts, W.G.; Palade, G.E. Neovasculature induced by vascular endothelial growth factor is fenestrated. Cancer Res. 1997, 57, 765–772. [Google Scholar]
- Portnow, J.; Badie, B.; Chen, M.; Liu, A.; Blanchard, S.; Synold, T.W. The Neuropharmacokinetics of Temozolomide in Patients with Resectable Brain Tumors: Potential Implications for the Current Approach to Chemoradiation. Clin. Cancer Res. 2009, 15, 7092–7098. [Google Scholar] [CrossRef]
- Tabatabaei, P.; Asklund, T.; Bergström, P.; Björn, E.; Johansson, M.; Bergenheim, A.T. Intratumoral retrograde microdialysis treatment of high-grade glioma with cisplatin. Acta Neurochir. 2020, 162, 3043–3053. [Google Scholar] [CrossRef]
- Krum, J.M.; Khaibullina, A. Inhibition of endogenous VEGF impedes revascularization and astroglial proliferation: Roles for VEGF in brain repair. Exp. Neurol. 2003, 181, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Rosenstein, J.M.; Krum, J.M.; Ruhrberg, C. VEGF in the nervous system. Organogenesis 2010, 6, 107–114. [Google Scholar] [CrossRef]
- Dvorak, H.F. Tumors: Wounds That Do Not Heal—A Historical Perspective with a Focus on the Fundamental Roles of Increased Vascular Permeability and Clotting. Semin. Thromb. Hemost. 2019, 45, 576–592. [Google Scholar] [CrossRef]
- Kreisl, T.N.; Kim, L.; Moore, K.; Duic, P.; Royce, C.; Stroud, I.; Garren, N.; Mackey, M.; Butman, J.A.; Camphausen, K.; et al. Phase II Trial of Single-Agent Bevacizumab Followed by Bevacizumab Plus Irinotecan at Tumor Progression in Recurrent Glioblastoma. J. Clin. Oncol. 2009, 27, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.J.; Burowski, N.A.; Chang, S.M.; Prados, M.D.; Clarke, J.; Polley, M.Y.C.; Sughrue, M.E.; McDermott, M.W.; Parsa, A.T.; Berger, M.S.; et al. Impact of bevacizumab chemotherapy on craniotomy wound healing Clinical article. J. Neurosurg. 2011, 114, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Riviere-Cazaux, C.; Neth, B.J.; Hoplin, M.D.; Wessel, B.; Miska, J.; Kizilbash, S.H.; Burns, T.C. Glioma Metabolic Feedback In Situ: A First-In-Human Pharmacodynamic Trial of Difluoromethylornithine + AMXT-1501 Through High–Molecular Weight Microdialysis. Neurosurgery 2023, 93, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Blakeley, J.O.; Olson, J.; Grossman, S.A.; He, X.; Weingart, J.; Supko, J.G. For the New Approaches to Brain Tumor Therapy (NABTT) Consortium Effect of blood brain barrier permeability in recurrent high grade gliomas on the intratumoral pharmacokinetics of methotrexate: A microdialysis study. J. Neuro-Oncol. 2008, 91, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Lynes, J.; Jackson, S.; Sanchez, V.; Dominah, G.; Wang, X.; Kuek, A.; Hayes, C.P.; Benzo, S.; Scott, G.C.; Chittiboina, P.; et al. Cytokine Microdialysis for Real-Time Immune Monitoring in Glioblastoma Patients Undergoing Checkpoint Blockade. Neurosurgery 2018, 84, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.; Weingart, J.; Nduom, E.K.; Harfi, T.T.; George, R.T.; McAreavey, D.; Ye, X.; Anders, N.M.; Peer, C.; Figg, W.D.; et al. The effect of an adenosine A2A agonist on intra-tumoral concentrations of temozolomide in patients with recurrent glioblastoma. Fluids Barriers CNS 2018, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Sottoriva, A.; Spiteri, I.; Piccirillo, S.G.M.; Touloumis, A.; Collins, V.P.; Marioni, J.C.; Curtis, C.; Watts, C.; Tavaré, S. Intratumor heterogeneity in human glioblastoma reflects cancer evolutionary dynamics. Proc. Natl. Acad. Sci. USA 2013, 110, 4009–4014. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.P.; Tirosh, I.; Trombetta, J.J.; Shalek, A.K.; Gillespie, S.M.; Wakimoto, H.; Cahill, D.P.; Nahed, B.V.; Curry, W.T.; Martuza, R.L.; et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 2014, 344, 1396–1401. [Google Scholar] [CrossRef]
- Karve, I.P.; Taylor, J.M.; Crack, P.J. The contribution of astrocytes and microglia to traumatic brain injury. Br. J. Pharmacol. 2015, 173, 692–702. [Google Scholar] [CrossRef]
- Banks, W.A.; Gray, A.M.; Erickson, M.A.; Salameh, T.S.; Damodarasamy, M.; Sheibani, N.; Meabon, J.S.; Wing, E.E.; Morofuji, Y.; Cook, D.G.; et al. Lipopolysaccharide-induced blood-brain barrier disruption: Roles of cyclooxygenase, oxidative stress, neuroinflammation, and elements of the neurovascular unit. J. Neuroinflamm. 2015, 12, 223. [Google Scholar] [CrossRef]
- Erickson, M.A.; Dohi, K.; Banks, W.A. Neuroinflammation: A Common Pathway in CNS Diseases as Mediated at the Blood-Brain Barrier. Neuroimmunomodulation 2012, 19, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Toyooka, T.; Nawashiro, H.; Shinomiya, N.; Yano, A.; Ooigawa, H.; Ohsumi, A.; Uozumi, Y.; Yanagawa, Y.; Matsuo, H.; Shima, K. Up-regulation of L type amino acid transporter 1 after spinal cord injury in rats. Acta Neurochir. Suppl. 2008, 102, 385–388. [Google Scholar] [PubMed]
- Nawashiro, H.; Otani, N.; Shinomiya, N.; Fukui, S.; Ooigawa, H.; Shima, K.; Matsuo, H.; Kanai, Y.; Endou, H. L-type amino acid transporter 1 as a potential molecular target in human astrocytic tumors. Int. J. Cancer 2006, 119, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaei, P.; Visse, E.; Bergström, P.; Brännström, T.; Siesjö, P.; Bergenheim, A.T. Radiotherapy induces an immediate inflammatory reaction in malignant glioma: A clinical microdialysis study. J. Neuro-Oncol. 2016, 131, 83–92. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, Y.; Rajani, K.; Riviere-Cazaux, C.; Rahman, M.; Olson, I.E.; Gharibi Loron, A.; Schroeder, M.A.; Rodriguez, M.; Warrington, A.E.; Burns, T.C. An Injury-like Signature of the Extracellular Glioma Metabolome. Cancers 2024, 16, 2705. https://doi.org/10.3390/cancers16152705
Ha Y, Rajani K, Riviere-Cazaux C, Rahman M, Olson IE, Gharibi Loron A, Schroeder MA, Rodriguez M, Warrington AE, Burns TC. An Injury-like Signature of the Extracellular Glioma Metabolome. Cancers. 2024; 16(15):2705. https://doi.org/10.3390/cancers16152705
Chicago/Turabian StyleHa, Yooree, Karishma Rajani, Cecile Riviere-Cazaux, Masum Rahman, Ian E. Olson, Ali Gharibi Loron, Mark A. Schroeder, Moses Rodriguez, Arthur E. Warrington, and Terry C. Burns. 2024. "An Injury-like Signature of the Extracellular Glioma Metabolome" Cancers 16, no. 15: 2705. https://doi.org/10.3390/cancers16152705
APA StyleHa, Y., Rajani, K., Riviere-Cazaux, C., Rahman, M., Olson, I. E., Gharibi Loron, A., Schroeder, M. A., Rodriguez, M., Warrington, A. E., & Burns, T. C. (2024). An Injury-like Signature of the Extracellular Glioma Metabolome. Cancers, 16(15), 2705. https://doi.org/10.3390/cancers16152705