

Radium-223 Treatment Produces Prolonged Suppression of Resident Osteoblasts and Decreased Bone Mineral Density in Trabecular Bone in Osteoblast Reporter Mice

 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Treatment of Mice with Ra-223 and Bioluminescence Measurement
2.3. µCT Analysis and GFP Quantification
2.4. Generation of TRAMP-BMP4 Cell Line
2.5. Mice with TRAMP-BMP4 Tumors
2.6. Statistical Analysis
3. Results
3.1. Ra-223 Treatment Decreases Tail Bone Luciferase Activity in a Col-Luc Osteoblast Reporter Mice
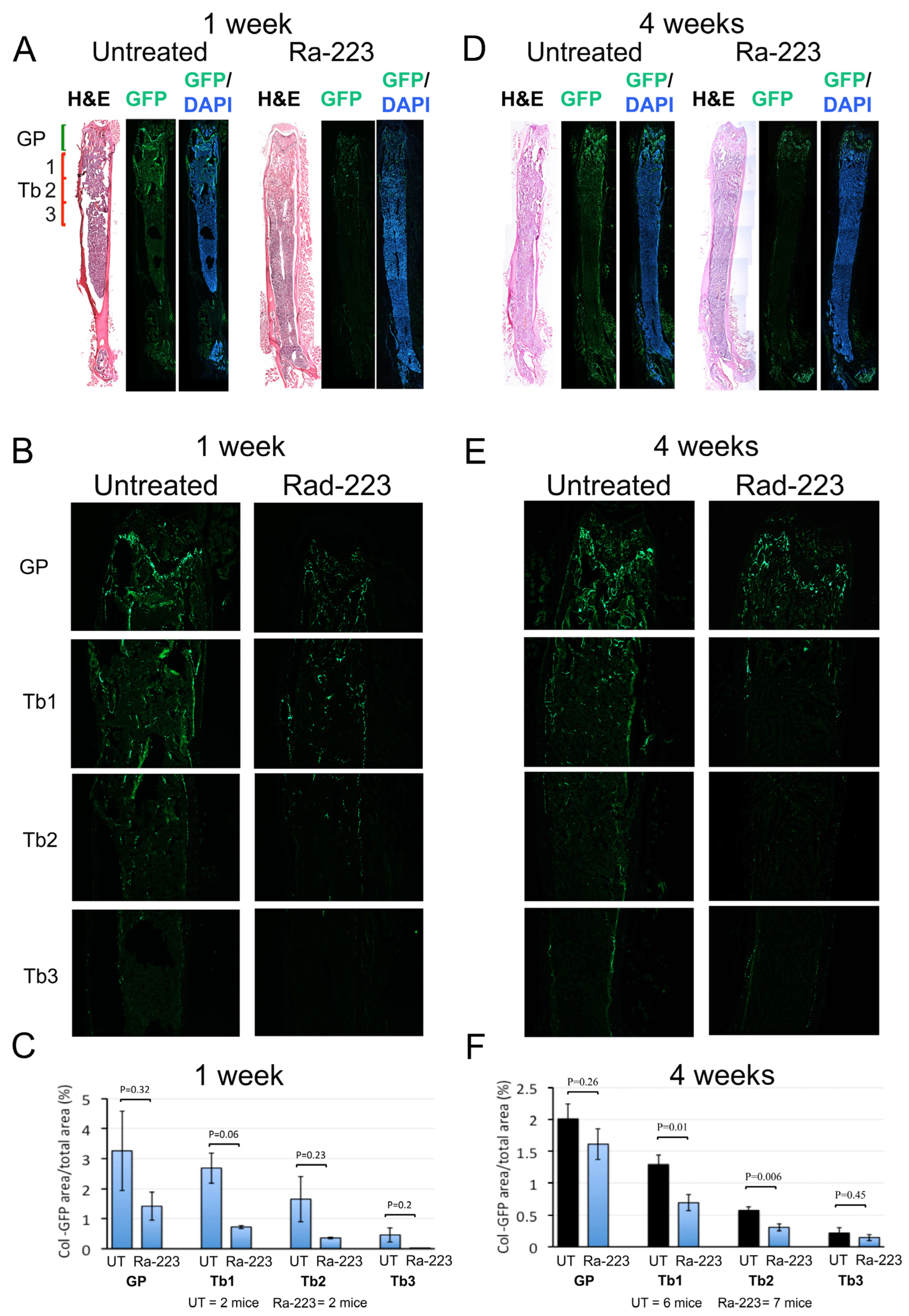
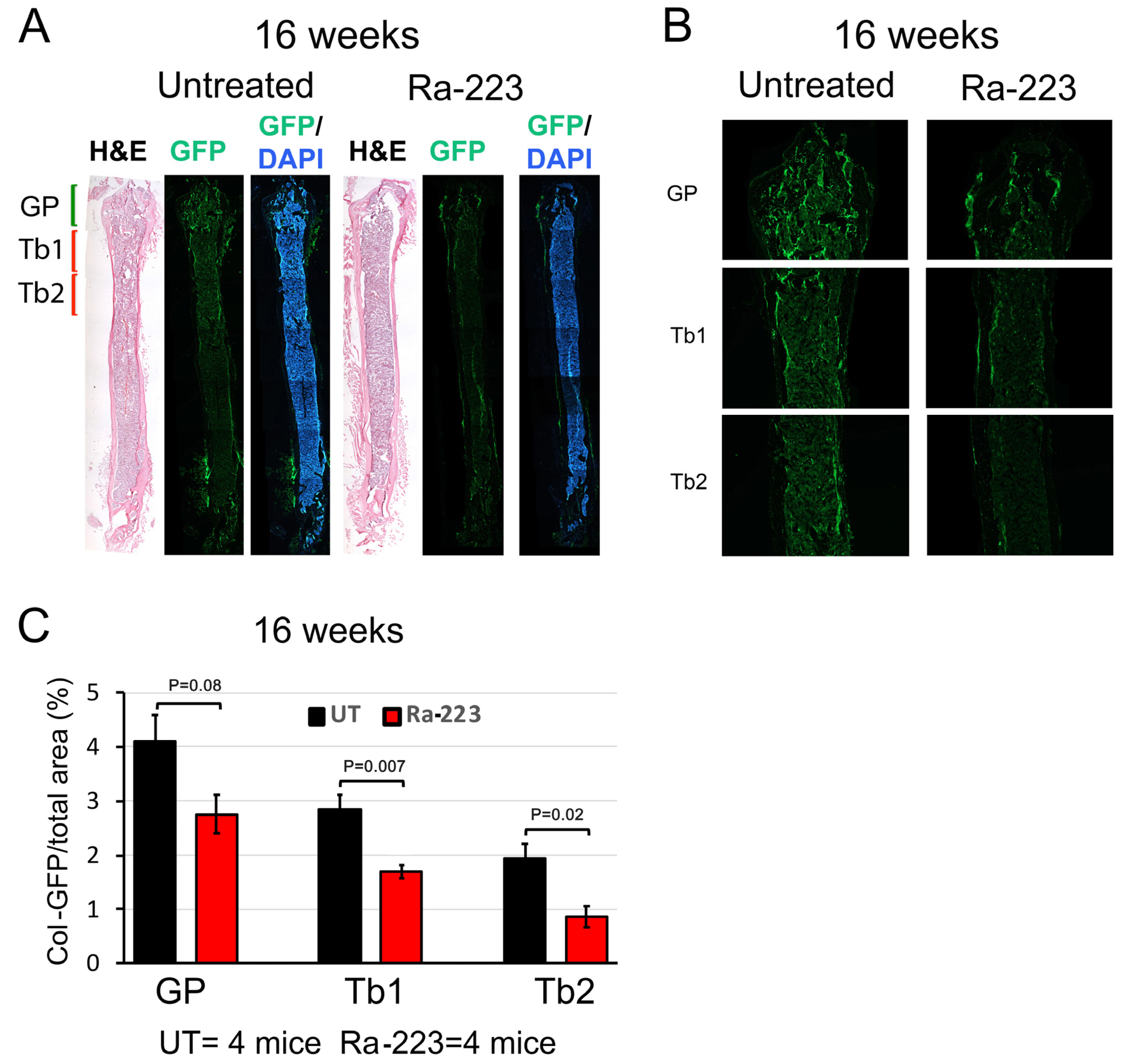
3.2. Ra-223 Decreases GFP-Labeled Osteoblasts in Col-GFP Mice
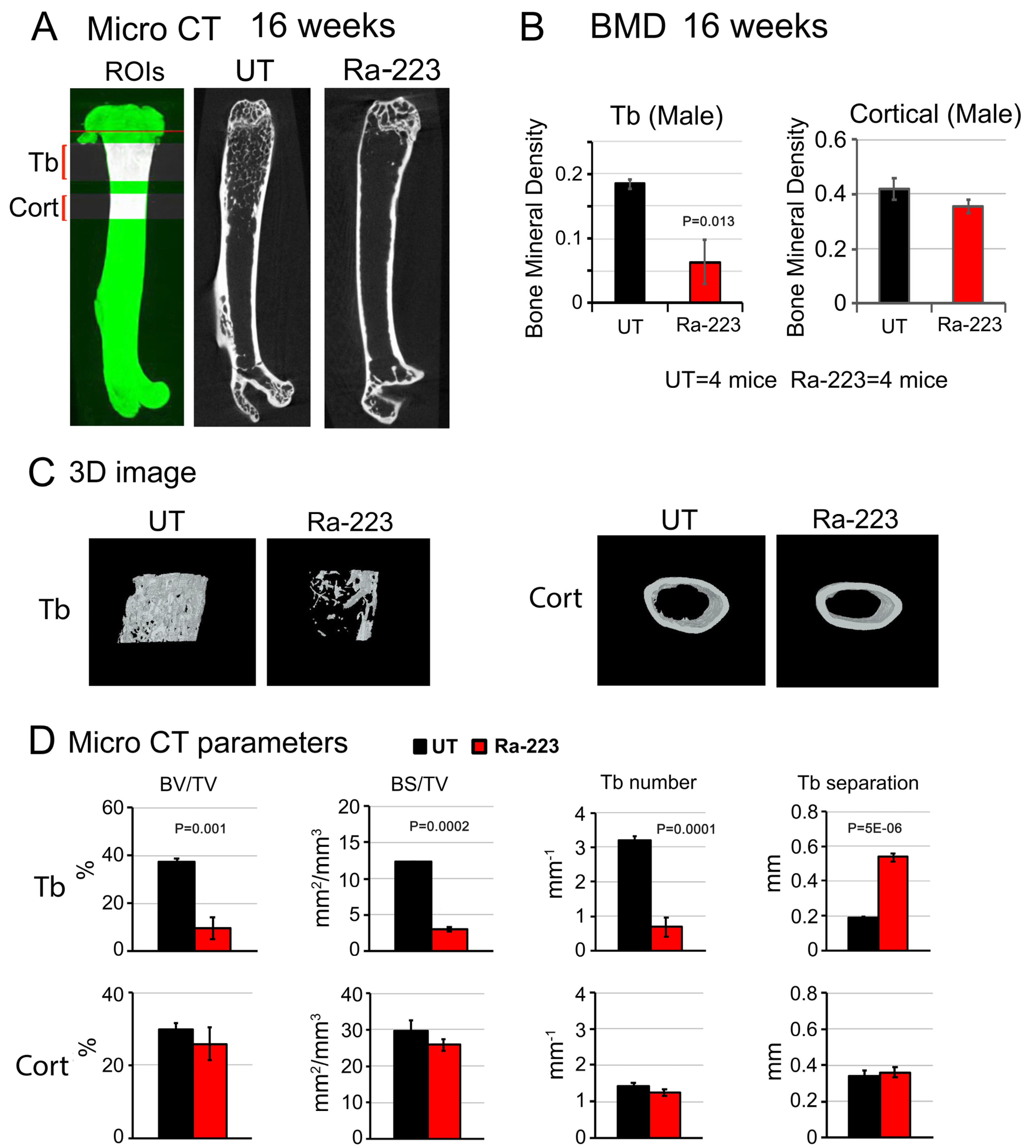
3.3. Ra-223 Decreases Bone Mineral Density of Col-GFP Mice
3.4. Ra-223 Decreases Tumor-Induced Osteoblasts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bubendorf, L.; Schopfer, A.; Wagner, U.; Sauter, G.; Moch, H.; Willi, N.; Gasser, T.C.; Mihatsch, M.J. Metastatic patterns of prostate cancer: An autopsy study of 1589 patients. Hum. Pathol. 2000, 31, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Cheng, C.J.; Bilen, M.A.; Lu, J.F.; Satcher, R.L.; Yu-Lee, L.Y.; Gallick, G.E.; Maity, S.N.; Lin, S.H. BMP4 promotes prostate tumor growth in bone through osteogenesis. Cancer Res. 2011, 71, 5194–5203. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Lin, S.C.; Yu, G.; Zhu, M.; Song, J.H.; Rivera, K.; Pappin, D.J.; Logothetis, C.J.; Panaretakis, T.; Wang, G.; et al. Prostate tumor-induced stromal reprogramming generates Tenascin C that promotes prostate cancer metastasis through YAP/TAZ inhibition. Oncogene 2022, 41, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Lee, Y.C.; Yu, G.; Cheng, C.J.; Zhou, X.; Chu, K.; Murshed, M.; Le, N.T.; Baseler, L.; Abe, J.I.; et al. Endothelial-to-Osteoblast Conversion Generates Osteoblastic Metastasis of Prostate Cancer. Dev. Cell 2017, 41, 467–480 e463. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Hall, C.L.; Escara-Wilke, J.; Mizokami, A.; Keller, J.M.; Keller, E.T. Prostate cancer induces bone metastasis through Wnt-induced bone morphogenetic protein-dependent and independent mechanisms. Cancer Res. 2008, 68, 5785–5794. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Keller, J.; Zhang, J.; Lu, Y.; Yao, Z.; Keller, E.T. Bone morphogenetic protein-6 promotes osteoblastic prostate cancer bone metastases through a dual mechanism. Cancer Res. 2005, 65, 8274–8285. [Google Scholar] [CrossRef]
- Fizazi, K.; Yang, J.; Peleg, S.; Sikes, C.R.; Kreimann, E.L.; Daliani, D.; Olive, M.; Raymond, K.A.; Janus, T.J.; Logothetis, C.J.; et al. Prostate cancer cells-osteoblast interaction shifts expression of growth/survival-related genes in prostate cancer and reduces expression of osteoprotegerin in osteoblasts. Clin. Cancer Res. 2003, 9, 2587–2597. [Google Scholar] [PubMed]
- Ribelli, G.; Simonetti, S.; Iuliani, M.; Rossi, E.; Vincenzi, B.; Tonini, G.; Pantano, F.; Santini, D. Osteoblasts Promote Prostate Cancer Cell Proliferation Through Androgen Receptor Independent Mechanisms. Front. Oncol. 2021, 11, 789885. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Lin, S.C.; Yu, G.; Cheng, C.J.; Liu, B.; Liu, H.C.; Hawke, D.H.; Parikh, N.U.; Varkaris, A.; Corn, P.; et al. Identification of Bone-Derived Factors Conferring De Novo Therapeutic Resistance in Metastatic Prostate Cancer. Cancer Res. 2015, 75, 4949–4959. [Google Scholar] [CrossRef]
- Bilen, M.A.; Johnson, M.M.; Mathew, P.; Pagliaro, L.C.; Araujo, J.C.; Aparicio, A.; Corn, P.G.; Tannir, N.M.; Wong, F.C.; Fisch, M.J.; et al. Randomized phase 2 study of bone-targeted therapy containing strontium-89 in advanced castrate-sensitive prostate cancer. Cancer 2015, 121, 69–76. [Google Scholar] [CrossRef]
- Tu, S.-M.; Delpassand, E.S.; Jones, D.; Amato, R.J.; Ellerhorst, J.; Logothetis, C.J. Strontium-89 combined with doxorubicin in the treatment of patients with androgen-independent prostate cancer. Urol. Oncol. 1996, 2, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Sartor, O.; Reid, R.H.; Hoskin, P.J.; Quick, D.P.; Ell, P.J.; Coleman, R.E.; Kotler, J.A.; Freeman, L.M.; Olivier, P. Quadramet 424Sm10/11 Study Group. Samarium-153-Lexidronam complex for treatment of painful bone metastases in hormone-refractory prostate cancer. Urology 2004, 63, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Serafini, A.N.; Houston, S.J.; Resche, I.; Quick, D.P.; Grund, F.M.; Ell, P.J.; Bertrand, A.; Ahmann, F.R.; Orihuela, E.; Reid, R.H.; et al. Palliation of pain associated with metastatic bone cancer using samarium-153 lexidronam: A double-blind placebo-controlled clinical trial. J. Clin. Oncol. 1998, 16, 1574–1581. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P. Samarium for osteoblastic bone metastases and osteosarcoma. Expert. Opin. Pharmacother. 2006, 7, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, G.; Fisher, D.R.; Roeske, J.C.; Bruland, O.S.; Larsen, R.H. Targeting of osseous sites with alpha-emitting 223Ra: Comparison with the beta-emitter 89Sr in mice. J. Nucl. Med. 2003, 44, 252–259. [Google Scholar] [PubMed]
- Parker, C.; Nilsson, S.; Heinrich, D.; Helle, S.I.; O‘Sullivan, J.M.; Fossa, S.D.; Chodacki, A.; Wiechno, P.; Logue, J.; Seke, M.; et al. Alpha emitter radium-223 and survival in metastatic prostate cancer. N. Engl. J. Med. 2013, 369, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Hijab, A.; Curcean, S.; Tunariu, N.; Tovey, H.; Alonzi, R.; Staffurth, J.; Blackledge, M.; Padhani, A.; Tree, A.; Stidwill, H.; et al. Fracture Risk in Men with Metastatic Prostate Cancer Treated With Radium-223. Clin. Genitourin. Cancer 2021, 19, e299–e305. [Google Scholar] [CrossRef]
- Smith, M.; Parker, C.; Saad, F.; Miller, K.; Tombal, B.; Ng, Q.S.; Boegemann, M.; Matveev, V.; Piulats, J.M.; Zucca, L.E.; et al. Addition of radium-223 to abiraterone acetate and prednisone or prednisolone in patients with castration-resistant prostate cancer and bone metastases (ERA 223): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Oncol. 2019, 20, 408–419. [Google Scholar] [CrossRef]
- Paindelli, C.; Casarin, S.; Wang, F.; Diaz-Gomez, L.; Zhang, J.; Mikos, A.G.; Logothetis, C.J.; Friedl, P.; Dondossola, E. Enhancing (223)Ra Treatment Efficacy by Anti-beta1 Integrin Targeting. J. Nucl. Med. 2022, 63, 1039–1045. [Google Scholar] [CrossRef]
- Vardaki, I.; Corn, P.; Gentile, E.; Song, J.H.; Madan, N.; Hoang, A.; Parikh, N.; Guerra, L.; Lee, Y.C.; Lin, S.C.; et al. Radium-223 Treatment Increases Immune Checkpoint Expression in Extracellular Vesicles from the Metastatic Prostate Cancer Bone Microenvironment. Clin. Cancer Res. 2021, 27, 3253–3264. [Google Scholar] [CrossRef]
- Suominen, M.I.; Fagerlund, K.M.; Rissanen, J.P.; Konkol, Y.M.; Morko, J.P.; Peng, Z.; Alhoniemi, E.J.; Laine, S.K.; Corey, E.; Mumberg, D.; et al. Radium-223 Inhibits Osseous Prostate Cancer Growth by Dual Targeting of Cancer Cells and Bone Microenvironment in Mouse Models. Clin. Cancer Res. 2017, 23, 4335–4346. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Huang, C.F.; Murshed, M.; Chu, K.; Araujo, J.C.; Ye, X.; deCrombrugghe, B.; Yu-Lee, L.Y.; Gallick, G.E.; Lin, S.H. Src family kinase/abl inhibitor dasatinib suppresses proliferation and enhances differentiation of osteoblasts. Oncogene 2010, 29, 3196–3207. [Google Scholar] [CrossRef] [PubMed]
- Foster, B.A.; Gingrich, J.R.; Kwon, E.D.; Madias, C.; Greenberg, N.M. Characterization of prostatic epithelial cell lines derived from transgenic adenocarcinoma of the mouse prostate (TRAMP) model. Cancer Res. 1997, 57, 3325–3330. [Google Scholar]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Rossert, J.; Eberspaecher, H.; de Crombrugghe, B. Separate cis-acting DNA elements of the mouse pro-alpha 1(I) collagen promoter direct expression of reporter genes to different type I collagen-producing cells in transgenic mice. J. Cell Biol. 1995, 129, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, S.C. Spread of prostatic cancer to bone. Urology 1983, 21, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Charhon, S.A.; Chapuy, M.C.; Delvin, E.E.; Valentin-Opran, A.; Edouard, C.M.; Meunier, P.J. Histomorphometric analysis of sclerotic bone metastases from prostatic carcinoma special reference to osteomalacia. Cancer 1983, 51, 918–924. [Google Scholar] [CrossRef] [PubMed]
- Logothetis, C.; Lin, S.-H. Osteoblasts in prostate cancer metastasis to bone. Nat. Rev. Cancer 2005, 5, 21–28. [Google Scholar] [CrossRef]
- Li, Z.; Mathew, P.; Yang, J.; Starbuck, M.-W.; Zurita, A.J.; Liu, J.; Sikes, C.; Multani, A.S.; Efstathiou, E.; Lopez, A.; et al. Androgen receptor–negative human prostate cancer cells induce osteogenesis through FGF9-mediated mechanisms. J. Clin. Investig. 2008, 118, 2697–2710. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Vessella, R.L.; Morrissey, C.; Brown, L.G.; Coleman, I.M.; Higano, C.S.; Mostaghel, E.A.; Zhang, X.; True, L.D.; Lam, H.M.; et al. LuCaP Prostate Cancer Patient-Derived Xenografts Reflect the Molecular Heterogeneity of Advanced Disease and Serve as Models for Evaluating Cancer Therapeutics. Prostate 2017, 77, 654–671. [Google Scholar] [CrossRef]
- Greenberg, N.M.; DeMayo, F.; Finegold, M.J.; Medina, D.; Tilley, W.D.; Aspinall, J.O.; Cunha, G.R.; Donjacour, A.A.; Matusik, R.J.; Rosen, J.M. Prostate cancer in a transgenic mouse. Proc. Natl. Acad. Sci. USA 1995, 92, 3439–3443. [Google Scholar] [CrossRef]
- Yu, G.; Corn, P.G.; Shen, P.; Song, J.H.; Lee, Y.C.; Lin, S.C.; Pan, J.; Agarwal, S.K.; Panaretakis, T.; Pacifici, M.; et al. Retinoic Acid Receptor Activation Reduces Metastatic Prostate Cancer Bone Lesions by Blocking the Endothelial-to-Osteoblast Transition. Cancer Res. 2022, 82, 3158–3171. [Google Scholar] [CrossRef]
- Al-Bari, A.A.; Al Mamun, A. Current advances in regulation of bone homeostasis. FASEB Bioadv. 2020, 2, 668–679. [Google Scholar] [CrossRef]
- Corral, D.A.; Amling, M.; Priemel, M.; Loyer, E.; Fuchs, S.; Ducy, P.; Baron, R.; Karsenty, G. Dissociation between bone resorption and bone formation in osteopenic transgenic mice. Proc. Natl. Acad. Sci. USA 1998, 95, 13835–13840. [Google Scholar] [CrossRef]
- Suominen, M.I.; Rissanen, J.P.; Kakonen, R.; Fagerlund, K.M.; Alhoniemi, E.; Mumberg, D.; Ziegelbauer, K.; Halleen, J.M.; Kakonen, S.M.; Scholz, A. Survival benefit with radium-223 dichloride in a mouse model of breast cancer bone metastasis. J. Natl. Cancer Inst. 2013, 105, 908–916. [Google Scholar] [CrossRef]
- Abou, D.S.; Ulmert, D.; Doucet, M.; Hobbs, R.F.; Riddle, R.C.; Thorek, D.L. Whole-Body and Microenvironmental Localization of Radium-223 in Naive and Mouse Models of Prostate Cancer Metastasis. J. Natl. Cancer Inst. 2016, 108, djv380. [Google Scholar] [CrossRef]
- Pan, T.; Lin, S.C.; Lee, Y.C.; Yu, G.; Song, J.H.; Pan, J.; Titus, M.; Satcher, R.L.; Panaretakis, T.; Logothetis, C.; et al. Statins reduce castration-induced bone marrow adiposity and prostate cancer progression in bone. Oncogene 2021, 40, 4592–4603. [Google Scholar] [CrossRef]
- Trieu, J.; Chang, M.; Rojas, V.; Varada, N.; Cao, Y.; Anderson, M.; Vogelzang, N.J. Lower Fracture Rates in Patients Treated with Radium-223, Abiraterone or Enzalutamide, When Given Concurrently with Bone Health Agents: A Real-World Analysis. Clin. Genitourin. Cancer 2022, 20, 399–403. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.-C.; Yu, G.; Corn, P.G.; Damasco, J.; Lee, Y.-C.; Song, J.H.; Navone, N.M.; Logothetis, C.J.; Melancon, M.P.; Panaretakis, T.; et al. Radium-223 Treatment Produces Prolonged Suppression of Resident Osteoblasts and Decreased Bone Mineral Density in Trabecular Bone in Osteoblast Reporter Mice. Cancers 2024, 16, 2603. https://doi.org/10.3390/cancers16142603
Lin S-C, Yu G, Corn PG, Damasco J, Lee Y-C, Song JH, Navone NM, Logothetis CJ, Melancon MP, Panaretakis T, et al. Radium-223 Treatment Produces Prolonged Suppression of Resident Osteoblasts and Decreased Bone Mineral Density in Trabecular Bone in Osteoblast Reporter Mice. Cancers. 2024; 16(14):2603. https://doi.org/10.3390/cancers16142603
Chicago/Turabian StyleLin, Song-Chang, Guoyu Yu, Paul G. Corn, Jossana Damasco, Yu-Chen Lee, Jian H. Song, Nora M. Navone, Christopher J. Logothetis, Marites P. Melancon, Theocharis Panaretakis, and et al. 2024. "Radium-223 Treatment Produces Prolonged Suppression of Resident Osteoblasts and Decreased Bone Mineral Density in Trabecular Bone in Osteoblast Reporter Mice" Cancers 16, no. 14: 2603. https://doi.org/10.3390/cancers16142603
APA StyleLin, S.-C., Yu, G., Corn, P. G., Damasco, J., Lee, Y.-C., Song, J. H., Navone, N. M., Logothetis, C. J., Melancon, M. P., Panaretakis, T., & Lin, S.-H. (2024). Radium-223 Treatment Produces Prolonged Suppression of Resident Osteoblasts and Decreased Bone Mineral Density in Trabecular Bone in Osteoblast Reporter Mice. Cancers, 16(14), 2603. https://doi.org/10.3390/cancers16142603