

Prognostic and Therapeutic Implications of Cell Division Cycle 20 Homolog in Breast Cancer

 , ,
, , 
Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. UALCAN; CDC20 Transcript and Protein Expressions in Tumors, BC Subtypes, and Normal Cells
2.2. KM Plotter; CDC20 Prognostic Value in BC and TNBC Patients
2.3. TIMER2.0; CDC20 Transcript Expression in Tumor and Normal Tissue
2.4. HPA Database; CDC20 Protein Expression and Associated Gene Clusters in BC and Normal Tissues
2.5. TNM-Plot; CDC20 Transcript Expression in BC and Normal Tissues
2.6. Bc-GenExMiner Database; CDC20 Transcript Expression among BC Subtypes
2.7. TIMER2.0; BC CDC20 Expression and Immune Infiltration Analysis
2.8. LinkedOmics, GO, KEGG, and PANTHER; CDC20 Attributes and Related Genes
2.9. STRING; CDC20 Related Gene Enrichment Analysis
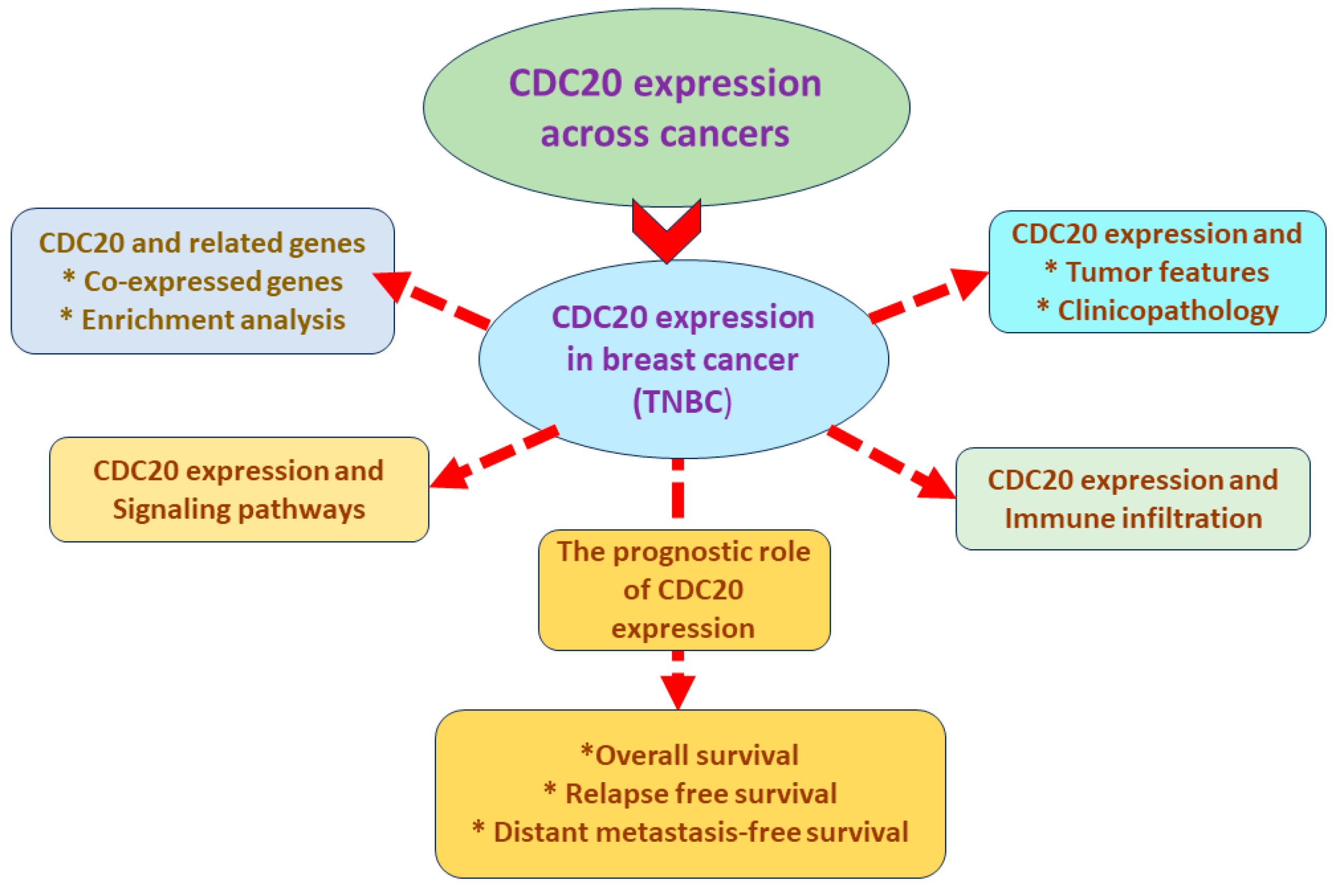
3. Results
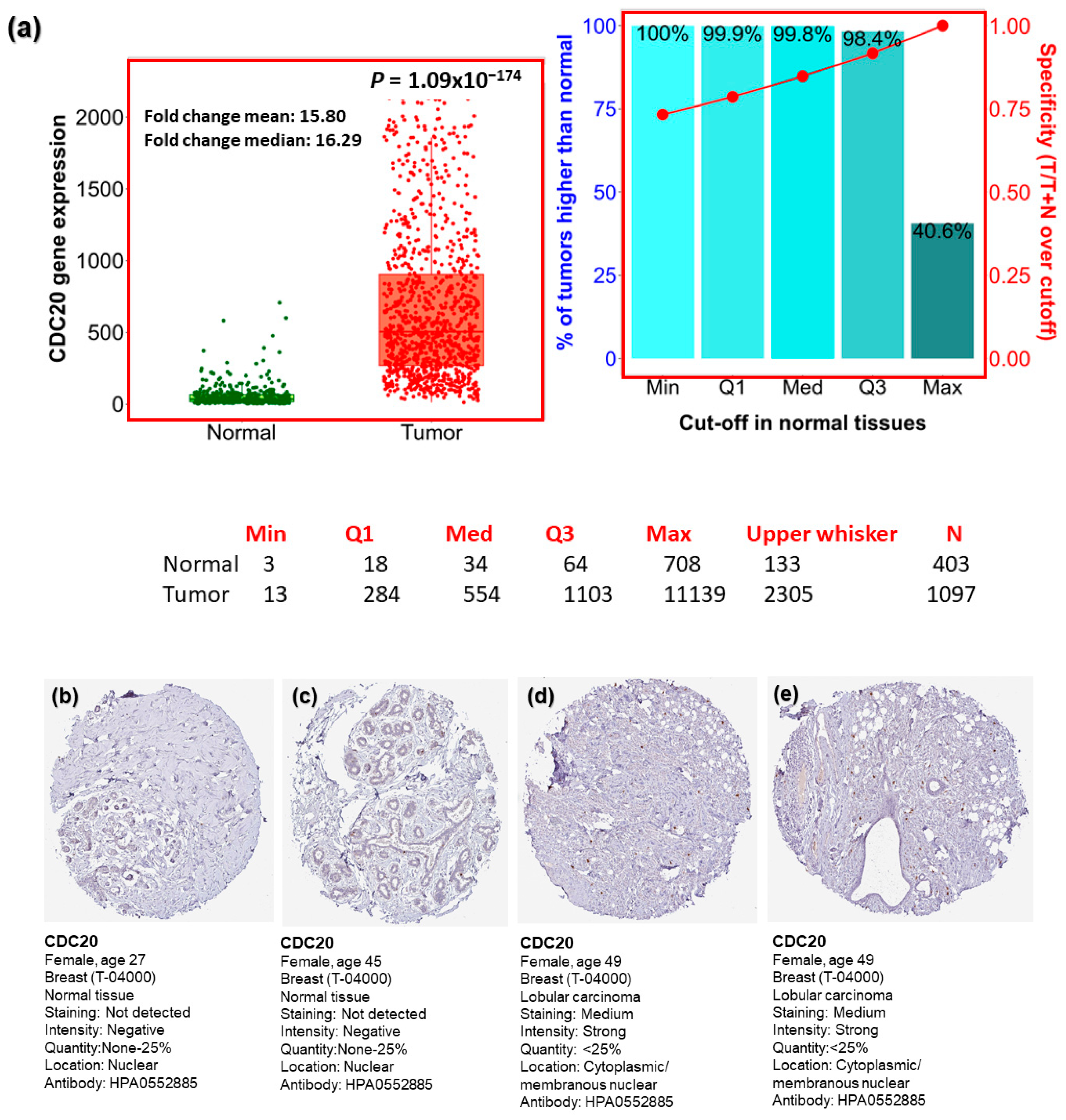
3.1. Analysis of CDC20 mRNA and Protein Expressions in BC Compared to Normal Tissues
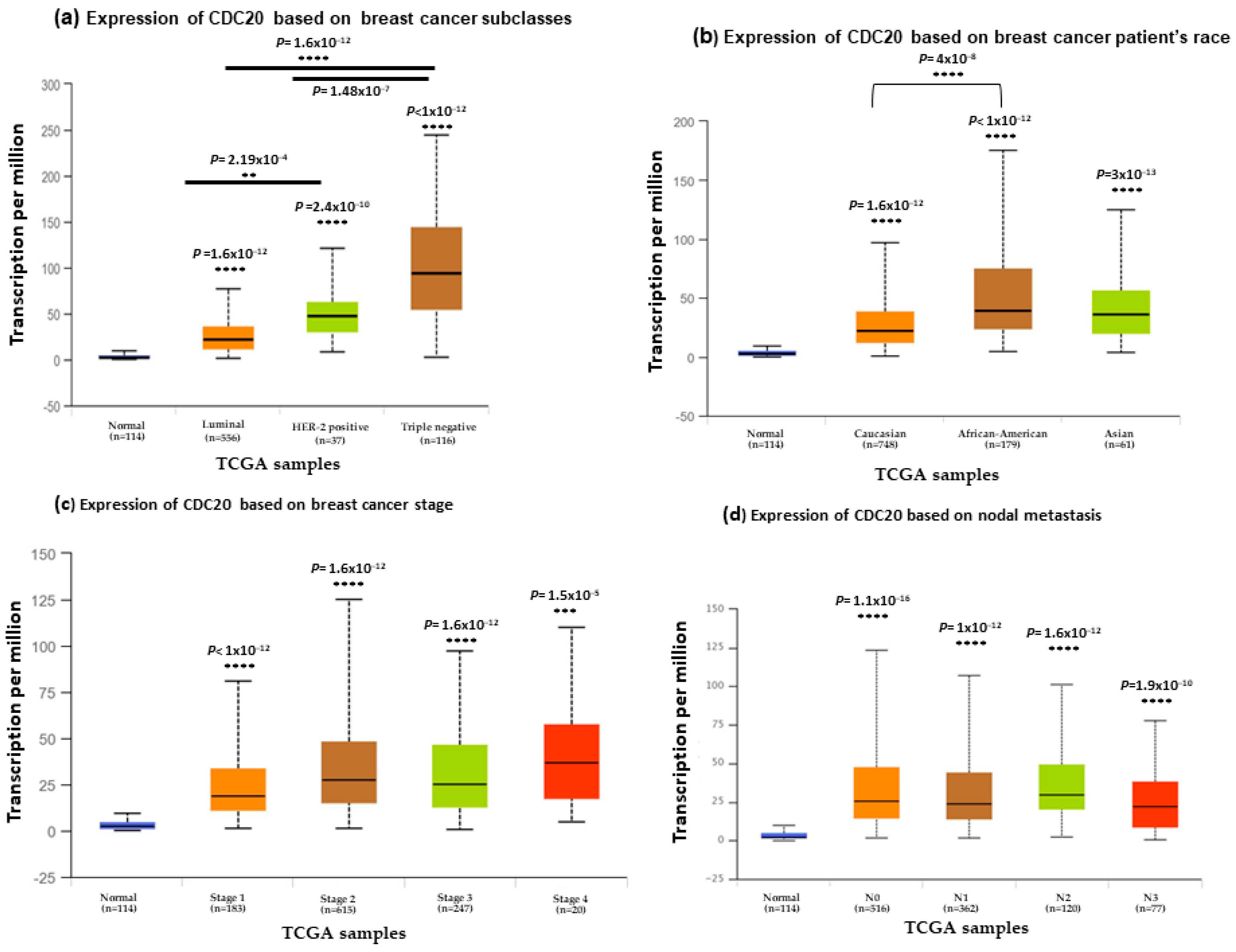
3.2. CDC20 Gene Expression Based on Hormone and BC1/2 Status
3.3. CDC20 mRNA Expression in Various Clinicopathological Features in BC
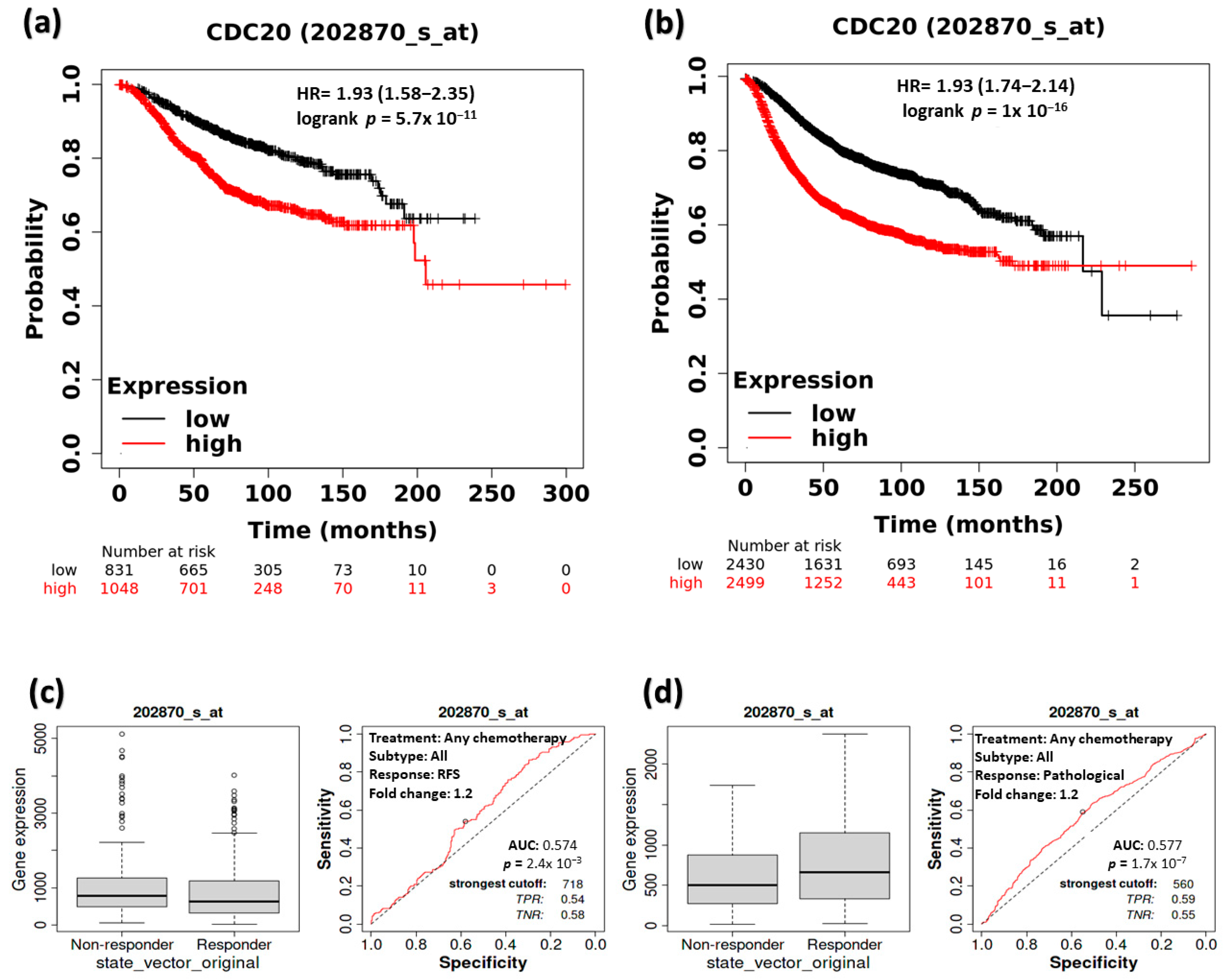
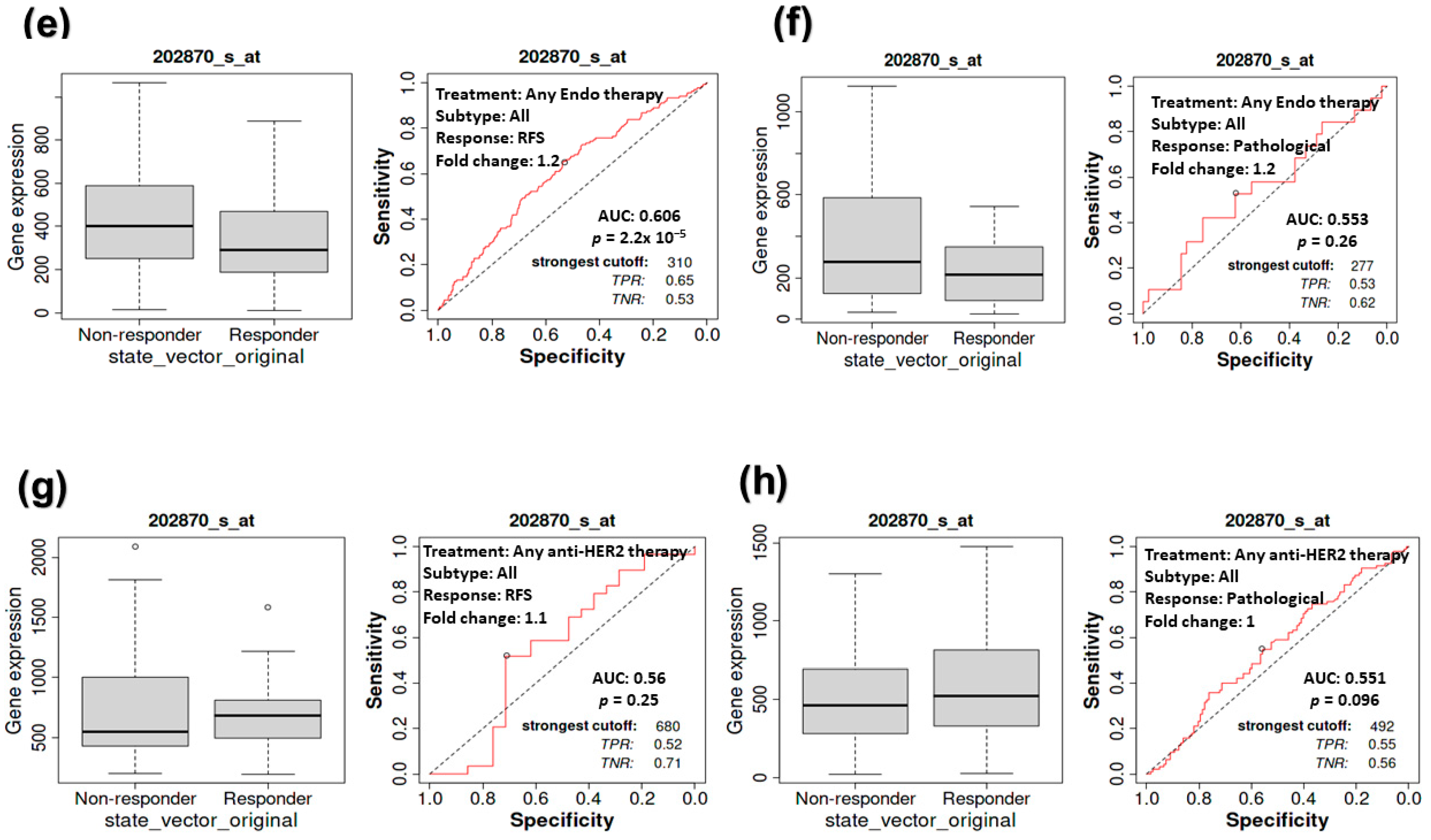
3.4. The Impact of CDC20 Expression on BC Patients’ RFS and Complete Pathological Response (CPR)
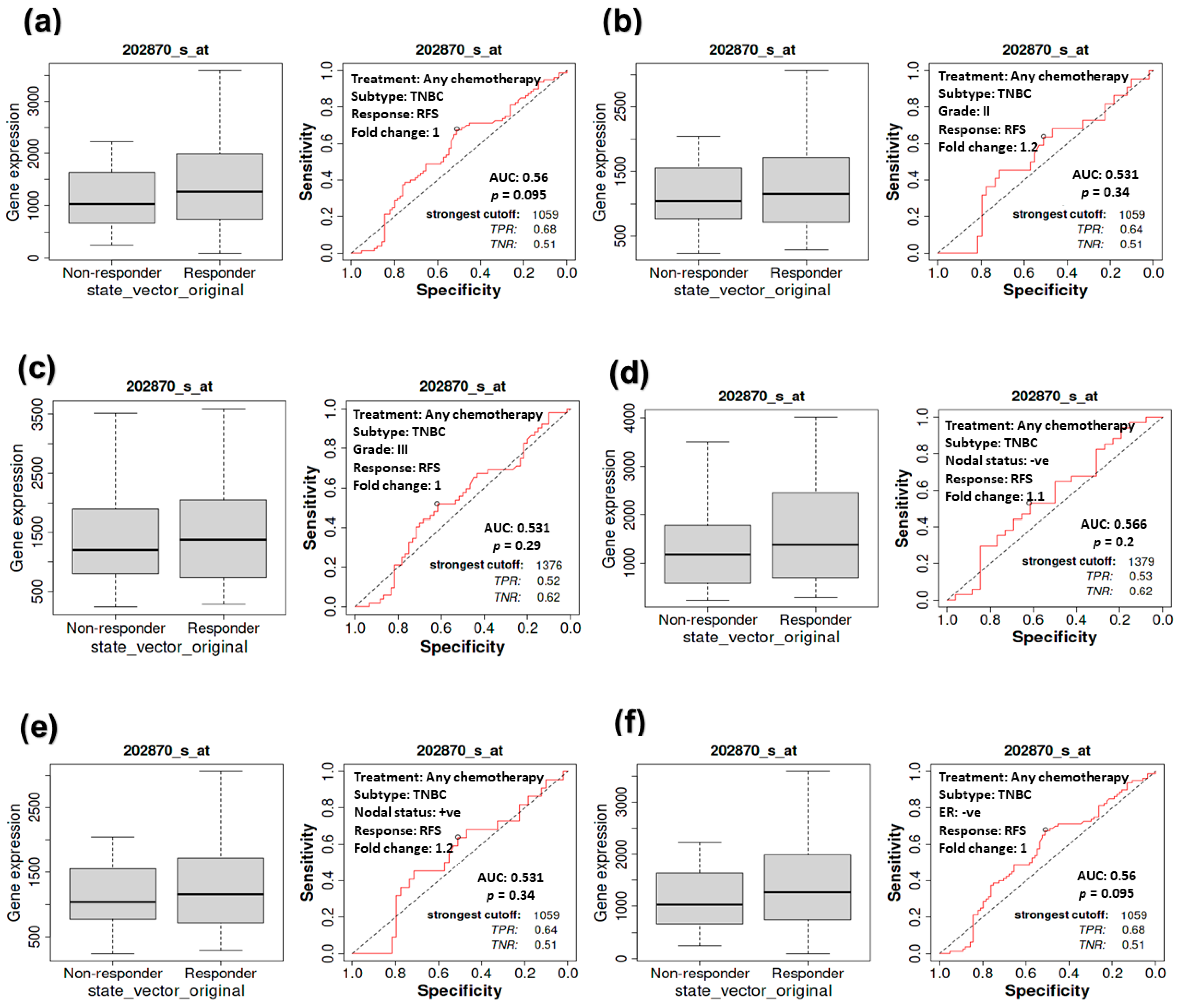
3.5. The Impact of CDC20 Expression on CPR in TNBC Subtype Patients
3.6. The Association between CDC20 Expression and RFS in TNBC Patients
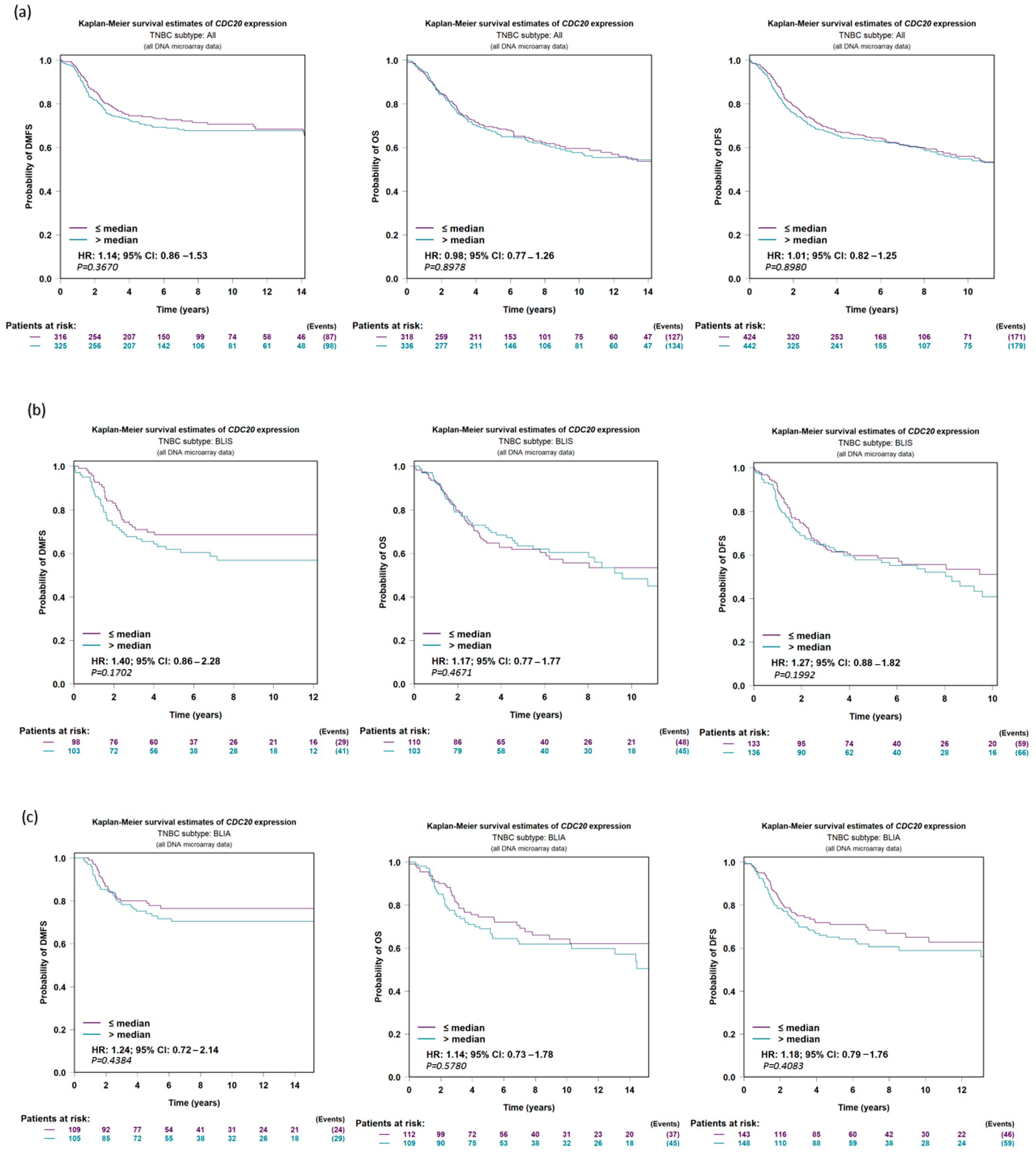
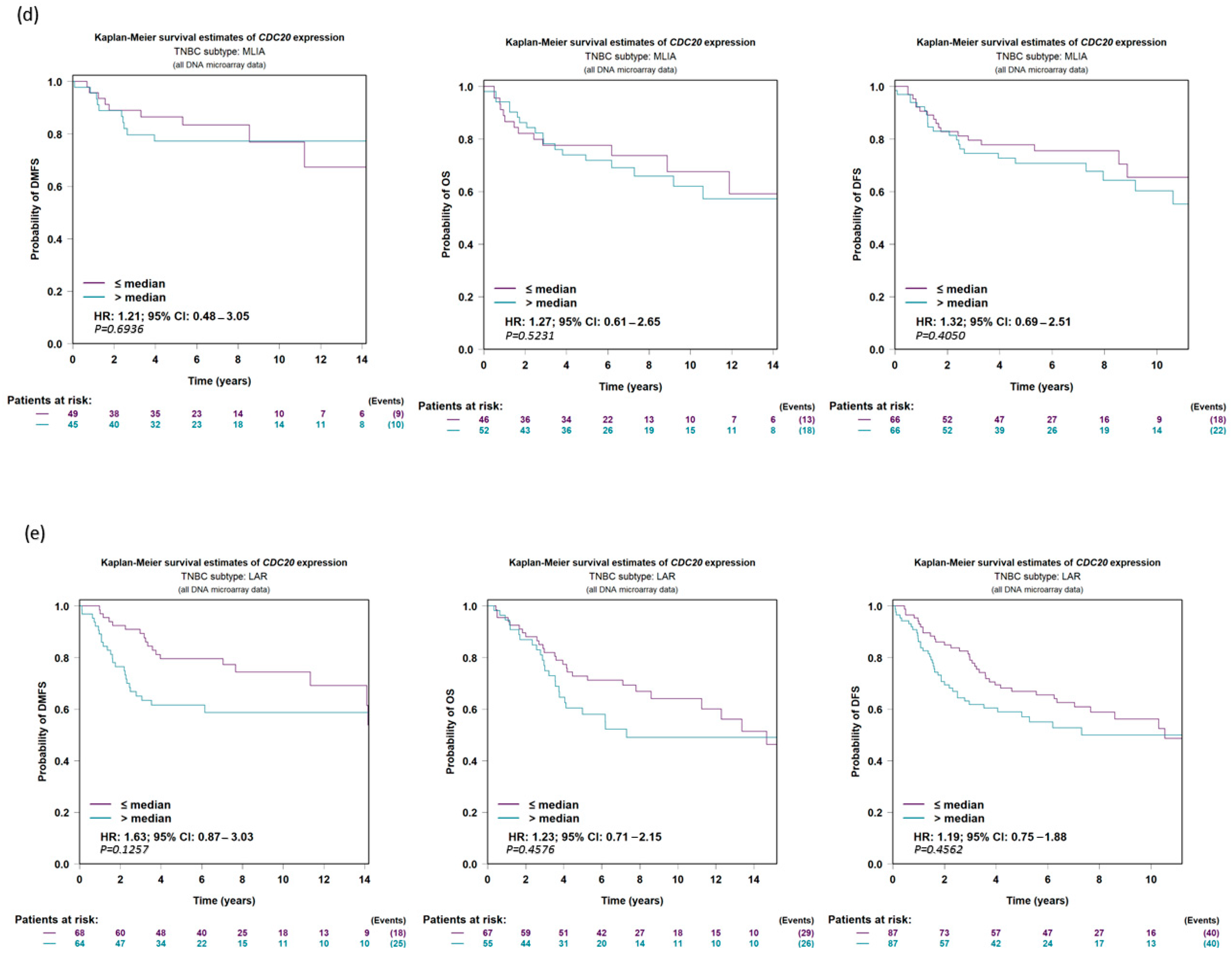
3.7. Expression of CDC20 in Relation to DFS, OS, and DMFS in TNBC Patients
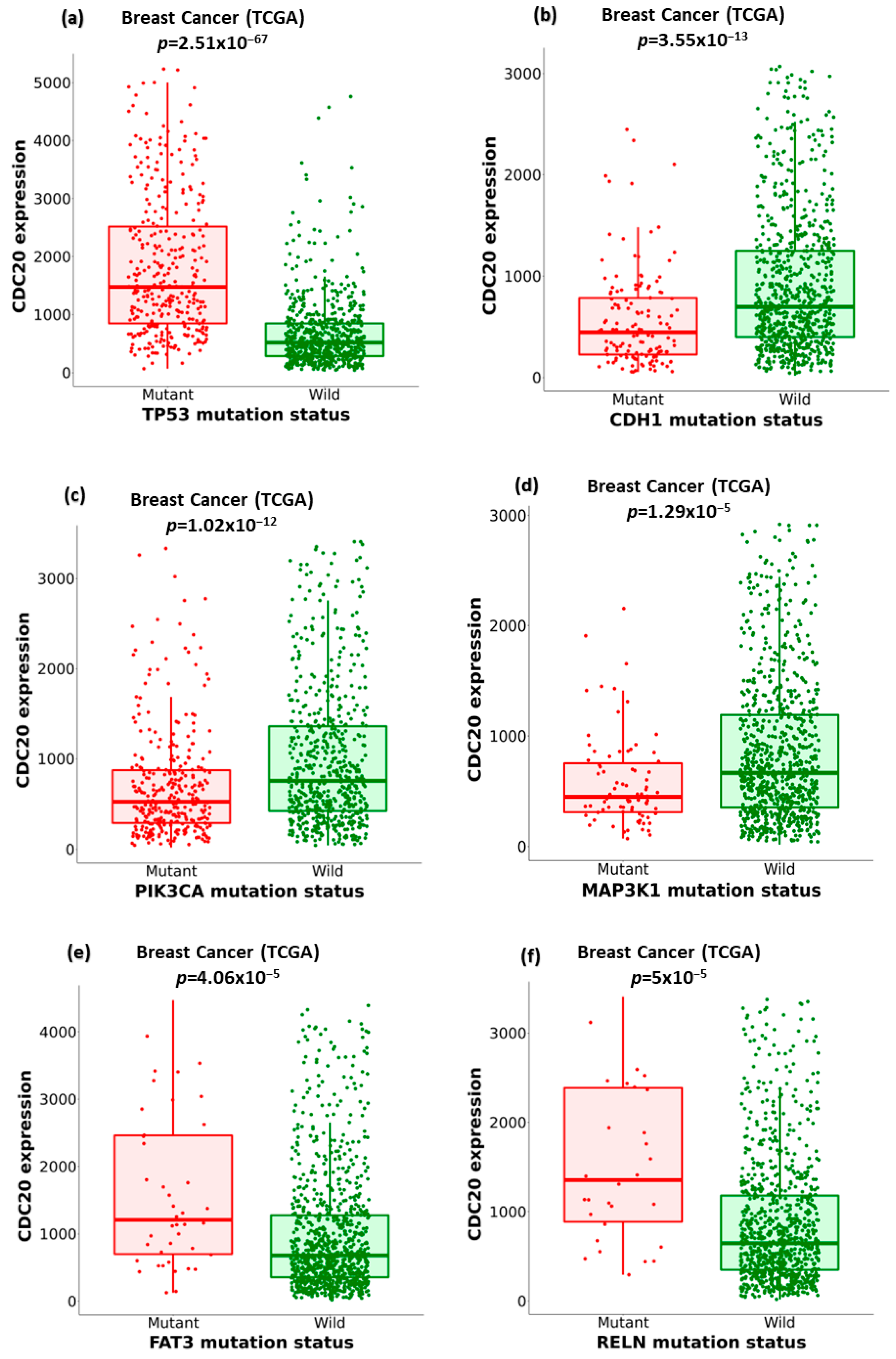
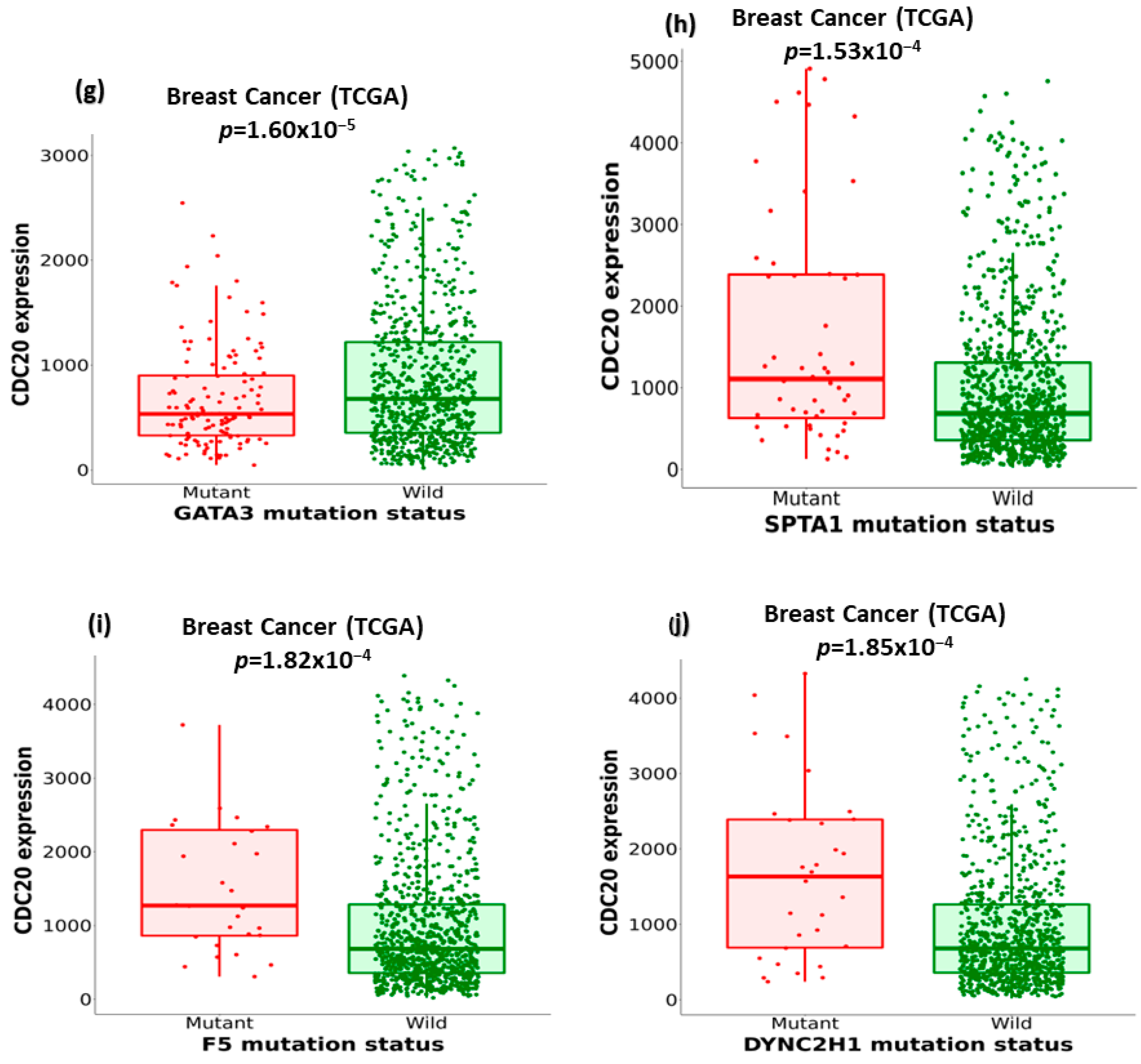
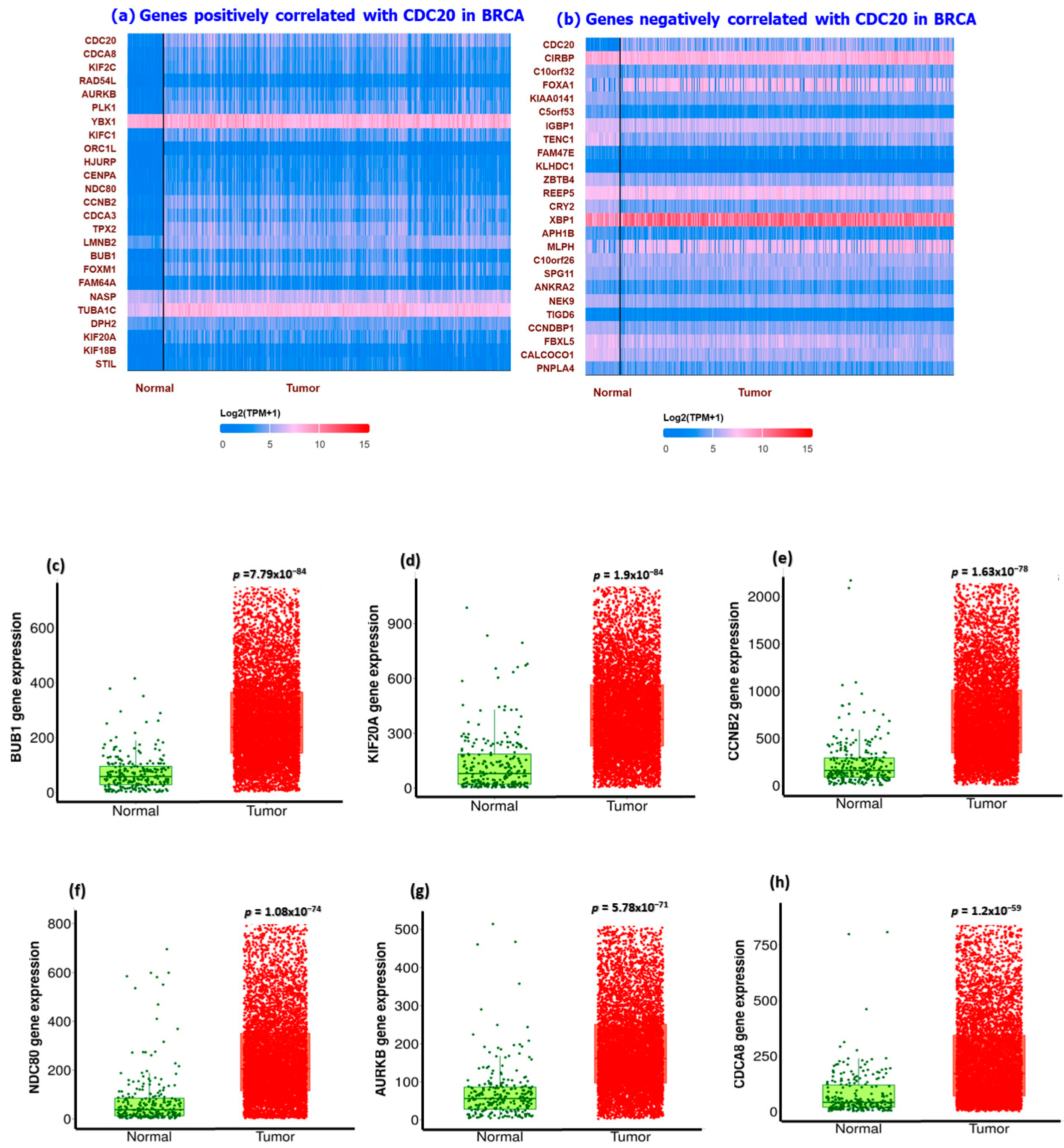
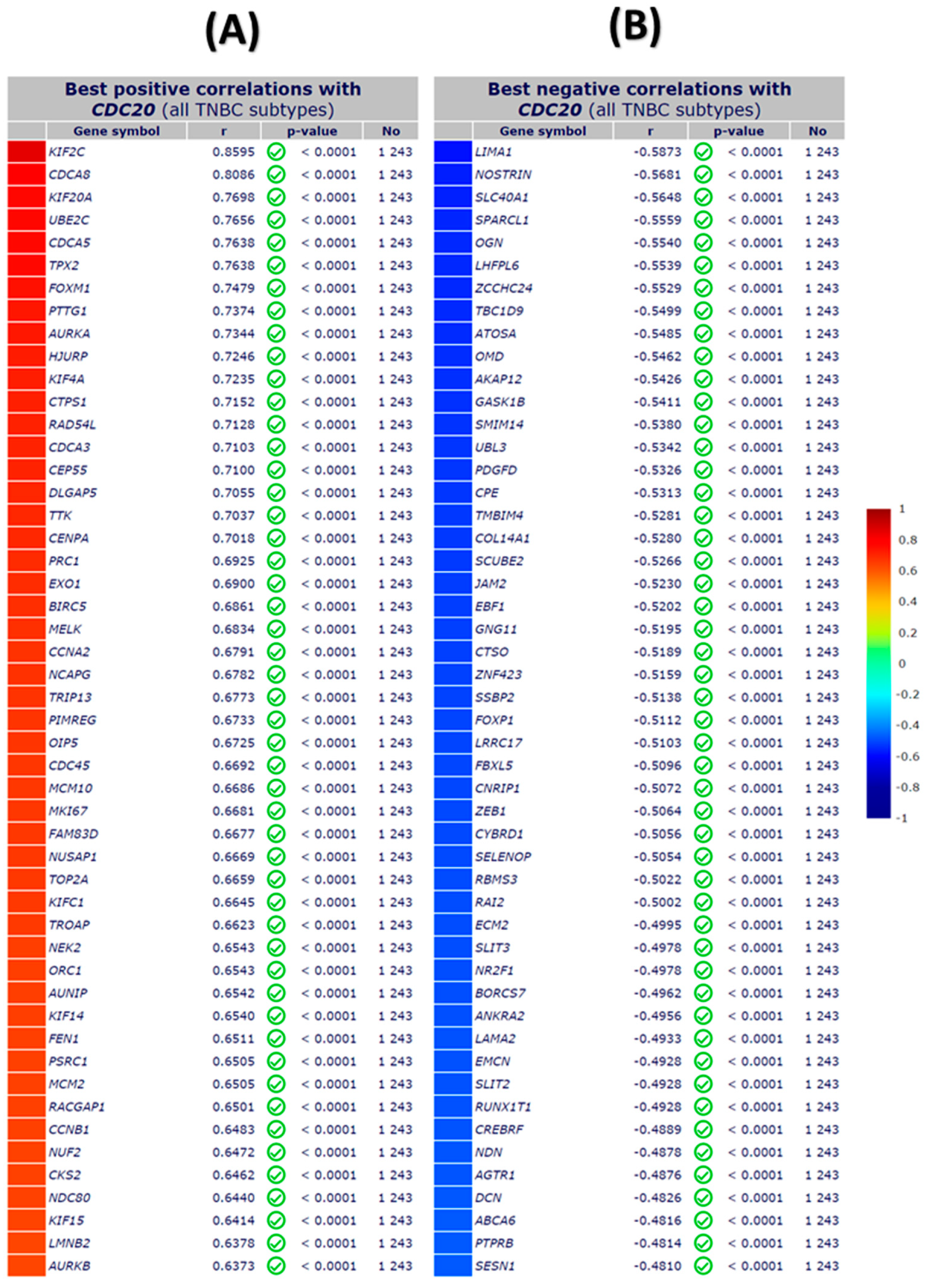
3.8. Association of Ten Commonly Mutated Genes with CDC20 mRNA Expression in BC
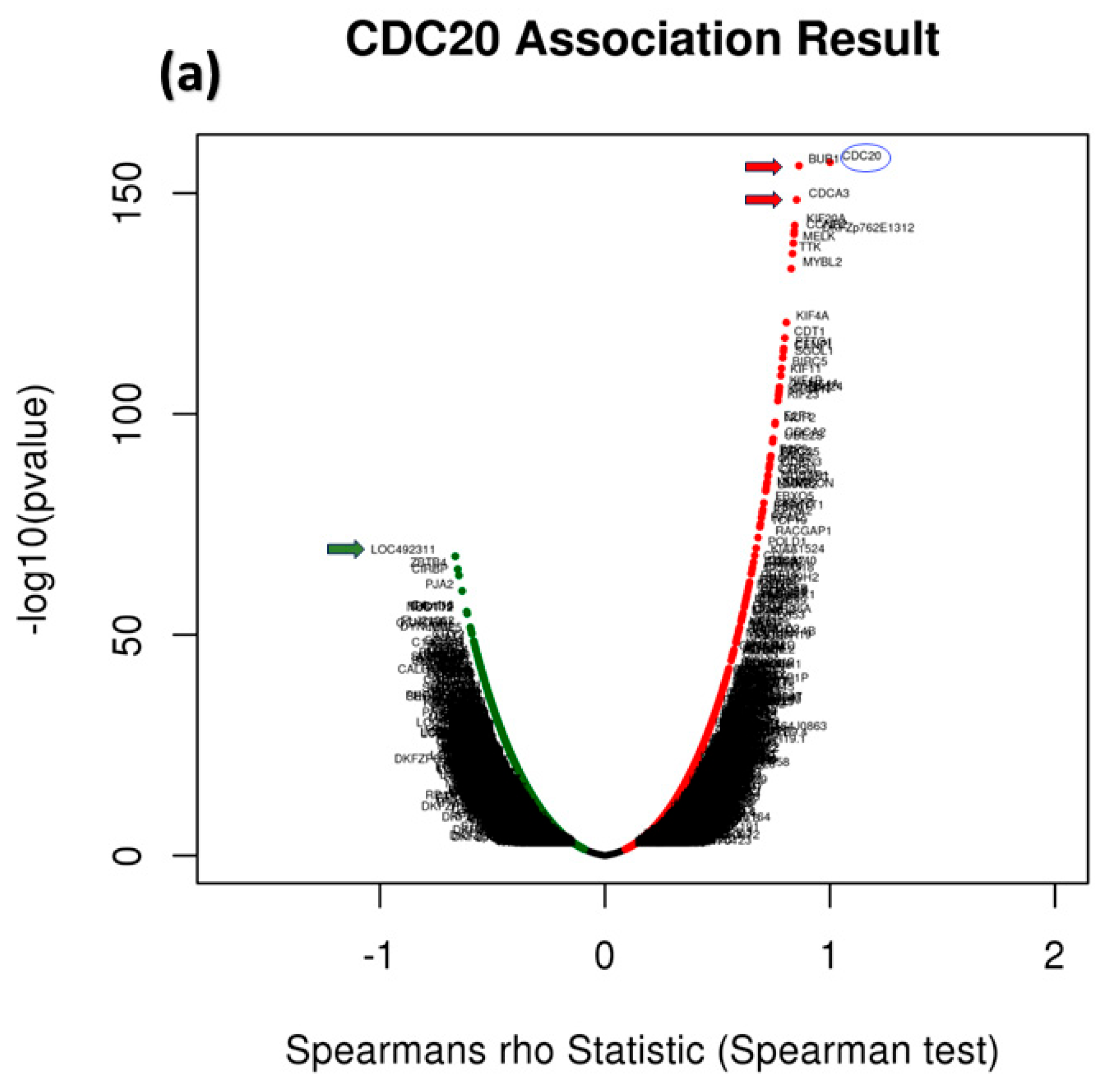
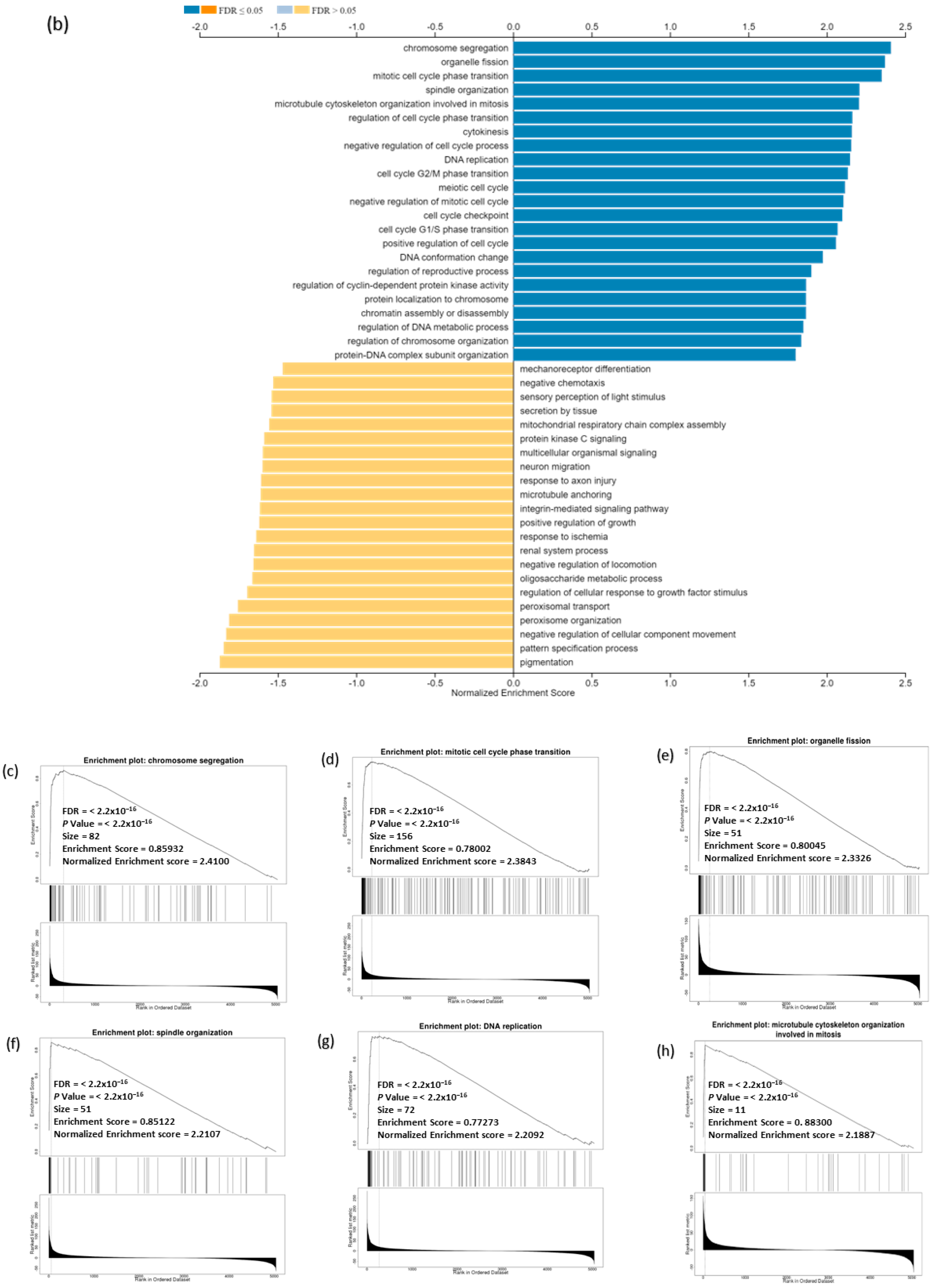
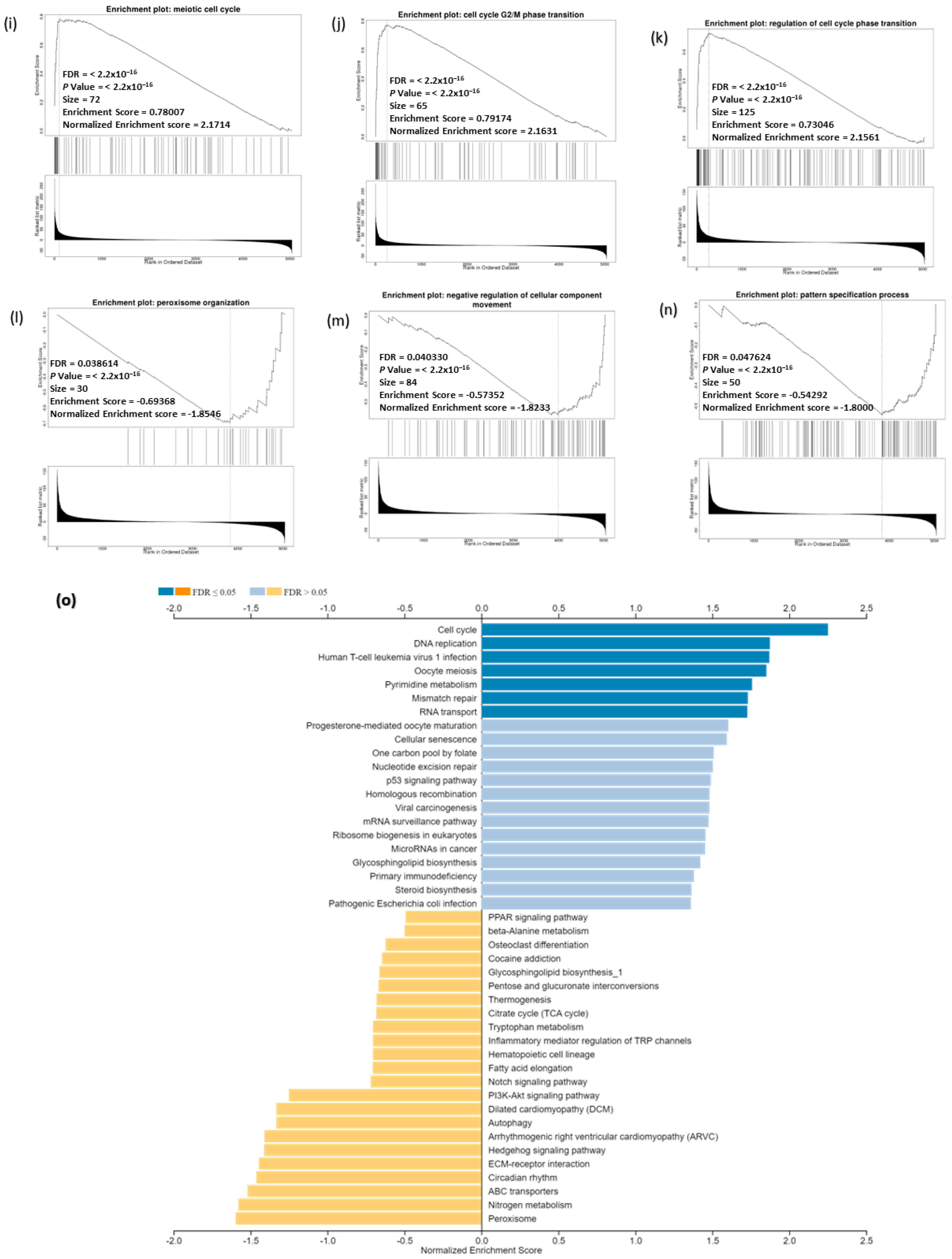
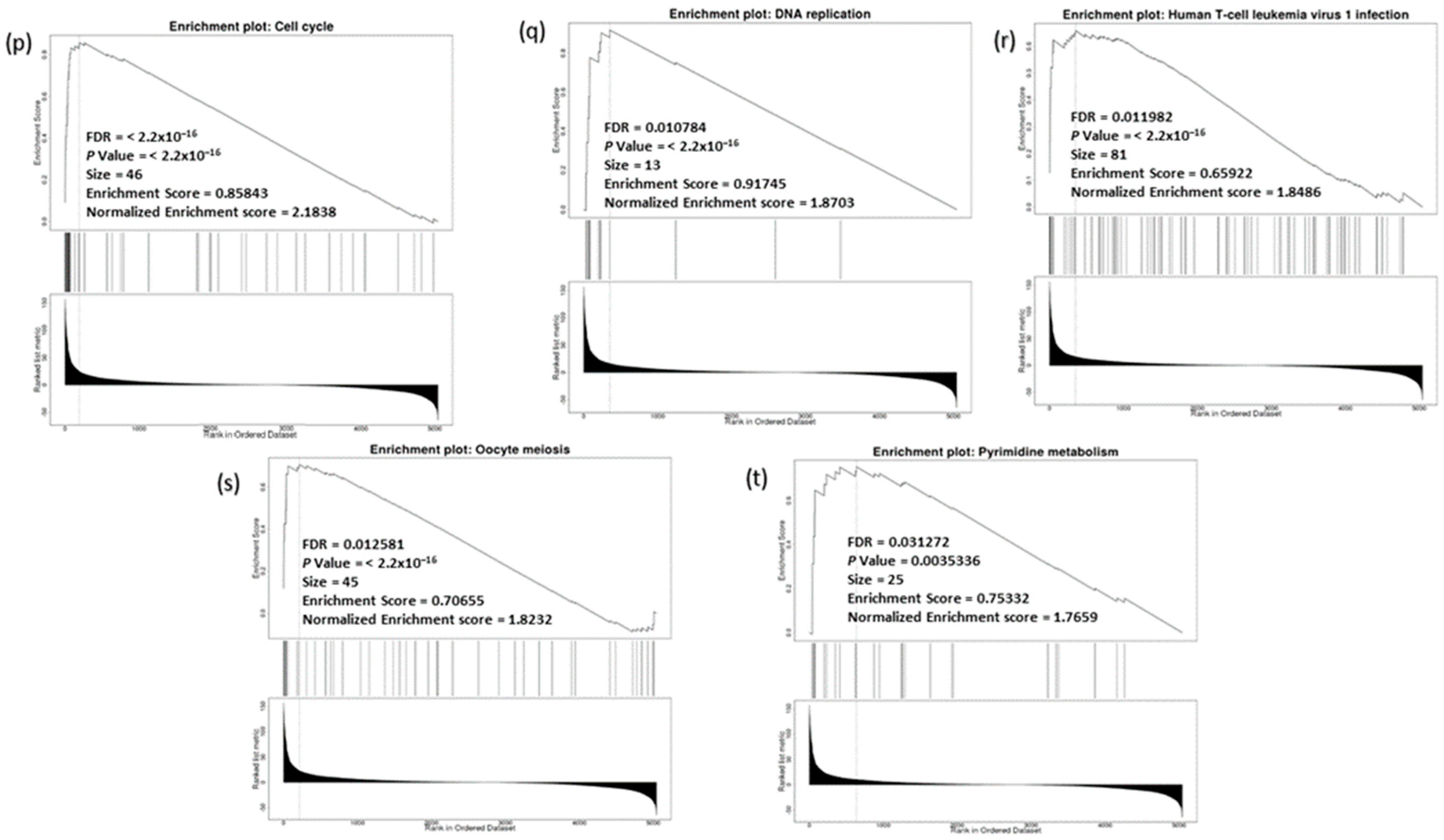
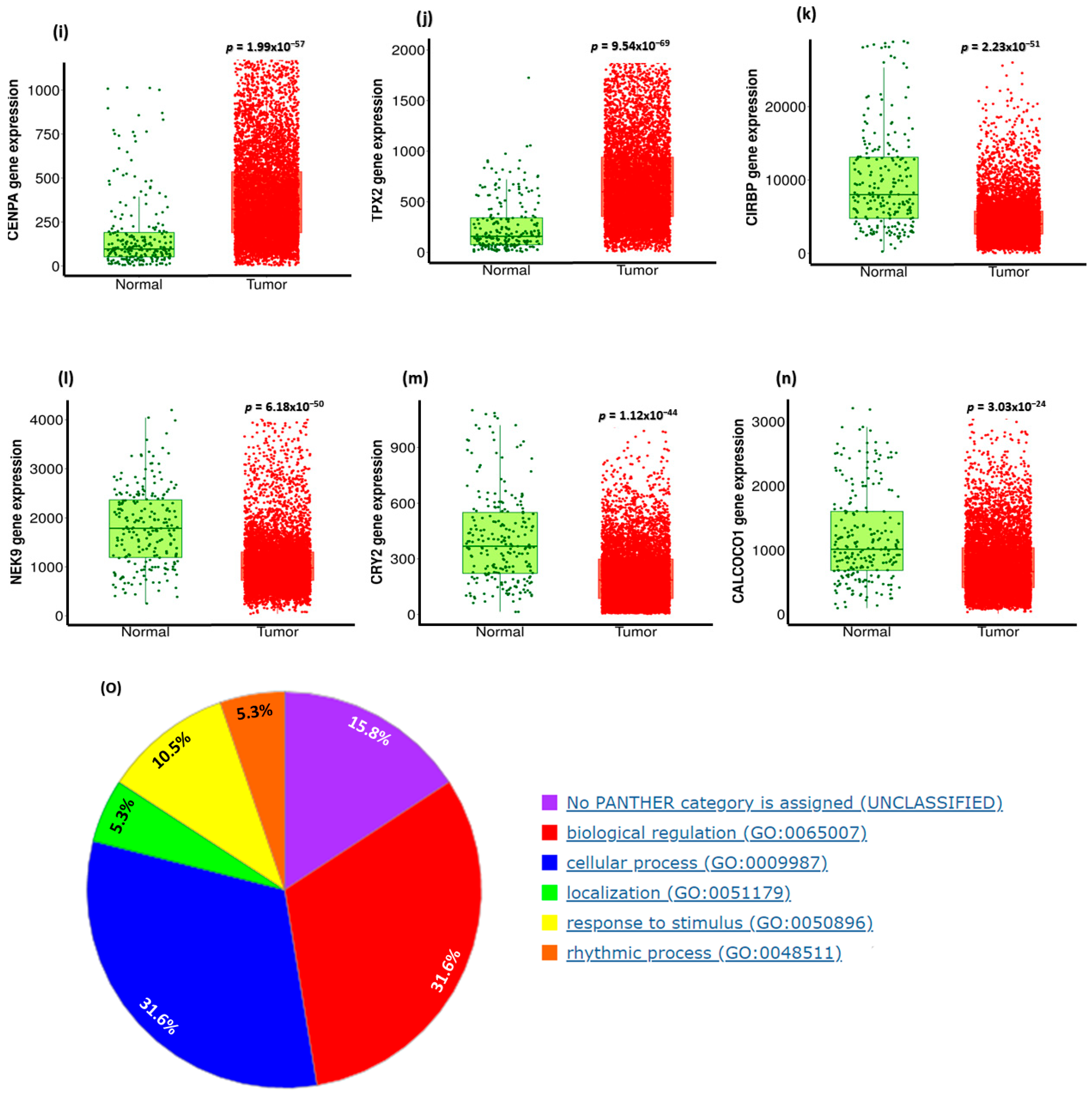
3.9. Analysis of GO and KEGG Biological Pathways of the Genes co-Expressed with CDC20 in BC
3.10. TNBC IHC Analysis
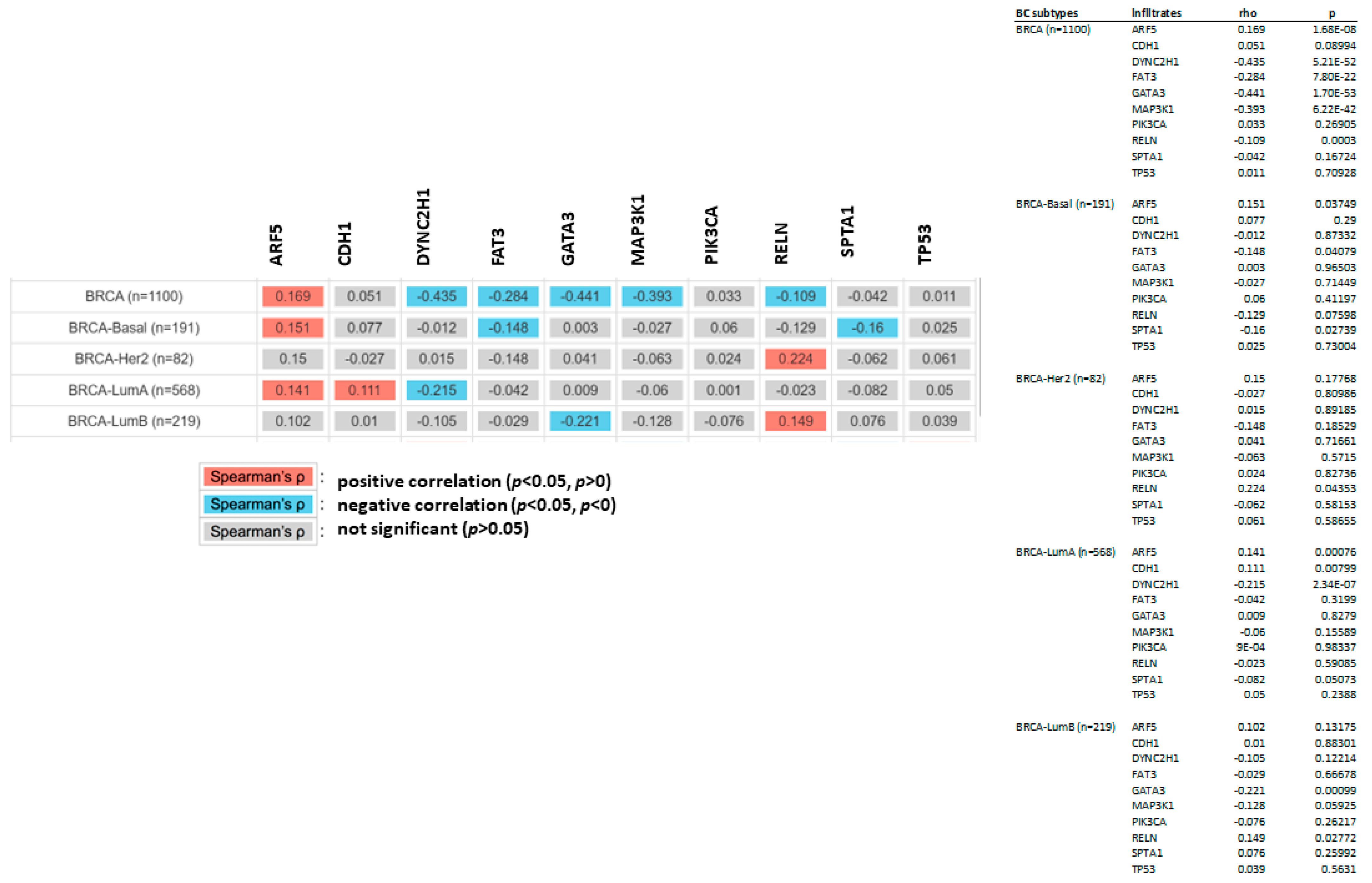
3.11. CDC20 Is Correlated with Tumor Purity and Immune Infiltration Levels in BC
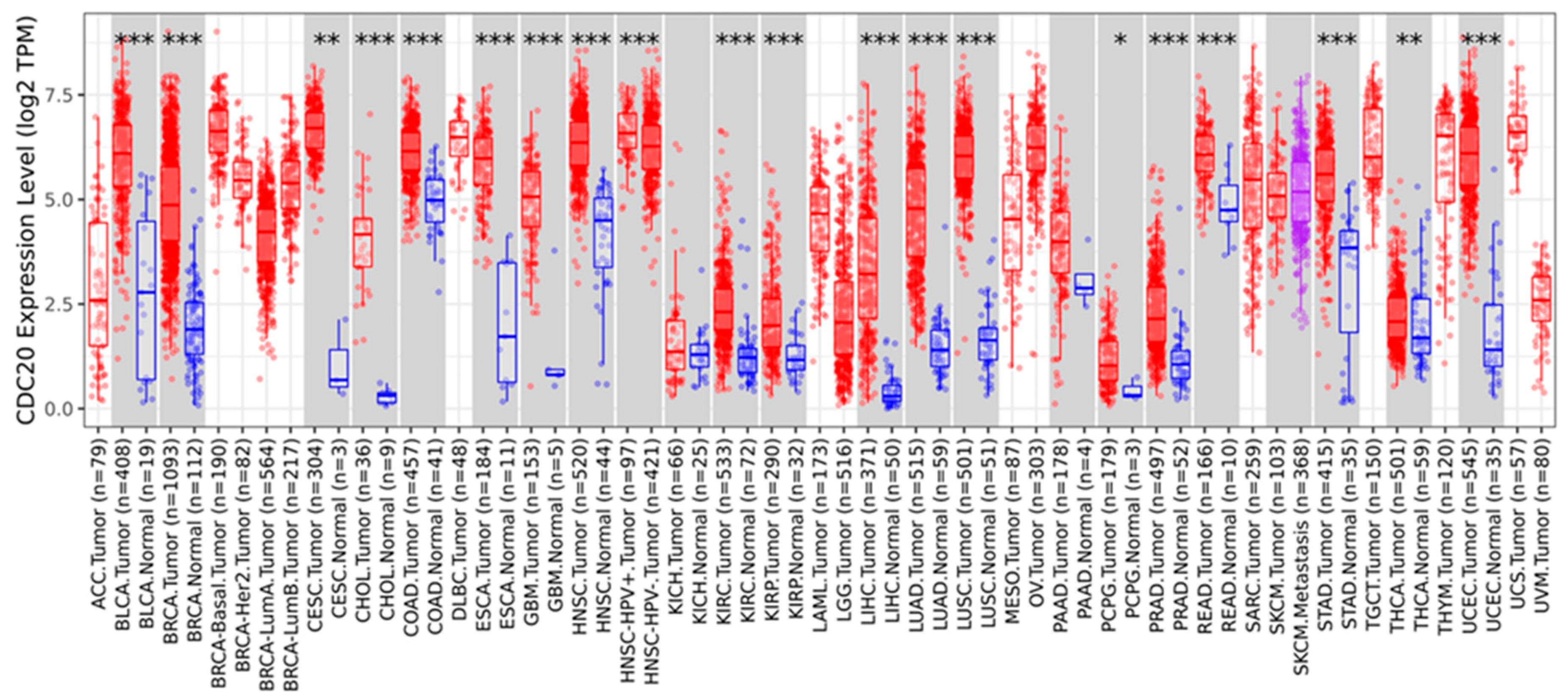
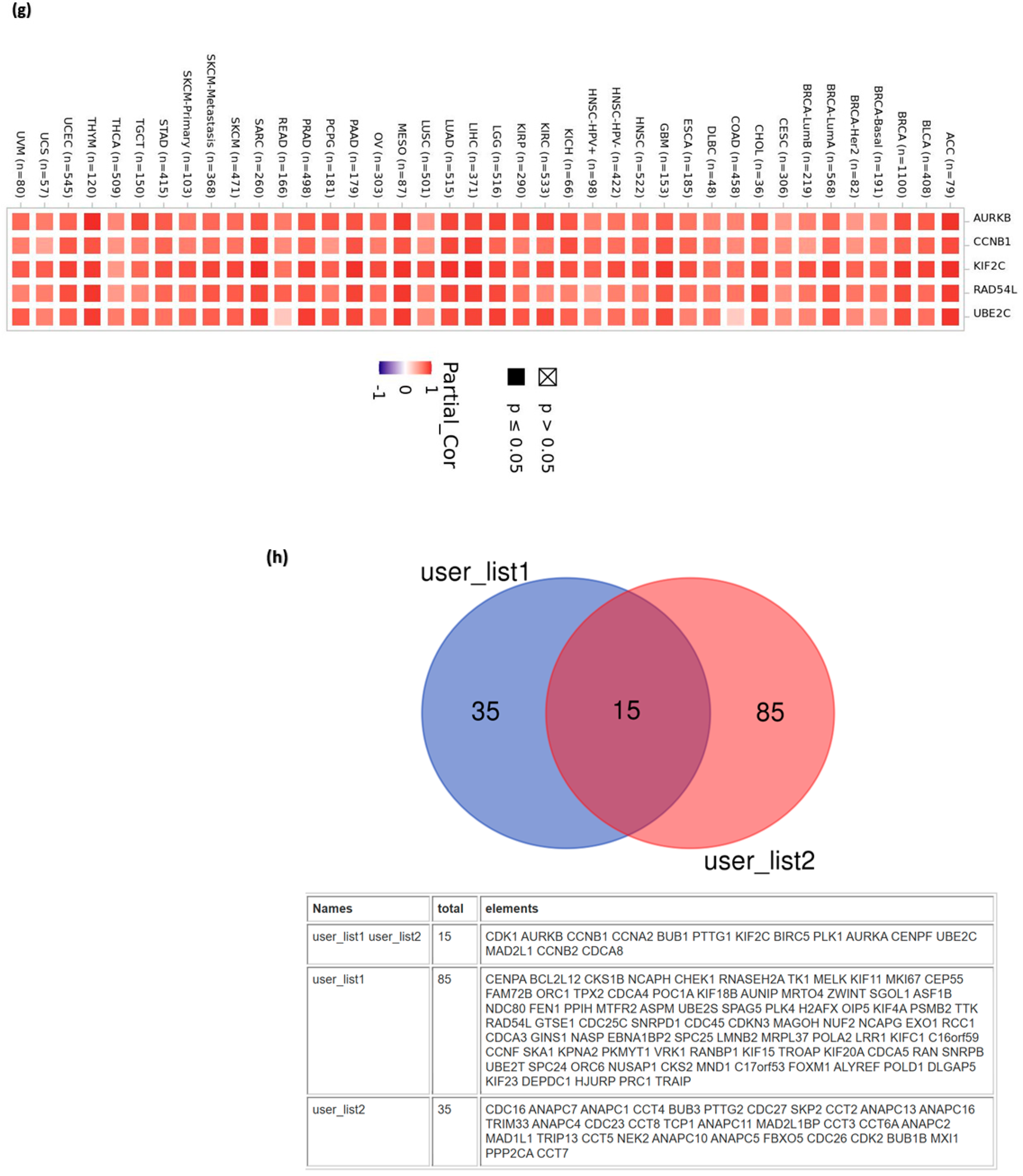
3.12. Pan-Cancer Analysis
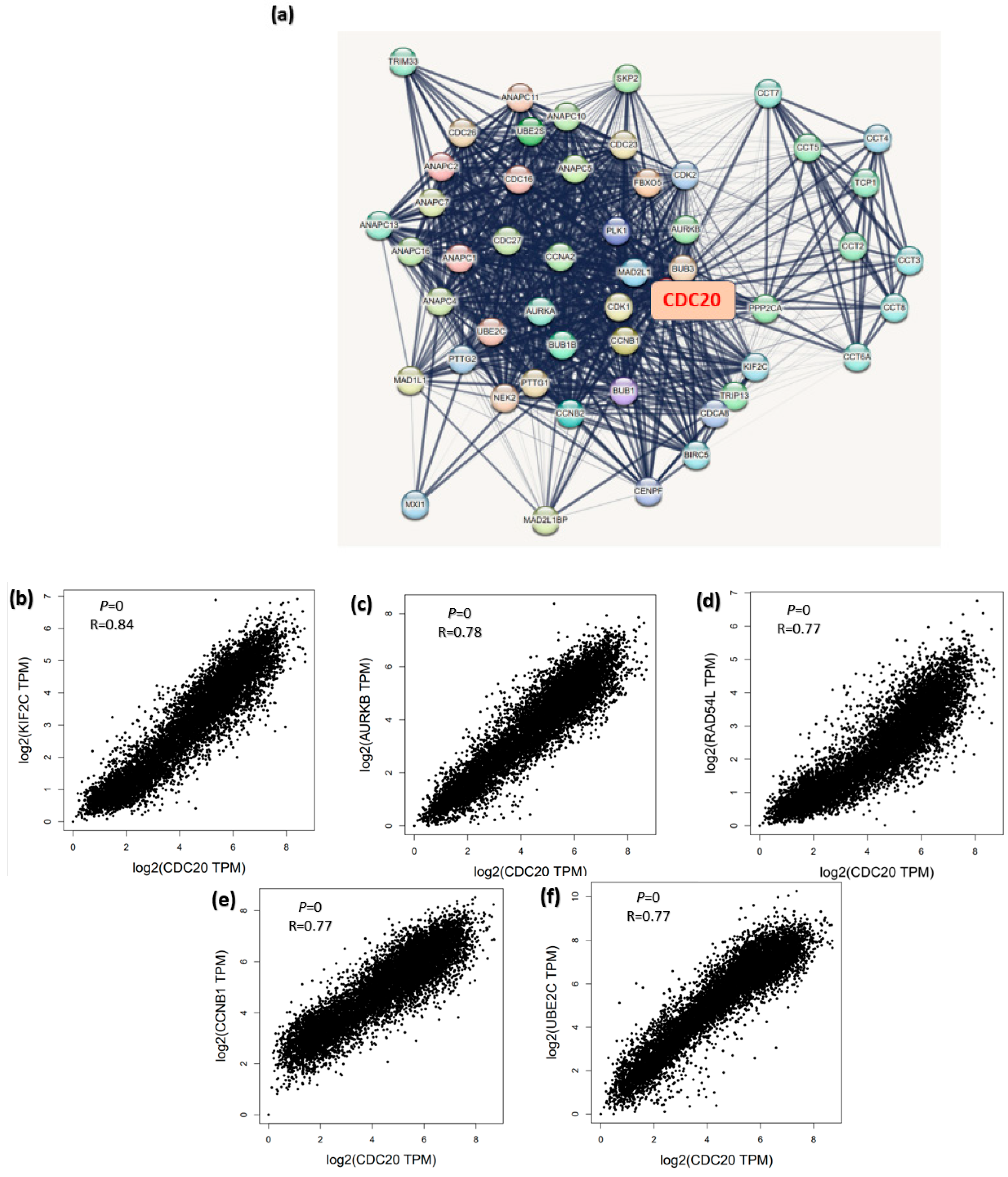
3.13. CDC20 Linked Gene Enrichment Analysis Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA A Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.A.; Oza, G.; Sharma, A.; Arriaga, L.G.; Hernández Hernández, J.M.; Rotello, V.M.; Ramirez, J.T. Triple-Negative Breast Cancer: A Review of Conventional and Advanced Therapeutic Strategies. Int. J. Environ. Res. Public Health 2020, 17, 2078. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.Y.S.; Tse, G.M. Molecular Classification of Breast Cancer. Adv. Anat. Pathol. 2020, 27, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Prat, A.; Pineda, E.; Adamo, B.; Galván, P.; Fernández, A.; Gaba, L.; Díez, M.; Viladot, M.; Arance, A.; Muñoz, M. Clinical implications of the intrinsic molecular subtypes of breast cancer. Breast 2015, 24 (Suppl. S2), S26–S35. [Google Scholar] [CrossRef] [PubMed]
- Karn, V.; Sandhya, S.; Hsu, W.; Parashar, D.; Singh, H.N.; Jha, N.K.; Gupta, S.; Dubey, N.K.; Kumar, S. CRISPR/Cas9 system in breast cancer therapy: Advancement, limitations and future scope. Cancer Cell Int. 2022, 22, 234. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.K.; Ko, T.-H.; Dubey, R.; Chouhan, M.; Tsai, L.-W.; Singh, H.N.; Chaubey, K.K.; Dayal, D.; Chiang, C.-W.; Kumar, S. CRISPR/Cas9 as a therapeutic tool for triple negative breast cancer: From bench to clinics. Front. Mol. Biosci. 2023, 10, 1214489. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-F.; Liao, Y.-Y.; Yang, M.; Peng, N.-F.; Xie, S.-R.; Xie, Y.-F. Discordances in ER, PR and HER2 receptors between primary and recurrent/metastatic lesions and their impact on survival in breast cancer patients. Med. Oncol. 2014, 31, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Aitken, S.J.; Thomas, J.S.; Langdon, S.P.; Harrison, D.J.; Faratian, D. Quantitative analysis of changes in ER, PR and HER2 expression in primary breast cancer and paired nodal metastases. Ann. Oncol. 2010, 21, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Lindström, L.S.; Karlsson, E.; Wilking, U.M.; Johansson, U.; Hartman, J.; Lidbrink, E.K.; Hatschek, T.; Skoog, L.; Bergh, J. Clinically used breast cancer markers such as estrogen receptor, progesterone receptor, and human epidermal growth factor receptor 2 are unstable throughout tumor progression. J. Clin. Oncol. 2012, 30, 2601–2608. [Google Scholar] [CrossRef] [PubMed]
- Jamdade, V.S.; Sethi, N.; Mundhe, N.A.; Kumar, P.; Lahkar, M.; Sinha, N. Therapeutic targets of triple-negative breast cancer: A review. Br. J. Pharmacol. 2015, 172, 4228–4237. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, X.; Zhou, W.H.; Liu, A.W.; Wu, J.B.; Deng, J.Y.; Yue, C.F.; Yang, S.B.; Wang, J.; Yuan, Z.Y.; et al. Aurora-A identifies early recurrence and poor prognosis and promises a potential therapeutic target in triple negative breast cancer. PLoS ONE 2013, 8, e56919. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Lowe, V.J.; Lee, S. Inhibition of Cdc20 suppresses the metastasis in triple negative breast cancer (TNBC). Breast Cancer 2021, 28, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Romecín, P.A.; Vinyoles, M.; López-Millán, B.; de la Guardia, R.D.; Atucha, N.M.; Querol, S.; Bueno, C.; Benitez, R.; Gonzalez-Rey, E.; Delgado, M. Robust in vitro and in vivo immunosuppressive and anti-inflammatory properties of inducible caspase-9-mediated apoptotic mesenchymal stromal/stem cell. Stem Cells Transl. Med. 2022, 11, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, M.; Mohamed, G.A.; Mohamed Saleem, M.A.; Ognjenovic, N.B.; Lu, H.; Kolling, F.W.; Wilkins, O.M.; Das, S.; LaCroix, I.S.; Nagaraj, S.H.; et al. Pharmacological induction of chromatin remodeling drives chemosensitization in triple-negative breast cancer. Cell Rep. Med. 2024, 5, 101504. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, F.; Huang, S.; Liu, L. PYCR3 Modulates mtDNA Copy Number to Drive Proliferation and Doxorubicin Resistance in Triple-Negative Breast Cancer. Int. J. Biochem. Cell Biol. 2024, 171, 106581. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Castro, A.C.; Lin, N.U.; Polyak, K. Insights into Molecular Classifications of Triple-Negative Breast Cancer: Improving Patient Selection for Treatment. Cancer Discov. 2019, 9, 176–198. [Google Scholar] [CrossRef] [PubMed]
- Lyman, G.H.; Greenlee, H.; Bohlke, K.; Bao, T.; DeMichele, A.M.; Deng, G.E.; Fouladbakhsh, J.M.; Gil, B.; Hershman, D.L.; Mansfield, S.; et al. Integrative Therapies During and After Breast Cancer Treatment: ASCO Endorsement of the SIO Clinical Practice Guideline. J. Clin. Oncol. 2018, 36, 2647–2655. [Google Scholar] [CrossRef]
- Wilson, E.A.; Sultana, N.; Shah, K.N.; Elford, H.L.; Faridi, J.S. Molecular Targeting of RRM2, NF-κB, and Mutant TP53 for the Treatment of Triple-Negative Breast Cancer. Mol. Cancer Ther. 2021, 20, 655–664. [Google Scholar] [CrossRef] [PubMed]
- García-Aranda, M.; Redondo, M. Immunotherapy: A challenge of breast cancer treatment. Cancers 2019, 11, 1822. [Google Scholar] [CrossRef] [PubMed]
- Mehanna, J.; Haddad, F.G.; Eid, R.; Lambertini, M.; Kourie, H.R. Triple-negative breast cancer: Current perspective on the evolving therapeutic landscape. Int. J. Women’s Health 2019, 11, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Nakhjavani, M.; Hardingham, J.E.; Palethorpe, H.M.; Price, T.J.; Townsend, A.R. Druggable molecular targets for the treatment of triple negative breast cancer. J. Breast Cancer 2019, 22, 341. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.J.; Walsh, S.; McDermott, E.W.; Crown, J. Biomarkers in breast cancer: Where are we and where are we going? Adv. Clin. Chem. 2015, 71, 1–23. [Google Scholar] [PubMed]
- Zang, Z.; Yin, Y.; Liu, C.; Zhu, Q.; Huang, X.; Li, H.; Yang, R. IL21R hypomethylation as a biomarker for distinguishing benign and malignant breast tumours. Epigenetics 2024, 19, 2352683. [Google Scholar] [CrossRef] [PubMed]
- Adinew, G.M.; Messeha, S.; Taka, E.; Soliman, K.F.A. The Prognostic and Therapeutic Implications of the Chemoresistance Gene BIRC5 in Triple-Negative Breast Cancer. Cancers 2022, 14, 5180. [Google Scholar] [CrossRef] [PubMed]
- Nath, P.; Alfarsi, L.H.; El-Ansari, R.; Masisi, B.K.; Erkan, B.; Fakroun, A.; Ellis, I.O.; Rakha, E.A.; Green, A.R. The amino acid transporter SLC7A11 expression in breast cancer. Cancer Biol. Ther. 2024, 25, 2291855. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Shi, J.; Zhang, J.; Lin, X.; Jiang, W.; Zhu, Y.; Song, Y.; Lu, Y.; Ma, Y. PKM2 is a potential prognostic biomarker and related to immune infiltration in lung cancer. Sci. Rep. 2023, 13, 22243. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Huang, R.; Wang, H.; Peng, Y.; Fan, Y.; Feng, Z.; Zeng, Z.; Ji, Y.; Wang, Y.; Lu, J. Prognostic and Immunological Role of PPP1R14A as a Pan-Cancer Analysis Candidate. Front. Genet 2022, 13, 842975. [Google Scholar] [CrossRef] [PubMed]
- Geng, R.; Zheng, Y.; Zhou, D.; Li, Q.; Li, R.; Guo, X. ZBTB7A, a potential biomarker for prognosis and immune infiltrates, inhibits progression of endometrial cancer based on bioinformatics analysis and experiments. Cancer Cell Int. 2020, 20, 542. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, J.; Jacobsen, F.W.; Hsu-Chen, J.; Wu, T.; Baum, L.G. A novel mammalian protein, p55CDC, present in dividing cells is associated with protein kinase activity and has homology to the Saccharomyces cerevisiae cell division cycle proteins Cdc20 and Cdc4. Mol. Cell Biol. 1994, 14, 3350–3363. [Google Scholar] [CrossRef]
- Weinstein, J. Cell cycle-regulated expression, phosphorylation, and degradation of p55Cdc. A mammalian homolog of CDC20/Fizzy/slp1. J. Biol. Chem. 1997, 272, 28501–28511. [Google Scholar] [CrossRef] [PubMed]
- Prinz, S.; Hwang, E.S.; Visintin, R.; Amon, A. The regulation of Cdc20 proteolysis reveals a role for APC components Cdc23 and Cdc27 during S phase and early mitosis. Curr. Biol. 1998, 8, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.M.; Bui, Q.T.; Kwak, M.; Lee, J.Y.; Lee, P.C. Targeting Cdc20 for cancer therapy. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188824. [Google Scholar] [CrossRef] [PubMed]
- Mondal, G.; Sengupta, S.; Panda, C.K.; Gollin, S.M.; Saunders, W.S.; Roychoudhury, S. Overexpression of Cdc20 leads to impairment of the spindle assembly checkpoint and aneuploidization in oral cancer. Carcinogenesis 2007, 28, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wan, L.; Zhong, J.; Inuzuka, H.; Liu, P.; Sarkar, F.H.; Wei, W. Cdc20: A potential novel therapeutic target for cancer treatment. Curr. Pharm Des. 2013, 19, 3210–3214. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, J.; Wan, L.; Zhou, X.; Wang, Z.; Wei, W. Targeting Cdc20 as a novel cancer therapeutic strategy. Pharmacol. Ther. 2015, 151, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; He, M.; Shah, A.A.; Wan, Y. Insights into APC/C: From cellular function to diseases and therapeutics. Cell Div. 2016, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Shang, G.; Ma, X.; Lv, G. Cell division cycle 20 promotes cell proliferation and invasion and inhibits apoptosis in osteosarcoma cells. Cell Cycle 2018, 17, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.J.; Hu, K.S.; Wang, D.S.; Zeng, Z.L.; Zhang, D.S.; Chen, D.L.; Bai, L.; Xu, R.H. CDC20 overexpression predicts a poor prognosis for patients with colorectal cancer. J. Transl. Med. 2013, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.Z.; Ma, Y.; Ji, B.; Liu, Y.; Hwu, P.; Abbruzzese, J.L.; Logsdon, C.; Wang, H. Increased CDC20 expression is associated with pancreatic ductal adenocarcinoma differentiation and progression. J. Hematol. Oncol. 2012, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Zhang, C.; Liu, X. Role of glucose metabolism and ATP in maintaining PINK1 levels during Parkin-mediated mitochondrial damage responses. J. Biol. Chem. 2015, 290, 904–917. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Meng, J. CDC20: A novel therapeutic target in cancer. Am. J. Transl. Res. 2023, 15, 678–693. [Google Scholar] [PubMed]
- Karra, H.; Repo, H.; Ahonen, I.; Löyttyniemi, E.; Pitkänen, R.; Lintunen, M.; Kuopio, T.; Söderström, M.; Kronqvist, P. Cdc20 and securin overexpression predict short-term breast cancer survival. Br. J. Cancer 2014, 110, 2905–2913. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Goel, M.K.; Khanna, P.; Kishore, J. Understanding survival analysis: Kaplan-Meier estimate. Int. J. Ayurveda Res. 2010, 1, 274. [Google Scholar] [PubMed]
- Györffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1,809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Barakat, A.; Mittal, A.; Ricketts, D.; Rogers, B.A. Understanding survival analysis: Actuarial life tables and the Kaplan–Meier plot. Br. J. Hosp. Med. 2019, 80, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Fekete, J.T.; Győrffy, B. ROCplot.org: Validating predictive biomarkers of chemotherapy/hormonal therapy/anti-HER2 therapy using transcriptomic data of 3,104 breast cancer patients. Int. J. Cancer 2019, 145, 3140–3151. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416.e411. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids. Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef] [PubMed]
- Bartha, Á.; Győrffy, B. TNMplot. com: A web tool for the comparison of gene expression in normal, tumor and metastatic tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef] [PubMed]
- Jézéquel, P.; Campone, M.; Gouraud, W.; Guérin-Charbonnel, C.; Leux, C.; Ricolleau, G.; Campion, L. bc-GenExMiner: An easy-to-use online platform for gene prognostic analyses in breast cancer. Breast Cancer Res. Treat 2012, 131, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Jézéquel, P.; Frénel, J.S.; Campion, L.; Guérin-Charbonnel, C.; Gouraud, W.; Ricolleau, G.; Campone, M. bc-GenExMiner 3.0: New mining module computes breast cancer gene expression correlation analyses. Database 2013, 2013, bas060. [Google Scholar] [CrossRef] [PubMed]
- Jézéquel, P.; Gouraud, W.; Ben Azzouz, F.; Guérin-Charbonnel, C.; Juin, P.P.; Lasla, H.; Campone, M. bc-GenExMiner 4.5: New mining module computes breast cancer differential gene expression analyses. Database 2021, 2021, baab007. [Google Scholar] [CrossRef] [PubMed]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res 2018, 46, D956–D963. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids. Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, N.; Zhu, L.; Zhen, D.; Li, M.; Chen, H.; Ye, M.; Wei, Y.; Shao, G. Multiple roles of apolipoprotein B mRNA editing enzyme catalytic subunit 3B (APOBEC3B) in human tumors: A pan-cancer analysis. BMC Bioinform. 2022, 23, 312. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.T.; Liu, T.T.; Wu, M.Y.; Chen, Q.X.; Zhuang, J.X.; Wang, Q. Identifying FBLN1 (Gene ID: 2192) as a Potential Melanoma Biomarker for Melanoma based on an Analysis of microRNA Expression Profiles in the GEO and TCGA Databases. Genet Test Mol. Biomark. 2021, 25, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Pal, T.; Bonner, D.; Kim, J.; Monteiro, A.N.; Kessler, L.; Royer, R.; Narod, S.A.; Vadaparampil, S.T. Early onset breast cancer in a registry-based sample of African-American women: BRCA mutation prevalence, and other personal and system-level clinical characteristics. Breast J. 2013, 19, 189–192. [Google Scholar] [CrossRef]
- Pal, T.; Bonner, D.; Cragun, D.; Monteiro, A.N.; Phelan, C.; Servais, L.; Kim, J.; Narod, S.A.; Akbari, M.R.; Vadaparampil, S.T. A high frequency of BRCA mutations in young black women with breast cancer residing in Florida. Cancer 2015, 121, 4173–4180. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.; McCarthy, A.M.; Kim, Y.; Armstrong, K. Predictors of BRCA 1/2 genetic testing among Black women with breast cancer: A population-based study. Cancer Med. 2017, 6, 1787–1798. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, C.E.; Siegel, R.L.; Sauer, A.G.; Miller, K.D.; Fedewa, S.A.; Alcaraz, K.I.; Jemal, A. Cancer statistics for African Americans, 2016: Progress and opportunities in reducing racial disparities. CA A Cancer J. Clin. 2016, 66, 290–308. [Google Scholar] [CrossRef] [PubMed]
- Alfarsi, L.H.; Ansari, R.E.; Craze, M.L.; Toss, M.S.; Masisi, B.; Ellis, I.O.; Rakha, E.A.; Green, A.R. CDC20 expression in oestrogen receptor positive breast cancer predicts poor prognosis and lack of response to endocrine therapy. Breast Cancer Res. Treat 2019, 178, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Xu, Y.; Woo, J.-H.; Wang, Y.; Bae, Y.K.; Yoon, D.-S.; Wersto, R.P.; Tully, E.; Wilsbach, K.; Gabrielson, E. Increased expression of mitotic checkpoint genes in breast cancer cells with chromosomal instability. Clin. Cancer Res. 2006, 12, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Momiyama, N.; Ueda, M.; Matsuyama, R.; Mori, R.; Fujii, Y.; Ichikawa, Y.; Endo, I.; Togo, S.; Shimada, H. Targeting of CDC20 via small interfering RNA causes enhancement of the cytotoxicity of chemoradiation. Anticancer. Res. 2008, 28, 1559–1563. [Google Scholar] [PubMed]
- Kidokoro, T.; Tanikawa, C.; Furukawa, Y.; Katagiri, T.; Nakamura, Y.; Matsuda, K. CDC20, a potential cancer therapeutic target, is negatively regulated by p53. Oncogene 2008, 27, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Hadjihannas, M.V.; Bernkopf, D.B.; Brückner, M.; Behrens, J. Cell cycle control of Wnt/β-catenin signalling by conductin/axin2 through CDC20. EMBO Rep. 2012, 13, 347–354. [Google Scholar] [CrossRef]
- Li, J.; Gao, J.Z.; Du, J.L.; Huang, Z.X.; Wei, L.X. Increased CDC20 expression is associated with development and progression of hepatocellular carcinoma. Int. J. Oncol. 2014, 45, 1547–1555. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.Y.; Wu, H.R.; Zhang, J.M.; Huang, G.R.; Ji, D.D. Expression characteristics of CDC20 in gastric cancer and its correlation with poor prognosis. Int. J. Clin. Exp. Pathol. 2014, 7, 722–727. [Google Scholar] [PubMed]
- Jouali, F.; El Ansari, F.Z.; Marchoudi, N.; Barakat, A.; Zmaimita, H.; Samlali, H.; Fekkak, J. EGFR, BRCA1, BRCA2 and TP53 genetic profile in Moroccan triple negative breast cancer cases. Int. J. Mol. Epidemiol. Genet 2020, 11, 16–25. [Google Scholar] [PubMed]
- Lane, D.P. Cancer. p53, guardian of the genome. Nature 1992, 358, 15–16. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.L.; Levine, A.J. The p53 pathway: Positive and negative feedback loops. Oncogene 2005, 24, 2899–2908. [Google Scholar] [CrossRef] [PubMed]
- Okamura, Y.; Nomoto, S.; Kanda, M.; Hayashi, M.; Nishikawa, Y.; Fujii, T.; Sugimoto, H.; Takeda, S.; Nakao, A. Reduced expression of reelin (RELN) gene is associated with high recurrence rate of hepatocellular carcinoma. Ann. Surg. Oncol. 2011, 18, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M. Function and cancer genomics of FAT family genes (review). Int. J. Oncol. 2012, 41, 1913–1918. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Cui, Y.; Shen, Y.; Li, Y.; Gao, X.; Xi, Y.; Wang, T. Identifying Mutually Exclusive Gene Sets with Prognostic Value and Novel Potential Driver Genes in Patients with Glioblastoma. Biomed. Res. Int. 2019, 2019, 4860367. [Google Scholar] [CrossRef] [PubMed]
- Menzl, I.; Lebeau, L.; Pandey, R.; Hassounah, N.B.; Li, F.W.; Nagle, R.; Weihs, K.; McDermott, K.M. Loss of primary cilia occurs early in breast cancer development. Cilia 2014, 3, 7. [Google Scholar] [CrossRef]
- Tinholt, M.; Viken, M.K.; Dahm, A.E.; Vollan, H.K.; Sahlberg, K.K.; Garred, O.; Børresen-Dale, A.L.; Jacobsen, A.F.; Kristensen, V.; Bukholm, I.; et al. Increased coagulation activity and genetic polymorphisms in the F5, F10 and EPCR genes are associated with breast cancer: A case-control study. BMC Cancer 2014, 14, 845. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, A.; Ramar, K.; Ramalingam, S. Computational Identification of Novel Stage-Specific Biomarkers in Colorectal Cancer Progression. PLoS ONE 2016, 11, e0156665. [Google Scholar] [CrossRef]
- Iwakawa, R.; Kohno, T.; Totoki, Y.; Shibata, T.; Tsuchihara, K.; Mimaki, S.; Tsuta, K.; Narita, Y.; Nishikawa, R.; Noguchi, M.; et al. Expression and clinical significance of genes frequently mutated in small cell lung cancers defined by whole exome/RNA sequencing. Carcinogenesis 2015, 36, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Tinholt, M.; Garred, Ø.; Borgen, E.; Beraki, E.; Schlichting, E.; Kristensen, V.; Sahlberg, K.K.; Iversen, N. Subtype-specific clinical and prognostic relevance of tumor-expressed F5 and regulatory F5 variants in breast cancer: The CoCaV study. J. Thromb Haemost 2018, 16, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Tinholt, M.; Stavik, B.; Tekpli, X.; Garred, Ø.; Borgen, E.; Kristensen, V.; Sahlberg, K.K.; Sandset, P.M.; Iversen, N. Coagulation factor V is a marker of tumor-infiltrating immune cells in breast cancer. Oncoimmunology 2020, 9, 1824644. [Google Scholar] [CrossRef]
- Gao, C.; Li, H.; Liu, C.; Xu, X.; Zhuang, J.; Zhou, C.; Liu, L.; Feng, F.; Sun, C. Tumor Mutation Burden and Immune Invasion Characteristics in Triple Negative Breast Cancer: Genome High-Throughput Data Analysis. Front. Immunol. 2021, 12, 650491. [Google Scholar] [CrossRef] [PubMed]
- Budczies, J.; Bockmayr, M.; Denkert, C.; Klauschen, F.; Lennerz, J.K.; Györffy, B.; Dietel, M.; Loibl, S.; Weichert, W.; Stenzinger, A. Classical pathology and mutational load of breast cancer—Integration of two worlds. J. Pathol. Clin. Res. 2015, 1, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, X.; An, N.; Liu, J. Bioinformatics analysis of BUB1 expression and gene regulation network in lung adenocarcinoma. Transl. Cancer Res. 2020, 9, 4820. [Google Scholar] [CrossRef] [PubMed]
- Baron, A.P.; von Schubert, C.; Cubizolles, F.; Siemeister, G.; Hitchcock, M.; Mengel, A.; Schröder, J.; Fernandez-Montalvan, A.; von Nussbaum, F.; Mumberg, D. Probing the catalytic functions of Bub1 kinase using the small molecule inhibitors BAY-320 and BAY-524. Elife 2016, 5, e12187. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, J.A.; van Heesbeen, R.G.; Blomen, V.A.; Janssen, L.M.; van Diemen, F.; Brummelkamp, T.R.; Medema, R.H. BUB1 is essential for the viability of human cells in which the spindle assembly checkpoint is compromised. Cell Rep. 2018, 22, 1424–1438. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y.; Han, Y.K.; Park, G.-Y.; Kim, S.D.; Geun Lee, C. Bub1 is required for maintaining cancer stem cells in breast cancer cell lines. Sci. Rep. 2015, 5, 15993. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, X.; Yang, M.; Yuan, D.; Ye, K.; Qu, X.; Wang, X. BUBs are new biomarkers of promoting tumorigenesis and affecting prognosis in breast cancer. Dis. Markers 2022, 2022, 2760432. [Google Scholar] [CrossRef] [PubMed]
- Indacochea, A.; Guerrero, S.; Ureña, M.; Araujo, F.; Coll, O.; LLeonart, M.E.; Gebauer, F. Cold-inducible RNA binding protein promotes breast cancer cell malignancy by regulating Cystatin C levels. Rna 2021, 27, 190–201. [Google Scholar] [CrossRef] [PubMed]
- García-Cárdenas, J.M.; Guerrero, S.; López-Cortés, A.; Armendáriz-Castillo, I.; Guevara-Ramírez, P.; Pérez-Villa, A.; Yumiceba, V.; Zambrano, A.K.; Leone, P.E.; Paz-y-Miño, C. Post-transcriptional regulation of colorectal cancer: A focus on RNA-binding proteins. Front. Mol. Biosci. 2019, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Järvelin, A.I.; Davis, I.; Bond, G.L.; Castello, A. Expanding horizons: New roles for non-canonical RNA-binding proteins in cancer. Curr. Opin. Genet Dev. 2018, 48, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Sokratous, G.; Polyzoidis, S.; Ashkan, K. Immune infiltration of tumor microenvironment following immunotherapy for glioblastoma multiforme. Hum. Vaccin. Immunother. 2017, 13, 2575–2582. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.B.; Zhu, B.; Lin, W.J.; Gao, H.Y.; Dai, H.; Zheng, L.; Shi, W.H.; Chen, W.X. Identification of prognostic significance of BIRC5 in breast cancer using integrative bioinformatics analysis. Biosci. Rep. 2020, 40, BSR20193678. [Google Scholar] [CrossRef] [PubMed]
- Parmar, M.B.; KC, R.B.; Löbenberg, R.; Uludağ, H. Additive Polyplexes to Undertake siRNA Therapy against CDC20 and Survivin in Breast Cancer Cells. Biomacromolecules 2018, 19, 4193–4206. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Zhang, Y.; Sun, Y. Comprehensive Analysis of the Expression Characteristics of the Enhancer of the Zeste Homolog 2 Gene in Pan-Cancer. Front. Genet 2021, 12, 658241. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Jedinak, A.; Sliva, D. Ganodermanontriol (GDNT) exerts its effect on growth and invasiveness of breast cancer cells through the down-regulation of CDC20 and uPA. Biochem. Biophys. Res. Commun. 2011, 415, 325–329. [Google Scholar] [CrossRef]
- Scott, R.E.; Ghule, P.N.; Stein, J.L.; Stein, G.S. Cell cycle gene expression networks discovered using systems biology: Significance in carcinogenesis. J. Cell Physiol. 2015, 230, 2533–2542. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutation of | Mean Expression (Mutant) | Mean Expression (Wild) | Mutant Number | Wild Number | FC (Mutant/Wild) | Direction | p-Value |
|---|---|---|---|---|---|---|---|
| TP53 | 1975.97 | 693.98 | 336 | 643 | 2.85 | up | 2.5 × 10−67 |
| CDH1 | 566.91 | 1227.02 | 138 | 841 | 2.17 | down | 3.55 × 10−13 |
| PIK3CA | 774.27 | 1311.07 | 323 | 656 | 1.69 | down | 1.02 × 10−12 |
| MAP3K1 | 657.46 | 1176.37 | 80 | 899 | 1.79 | down | 1.29 × 10−5 |
| FAT3 | 1827.86 | 1101.31 | 44 | 935 | 1.66 | up | 4.06 × 10−5 |
| R | 1782.88 | 1112.04 | 32 | 947 | 1.6 | up | 5 × 10−5 |
| SPTA1 | 1919.39 | 1088.12 | 54 | 925 | 1.76 | up | 1.53 × 10−4 |
| GATA3 | 709.19 | 1198.43 | 129 | 850 | 1.69 | down | 1.6 × 10−4 |
| F5 | 1708.62 | 1116.43 | 29 | 950 | 1.53 | up | 1.82 × 10−4 |
| DYNC2H1 | 1964.19 | 1105.91 | 32 | 947 | 1.78 | up | 1.85 × 10−4 |
| Tumor | Normal | Expression | p-Value |
|---|---|---|---|
| BLCA. Tumor (n = 408) | BLCA. Normal (n = 19) | Upregulation | 3.5 × 10−10 |
| BC. Tumor (n = 1093) | BC. Normal (n = 112) | Upregulation | 1.81 × 10−58 |
| CESC. Tumor (n = 304) | CESC. Normal (n = 3) | Upregulation | 2.9 × 10−3 |
| CHOL. Tumor (n = 36) | CHOL. Normal (n = 9) | Upregulation | 2.26 × 10−9 |
| COAD. Tumor (n = 457) | COAD. Normal (n = 41) | Upregulation | 7.13 × 10−17 |
| ESCA. Tumor (n = 184) | ESCA. Normal (n = 11) | Upregulation | 4.11 × 10−8 |
| GBM. Tumor (n = 153) | GBM. Normal (n = 5) | Upregulation | 4.3 × 10−4 |
| HNSC-HPV+. Tumor (n = 97) | HNSC-HPV-. Tumor (n = 421) | Upregulation | 1.4 × 10−5 |
| HNSC. Tumor (n = 520) | HNSC. Normal (n = 44) | Upregulation | 4.34 × 10−24 |
| KICH. Tumor (n = 66) | KICH. Normal (n = 25) | Upregulation | 0.343689 |
| KIRC. Tumor (n = 533) | KIRC. Normal (n = 72) | Upregulation | 3.24 × 10−26 |
| KIRP. Tumor (n = 290) | KIRP. Normal (n = 32) | Upregulation | 1.43 × 10−8 |
| LIHC. Tumor (n = 371) | LIHC. Normal (n = 50) | Upregulation | 5.68 × 10−28 |
| LUAD. Tumor (n = 515) | LUAD. Normal (n = 59) | Upregulation | 1.96 × 10−34 |
| LUSC. Tumor (n = 501) | LUSC. Normal (n = 51) | Upregulation | 1.18 × 10−31 |
| PAAD. Tumor (n = 178) | PAAD. Normal (n = 4) | Upregulation | 6.897 × 10−2 |
| PCPG. Tumor (n = 179) | PCPG. Normal (n = 3) | Upregulation | 2.787 × 10−2 |
| PRAD. Tumor (n = 497) | PRAD. Normal (n = 52) | Upregulation | 1.21 × 10−17 |
| READ. Tumor (n = 166) | READ. Normal (n = 10) | Upregulation | 9.07 × 10−5 |
| SKCM. Tumor (n = 103) | SKCM. Metastasis (n = 368) | Upregulation | 0.313568 |
| STAD. Tumor (n = 415) | STAD. Normal (n = 35) | Upregulation | 2.37 × 10−15 |
| THCA. Tumor (n = 501) | THCA. Normal (n = 59) | Upregulation | 3.99 × 10−3 |
| UCEC. Tumor (n = 545) | UCEC. Normal (n = 35) | Upregulation | 7.21 × 10−22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messeha, S.S.; Zarmouh, N.O.; Maku, H.; Gendy, S.; Yedjou, C.G.; Elhag, R.; Latinwo, L.; Odewumi, C.; Soliman, K.F.A. Prognostic and Therapeutic Implications of Cell Division Cycle 20 Homolog in Breast Cancer. Cancers 2024, 16, 2546. https://doi.org/10.3390/cancers16142546
Messeha SS, Zarmouh NO, Maku H, Gendy S, Yedjou CG, Elhag R, Latinwo L, Odewumi C, Soliman KFA. Prognostic and Therapeutic Implications of Cell Division Cycle 20 Homolog in Breast Cancer. Cancers. 2024; 16(14):2546. https://doi.org/10.3390/cancers16142546
Chicago/Turabian StyleMesseha, Samia S., Najla O. Zarmouh, Henrietta Maku, Sherif Gendy, Clement G. Yedjou, Rashid Elhag, Lekan Latinwo, Caroline Odewumi, and Karam F. A. Soliman. 2024. "Prognostic and Therapeutic Implications of Cell Division Cycle 20 Homolog in Breast Cancer" Cancers 16, no. 14: 2546. https://doi.org/10.3390/cancers16142546
APA StyleMesseha, S. S., Zarmouh, N. O., Maku, H., Gendy, S., Yedjou, C. G., Elhag, R., Latinwo, L., Odewumi, C., & Soliman, K. F. A. (2024). Prognostic and Therapeutic Implications of Cell Division Cycle 20 Homolog in Breast Cancer. Cancers, 16(14), 2546. https://doi.org/10.3390/cancers16142546