
SMAD7 Sustains XIAP Expression and Migration of Colorectal Carcinoma Cells

 ,
,  ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Transfection Protocol
2.3. Wound Scratch Assay
2.4. Real-Time Migration by xCELLigence System
2.5. Confocal Microscopy
2.6. Western Blotting
2.7. Flow Cytometry
2.8. Real-Time PCR
2.9. Transcriptome Analysis
2.10. Gene Expression Profiling Interactive Analysis (GEPIA)
2.11. Statistical Analysis
3. Results
3.1. SMAD7 Knockdown Reduces CRC Cell Migration
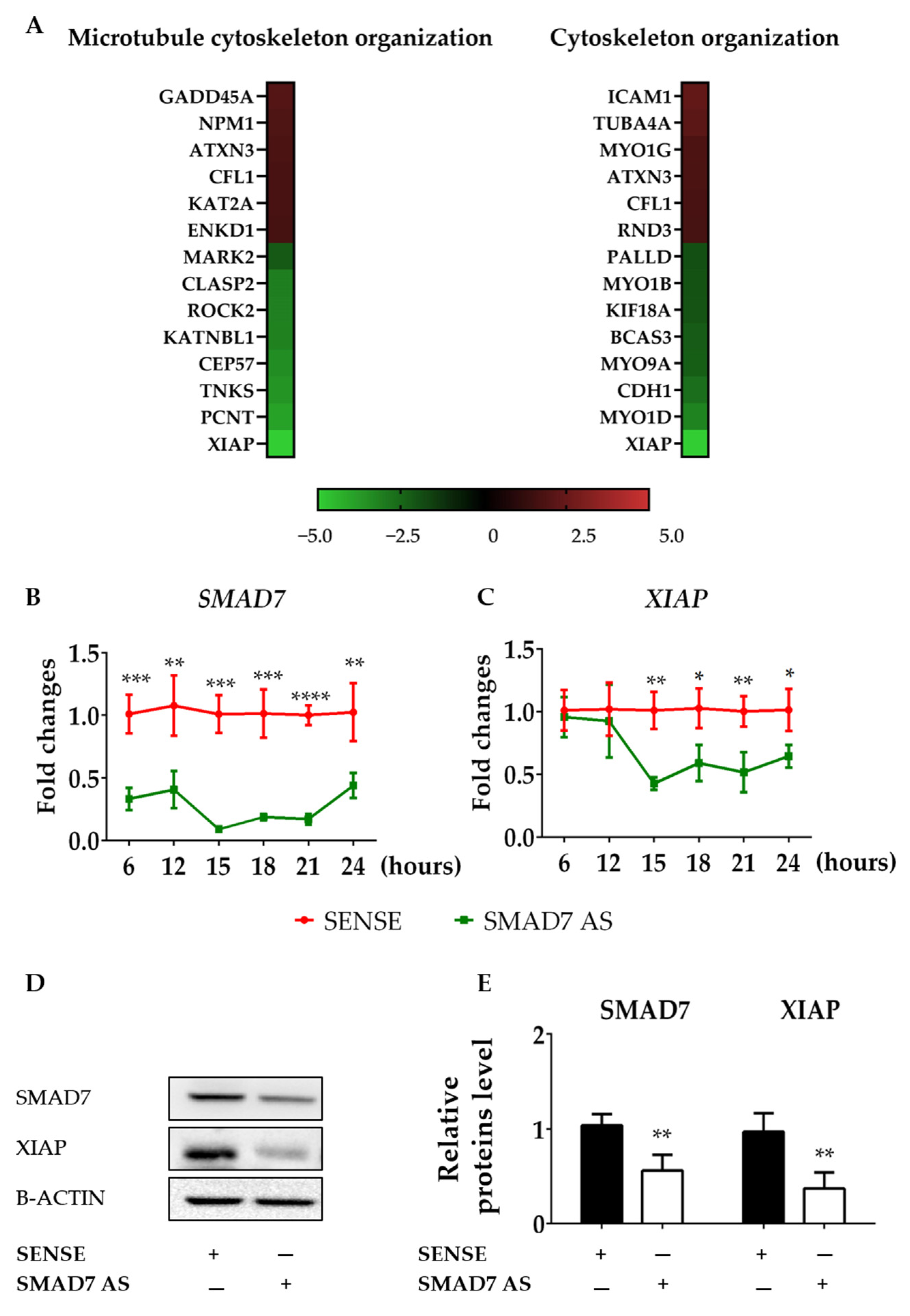
3.2. SMAD7-Deficient CRC Cells Have Reduced Levels of XIAP
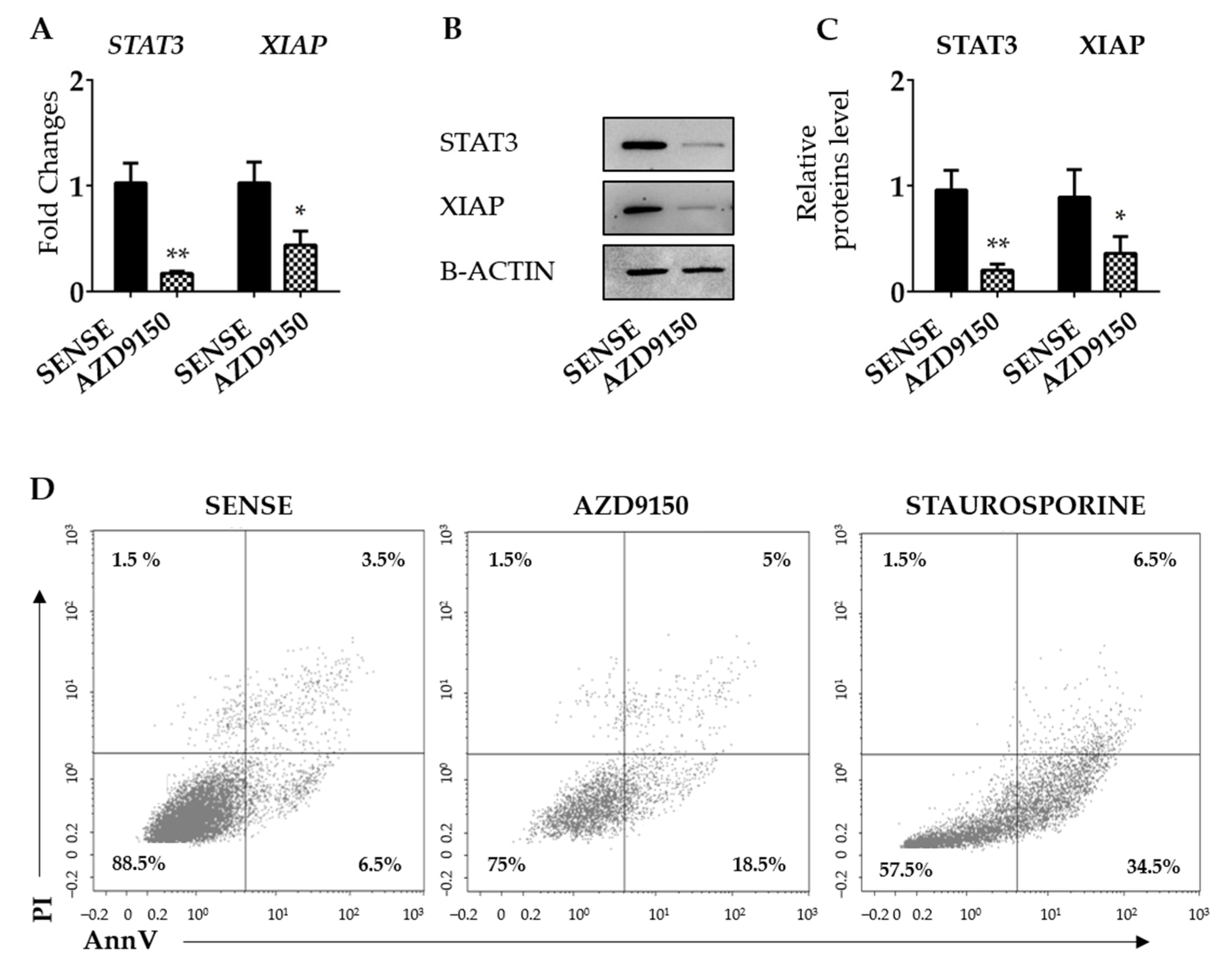
3.3. XIAP Is Regulated by STAT3 in CRC Cells
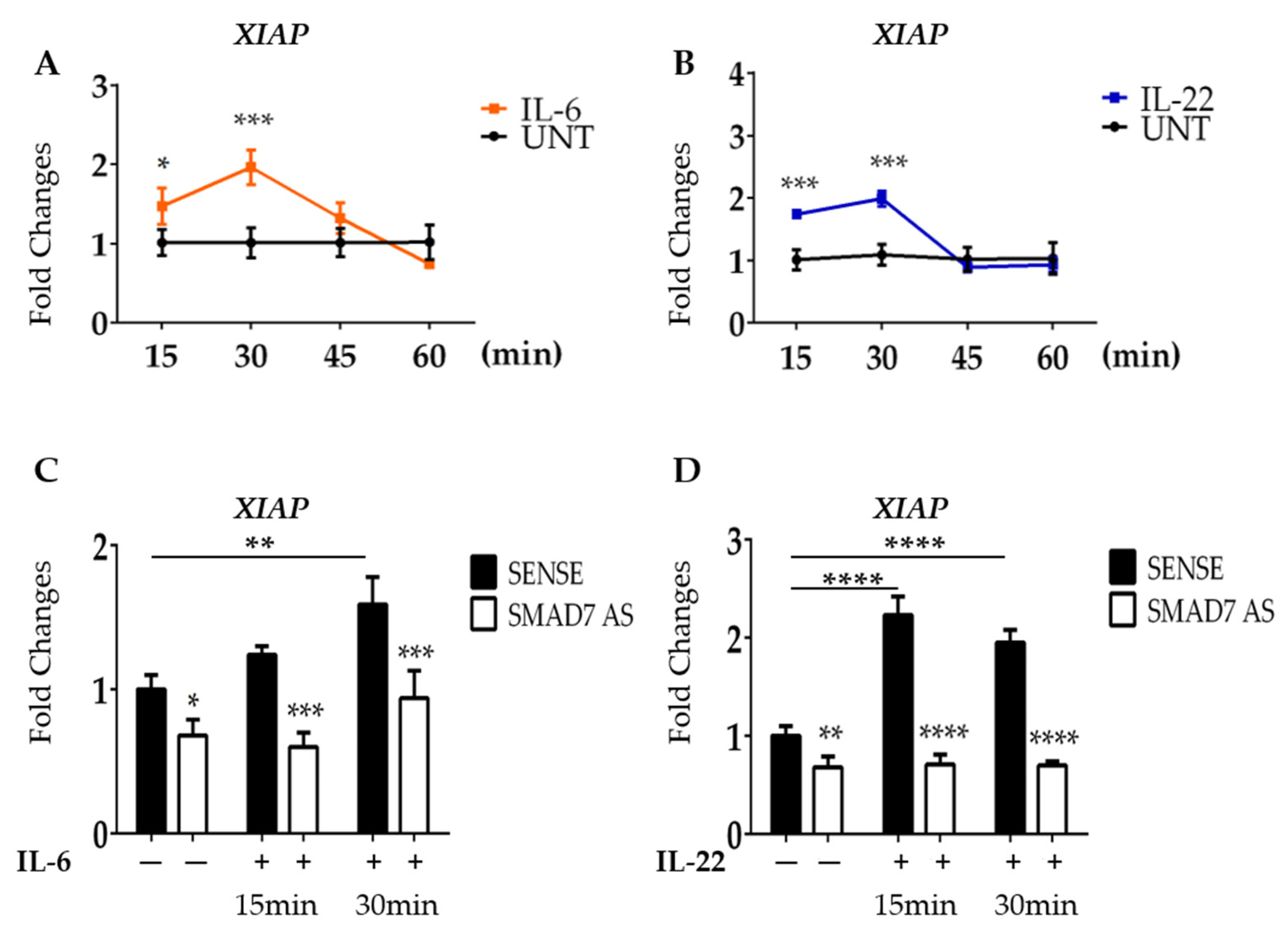
3.4. SMAD7-Deficient Cells Fail to Up-Regulate XIAP following IL-6 and IL-22 Stimulation
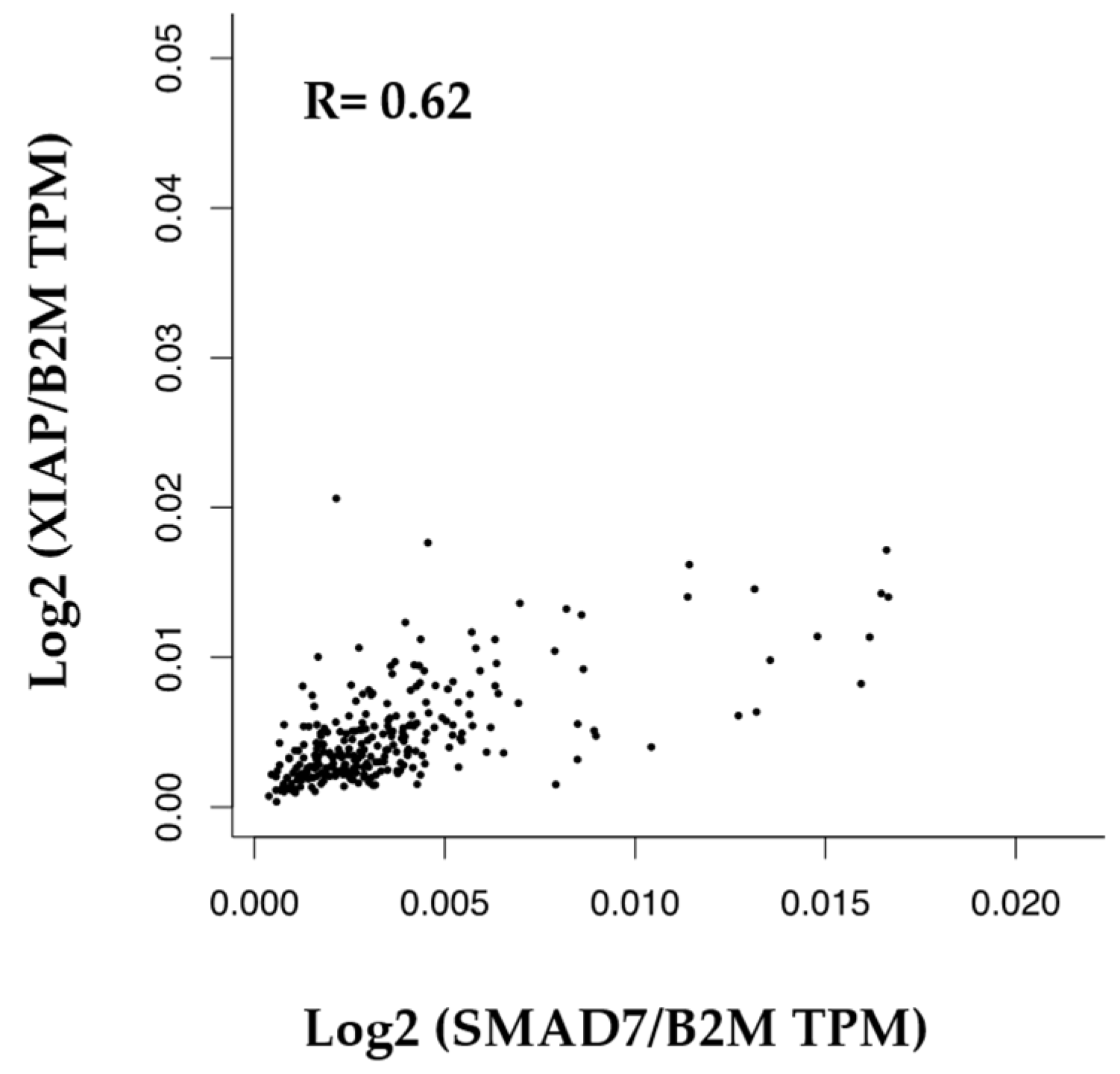
3.5. Correlation between SMAD7 and XIAP in Human CRC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hossain, M.S.; Karuniawati, H.; Jairoun, A.A.; Urbi, Z.; Ooi, D.J.; John, A.; Lim, Y.C.; Kibria, K.M.K.; Mohiuddin, A.K.M.; Ming, L.C.; et al. Colorectal Cancer: A Review of Carcinogenesis, Global Epidemiology, Current Challenges, Risk Factors, Preventive and Treatment Strategies. Cancers 2022, 14, 1732. [Google Scholar] [CrossRef] [PubMed]
- Al-Joufi, F.; Setia, A.; Salem-Bekhit, M.; Sahu, R.; Alqahtani, F.; Widyowati, R.; Aleanizy, F. Molecular Pathogenesis of Colorectal Cancer with an Emphasis on Recent Advances in Biomarkers, as Well as Nanotechnology-Based Diagnostic and Therapeutic Approaches. Nanomaterials 2022, 12, 169. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Castelli, G.; Pelosi, E. Genetic Alterations of Metastatic Colorectal Cancer. Biomedicines 2020, 8, 414. [Google Scholar] [CrossRef] [PubMed]
- Leowattana, W.; Leowattana, P.; Leowattana, T. Systemic Treatment for Metastatic Colorectal Cancer. World J. Gastroenterol. 2023, 29, 1569–1588. [Google Scholar] [CrossRef] [PubMed]
- Hirano, H.; Takashima, A.; Hamaguchi, T.; Shida, D.; Kanemitsu, Y.; the Colorectal Cancer Study Group (CCSG) of the Japan Clinical Oncology Group (JCOG). Current Status and Perspectives of Immune Checkpoint Inhibitors for Colorectal Cancer. Jpn. J. Clin. Oncol. 2021, 51, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Overman, M.J.; Ernstoff, M.S.; Morse, M.A. Where We Stand With Immunotherapy in Colorectal Cancer: Deficient Mismatch Repair, Proficient Mismatch Repair, and Toxicity Management. Am. Soc. Clin. Oncol. Educ. Book. 2018, 38, 239–247. [Google Scholar] [CrossRef]
- Troncone, E.; Monteleone, G. Smad7 and Colorectal Carcinogenesis: A Double-Edged Sword. Cancers 2019, 11, 612. [Google Scholar] [CrossRef] [PubMed]
- Ullman, T.A.; Itzkowitz, S.H. Intestinal Inflammation and Cancer. Gastroenterology 2011, 140, 1807–1816.e1. [Google Scholar] [CrossRef] [PubMed]
- Carethers, J.M.; Jung, B.H. Genetics and Genetic Biomarkers in Sporadic Colorectal Cancer. Gastroenterology 2015, 149, 1177–1190.e3. [Google Scholar] [CrossRef]
- Monahan, K.J.; Swinyard, O.; Latchford, A. Biology of Precancers and Opportunities for Cancer Interception: Lesson from Colorectal Cancer Susceptibility Syndromes. Cancer Prev. Res. 2023, 16, 421–427. [Google Scholar] [CrossRef]
- Thapa, N.; Wen, T.; Cryns, V.L.; Anderson, R.A. Regulation of Cell Adhesion and Migration via Microtubule Cytoskeleton Organization, Cell Polarity, and Phosphoinositide Signaling. Biomolecules 2023, 13, 1430. [Google Scholar] [CrossRef] [PubMed]
- Fife, C.M.; McCarroll, J.A.; Kavallaris, M. Movers and Shakers: Cell Cytoskeleton in Cancer Metastasis. Br. J. Pharmacol. 2014, 171, 5507–5523. [Google Scholar] [CrossRef] [PubMed]
- Hall, A. The Cytoskeleton and Cancer. Cancer Metastasis Rev. 2009, 28, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, D.; Luo, W.; Yu, Y.; Yu, J.; Li, J.; Zhang, X.; Zhang, B.; Chen, J.; Wu, X.-R.; et al. X-Linked Inhibitor of Apoptosis Protein (XIAP) Mediates Cancer Cell Motility via Rho GDP Dissociation Inhibitor (RhoGDI)-Dependent Regulation of the Cytoskeleton. J. Biol. Chem. 2011, 286, 15630–15640. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, B.; Ferreira, C.; Maia, R.; Nestal De Moraes, G. Subcellular Localization of X-Linked Inhibitor of Apoptosis Protein (XIAP) in Cancer: Does That Matter? BBA Adv. 2022, 2, 100050. [Google Scholar] [CrossRef] [PubMed]
- Guoan, X.; Xiaomin, W.; Hanning, W.; Kaiyun, C.; Hao, L. Expression of X-linked Inhibitor of Apoptosis Protein in Human Colorectal Cancer and Its Correlation with Prognosis. J. Surg. Oncol. 2009, 100, 708–712. [Google Scholar] [CrossRef]
- Gargalionis, A.N.; Papavassiliou, K.A.; Papavassiliou, A.G. Targeting STAT3 Signaling Pathway in Colorectal Cancer. Biomedicines 2021, 9, 1016. [Google Scholar] [CrossRef] [PubMed]
- Sim, D.Y.; Lee, H.-J.; Jung, J.H.; Im, E.; Hwang, J.; Kim, D.S.; Kim, S.-H. Suppression of STAT3 Phosphorylation and RelA/P65 Acetylation Mediated by MicroRNA134 Plays a Pivotal Role in the Apoptotic Effect of Lambertianic Acid. Int. J. Mol. Sci. 2019, 20, 2993. [Google Scholar] [CrossRef]
- Stolfi, C.; De Simone, V.; Colantoni, A.; Franzè, E.; Ribichini, E.; Fantini, M.C.; Caruso, R.; Monteleone, I.; Sica, G.S.; Sileri, P.; et al. A Functional Role for Smad7 in Sustaining Colon Cancer Cell Growth and Survival. Cell Death Dis. 2014, 5, e1073. [Google Scholar] [CrossRef]
- Nakao, A.; Afrakhte, M.; Morn, A.; Nakayama, T.; Christian, J.L.; Heuchel, R.; Itoh, S.; Kawabata, M.; Heldin, N.-E.; Heldin, C.-H.; et al. Identification of Smad7, a TGFβ-Inducible Antagonist of TGF-β Signalling. Nature 1997, 389, 631–635. [Google Scholar] [CrossRef]
- Massagué, J. TGFβ in Cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Levy, L.; Hill, C. Alterations in Components of the TGF-β Superfamily Signaling Pathways in Human Cancer. Cytokine Growth Factor. Rev. 2006, 17, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Maresca, C.; Di Maggio, G.; Stolfi, C.; Laudisi, F.; Colella, M.; Pacifico, T.; Di Grazia, A.; Di Fusco, D.; Congiu, D.; Guida, A.M.; et al. Smad7 Sustains Stat3 Expression and Signaling in Colon Cancer Cells. Cancers 2022, 14, 4993. [Google Scholar] [CrossRef] [PubMed]
- Pagliara, V.; De Rosa, M.; Di Donato, P.; Nasso, R.; D’Errico, A.; Cammarota, F.; Poli, A.; Masullo, M.; Arcone, R. Inhibition of Interleukin-6-Induced Matrix Metalloproteinase-2 Expression and Invasive Ability of Lemon Peel Polyphenol Extract in Human Primary Colon Cancer Cells. Molecules 2021, 26, 7076. [Google Scholar] [CrossRef]
- Colella, M.; Cuomo, D.; Nittoli, V.; Amoresano, A.; Porciello, A.; Reale, C.; Roberto, L.; Russo, F.; Russo, N.A.; De Felice, M.; et al. A Cross-Species Analysis Reveals Dysthyroidism of the Ovaries as a Common Trait of Premature Ovarian Aging. Int. J. Mol. Sci. 2023, 24, 3054. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Jiang, Q.; Zhai, X.; Liu, L.; Hong, Y. Screening and Validation of the Optimal Panel of Reference Genes in Colonic Epithelium and Relative Cancer Cell Lines. Sci. Rep. 2023, 13, 17777. [Google Scholar] [CrossRef]
- Rácz, G.A.; Nagy, N.; Tóvári, J.; Apáti, Á.; Vértessy, B.G. Identification of New Reference Genes with Stable Expression Patterns for Gene Expression Studies Using Human Cancer and Normal Cell Lines. Sci. Rep. 2021, 11, 19459. [Google Scholar] [CrossRef]
- Laudisi, F.; Stolfi, C.; Bevivino, G.; Maresca, C.; Franzè, E.; Troncone, E.; Lolli, E.; Marafini, I.; Pietrucci, D.; Teofani, A.; et al. GATA6 Deficiency Leads to Epithelial Barrier Dysfunction and Enhances Susceptibility to Gut Inflammation. J. Crohn’s Colitis 2022, 16, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for Gene List Enrichment Analysis and Candidate Gene Prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef]
- Li, C.; Tang, Z.; Zhang, W.; Ye, Z.; Liu, F. GEPIA2021: Integrating Multiple Deconvolution-Based Analysis into GEPIA. Nucleic Acids Res. 2021, 49, W242–W246. [Google Scholar] [CrossRef]
- Di Fusco, D.; Segreto, M.T.; Di Maggio, G.; Iannucci, A.; Maresca, C.; Di Grazia, A.; Colella, M.; Stolfi, C.; Monteleone, G.; Monteleone, I. Insulin-like Growth Factor II mRNA-Binding Protein 1 Regulates Pancreatic Cancer Cell Growth through the Surveillance of CDC25A mRNA. Cancers 2023, 15, 4983. [Google Scholar] [CrossRef] [PubMed]
- Svitkina, T. The Actin Cytoskeleton and Actin-Based Motility. Cold Spring Harb. Perspect. Biol. 2018, 10, a018267. [Google Scholar] [CrossRef]
- Yu, H.; Lee, H.; Herrmann, A.; Buettner, R.; Jove, R. Revisiting STAT3 Signalling in Cancer: New and Unexpected Biological Functions. Nat. Rev. Cancer 2014, 14, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Corvinus, F.M.; Orth, C.; Moriggl, R.; Tsareva, S.A.; Wagner, S.; Pfitzner, E.B.; Baus, D.; Kaufman, R.; Huber, L.A.; Zatloukal, K.; et al. Persistent STAT3 Activation in Colon Cancer Is Associated with Enhanced Cell Proliferation and Tumor Growth. Neoplasia 2005, 7, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Tolomeo, M.; Cascio, A. The Multifaced Role of STAT3 in Cancer and Its Implication for Anticancer Therapy. Int. J. Mol. Sci. 2021, 22, 603. [Google Scholar] [CrossRef] [PubMed]
- Laudisi, F.; Cherubini, F.; Di Grazia, A.; Dinallo, V.; Di Fusco, D.; Franzè, E.; Ortenzi, A.; Salvatori, I.; Scaricamazza, S.; Monteleone, I.; et al. Progranulin Sustains STAT 3 Hyper-activation and Oncogenic Function in Colorectal Cancer Cells. Mol. Oncol. 2019, 13, 2142–2159. [Google Scholar] [CrossRef] [PubMed]
- De Simone, V.; Franzè, E.; Ronchetti, G.; Colantoni, A.; Fantini, M.C.; Di Fusco, D.; Sica, G.S.; Sileri, P.; MacDonald, T.T.; Pallone, F.; et al. Th17-Type Cytokines, IL-6 and TNF-α Synergistically Activate STAT3 and NF-kB to Promote Colorectal Cancer Cell Growth. Oncogene 2015, 34, 3493–3503. [Google Scholar] [CrossRef]
- Jiang, R.; Wang, H.; Deng, L.; Hou, J.; Shi, R.; Yao, M.; Gao, Y.; Yao, A.; Wang, X.; Yu, L.; et al. IL-22 Is Related to Development of Human Colon Cancer by Activation of STAT3. BMC Cancer 2013, 13, 59. [Google Scholar] [CrossRef]
- Rosic, J.; Dragicevic, S.; Miladinov, M.; Despotovic, J.; Bogdanovic, A.; Krivokapic, Z.; Nikolic, A. SMAD7 and SMAD4 Expression in Colorectal Cancer Progression and Therapy Response. Exp. Mol. Pathol. 2021, 123, 104714. [Google Scholar] [CrossRef]
- Halder, S.K.; Rachakonda, G.; Deane, N.G.; Datta, P.K. Smad7 Induces Hepatic Metastasis in Colorectal Cancer. Br. J. Cancer 2008, 99, 957–965. [Google Scholar] [CrossRef]
- Troncone, E.; Marafini, I.; Stolfi, C.; Monteleone, G. Involvement of Smad7 in Inflammatory Diseases of the Gut and Colon Cancer. Int. J. Mol. Sci. 2021, 22, 3922. [Google Scholar] [CrossRef] [PubMed]
- Planchon, S.; Fiocchi, C.; Takafuji, V.; Roche, J.K. Transforming Growth Factor-?1 Preserves Epithelial Barrier Function: Identification of Receptors, Biochemical Intermediates, and Cytokine Antagonists. J. Cell. Physiol. 1999, 181, 55–66. [Google Scholar] [CrossRef]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [PubMed]
- Mondal, C.; Di Martino, J.S.; Bravo-Cordero, J.J. Actin Dynamics during Tumor Cell Dissemination. In International Review of Cell and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 360, pp. 65–98. ISBN 978-0-12-824138-7. [Google Scholar]
- Deveraux, Q.L. Cleavage of Human Inhibitor of Apoptosis Protein XIAP Results in Fragments with Distinct Specificities for Caspases. EMBO J. 1999, 18, 5242–5251. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.C.; Wilkinson, A.S.; Scott, F.L.; Csomos, R.A.; Salvesen, G.S.; Duckett, C.S. Neutralization of Smac/Diablo by Inhibitors of Apoptosis (IAPs). J. Biol. Chem. 2004, 279, 51082–51090. [Google Scholar] [CrossRef] [PubMed]
- Schimmer, A.D.; Dalili, S.; Batey, R.A.; Riedl, S.J. Targeting XIAP for the Treatment of Malignancy. Cell Death Differ. 2006, 13, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Morizane, Y. X-Linked Inhibitor of Apoptosis Functions as Ubiquitin Ligase toward Mature Caspase-9 and Cytosolic Smac/DIABLO. J. Biochem. 2005, 137, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Kleinberg, L.; Lie, A.K.; Flørenes, V.A.; Nesland, J.M.; Davidson, B. Expression of Inhibitor-of-Apoptosis Protein Family Members in Malignant Mesothelioma. Hum. Pathol. 2007, 38, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Kleinberg, L.; Flørenes, V.A.; Silins, I.; Haug, K.; Trope, C.G.; Nesland, J.M.; Davidson, B. Nuclear Expression of Survivin Is Associated with Improved Survival in Metastatic Ovarian Carcinoma. Cancer 2007, 109, 228–238. [Google Scholar] [CrossRef]
- Akyurek, N.; Ren, Y.; Rassidakis, G.Z.; Schlette, E.J.; Medeiros, L.J. Expression of Inhibitor of Apoptosis Proteins in B-cell non-Hodgkin and Hodgkin Lymphomas. Cancer 2006, 107, 1844–1851. [Google Scholar] [CrossRef]
- Gordon, G.; Mani, M.; Mukhopadhyay, L.; Dong, L.; Edenfield, H.; Glickman, J.; Yeap, B.; Sugarbaker, D.; Bueno, R. Expression Patterns of Inhibitor of Apoptosis Proteins in Malignant Pleural Mesothelioma. J. Pathol. 2007, 211, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, B.; Qu, B.; Xing, H.; Gao, S.; Yin, J.; Wang, X.; Cheng, Y. Silencing STAT3 May Inhibit Cell Growth through Regulating Signaling Pathway, Telomerase, Cell Cycle, Apoptosis and Angiogenesis in Hepatocellular Carcinoma: Potential Uses for Gene Therapy. Neoplasma 2011, 58, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Ravi, R.; Jain, A.J.; Schulick, R.D.; Pham, V.; Prouser, T.S.; Allen, H.; Mayer, E.G.; Yu, H.; Pardoll, D.M.; Ashkenazi, A.; et al. Elimination of Hepatic Metastases of Colon Cancer Cells via P53-Independent Cross-Talk between Irinotecan and Apo2 Ligand/TRAIL. Cancer Res. 2004, 64, 9105–9114. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.; Chu, B.; Wei, S.; Djeu, J.; Epling-Burnette, P.K.; Loughran, T.; Jove, R.; Pledger, W.J. Roscovitine Inhibits STAT5 Activity and Induces Apoptosis in the Human Leukemia Virus Type 1-Transformed Cell Line MT-2. Cancer Res. 2003, 63, 8523–8530. [Google Scholar] [PubMed]
- Liongue, C.; Sobah, M.L.; Ward, A.C. Signal Transducer and Activator of Transcription Proteins at the Nexus of Immunodeficiency, Autoimmunity and Cancer. Biomedicines 2023, 12, 45. [Google Scholar] [CrossRef] [PubMed]
- Carter, B.Z.; Milella, M.; Tsao, T.; McQueen, T.; Schober, W.D.; Hu, W.; Dean, N.M.; Steelman, L.; McCubrey, J.A.; Andreeff, M. Regulation and Targeting of Antiapoptotic XIAP in Acute Myeloid Leukemia. Leukemia 2003, 17, 2081–2089. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Tanaka, T.; Sadik, G.; Baba, M.; Maruyama, D.; Yanagida, K.; Kodama, T.; Morihara, T.; Tagami, S.; Okochi, M.; et al. Protein Kinase C Stabilizes X-linked Inhibitor of Apoptosis Protein (XIAP) through Phosphorylation at Ser 87 to Suppress Apoptotic Cell Death. Psychogeriatrics 2011, 11, 90–97. [Google Scholar] [CrossRef]
- Zhou, H.-Z.; Zeng, H.-Q.; Yuan, D.; Ren, J.-H.; Cheng, S.-T.; Yu, H.-B.; Ren, F.; Wang, Q.; Qin, Y.-P.; Huang, A.-L.; et al. NQO1 Potentiates Apoptosis Evasion and Upregulates XIAP via Inhibiting Proteasome-Mediated Degradation SIRT6 in Hepatocellular Carcinoma. Cell Commun. Signal. 2019, 17, 168. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colella, M.; Iannucci, A.; Maresca, C.; Albano, F.; Mazzoccoli, C.; Laudisi, F.; Monteleone, I.; Monteleone, G. SMAD7 Sustains XIAP Expression and Migration of Colorectal Carcinoma Cells. Cancers 2024, 16, 2370. https://doi.org/10.3390/cancers16132370
Colella M, Iannucci A, Maresca C, Albano F, Mazzoccoli C, Laudisi F, Monteleone I, Monteleone G. SMAD7 Sustains XIAP Expression and Migration of Colorectal Carcinoma Cells. Cancers. 2024; 16(13):2370. https://doi.org/10.3390/cancers16132370
Chicago/Turabian StyleColella, Marco, Andrea Iannucci, Claudia Maresca, Francesco Albano, Carmela Mazzoccoli, Federica Laudisi, Ivan Monteleone, and Giovanni Monteleone. 2024. "SMAD7 Sustains XIAP Expression and Migration of Colorectal Carcinoma Cells" Cancers 16, no. 13: 2370. https://doi.org/10.3390/cancers16132370
APA StyleColella, M., Iannucci, A., Maresca, C., Albano, F., Mazzoccoli, C., Laudisi, F., Monteleone, I., & Monteleone, G. (2024). SMAD7 Sustains XIAP Expression and Migration of Colorectal Carcinoma Cells. Cancers, 16(13), 2370. https://doi.org/10.3390/cancers16132370