
Understanding the Epitranscriptome for Avant-Garde Brain Tumour Diagnostics

 , , ,
, , ,
Abstract
Simple Summary
Abstract
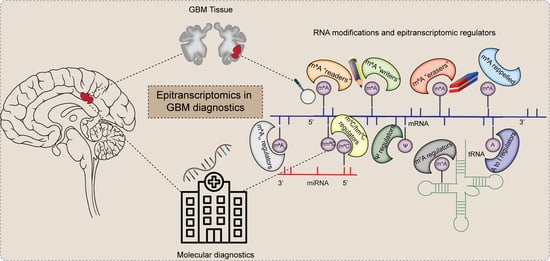
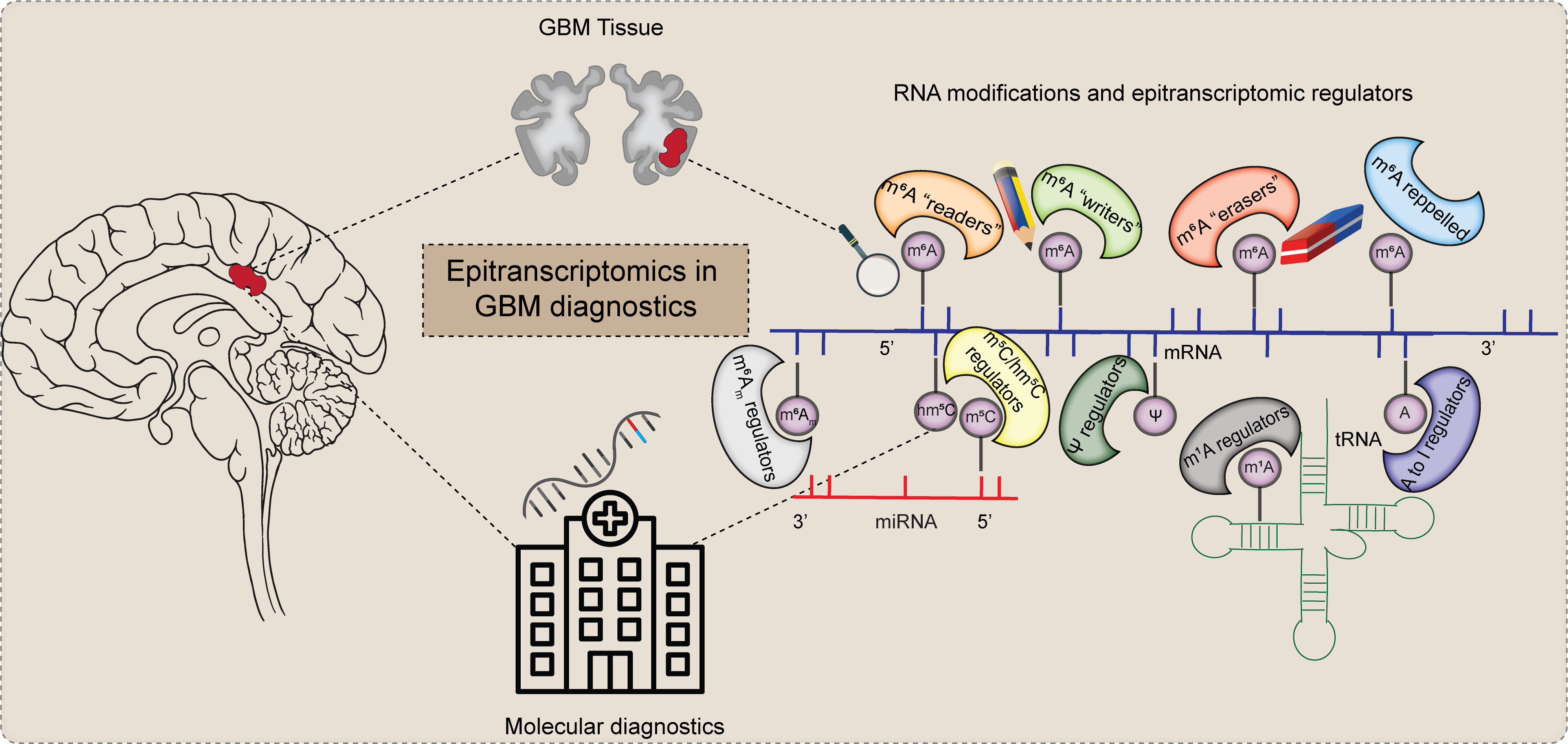
1. Introduction
2. Epitranscriptomic Diversity and the Cellular Roles of RNA Modifications
2.1. N6-Methyladenosine (m6A) RNA Modification
2.1.1. m6A Writers
2.1.2. m6A Readers
2.1.3. m6A ‘Erasers’
2.1.4. Repelled Proteins
2.2. Other RNA Modifications
2.2.1. N6,2-O-Dimethyladenosine: m6Am Modifications
2.2.2. N1-Methyladenosine: m1A Modifications
2.2.3. 5-Methylcytosine (m5C) and 5-Hydroxymethylcytidine (hm5C) Modifications
2.2.4. Adenosine-to-Inosine (A-to-I) Modifications
2.2.5. Pseudouridine (Ψ) Modifications
3. Epitranscriptomics in Neurobiology and Brain Cancer
3.1. m6A Regulators in Neural Development and Physiology
3.2. m6A Regulators in Glioma Stem-like Cells (GSCs) and Tumourigenicity
3.3. m6A Modifications and Regulators in Glioblastoma with Potential Therapeutic Implications for Improving Treatment Response
3.4. Other RNA Modifications Implicated in Glioblastoma Biology and Treatment
4. Epitranscriptomics in Diagnosis
4.1. RNA Methylation Detection Methods
4.2. Epitranscriptomics in Current Diagnostic Approaches
4.3. Epitranscriptomics in Glioma Diagnostics

{kind=link}
{kind=link}
{kind=link}
| Increased m6A Regulator(s) | Decreased m6A Regulator(s) | Data Used | Observations/Role | Ref. |
|---|---|---|---|---|
| YTHDF2 | TCGA, REMBRANDT French, Kawaguchi, Paugh | YTHDF2 is linked to glioma malignancy and invasiveness. | [198] | |
| YTHDF2, YTHDF1, METTL3, RBM15, HNRNPC | ALKBH5, WTAP, YTHDC2, ZC3H13 METTL14, FTO | TCGA | YTHDF1 overexpression correlates with the advanced stage of disease. YTHDF1 contributes to glioma progression. | [100] |
| eIF3e | Oncomine, TCGA | eIF3 subunits show varied expression in distinct regions of GBM tumours. eIF3e proteins expression correlates with glioma grade, highest expression in GBM, and increases in recurrences. eIF3e upregulation in recurrences may have a role in treatment resistance. | [211] | |
| eIF3b, eIF3i, eIF3k and eIF3m (poor OS) | eIF3a and eIF3l (better OS) | CGGA, TCGA | Expression of eIF3d, eIF3e, eIF3f, eIF3h, and eIF3l correlates with the IDH-mutant status of gliomas. eIF3i and eIF3k expressions increase with tumour grade and are associated with poor OS. eIF3i is an independent prognostic factor in IDH-mutant LGG and can predict the 1p/19q codeletion status in IDH-mutant LGG. High eIF3i expression correlates with cell proliferation, mRNA processing, translation, T-cell receptor signalling, NF-kB signalling, and many others. | [202] |
| METTL3 | CGGA, TCGA | METTL3 promotes the malignant progression of gliomas in vitro and in vivo. METTL3 correlates with poor OS in IDH-wildtype but not in IDH-mutant gliomas. | [205] | |
| eIF3A, FMR1, FTO, METTL14, METTL16, METTL3, RBMX, YTHDC, YTHDF3 and ZC3H13 (IDH-mutant vs. IDH-wildtype) | ALKBH5, IGF2BP2, IGF2BP3, RBM15, WTAP and YTHDF1 (IDH-mutant vs. IDH-wildtype) | CGGA, TCGA, REMBRANDT | Expression of m6A regulators is associated with Prognosis, grade, IDH, and 1p/19q status. Lower m6A regulators expression (except FTO) is associated with longer OS. A prognostic risk signature—ALKBH5, IGF2BP3, KIAA1429, and YTHDF2. | [101] |
| ALBKH5, RBM15, YTHDF and WTAP (increased tumour grade) | FTO | CGGA, TCGA | RBM15, METTL3, METTL14, ALKBH5, FTO, YTHDC1, and YTHDF2 are significantly differentially expressed between IDH-mutant and IDH-wildtype LGG. METTL3, FTO, and YTHDC1 are significantly differentially expressed between IDH-mutant and IDH-wildtype GBM. The risk signature comprises RBM15, WTAP, ALBKH5, FTO, YTHDC1, YTHDF1, and YTHDF2, all of which are independent prognostic markers and predictive of clinicopathological features and treatment sensitivity. | [105] |
| RBM15, RBM15B, METTL3, METTL14, WTAP, HNRNPA2B1, HNRNPC, YTHDF1, YTHDF2, YTHDF3, and YTHDC2 (gliomas vs. control) ALKBH5, RBM15, WTAP and YTHDF2 (in GBM vs. LGG) | FTO and ZC3H13 (gliomas vs. control) FTO, KIAA1429, METTL3, ZC3H13, HNRNPC, and YTHDC2 (in GBM vs. LGG) | CGGA | Four m6A-related lncRNAs that have prognostic values: LINC00900 and MIR155HG, increased in higher-grade tumours, while MIR9-3HG and LINC00515 have lower expression in HGG vs. LGG. | [107] |
| IGF2BP3 | YTHDC2 | TCGA, CGGA cBioportal | IGF2BP3 expression increases with tumour grade and correlates with shorter OS. YTHDC2 and IGF2BP3 are negative and positive prognostic factors for OS. | [212] |
| HNRNPC, WTAP, YTHDF2 and, YTHDF1 | TCGA | Defined prognostic risk signature: HNRNPC, ZC3H13, and YTHDF2. HNRNPC plays an important role in malignancy and contributes to the development of gliomas. High expression of HNRNPC correlates with a favourable prognosis. | [213] | |
| LINC00265 | C6orf3, GDNF-AS1, LINC00925, LINC00237 | TCGA, CGGA | Twenty-four prognostic m6A-related lncRNAs were identified as prognostic lncRNAs. m6A-related lncRNA prognostic signature (m6A-LPS). | [107] |
| METTL14, IGF2BP2, IGF2BP3, HNRNPA2B1, YTHDF1, YTHDF3,HNRNPC, RBMX, WTAP, YTHDF2, and IGF2BP1 | TCGA, CGGA | Defined prognostic risk signature: ALKBH5, IGF2BP2, IGF2BP3, HNRNPA2B1, YTHDF1, YTHDF2, RBM15, and WTAP. | [206] |
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goldberg, A.D.; Allis, C.D.; Bernstein, E. Epigenetics: A landscape takes shape. Cell 2007, 128, 635–638. [Google Scholar] [CrossRef]
- Livneh, I.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G.; Dominissini, D. The m(6)A epitranscriptome: Transcriptome plasticity in brain development and function. Nat. Rev. Neurosci. 2020, 21, 36–51. [Google Scholar] [CrossRef] [PubMed]
- He, C. Grand challenge commentary: RNA epigenetics? Nat. Chem. Biol. 2010, 6, 863–865. [Google Scholar] [CrossRef] [PubMed]
- Galardi, S.; Michienzi, A.; Ciafrè, S.A. Insights into the Regulatory Role of m(6)A Epitranscriptome in Glioblastoma. Int. J. Mol. Sci. 2020, 21, 2816. [Google Scholar] [CrossRef]
- Desrosiers, R.; Friderici, K.; Rottman, F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc. Natl. Acad. Sci. USA 1974, 71, 3971–3975. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, K.P. RNA synthesis and processing reactions in a subcellular system from mouse L cells. Hoppe Seylers Z. Physiol. Chem. 1982, 363, 33–43. [Google Scholar] [CrossRef]
- Perry, R.; Kelley, D.J.C. Existence of Methylated Messenger RNA in Mouse L Cells. Cell 1974, 1, 37–42. [Google Scholar] [CrossRef]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vågbø, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N(6)-methyladenosine Modulates Messenger RNA Translation Efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. YTHDF3 facilitates translation and decay of N(6)-methyladenosine-modified RNA. Cell Res. 2017, 27, 315–328. [Google Scholar] [CrossRef]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.B.; Jaffrey, S.R. 5’ UTR m(6)A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Choe, J.; Du, P.; Triboulet, R.; Gregory, R.I. The m(6)A Methyltransferase METTL3 Promotes Translation in Human Cancer Cells. Mol. Cell 2016, 62, 335–345. [Google Scholar] [CrossRef]
- Louloupi, A.; Ntini, E.; Conrad, T.; Ørom, U.A.V. Transient N-6-Methyladenosine Transcriptome Sequencing Reveals a Regulatory Role of m6A in Splicing Efficiency. Cell Rep. 2018, 23, 3429–3437. [Google Scholar] [CrossRef]
- Ke, S.; Alemu, E.A.; Mertens, C.; Gantman, E.C.; Fak, J.J.; Mele, A.; Haripal, B.; Zucker-Scharff, I.; Moore, M.J.; Park, C.Y.; et al. A majority of m6A residues are in the last exons, allowing the potential for 3’ UTR regulation. Genes Dev. 2015, 29, 2037–2053. [Google Scholar] [CrossRef] [PubMed]
- Ping, X.L.; Sun, B.F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Alarcon, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m(6)A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef]
- Kim, T.Y.; Zhong, S.; Fields, C.R.; Kim, J.H.; Robertson, K.D. Epigenomic profiling reveals novel and frequent targets of aberrant DNA methylation-mediated silencing in malignant glioma. Cancer Res. 2006, 66, 7490–7501. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Cui, H. The Emerging Roles of RNA Modifications in Glioblastoma. Cancers 2020, 12, 736. [Google Scholar] [CrossRef]
- Das, K.K.; Kumar, R. Pediatric Glioblastoma. In Glioblastoma; De Vleeschouwer, S., Ed.; Codon Publications: Brisbane, Austrilia, 2017. [Google Scholar]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization classification of tumors of the central nervous system: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA Modifications in Gene Expression Regulation. Cell 2017, 169, 1187–1200. [Google Scholar] [CrossRef]
- Wei, C.M.; Gershowitz, A.; Moss, B. Methylated nucleotides block 5′ terminus of HeLa cell messenger RNA. Cell 1975, 4, 379–386. [Google Scholar] [CrossRef]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, W.V.; Bell, T.A.; Schaening, C. Messenger RNA modifications: Form, distribution, and function. Science 2016, 352, 1408–1412. [Google Scholar] [CrossRef]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Śledź, P.; Jinek, M. Structural insights into the molecular mechanism of the m(6)A writer complex. eLife 2016, 5, e18434. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.; Mumbach, M.R.; Jovanovic, M.; Wang, T.; Maciag, K.; Bushkin, G.G.; Mertins, P.; Ter-Ovanesyan, D.; Habib, N.; Cacchiarelli, D.; et al. Perturbation of m6A writers reveals two distinct classes of mRNA methylation at internal and 5′ sites. Cell Rep. 2014, 8, 284–296. [Google Scholar] [CrossRef]
- Wei, X.; Huo, Y.; Pi, J.; Gao, Y.; Rao, S.; He, M.; Wei, Q.; Song, P.; Chen, Y.; Lu, D.; et al. METTL3 preferentially enhances non-m(6)A translation of epigenetic factors and promotes tumourigenesis. Nat. Cell Biol. 2022, 24, 1278–1290. [Google Scholar] [CrossRef] [PubMed]
- Nance, D.J.; Satterwhite, E.R.; Bhaskar, B.; Misra, S.; Carraway, K.R.; Mansfield, K.D. Characterization of METTL16 as a cytoplasmic RNA binding protein. PloS ONE 2020, 15, e0227647. [Google Scholar] [CrossRef]
- Warda, A.S.; Kretschmer, J.; Hackert, P.; Lenz, C.; Urlaub, H.; Höbartner, C.; Sloan, K.E.; Bohnsack, M.T. Human METTL16 is a N(6)-methyladenosine (m(6)A) methyltransferase that targets pre-mRNAs and various non-coding RNAs. EMBO Rep. 2017, 18, 2004–2014. [Google Scholar] [CrossRef]
- Pendleton, K.E.; Chen, B.; Liu, K.; Hunter, O.V.; Xie, Y.; Tu, B.P.; Conrad, N.K. The U6 snRNA m(6)A Methyltransferase METTL16 Regulates SAM Synthetase Intron Retention. Cell 2017, 169, 824–835.e814. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Dong, L.; Li, Y.; Gao, M.; He, P.C.; Liu, W.; Wei, J.; Zhao, Z.; Gao, L.; Han, L.; et al. METTL16 exerts an m(6)A-independent function to facilitate translation and tumorigenesis. Nat. Cell Biol. 2022, 24, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Lv, R.; Ma, H.; Shen, H.; He, C.; Wang, J.; Jiao, F.; Liu, H.; Yang, P.; Tan, L.; et al. Zc3h13 Regulates Nuclear RNA m(6)A Methylation and Mouse Embryonic Stem Cell Self-Renewal. Mol. Cell 2018, 69, 1028–1038.e1026. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Liu, J.; Cui, X.; Cao, J.; Luo, G.; Zhang, Z.; Cheng, T.; Gao, M.; Shu, X.; Ma, H.; et al. VIRMA mediates preferential m(6)A mRNA methylation in 3’UTR and near stop codon and associates with alternative polyadenylation. Cell Discov. 2018, 4, 10. [Google Scholar] [CrossRef]
- Horiuchi, K.; Kawamura, T.; Iwanari, H.; Ohashi, R.; Naito, M.; Kodama, T.; Hamakubo, T. Identification of Wilms’ tumor 1-associating protein complex and its role in alternative splicing and the cell cycle. J. Biol. Chem. 2013, 288, 33292–33302. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.P.; Chen, C.K.; Pickering, B.F.; Chow, A.; Jackson, C.; Guttman, M.; Jaffrey, S.R. m(6)A RNA methylation promotes XIST-mediated transcriptional repression. Nature 2016, 537, 369–373. [Google Scholar] [CrossRef]
- Liu, J.; Harada, B.T.; He, C. Regulation of Gene Expression by N(6)-methyladenosine in Cancer. Trends Cell Biol. 2019, 29, 487–499. [Google Scholar] [CrossRef]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef]
- Roundtree, I.A.; Luo, G.Z.; Zhang, Z.; Wang, X.; Zhou, T.; Cui, Y.; Sha, J.; Huang, X.; Guerrero, L.; Xie, P.; et al. YTHDC1 mediates nuclear export of N(6)-methyladenosine methylated mRNAs. eLife 2017, 6, e31311. [Google Scholar] [CrossRef]
- Roundtree, I.A.; He, C. Nuclear m(6)A Reader YTHDC1 Regulates mRNA Splicing. Trends Genet. 2016, 32, 320–321. [Google Scholar] [CrossRef]
- Wojtas, M.N.; Pandey, R.R.; Mendel, M.; Homolka, D.; Sachidanandam, R.; Pillai, R.S. Regulation of m(6)A Transcripts by the 3′→5′ RNA Helicase YTHDC2 Is Essential for a Successful Meiotic Program in the Mammalian Germline. Mol. Cell 2017, 68, 374–387.e312. [Google Scholar] [CrossRef]
- Hsu, P.J.; Zhu, Y.; Ma, H.; Guo, Y.; Shi, X.; Liu, Y.; Qi, M.; Lu, Z.; Shi, H.; Wang, J.; et al. Ythdc2 is an N(6)-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res. 2017, 27, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Kang, Y.; Wang, M.; Li, Y.; Xu, T.; Yang, W.; Song, H.; Wu, H.; Shu, Q.; Jin, P. Fragile X mental retardation protein modulates the stability of its m6A-marked messenger RNA targets. Hum. Mol. Genet. 2018, 27, 3936–3950. [Google Scholar] [CrossRef]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef]
- Liu, N.; Zhou, K.I.; Parisien, M.; Dai, Q.; Diatchenko, L.; Pan, T. N6-methyladenosine alters RNA structure to regulate binding of a low-complexity protein. Nucleic Acids Res. 2017, 45, 6051–6063. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Liu, F.; Lu, Z.; Fei, Q.; Ai, Y.; He, P.C.; Shi, H.; Cui, X.; Su, R.; Klungland, A.; et al. Differential m(6)A, m(6)A(m), and m(1)A Demethylation Mediated by FTO in the Cell Nucleus and Cytoplasm. Mol. Cell 2018, 71, 973–985.e975. [Google Scholar] [CrossRef]
- Mauer, J.; Luo, X.; Blanjoie, A.; Jiao, X.; Grozhik, A.V.; Patil, D.P.; Linder, B.; Pickering, B.F.; Vasseur, J.J.; Chen, Q.; et al. Reversible methylation of m(6)A(m) in the 5’ cap controls mRNA stability. Nature 2017, 541, 371–375. [Google Scholar] [CrossRef]
- Edupuganti, R.R.; Geiger, S.; Lindeboom, R.G.H.; Shi, H.; Hsu, P.J.; Lu, Z.; Wang, S.Y.; Baltissen, M.P.A.; Jansen, P.; Rossa, M.; et al. N(6)-methyladenosine (m(6)A) recruits and repels proteins to regulate mRNA homeostasis. Nat. Struct. Mol. Biol. 2017, 24, 870–878. [Google Scholar] [CrossRef]
- Lee, Y.; Choe, J.; Park, O.H.; Kim, Y.K. Molecular Mechanisms Driving mRNA Degradation by m(6)A Modification. Trends Genet. 2020, 36, 177–188. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Toth, J.I.; Petroski, M.D.; Zhang, Z.; Zhao, J.C. N6-methyladenosine modification destabilizes developmental regulators in embryonic stem cells. Nat. Cell Biol. 2014, 16, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, K.; Cai, J.; Zhang, M.; Zhang, X.; Xiong, X.; Meng, H.; Xu, X.; Huang, Z.; Peng, J.; et al. Landscape and Regulation of m(6)A and m(6)Am Methylome across Human and Mouse Tissues. Mol. Cell 2020, 77, 426–440.e426. [Google Scholar] [CrossRef] [PubMed]
- Akichika, S.; Hirano, S.; Shichino, Y.; Suzuki, T.; Nishimasu, H.; Ishitani, R.; Sugita, A.; Hirose, Y.; Iwasaki, S.; Nureki, O.; et al. Cap-specific terminal N (6)-methylation of RNA by an RNA polymerase II-associated methyltransferase. Science 2019, 363. [Google Scholar] [CrossRef]
- Asif, S.; Fatima, R.; Krc, R.; Bennett, J.; Raza, S. Comparative proteogenomic characterization of glioblastoma. CNS Oncol. 2019, 8, Cns37. [Google Scholar] [CrossRef]
- Zhang, C.; Jia, G. Reversible RNA Modification N(1)-methyladenosine (m(1)A) in mRNA and tRNA. Genom. Proteom. Bioinform. 2018, 16, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Portela, A.; Esteller, M. Epigenetic modifications and human disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Yang, W.L.; Zhao, Y.L.; Yang, Y.G. Dynamic transcriptomic m(5) C and its regulatory role in RNA processing. Wiley Interdiscip. Rev. RNA 2021, 12, e1639. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.; Sun, B.F.; Chen, Y.S.; Xu, J.W.; Lai, W.Y.; Li, A.; Wang, X.; Bhattarai, D.P.; Xiao, W.; et al. 5-methylcytosine promotes mRNA export - NSUN2 as the methyltransferase and ALYREF as an m(5)C reader. Cell Res. 2017, 27, 606–625. [Google Scholar] [CrossRef]
- Fu, L.; Guerrero, C.R.; Zhong, N.; Amato, N.J.; Liu, Y.; Liu, S.; Cai, Q.; Ji, D.; Jin, S.G.; Niedernhofer, L.J.; et al. Tet-mediated formation of 5-hydroxymethylcytosine in RNA. J. Am. Chem. Soc. 2014, 136, 11582–11585. [Google Scholar] [CrossRef]
- Bass, B.L. RNA editing by adenosine deaminases that act on RNA. Annu. Rev. Biochem. 2002, 71, 817–846. [Google Scholar] [CrossRef] [PubMed]
- Gerber, A.P.; Keller, W. RNA editing by base deamination: More enzymes, more targets, new mysteries. Trends Biochem. Sci. 2001, 26, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Charette, M.; Gray, M.W. Pseudouridine in RNA: What, where, how, and why. IUBMB Life 2000, 49, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Carlile, T.M.; Rojas-Duran, M.F.; Zinshteyn, B.; Shin, H.; Bartoli, K.M.; Gilbert, W.V. Pseudouridine profiling reveals regulated mRNA pseudouridylation in yeast and human cells. Nature 2014, 515, 143–146. [Google Scholar] [CrossRef]
- Blanco, S.; Dietmann, S.; Flores, J.V.; Hussain, S.; Kutter, C.; Humphreys, P.; Lukk, M.; Lombard, P.; Treps, L.; Popis, M.; et al. Aberrant methylation of tRNAs links cellular stress to neuro-developmental disorders. Embo J. 2014, 33, 2020–2039. [Google Scholar] [CrossRef]
- Park, C.W.; Lee, S.M.; Yoon, K.J. Epitranscriptomic regulation of transcriptome plasticity in development and diseases of the brain. BMB Rep. 2020, 53, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Pupak, A.; Singh, A.; Sancho-Balsells, A.; Alcalá-Vida, R.; Espina, M.; Giralt, A.; Martí, E.; Ørom, U.A.V.; Ginés, S.; Brito, V. Altered m6A RNA methylation contributes to hippocampal memory deficits in Huntington’s disease mice. Cell. Mol. Life Sci. CMLS 2022, 79, 416. [Google Scholar] [CrossRef] [PubMed]
- Ming, Y.; Deng, Z.; Tian, X.; Jia, Y.; Ning, M.; Cheng, S. m6A Methyltransferase METTL3 Reduces Hippocampal Neuron Apoptosis in a Mouse Model of Autism Through the MALAT1/SFRP2/Wnt/β-catenin Axis. Psychiatry Investig. 2022, 19, 771–787. [Google Scholar] [CrossRef] [PubMed]
- Vissers, C.; Sinha, A.; Ming, G.L.; Song, H. The epitranscriptome in stem cell biology and neural development. Neurobiol. Dis. 2020, 146, 105139. [Google Scholar] [CrossRef]
- Chokkalla, A.K.; Mehta, S.L.; Vemuganti, R. Epitranscriptomic regulation by m(6)A RNA methylation in brain development and diseases. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2020, 40, 2331–2349. [Google Scholar] [CrossRef]
- Nie, Y.; Tian, G.G.; Zhang, L.; Lee, T.; Zhang, Z.; Li, J.; Sun, T. Identifying cortical specific long noncoding RNAs modified by m(6)A RNA methylation in mouse brains. Epigenetics 2021, 16, 1260–1276. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Lv, H.; Zhang, W.; Ma, C.; He, X.; Zhao, S.; Zhang, Z.W.; Zeng, Y.X.; Song, S.; Niu, Y.; et al. Region-specific RNA m(6)A methylation represents a new layer of control in the gene regulatory network in the mouse brain. Open Biol. 2017, 7, 170166. [Google Scholar] [CrossRef]
- Ma, C.; Chang, M.; Lv, H.; Zhang, Z.W.; Zhang, W.; He, X.; Wu, G.; Zhao, S.; Zhang, Y.; Wang, D.; et al. RNA m(6)A methylation participates in regulation of postnatal development of the mouse cerebellum. Genome Biol. 2018, 19, 68. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chen, M.; Huang, H.; Zhu, J.; Song, H.; Zhu, J.; Park, J.; Ji, S.J. Dynamic m6A modification regulates local translation of mRNA in axons. Nucleic Acids Res. 2018, 46, 1412–1423. [Google Scholar] [CrossRef] [PubMed]
- Merkurjev, D.; Hong, W.T.; Iida, K.; Oomoto, I.; Goldie, B.J.; Yamaguti, H.; Ohara, T.; Kawaguchi, S.Y.; Hirano, T.; Martin, K.C.; et al. Synaptic N(6)-methyladenosine (m(6)A) epitranscriptome reveals functional partitioning of localized transcripts. Nat. Neurosci. 2018, 21, 1004–1014. [Google Scholar] [CrossRef]
- Choi, H.; Baek, S.; Cho, B.; Kim, S.; Kim, J.; Chang, Y.; Shin, J.; Kim, J. Epitranscriptomic N(6)-Methyladenosine Modification Is Required for Direct Lineage Reprogramming into Neurons. ACS Chem. Biol. 2020, 15, 2087–2097. [Google Scholar] [CrossRef]
- Laug, D.; Glasgow, S.M.; Deneen, B. A glial blueprint for gliomagenesis. Nat. Rev. Neurosci. 2018, 19, 393–403. [Google Scholar] [CrossRef]
- Yoon, K.J.; Ringeling, F.R.; Vissers, C.; Jacob, F.; Pokrass, M.; Jimenez-Cyrus, D.; Su, Y.; Kim, N.S.; Zhu, Y.; Zheng, L.; et al. Temporal Control of Mammalian Cortical Neurogenesis by m(6)A Methylation. Cell 2017, 171, 877–889.e817. [Google Scholar] [CrossRef]
- Xu, H.; Dzhashiashvili, Y.; Shah, A.; Kunjamma, R.B.; Weng, Y.L.; Elbaz, B.; Fei, Q.; Jones, J.S.; Li, Y.I.; Zhuang, X.; et al. m(6)A mRNA Methylation Is Essential for Oligodendrocyte Maturation and CNS Myelination. Neuron 2020, 105, 293–309.e295. [Google Scholar] [CrossRef]
- Wu, R.; Li, A.; Sun, B.; Sun, J.G.; Zhang, J.; Zhang, T.; Chen, Y.; Xiao, Y.; Gao, Y.; Zhang, Q.; et al. A novel m(6)A reader Prrc2a controls oligodendroglial specification and myelination. Cell Res. 2019, 29, 23–41. [Google Scholar] [CrossRef]
- Cui, Q.; Shi, H.; Ye, P.; Li, L.; Qu, Q.; Sun, G.; Sun, G.; Lu, Z.; Huang, Y.; Yang, C.G.; et al. m(6)A RNA Methylation Regulates the Self-Renewal and Tumorigenesis of Glioblastoma Stem Cells. Cell Rep. 2017, 18, 2622–2634. [Google Scholar] [CrossRef]
- Zepecki, J.P.; Karambizi, D.; Fajardo, J.E.; Snyder, K.M.; Guetta-Terrier, C.; Tang, O.Y.; Chen, J.S.; Sarkar, A.; Fiser, A.; Toms, S.A.; et al. miRNA-mediated loss of m6A increases nascent translation in glioblastoma. PLoS Genet. 2021, 17, e1009086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhao, B.S.; Zhou, A.; Lin, K.; Zheng, S.; Lu, Z.; Chen, Y.; Sulman, E.P.; Xie, K.; Bögler, O.; et al. m(6)A Demethylase ALKBH5 Maintains Tumorigenicity of Glioblastoma Stem-like Cells by Sustaining FOXM1 Expression and Cell Proliferation Program. Cancer Cell 2017, 31, 591–606.e596. [Google Scholar] [CrossRef] [PubMed]
- Gong, A.H.; Wei, P.; Zhang, S.; Yao, J.; Yuan, Y.; Zhou, A.D.; Lang, F.F.; Heimberger, A.B.; Rao, G.; Huang, S. FoxM1 Drives a Feed-Forward STAT3-Activation Signaling Loop That Promotes the Self-Renewal and Tumorigenicity of Glioblastoma Stem-like Cells. Cancer Res. 2015, 75, 2337–2348. [Google Scholar] [CrossRef]
- Liu, M.; Dai, B.; Kang, S.H.; Ban, K.; Huang, F.J.; Lang, F.F.; Aldape, K.D.; Xie, T.X.; Pelloski, C.E.; Xie, K.; et al. FoxM1B is overexpressed in human glioblastomas and critically regulates the tumorigenicity of glioma cells. Cancer Res. 2006, 66, 3593–3602. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, C.; Zhang, G. m6A RNA Methylation Controls Proliferation of Human Glioma Cells by Influencing Cell Apoptosis. Cytogenet. Genome Res. 2019, 159, 119–125. [Google Scholar] [CrossRef]
- Malacrida, A.; Rivara, M.; Di Domizio, A.; Cislaghi, G.; Miloso, M.; Zuliani, V.; Nicolini, G. 3D proteome-wide scale screening and activity evaluation of a new ALKBH5 inhibitor in U87 glioblastoma cell line. Bioorg. Med. Chem. 2020, 28, 115300. [Google Scholar] [CrossRef]
- Su, R.; Dong, L.; Li, C.; Nachtergaele, S.; Wunderlich, M.; Qing, Y.; Deng, X.; Wang, Y.; Weng, X.; Hu, C.; et al. R-2HG Exhibits Anti-tumor Activity by Targeting FTO/m(6)A/MYC/CEBPA Signaling. Cell 2018, 172, 90–105.e123. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Xue, Y.; Zheng, J.; Liu, X.; Ma, J.; Liu, Y. WTAP Expression Predicts Poor Prognosis in Malignant Glioma Patients. J. Mol. Neurosci. 2016, 60, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, A.; Patil, V.; Arora, A.; Hegde, A.S.; Arivazhagan, A.; Santosh, V.; Somasundaram, K. Essential role of METTL3-mediated m(6)A modification in glioma stem-like cells maintenance and radioresistance. Oncogene 2018, 37, 522–533. [Google Scholar] [CrossRef]
- Visvanathan, A.; Patil, V.; Abdulla, S.; Hoheisel, J.D.; Somasundaram, K. N⁶-Methyladenosine Landscape of Glioma Stem-Like Cells: METTL3 Is Essential for the Expression of Actively Transcribed Genes and Sustenance of the Oncogenic Signaling. Genes 2019, 10, 141. [Google Scholar] [CrossRef]
- Li, F.; Yi, Y.; Miao, Y.; Long, W.; Long, T.; Chen, S.; Cheng, W.; Zou, C.; Zheng, Y.; Wu, X.; et al. N(6)-Methyladenosine Modulates Nonsense-Mediated mRNA Decay in Human Glioblastoma. Cancer Res. 2019, 79, 5785–5798. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Teng, X.; Zhao, F.; Ma, C.; Zhang, J.; Xiao, L.F.; Wang, Y.; Chang, M.; Tian, Y.; Li, C.; et al. METTL3 regulates m(6)A methylation of PTCH1 and GLI2 in Sonic hedgehog signaling to promote tumor progression in SHH-medulloblastoma. Cell Rep. 2022, 41, 111530. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yin, X.; Zhao, W.; Xu, W.; Chen, L. Molecular mechanism of m(6)A methylation of circDLC1 mediated by RNA methyltransferase METTL3 in the malignant proliferation of glioma cells. Cell Death Discov. 2022, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Lin, Y.; Wei, N. N6-methyladenosine-modified HOTAIRM1 promotes vasculogenic mimicry formation in glioma. Cancer Sci. 2023, 114, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.Q.; Chen, W.; Zhou, J.; Shen, Q.; Sun, Y.; Li, T.; Wang, S.C. N(6)-adenosine-methyltransferase-14 promotes glioma tumorigenesis by repressing argininosuccinate synthase 1 expression in an m6A-dependent manner. Bioengineered 2022, 13, 1858–1871. [Google Scholar] [CrossRef]
- Xiao, L.; Li, X.; Mu, Z.; Zhou, J.; Zhou, P.; Xie, C.; Jiang, S. FTO Inhibition Enhances the Antitumor Effect of Temozolomide by Targeting MYC-miR-155/23a Cluster-MXI1 Feedback Circuit in Glioma. Cancer Res. 2020, 80, 3945–3958. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Dong, L.; Li, Y.; Gao, M.; Han, L.; Wunderlich, M.; Deng, X.; Li, H.; Huang, Y.; Gao, L.; et al. Targeting FTO Suppresses Cancer Stem Cell Maintenance and Immune Evasion. Cancer Cell 2020, 38, 79–96.e11. [Google Scholar] [CrossRef]
- Xu, C.; Yuan, B.; He, T.; Ding, B.; Li, S. Prognostic values of YTHDF1 regulated negatively by mir-3436 in Glioma. J. Cell. Mol. Med. 2020, 24, 7538–7549. [Google Scholar] [CrossRef]
- Xu, S.; Tang, L.; Dai, G.; Luo, C.; Liu, Z. Expression of m6A Regulators Correlated with Immune Microenvironment Predicts Therapeutic Efficacy and Prognosis in Gliomas. Front. Cell Dev. Biol. 2020, 8, 594112. [Google Scholar] [CrossRef]
- Takahashi, H.; Hase, H.; Yoshida, T.; Tashiro, J.; Hirade, Y.; Kitae, K.; Tsujikawa, K. Discovery of two novel ALKBH5 selective inhibitors that exhibit uncompetitive or competitive type and suppress the growth activity of glioblastoma multiforme. Chem. Biol. Drug Des. 2022, 100, 1–12. [Google Scholar] [CrossRef]
- Kowalski-Chauvel, A.; Lacore, M.G.; Arnauduc, F.; Delmas, C.; Toulas, C.; Cohen-Jonathan-Moyal, E.; Seva, C. The m6A RNA Demethylase ALKBH5 Promotes Radioresistance and Invasion Capability of Glioma Stem Cells. Cancers 2020, 13, 40. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Xu, N.; Zhou, J.; He, Z.; Lenahan, C.; Wang, C.; Ji, H.; Liu, B.; Zou, Y.; Zeng, H.; et al. ALKBH5 promotes PD-L1-mediated immune escape through m6A modification of ZDHHC3 in glioma. Cell Death Discov. 2022, 8, 497. [Google Scholar] [CrossRef]
- Chai, R.C.; Wu, F.; Wang, Q.X.; Zhang, S.; Zhang, K.N.; Liu, Y.Q.; Zhao, Z.; Jiang, T.; Wang, Y.Z.; Kang, C.S. m(6)A RNA methylation regulators contribute to malignant progression and have clinical prognostic impact in gliomas. Aging 2019, 11, 1204–1225. [Google Scholar] [CrossRef]
- Tao, M.; Li, X.; He, L.; Rong, X.; Wang, H.; Pan, J.; Lu, Z.; Zhang, X.; Peng, Y. Decreased RNA m(6)A methylation enhances the process of the epithelial mesenchymal transition and vasculogenic mimicry in glioblastoma. Am. J. Cancer Res. 2022, 12, 893–906. [Google Scholar]
- Wang, W.; Li, J.; Lin, F.; Guo, J.; Zhao, J. Identification of N(6)-methyladenosine-related lncRNAs for patients with primary glioblastoma. Neurosurg. Rev. 2021, 44, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Yarmishyn, A.A.; Yang, Y.P.; Lu, K.H.; Chen, Y.C.; Chien, Y.; Chou, S.J.; Tsai, P.H.; Ma, H.I.; Chien, C.S.; Chen, M.T.; et al. Musashi-1 promotes cancer stem cell properties of glioblastoma cells via upregulation of YTHDF1. Cancer Cell Int. 2020, 20, 597. [Google Scholar] [CrossRef]
- Chai, R.C.; Chang, Y.Z.; Chang, X.; Pang, B.; An, S.Y.; Zhang, K.N.; Chang, Y.H.; Jiang, T.; Wang, Y.Z. YTHDF2 facilitates UBXN1 mRNA decay by recognizing METTL3-mediated m(6)A modification to activate NF-κB and promote the malignant progression of glioma. J. Hematol. Oncol. 2021, 14, 109. [Google Scholar] [CrossRef] [PubMed]
- Dixit, D.; Prager, B.C.; Gimple, R.C.; Poh, H.X.; Wang, Y.; Wu, Q.; Qiu, Z.; Kidwell, R.L.; Kim, L.J.Y.; Xie, Q.; et al. The RNA m6A Reader YTHDF2 Maintains Oncogene Expression and Is a Targetable Dependency in Glioblastoma Stem Cells. Cancer Discov. 2021, 11, 480–499. [Google Scholar] [CrossRef]
- Gao, S.; Zhou, J.; Hu, Z.; Zhang, S.; Wu, Y.; Musunuru, P.P.; Zhang, T.; Yang, L.; Luo, X.; Bai, J.; et al. Effects of the m6Am methyltransferase PCIF1 on cell proliferation and survival in gliomas. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166498. [Google Scholar] [CrossRef]
- Cheray, M.; Etcheverry, A.; Jacques, C.; Pacaud, R.; Bougras-Cartron, G.; Aubry, M.; Denoual, F.; Peterlongo, P.; Nadaradjane, A.; Briand, J.; et al. Cytosine methylation of mature microRNAs inhibits their functions and is associated with poor prognosis in glioblastoma multiforme. Mol. Cancer 2020, 19, 36. [Google Scholar] [CrossRef]
- Carella, A.; Tejedor, J.R.; García, M.G.; Urdinguio, R.G.; Bayón, G.F.; Sierra, M.; López, V.; García-Toraño, E.; Santamarina-Ojeda, P.; Pérez, R.F.; et al. Epigenetic downregulation of TET3 reduces genome-wide 5hmC levels and promotes glioblastoma tumorigenesis. Int. J. Cancer 2020, 146, 373–387. [Google Scholar] [CrossRef] [PubMed]
- García, M.G.; Carella, A.; Urdinguio, R.G.; Bayón, G.F.; Lopez, V.; Tejedor, J.R.; Sierra, M.I.; García-Toraño, E.; Santamarina, P.; Perez, R.F.; et al. Epigenetic dysregulation of TET2 in human glioblastoma. Oncotarget 2018, 9, 25922–25934. [Google Scholar] [CrossRef]
- Takai, H.; Masuda, K.; Sato, T.; Sakaguchi, Y.; Suzuki, T.; Suzuki, T.; Koyama-Nasu, R.; Nasu-Nishimura, Y.; Katou, Y.; Ogawa, H.; et al. 5-Hydroxymethylcytosine plays a critical role in glioblastomagenesis by recruiting the CHTOP-methylosome complex. Cell Rep. 2014, 9, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Paz, N.; Levanon, E.Y.; Amariglio, N.; Heimberger, A.B.; Ram, Z.; Constantini, S.; Barbash, Z.S.; Adamsky, K.; Safran, M.; Hirschberg, A.; et al. Altered adenosine-to-inosine RNA editing in human cancer. Genome Res. 2007, 17, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Li, Z.; Du, C.; Qi, B.; Zhao, X.; Wang, L.; Bi, L.; Wang, G.; Zhang, X.; Su, X.; et al. Abnormal expression of an ADAR2 alternative splicing variant in gliomas downregulates adenosine-to-inosine RNA editing. Acta Neurochir. 2014, 156, 1135–1142. [Google Scholar] [CrossRef]
- Galeano, F.; Rossetti, C.; Tomaselli, S.; Cifaldi, L.; Lezzerini, M.; Pezzullo, M.; Boldrini, R.; Massimi, L.; Di Rocco, C.M.; Locatelli, F.; et al. ADAR2-editing activity inhibits glioblastoma growth through the modulation of the CDC14B/Skp2/p21/p27 axis. Oncogene 2013, 32, 998–1009. [Google Scholar] [CrossRef]
- Rintala-Dempsey, A.C.; Kothe, U. Eukaryotic stand-alone pseudouridine synthases - RNA modifying enzymes and emerging regulators of gene expression? RNA Biol. 2017, 14, 1185–1196. [Google Scholar] [CrossRef]
- Miao, F.A.; Chu, K.; Chen, H.R.; Zhang, M.; Shi, P.C.; Bai, J.; You, Y.P. Increased DKC1 expression in glioma and its significance in tumor cell proliferation, migration and invasion. Invest. New Drugs 2019, 37, 1177–1186. [Google Scholar] [CrossRef]
- Cui, Q.; Yin, K.; Zhang, X.; Ye, P.; Chen, X.; Chao, J.; Meng, H.; Wei, J.; Roeth, D.; Li, L.; et al. Targeting PUS7 suppresses tRNA pseudouridylation and glioblastoma tumorigenesis. Nat. Cancer 2021, 2, 932–949. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, L.; Li, X. Detection technologies for RNA modifications. Exp. Mol. Med. 2022, 54, 1601–1616. [Google Scholar] [CrossRef]
- Du, Y.; Xia, M.; Hu, Z. Analysis of N 6-methyladenosine RNA Modification Levels by Dot Blotting. Bio-Protoc. 2022, 12, e4565. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liang, Z.; Yu, H. Dot Blot Analysis of N6-methyladenosine RNA Modification Levels. Bio-Protoc. 2017, 7, e2095. [Google Scholar] [CrossRef]
- Heiss, M.; Borland, K.; Yoluç, Y.; Kellner, S. Quantification of Modified Nucleosides in the Context of NAIL-MS. Methods Mol. Biol. 2021, 2298, 279–306. [Google Scholar] [CrossRef]
- Keith, G. Mobilities of modified ribonucleotides on two-dimensional cellulose thin-layer chromatography. Biochimie 1995, 77, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Olazagoitia-Garmendia, A.; Castellanos-Rubio, A. Relative Quantification of Residue-Specific m(6)A RNA Methylation Using m(6)A-RT-QPCR. Methods Mol. Biol. 2021, 2298, 185–195. [Google Scholar] [CrossRef]
- Shu, X.; Cao, J.; Cheng, M.; Xiang, S.; Gao, M.; Li, T.; Ying, X.; Wang, F.; Yue, Y.; Lu, Z.; et al. A metabolic labeling method detects m(6)A transcriptome-wide at single base resolution. Nat. Chem. Biol. 2020, 16, 887–895. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, L.Q.; Zhao, Y.L.; Yang, C.G.; Roundtree, I.A.; Zhang, Z.; Ren, J.; Xie, W.; He, C.; Luo, G.Z. Single-base mapping of m(6)A by an antibody-independent method. Sci. Adv. 2019, 5, eaax0250. [Google Scholar] [CrossRef]
- Garcia-Campos, M.A.; Edelheit, S.; Toth, U.; Safra, M.; Shachar, R.; Viukov, S.; Winkler, R.; Nir, R.; Lasman, L.; Brandis, A.; et al. Deciphering the “m(6)A Code” via Antibody-Independent Quantitative Profiling. Cell 2019, 178, 731–747.e716. [Google Scholar] [CrossRef]
- Meyer, K.D. DART-seq: An antibody-free method for global m(6)A detection. Nat. Methods 2019, 16, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Linder, B.; Grozhik, A.V.; Olarerin-George, A.O.; Meydan, C.; Mason, C.E.; Jaffrey, S.R. Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome. Nat. Methods 2015, 12, 767–772. [Google Scholar] [CrossRef]
- Ge, R.; Ye, C.; Peng, Y.; Dai, Q.; Zhao, Y.; Liu, S.; Wang, P.; Hu, L.; He, C. m(6)A-SAC-seq for quantitative whole transcriptome m(6)A profiling. Nat. Protoc. 2023, 8, 626–657. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.L.; Liu, S.; Ge, R.; Wu, Y.; He, C.; Chen, M.; Tang, W. Transcriptome-wide profiling and quantification of N(6)-methyladenosine by enzyme-assisted adenosine deamination. Nat. Biotechnol. 2023, Online ahead of print. [Google Scholar] [CrossRef]
- Liu, C.; Sun, H.; Yi, Y.; Shen, W.; Li, K.; Xiao, Y.; Li, F.; Li, Y.; Hou, Y.; Lu, B.; et al. Absolute quantification of single-base m(6)A methylation in the mammalian transcriptome using GLORI. Nat. Biotechnol. 2022, Online ahead of print. [Google Scholar] [CrossRef]
- Liu, H.; Begik, O.; Lucas, M.C.; Ramirez, J.M.; Mason, C.E.; Wiener, D.; Schwartz, S.; Mattick, J.S.; Smith, M.A.; Novoa, E.M. Accurate detection of m(6)A RNA modifications in native RNA sequences. Nat. Commun. 2019, 10, 4079. [Google Scholar] [CrossRef]
- Begik, O.; Mattick, J.S.; Novoa, E.M. Exploring the epitranscriptome by native RNA sequencing. RNA 2022, 28, 1430–1439. [Google Scholar] [CrossRef] [PubMed]
- Leger, A.; Amaral, P.P.; Pandolfini, L.; Capitanchik, C.; Capraro, F.; Miano, V.; Migliori, V.; Toolan-Kerr, P.; Sideri, T.; Enright, A.J.; et al. RNA modifications detection by comparative Nanopore direct RNA sequencing. Nat. Commun. 2021, 12, 7198. [Google Scholar] [CrossRef] [PubMed]
- Jenjaroenpun, P.; Wongsurawat, T.; Wadley, T.D.; Wassenaar, T.M.; Liu, J.; Dai, Q.; Wanchai, V.; Akel, N.S.; Jamshidi-Parsian, A.; Franco, A.T.; et al. Decoding the epitranscriptional landscape from native RNA sequences. Nucleic Acids Res. 2021, 49, e7. [Google Scholar] [CrossRef]
- Pryszcz, L.P.; Novoa, E.M. ModPhred: An integrative toolkit for the analysis and storage of nanopore sequencing DNA and RNA modification data. Bioinformatics 2021, 38, 257–260. [Google Scholar] [CrossRef]
- Cozzuto, L.; Liu, H.; Pryszcz, L.P.; Pulido, T.H.; Delgado-Tejedor, A.; Ponomarenko, J.; Novoa, E.M. MasterOfPores: A Workflow for the Analysis of Oxford Nanopore Direct RNA Sequencing Datasets. Front. Genet. 2020, 11, 211. [Google Scholar] [CrossRef]
- Pei, Y.; Lou, X.; Li, K.; Xu, X.; Guo, Y.; Xu, D.; Yang, Z.; Xu, D.; Cui, W.; Zhang, D. Peripheral Blood Leukocyte N6-methyladenosine is a Noninvasive Biomarker for Non-small-cell Lung Carcinoma. OncoTargets Ther. 2020, 13, 11913–11921. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Fan, X.; Zhang, R.; Wu, G. Upregulated N6-Methyladenosine RNA in Peripheral Blood: Potential Diagnostic Biomarker for Breast Cancer. Cancer Res. Treat. 2021, 53, 399–408. [Google Scholar] [CrossRef]
- Ge, L.; Zhang, N.; Chen, Z.; Song, J.; Wu, Y.; Li, Z.; Chen, F.; Wu, J.; Li, D.; Li, J.; et al. Level of N6-Methyladenosine in Peripheral Blood RNA: A Novel Predictive Biomarker for Gastric Cancer. Clin. Chem. 2020, 66, 342–351. [Google Scholar] [CrossRef]
- Luo, Q.; Gao, Y.; Zhang, L.; Rao, J.; Guo, Y.; Huang, Z.; Li, J. Decreased ALKBH5, FTO, and YTHDF2 in Peripheral Blood Are as Risk Factors for Rheumatoid Arthritis. BioMed Res. Int. 2020, 2020, 5735279. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, S.; Liao, F.; Yang, J.; Liang, T.; Yang, Y.; Huang, X.; Gu, L.; Su, L. Comprehensive Analysis of Blood-Based m6A Methylation in Human Ischemic Stroke. Mol. Neurobiol. 2023, 60, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Huang, W.; Huang, J.T.; Xiong, J.; Yang, Y.; Wu, K.; Jia, G.F.; Chen, J.; Feng, Y.Q.; Yuan, B.F.; et al. Decreased N(6)-methyladenosine in peripheral blood RNA from diabetic patients is associated with FTO expression rather than ALKBH5. J. Clin. Endocrinol. Metab. 2015, 100, E148–E154. [Google Scholar] [CrossRef]
- Min, K.W.; Zealy, R.W.; Davila, S.; Fomin, M.; Cummings, J.C.; Makowsky, D.; McDowell, C.H.; Thigpen, H.; Hafner, M.; Kwon, S.H.; et al. Profiling of m6A RNA modifications identified an age-associated regulation of AGO2 mRNA stability. Aging Cell 2018, 17, e12753. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Xie, Z.; Liao, Y.; Jiang, J.; Dong, W.; Yin, F.; Li, W.X.; Ye, L.; Lin, J.; Liang, H. Systematic analysis of clinical relevance and molecular characterization of m(6)A in COVID-19 patients. Genes Dis. 2022, 9, 1170–1173. [Google Scholar] [CrossRef]
- Kupsco, A.; Gonzalez, G.; Baker, B.H.; Knox, J.M.; Zheng, Y.; Wang, S.; Chang, D.; Schwartz, J.; Hou, L.; Wang, Y.; et al. Associations of smoking and air pollution with peripheral blood RNA N(6)-methyladenosine in the Beijing truck driver air pollution study. Environ. Int. 2020, 144, 106021. [Google Scholar] [CrossRef] [PubMed]
- Naren, D.; Yan, T.; Gong, Y.; Huang, J.; Zhang, D.; Sang, L.; Zheng, X.; Li, Y. High Wilms’ tumor 1 associating protein expression predicts poor prognosis in acute myeloid leukemia and regulates m(6)A methylation of MYC mRNA. J. Cancer Res. Clin. Oncol. 2021, 147, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Huang, Z.; Jiang, P.; Wu, R.; Jiang, H.; Luo, C.; Hong, H.; Yin, H. Elevated N6-Methyladenosine RNA Levels in Peripheral Blood Immune Cells: A Novel Predictive Biomarker and Therapeutic Target for Colorectal Cancer. Front. Immunol 2021, 12, 760747. [Google Scholar] [CrossRef]
- Wang, S.; Xie, X.; Li, C.; Jia, J.; Chen, C. Integrative network analysis of N(6) methylation-related genes reveal potential therapeutic targets for spinal cord injury. Math. Biosci. Eng. 2021, 18, 8174–8187. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Rao, J.; Zhang, L.; Fu, B.; Guo, Y.; Huang, Z.; Li, J. The study of METTL14, ALKBH5, and YTHDF2 in peripheral blood mononuclear cells from systemic lupus erythematosus. Mol. Genet. Genom. Med. 2020, 8, e1298. [Google Scholar] [CrossRef]
- Chang, J.S.; Lin, Z.X.; Liu, Y.J.; Yang, S.M.; Zhang, Y.; Yu, X.Y. Ultra performance liquid chromatography-tandem mass spectrometry assay for the quantification of RNA and DNA methylation. J. Pharm. Biomed. Anal. 2021, 197, 113969. [Google Scholar] [CrossRef]
- Guo, W.; Huai, Q.; Zhang, G.; Guo, L.; Song, P.; Xue, X.; Tan, F.; Xue, Q.; Gao, S.; He, J. Elevated Heterogeneous Nuclear Ribonucleoprotein C Expression Correlates With Poor Prognosis in Patients With Surgically Resected Lung Adenocarcinoma. Front. Oncol. 2020, 10, 598437. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Huai, Q.; Wan, H.; Guo, L.; Song, P.; Gao, S.; He, J. Prognostic Impact of IGF2BP3 Expression in Patients with Surgically Resected Lung Adenocarcinoma. DNA Cell Biol. 2021, 40, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, M.; Hu, D. Deciphering N(6)-Methyladenosine-Related Genes Signature to Predict Survival in Lung Adenocarcinoma. BioMed Res. Int. 2020, 2020, 2514230. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Liu, L.; Li, J.; Hu, Q.; Sun, R. Expression and Prognostic Significance of m6A-Related Genes in Lung Adenocarcinoma. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e919644. [Google Scholar] [CrossRef]
- Luo, Q.; Fu, B.; Zhang, L.; Guo, Y.; Huang, Z.; Li, J. Decreased Peripheral Blood ALKBH5 Correlates with Markers of Autoimmune Response in Systemic Lupus Erythematosus. Dis. Markers 2020, 2020, 8193895. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Gan, L.; Sun, J. Identification and Validation of Three m6A Regulators: FTO, HNRNPC, and HNRNPA2B1 as Potential Biomarkers for Endometriosis. Genes 2022, 14, 86. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.C.; Sun, Q.; Su, W.J.; Li, J.C.; Liu, Y.S.; Zhang, K.; Yang, L.Q. Analysis of m(6)A modulator-mediated methylation modification patterns and the tumor microenvironment in lung adenocarcinoma. Sci. Rep. 2022, 12, 20684. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guo, X.; Zhao, M.; Ao, H.; Leng, X.; Liu, M.; Wu, C.; Ma, J.; Zhu, J. Contributions and prognostic values of m(6) A RNA methylation regulators in non-small-cell lung cancer. J. Cell. Physiol. 2020, 235, 6043–6057. [Google Scholar] [CrossRef]
- Gu, Z.; Yang, Y.; Ma, Q.; Wang, H.; Zhao, S.; Qi, Y.; Li, Y. HNRNPC, a predictor of prognosis and immunotherapy response based on bioinformatics analysis, is related to proliferation and invasion of NSCLC cells. Respir. Res. 2022, 23, 362. [Google Scholar] [CrossRef]
- Xu, F.; Zhang, H.; Chen, J.; Lin, L.; Chen, Y. Immune signature of T follicular helper cells predicts clinical prognostic and therapeutic impact in lung squamous cell carcinoma. Int. Immunopharmacol. 2020, 81, 105932. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Y.; Chen, J.; Peng, C.; Zhang, Y.; Tong, R.; Cheng, Q.; Yang, B.; Feng, X.; Lu, Y.; et al. ALKBH5 suppresses malignancy of hepatocellular carcinoma via m(6)A-guided epigenetic inhibition of LYPD1. Mol. Cancer 2020, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chen, Y.; Mao, Q.; Jiang, X.; Jiang, W.; Chen, J.; Xu, W.; Zhong, L.; Sun, X. Overexpression of YTHDF1 is associated with poor prognosis in patients with hepatocellular carcinoma. Cancer Biomark. Sect. A Dis. Markers 2018, 21, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Qu, N.; Qin, S.; Zhang, X.; Bo, X.; Liu, Z.; Tan, C.; Wen, G.; Jiang, H. Multiple m(6)A RNA methylation modulators promote the malignant progression of hepatocellular carcinoma and affect its clinical prognosis. BMC Cancer 2020, 20, 165. [Google Scholar] [CrossRef]
- Li, Z.; Li, F.; Peng, Y.; Fang, J.; Zhou, J. Identification of three m6A-related mRNAs signature and risk score for the prognostication of hepatocellular carcinoma. Cancer Med. 2020, 9, 1877–1889. [Google Scholar] [CrossRef]
- Lu, J.; Wang, H.; Cao, W.; Chen, D.; He, Z.; Xu, J. Construction of a m5C-related long non-coding RNA signature for the prognosis of hepatocellular carcinoma. Hum. Cell 2022, Online ahead of print. [Google Scholar] [CrossRef]
- Liu, T.; Yang, S.; Cheng, Y.P.; Kong, X.L.; Du, D.D.; Wang, X.; Bai, Y.F.; Yin, L.H.; Pu, Y.P.; Liang, G.Y. The N6-Methyladenosine (m6A) Methylation Gene YTHDF1 Reveals a Potential Diagnostic Role for Gastric Cancer. Cancer Manag. Res. 2020, 12, 11953–11964. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, Q.; Li, B.; Wang, D.; Wang, L.; Zhou, Y.L. m(6)A regulator-mediated methylation modification patterns and tumor microenvironment infiltration characterization in gastric cancer. Mol. Cancer 2020, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chen, C.; Ding, Q.; Zhao, Y.; Wang, Z.; Chen, J.; Jiang, Z.; Zhang, Y.; Xu, G.; Zhang, J.; et al. METTL3-mediated m(6)A modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance. Gut 2020, 69, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, D.; Wang, F.; Xu, Y.; Yu, H.; Zhang, H. Expression of Demethylase Genes, FTO and ALKBH1, Is Associated with Prognosis of Gastric Cancer. Dig. Dis. Sci. 2019, 64, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Feng, S.; Sun, Y.; Jiang, Y.; Tang, C.; Sun, D. Identification of a five m6A-relevant mRNAs signature and risk score for the prognostication of gastric cancer. J. Gastrointest. Oncol 2022, 13, 2234–2248. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wang, Z.; Li, H.; Zhang, H.; Luo, L. Gene Signature and Identification of Clinical Trait-Related m(6) A Regulators in Pancreatic Cancer. Front. Genet. 2020, 11, 522. [Google Scholar] [CrossRef]
- Zhao, Z.; Ju, Q.; Ji, J.; Li, Y.; Zhao, Y. N6-Methyladenosine Methylation Regulator RBM15 is a Potential Prognostic Biomarker and Promotes Cell Proliferation in Pancreatic Adenocarcinoma. Front. Mol. Biosci. 2022, 9, 842833. [Google Scholar] [CrossRef]
- Xu, D.; Shao, J.; Song, H.; Wang, J. The YTH Domain Family of N6-Methyladenosine “Readers” in the Diagnosis and Prognosis of Colonic Adenocarcinoma. BioMed Res. Int. 2020, 2020, 9502560. [Google Scholar] [CrossRef]
- Yang, P.; Wang, Q.; Liu, A.; Zhu, J.; Feng, J. ALKBH5 Holds Prognostic Values and Inhibits the Metastasis of Colon Cancer. Pathol. Oncol. Res. POR 2020, 26, 1615–1623. [Google Scholar] [CrossRef]
- Liao, X.; Chen, L.; Liu, J.; Hu, H.; Hou, D.; You, R.; Wang, X.; Huang, H. m(6)A RNA methylation regulators predict prognosis and indicate characteristics of tumour microenvironment infiltration in acute myeloid leukaemia. Epigenetics 2022, 1–20, Online ahead of print. [Google Scholar] [CrossRef]
- Wang, Z.; Cheng, H.; Xu, H.; Yu, X.; Sui, D. A five-gene signature derived from m6A regulators to improve prognosis prediction of neuroblastoma. Cancer Biomark. Sect. A Dis. Markers 2020, 28, 275–284. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, J.N.; Wang, E.H.; Gong, C.J.; Lan, K.F.; Ding, X. The m6A reader protein YTHDC2 is a potential biomarker and associated with immune infiltration in head and neck squamous cell carcinoma. PeerJ 2020, 8, e10385. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, J.; Liu, M.; Feng, Q.; Peng, H. IGF2BP2 serves as a core m6A regulator in head and neck squamous cell carcinoma. Biosci. Rep. 2022, 42, BSR20221311. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Wan, Q.; Lu, J. The prognostic values of m6A RNA methylation regulators in uveal melanoma. BMC Cancer 2020, 20, 674. [Google Scholar] [CrossRef]
- Li, T.; Gu, M.; Deng, A.; Qian, C. Increased expression of YTHDF1 and HNRNPA2B1 as potent biomarkers for melanoma: A systematic analysis. Cancer Cell Int. 2020, 20, 239. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Bai, J.; Zhang, Y.; Xue, Y.; Peng, Q. N(6)-Methyladenosine regulator RBM15B acts as an independent prognostic biomarker and its clinical significance in uveal melanoma. Front. Immunol. 2022, 13, 918522. [Google Scholar] [CrossRef]
- Li, J.; Rao, B.; Yang, J.; Liu, L.; Huang, M.; Liu, X.; Cui, G.; Li, C.; Han, Q.; Yang, H.; et al. Dysregulated m6A-Related Regulators Are Associated with Tumor Metastasis and Poor Prognosis in Osteosarcoma. Front. Oncol. 2020, 10, 769. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Shan, H.; Zhang, Y.; Fan, Y.; Wu, B. m(6)A RNA methylation regulators have prognostic value in papillary thyroid carcinoma. Am. J. Otolaryngol. 2020, 41, 102547. [Google Scholar] [CrossRef]
- Li, K.; Luo, H.; Luo, H.; Zhu, X. Clinical and prognostic pan-cancer analysis of m6A RNA methylation regulators in four types of endocrine system tumors. Aging 2020, 12, 23931–23944. [Google Scholar] [CrossRef]
- Fu, Y.; Sun, S.; Bi, J.; Kong, C.; Yin, L. Expression patterns and prognostic value of m6A RNA methylation regulators in adrenocortical carcinoma. Medicine 2021, 100, e25031. [Google Scholar] [CrossRef]
- Zhang, Q.J.; Luan, J.C.; Song, L.B.; Cong, R.; Ji, C.J.; Zhou, X.; Xia, J.D.; Song, N.H. m6A RNA methylation regulators correlate with malignant progression and have potential predictive values in clear cell renal cell carcinoma. Exp. Cell Res. 2020, 392, 112015. [Google Scholar] [CrossRef]
- Zheng, Z.; Mao, S.; Guo, Y.; Zhang, W.; Liu, J.; Li, C.; Yao, X. N6-methyladenosine RNA methylation regulators participate in malignant progression and have prognostic value in clear cell renal cell carcinoma. Oncol. Rep. 2020, 43, 1591–1605. [Google Scholar] [CrossRef]
- Strick, A.; von Hagen, F.; Gundert, L.; Klümper, N.; Tolkach, Y.; Schmidt, D.; Kristiansen, G.; Toma, M.; Ritter, M.; Ellinger, J. The N(6) -methyladenosine (m(6) A) erasers alkylation repair homologue 5 (ALKBH5) and fat mass and obesity-associated protein (FTO) are prognostic biomarkers in patients with clear cell renal carcinoma. BJU Int. 2020, 125, 617–624. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, H.; Chen, Q.; Wan, Z.; Gao, X.; Qian, W. Identification of METTL14 in Kidney Renal Clear Cell Carcinoma Using Bioinformatics Analysis. Dis. Markers 2019, 2019, 5648783. [Google Scholar] [CrossRef]
- Wu, J.; Wang, X.; Xu, H.; Tian, J.; Ji, H.; Zhu, J.; Guo, H.; Chen, Z. Bioinformatics analysis of the correlation between m6A RNA methylation regulators and the immune infiltration and prognosis of bladder cancer. Ann. Transl. Med. 2022, 10, 1386. [Google Scholar] [CrossRef]
- Chen, M.; Nie, Z.Y.; Wen, X.H.; Gao, Y.H.; Cao, H.; Zhang, S.F. m6A RNA methylation regulators can contribute to malignant progression and impact the prognosis of bladder cancer. Biosci. Rep. 2019, 39, BSR20192892. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- Fang, R.; Chen, X.; Zhang, S.; Shi, H.; Ye, Y.; Shi, H.; Zou, Z.; Li, P.; Guo, Q.; Ma, L.; et al. EGFR/SRC/ERK-stabilized YTHDF2 promotes cholesterol dysregulation and invasive growth of glioblastoma. Nat. Commun. 2021, 12, 177. [Google Scholar] [CrossRef] [PubMed]
- Hershey, J.W. The role of eIF3 and its individual subunits in cancer. Biochim. Biophys. Acta 2015, 1849, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhang, Y.; Yao, S.; Shi, H.; Huang, X.; Li, Y.; Wei, Y.; Lin, S. The translation initiation factor eIF3i up-regulates vascular endothelial growth factor A, accelerates cell proliferation, and promotes angiogenesis in embryonic development and tumorigenesis. J. Biol. Chem. 2014, 289, 28310–28323. [Google Scholar] [CrossRef]
- Hu, X.; Martinez-Ledesma, E.; Zheng, S.; Kim, H.; Barthel, F.; Jiang, T.; Hess, K.R.; Verhaak, R.G.W. Multigene signature for predicting prognosis of patients with 1p19q co-deletion diffuse glioma. Neuro Oncol. 2017, 19, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.C.; Wang, N.; Chang, Y.Z.; Zhang, K.N.; Li, J.J.; Niu, J.J.; Wu, F.; Liu, Y.Q.; Wang, Y.Z. Systematically profiling the expression of eIF3 subunits in glioma reveals the expression of eIF3i has prognostic value in IDH-mutant lower grade glioma. Cancer Cell Int. 2019, 19, 155. [Google Scholar] [CrossRef]
- Bai, Z.; Wang, X.; Zhang, Z. Establishment and Validation of a 5 m6A RNA Methylation Regulatory Gene Prognostic Model in Low-Grade Glioma. Front. Genet. 2022, 13, 655169. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, N.; Wu, J.; Gao, X.; Zhao, H.; Liu, Z.; Yan, X.; Dong, J.; Wang, F.; Ba, Y.; et al. 5-Methylcytosine Related LncRNAs Reveal Immune Characteristics, Predict Prognosis and Oncology Treatment Outcome in Lower-Grade Gliomas. Front. Immunol. 2022, 13, 844778. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.Z.; Chai, R.C.; Pang, B.; Chang, X.; An, S.Y.; Zhang, K.N.; Jiang, T.; Wang, Y.Z. METTL3 enhances the stability of MALAT1 with the assistance of HuR via m6A modification and activates NF-κB to promote the malignant progression of IDH-wildtype glioma. Cancer Lett. 2021, 511, 36–46. [Google Scholar] [CrossRef]
- Guan, S.; He, Y.; Su, Y.; Zhou, L. A Risk Signature Consisting of Eight m(6)A Methylation Regulators Predicts the Prognosis of Glioma. Cell. Mol. Neurobiol. 2022, 42, 2733–2743. [Google Scholar] [CrossRef]
- Wei, C.; Wang, B.; Peng, D.; Zhang, X.; Li, Z.; Luo, L.; He, Y.; Liang, H.; Du, X.; Li, S.; et al. Pan-Cancer Analysis Shows That ALKBH5 Is a Potential Prognostic and Immunotherapeutic Biomarker for Multiple Cancer Types Including Gliomas. Front. Immunol. 2022, 13, 849592. [Google Scholar] [CrossRef]
- Lin, S.; Xu, H.; Zhang, A.; Ni, Y.; Xu, Y.; Meng, T.; Wang, M.; Lou, M. Prognosis Analysis and Validation of m(6)A Signature and Tumor Immune Microenvironment in Glioma. Front. Oncol. 2020, 10, 541401. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cao, H.; Zhong, Y.; Ji, P.; Chen, F. The Role of m6A Regulator-Mediated Methylation Modification and Tumor Microenvironment Infiltration in Glioblastoma Multiforme. Front. Cell Dev. Biol. 2022, 10, 842835. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.; Li, Y.; Wu, J.; Zhang, B.; Xie, S.; Zheng, X.; Jiang, Z. An m6A/m5C/m1A/m7G-Related Long Non-coding RNA Signature to Predict Prognosis and Immune Features of Glioma. Front. Genet. 2022, 13, 903117. [Google Scholar] [CrossRef]
- Bertorello, J.; Sesen, J.; Gilhodes, J.; Evrard, S.; Courtade-Saïdi, M.; Augustus, M.; Uro-Coste, E.; Toulas, C.; Moyal, E.C.; Seva, C.; et al. Translation reprogramming by eIF3 linked to glioblastoma resistance. NAR Cancer 2020, 2, zcaa020. [Google Scholar] [CrossRef]
- Sun, C.; Zheng, X.; Sun, Y.; Yu, J.; Sheng, M.; Yan, S.; Zhu, Q.; Lan, Q. Identification of IGF2BP3 as an Adverse Prognostic Biomarker of Gliomas. Front. Genet. 2021, 12, 743738. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.C.; Chen, S.H.; Shen, X.L.; Li, D.C.; Liu, H.Y.; Ji, Y.L.; Li, M.; Yu, K.; Yang, H.; Chen, J.J.; et al. M6A RNA Methylation Regulator HNRNPC Contributes to Tumorigenesis and Predicts Prognosis in Glioblastoma Multiforme. Front. Oncol. 2020, 10, 536875. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.; Shi, L.; Ye, Y.; Shi, H.; Zeng, L.; Tiwary, S.; Huse, J.T.; Huo, L.; Ma, L.; Ma, Y.; et al. YTHDF3 Induces the Translation of m(6)A-Enriched Gene Transcripts to Promote Breast Cancer Brain Metastasis. Cancer Cell 2020, 38, 857–871.e857. [Google Scholar] [CrossRef] [PubMed]


| Disease/Condition | Upregulated | Downregulated | m6A Levels | Ref. |
|---|---|---|---|---|
| Non-small-cell lung carcinoma | METTL3, METTL14, RBM15 | ALKBH5, FTO | ↑ | [142] |
| Breast cancer | METTL14 | FTO | ↑ | [143] |
| Gastric cancer | ALKBH5, FTO | ↑ | [144] | |
| Acute myeloid leukaemia | WTAP | [151] | ||
| Colorectal cancer | IGF2BP2 | ↑ | [152] | |
| Rheumatoid arthritis | ALKBH5, FTO, YTHDF2 | ↑ | [145] | |
| Diabetes | FTO | ↓ | [147] | |
| Spinal cord injury | FTO, METTL14, RBMX, YTHDF2, YTHDC2, HNRNPA2B1 | [153] | ||
| Systemic lupus erythematosus | METTL3, METTL14, WTAP, FTO, ALKBH5, YTHDF2 | [154] | ||
| COVID-19 | METTL3, FTO | ↓ | [149] | |
| Aging | ↓ | [148] | ||
| Smoking and air pollution | ↓ | [150] | ||
| Myocardial infarction (in rats) | ↑ | [155] | ||
| Ischemic stroke | ↑ | [146] |
| Disease/Condition | RNA Modification or Epitranscriptomic Regulator | Sample Type | Observations/Role | Ref. |
|---|---|---|---|---|
| Smoking and air pollution | Global m6A levels | Peripheral blood | Decreased global m6A levels were found in smokers compared to non-smokers. | [150] |
| Aging | Global m6A levels | PBMCs | Decrease in overall m6A with aging. m6A-modified transcripts have higher expression than the nonmodified ones. DROSHA and AGO2 have high methylation levels in younger subjects. | [148] |
| COVID-19 | m6A levels, METTL3, FTO | PBMCs | Increased METTL3 and FTO in COVID-19 patients. m6A modification has an essential role in the clinical status of COVID-19 patients | [149] |
| Spinal cord injury (SCI) | FTO, METTL14, RBMX, YTHDF2, YTHDC2, HNRNPA2B1 | PBMCs, GEO data | METTL14, FTO, RBMX, YTHDF2, HNRNPA2B1, and YTHDC2 downregulated in SCI. AKT2/3 and PIK3R1 are potential m6A-related therapeutic targets. | [153] |
| Myocardial infarction (MI) | 5mdC, 5mrC, m6A levels | Heart tissue and blood (rats) | Increased levels of 5mdC, 5mrC, and m6A in heart tissue eight weeks after surgery. | [155] |
| Rheumatoid arthritis | ALKBH5, FTO, YTHDF2, m6A levels | Peripheral blood | Decreased ALKBH5, FTO, and YTHDF2 as risk factors. Global m6A is increased and negatively correlates with decreased FTO gene expression. | [145] |
| Type 2 diabetes (T2DM) | FTO | Peripheral blood | m6A levels are lower in T2DM patients, probably caused by the higher FTO expression. Higher FTO level is associated with T2DM risk. | [147] |
| Systemic lupus erythematosus (SLE) | METTL3, WTAP, FTO, ALKBH5, YTHDF2 | Peripheral blood | Decreased METTL3, WTAP, FTO, ALKBH5, and YTHDF2 gene expression in the blood of SLE patients. ALKBH5 as a risk factor and involved in SLE pathogenesis. | [160] |
| Systemic lupus erythematosus (SLE) | METTL14, ALKBH5, YTHDF2 | PBMCs | Decreased expression of METTL14, ALKBH5, and YTHDF2 in SLE. | [154] |
| Endometriosis | FTO, HNRNPC, HNRNPA2B1 | GSE6364 data | FTO, HNRNPC, and HNRNPA2B1 have biomarker potential. | [161] |
| Lung adenocarcinoma | HNRNPC, METTL3, YTHDC2, KIAA1429, ALKBH5, YTHDF1 | Tissue | HNRNPC, METTL3, YTHDC2, KIAA1429, ALKBH5, and YTHDF1 linked to clinical features, pathological stages, gender, and survival. | [158] |
| Lung adenocarcinoma | HNRNPC | Tissue | HNRNPC high expression correlates with gender, age, ethnicity, lymph node metastasis, smoking history, TNM staging, and poor prognosis. | [156] |
| Lung adenocarcinoma | IGF2BP3 | Tissue | IGF2BP3 correlates with poor prognosis, tumour length, differentiation, T stage, and gender. IGF2BP3 is an independent prognosis factor and potential oncogene. | [157] |
| Lung adenocarcinoma | KIAA1429, RBM15, METTL3, HNRNPC, HNRNPA2B1, YTHDF1, YTHDF2 | Tissue | METTL3, YTHDF1, and YTHDF2 are prognostic biomarkers and suggest better OS and RFS. | [159] |
| Lung adenocarcinoma | METL3, VIRMA, RBM15, YTHDF1, YTHDF2, LRPPRC, HNRNPA2B, IGFBP3, RBMX, FTO, ALKBH5, WTAP, METTL16, METTL14, ZC3H13 | Tissue | Risk factors that predict worse prognosis. | [162] |
| Non-small-cell lung carcinoma (NSCLC) | m6A levels, METTL3, METTL14, RBM15, ALKBH5, FTO | Peripheral blood | Leukocyte m6A levels potential biomarker for NSCLC screening, diagnosis, and monitoring. | [142] |
| Non-small cell lung cancer (NSCLC) | YTHDC2, METTL3, RBM15, HNRNPC, YTHDF2, YTHDF1, ZC3H13 | Tissue | Gene signatures classify prognostic groups. | [163] |
| Non-small cell lung cancer (NSCLC) | HNRNPC | Tissue | HNRNPC predicts poor prognosis and correlates with lymph node metastasis and tumour invasion. | [164] |
| Lung squamous cell carcinoma | ALKBH5, METTL3, HNRNPC, KIAA1429 | Tissue | T follicular helper cells have a prognostic signature role in predicting the survival and treatment response. | [165] |
| Breast cancer | METTL14, FTO, m6A levels | Peripheral blood | m6A levels are elevated in advanced tumour stages. | [143] |
| Hepatocellular carcinoma (HCC) | ALKBH5 | Tissue and cells | Loss of ALKBH5 is an independent prognostic factor. ALKBH5 inhibits HCC proliferation in vitro and in vivo. | [166] |
| Hepatocellular carcinoma (HCC) | YTHDF1 | Tissue | YTHDF1 is upregulated in HCC and correlates with stage. Lower YTHDF1 expression results in better OS. YTHDF1 is involved in HCC cell cycle progression and metabolism regulation. | [167] |
| Hepatocellular carcinoma (HC) | YTHDF1, YTHDF2, METTL3, KIAA1429 | Tissue | m6A regulators differentially expressed in HC. Independent prognostic risk signature: YTHDF1, YTHDF2, METTL3, and KIAA1429. | [168] |
| Hepatocellular carcinoma (HCC) | METTL14 | Tissue | METTL14 expression correlates with the expression and regulates m6A methylation of hub genes, CSAD, GOT2, and SOCS2. | [169] |
| Hepatocellular carcinoma (HCC) | m5C-related lncRNAs | Tissue | Prognosis value established for 8 m5C-related lncRNAs. | [170] |
| Gastric cancer (GC) and benign gastric disease (BGD) | m6A levels, ALKBH5, FTO | Peripheral blood | m6A levels are elevated in advanced tumour stages. m6A decreases after surgery. FTO in stage IV disease < stage I. | [144] |
| Gastric cancer | m6A levels, YTHDF1 | Tissue and cell | Constructed a diagnostic m6A score that can distinguish cancer from normal tissue. YTHDF1 expression correlates with high-risk subtype patients, and it is a possible oncogene. | [171] |
| Gastric cancer (GC) | m6A levels | Tissue | m6A score is an independent prognostic biomarker. Lower m6A scores have EBV and MSI patients that are sensitive to checkpoint immunotherapy. Negative correlation between m6A score and mutation. EMT has the lowest m6A score. | [172] |
| Gastric cancer (GC) | m6A levels, METTL3 | Tissue | High METTL3 expression increases m6A levels and is associated with GC proliferation, liver metastasis, and poor prognosis. | [173] |
| Gastric cancer (GC) | FTO, ALKBH1 | Tissue | High expression of FTO and ALKBH1 transcripts associated with low survival. Low ALKBH1 protein expression correlates with larger tumour size and advanced TNM stages. Low FTO protein expression correlates with shorter OS. | [174] |
| Gastric cancer (GC) | RBM15, WTAP, METTL3, YTHDF2, YTHDF1, YTHDC1, YTHDC2, KIAA1429, ZC3H13, HNRNPC | Tissue | Hub genes associated with m6A regulators have prognostic values: AARD, ASPN, SLAMF9, MIR3117, and DUSP1. ASPN is also upregulated in GC cells. | [175] |
| Pancreatic cancer | KIAA1429, HNRNPC, METTL3, YTHDF1, IGF2BP2, IGF2BP3 | Tissue, cell line | m6A-regulator risk signature. | [176] |
| Pancreatic adenocarcinoma | RBM15 | Tissue | Various significant prognostic parameters. | [177] |
| Colonic adenocarcinoma | YTHDF1, METTL3, KIAA1429, YTHDF3, YTHDC2, METTL14, ALKBH5 | Tissue | YTHDF1, YTHDF3, and YTHDC2 are promising biomarkers for detection, progression, and prognosis. | [178] |
| Colon cancer | ALKBH5 | Tissue, cells | ALKBH5 has a tumour suppressor role in CC. Overexpression of ALKBH5 can inhibit CC invasion and metastasis and has prognostic significance. | [179] |
| Colorectal cancer (CRC) | m6A levels, IGF2BP2 | PBMCs, GEO data | m6A in the blood is a prospective biomarker for CRC and a possible therapeutic target. IGF2BP2 has high expression in CRC blood. Monocytes have the most m6A modification. | [152] |
| Colorectal adenocarcinoma | METTL3, YTHDF1, IGF2BP1, IGF2BP3, EIF3B, HNRNPA2B, YTHDF1, IGF2BP1, IGF2BP3 | Tissue | Potential biomarkers YTHDF1, IGF2BP1, IGF2BP3, and EIF3B. | |
| Acute myeloid leukaemia | WTAP | Peripheral blood, bone marrow cells | Patients were classified into two risk groups based on WTAP expression. High WTAP more common in older patients. | [151] |
| Acute myeloid leukaemia | ZC3H13, RBM15, LRPPRC, METTL14, YTHDC2 | Tissue | METTL14, YTHDC2, ZC3H13, and RBM15 expression correlates with OS. | [180] |
| Neuroblastoma | METT14, WTAP, HNRNPC, YTHDF1, IGF2BP2 | Tissue | Risk prediction signature: METT14, WTAP, HNRNPC, YTHDF1, and IGF2BP2. | [181] |
| Head and neck squamous cell carcinoma. | YTHDC2 | Tissue | YTHDC2 correlates with prognosis and immune infiltration level (CD4+ T cell subpopulation). YTHDC2 has a possible tumour suppressor role. | [182] |
| Head and neck squamous cell carcinoma | IGF2BP2 | Tissue | IGF2BP2 was identified as a hub m6A regulator, and its high expressions are correlated with poor prognosis. | [183] |
| Melanoma | ALKBH5, YTHDF1, KIAA1429 | Tissue | Prognostic risk signature: ALKBH5, YTHDF1, and KIAA1429 divides patients into high- and low-risk OS groups. | [184] |
| Melanoma | YTHDF1, HNRNPA2B1 | Tissue | Tumour stage and treatment response differ between patients with/without mutations in m6A regulatory genes. | [185] |
| Uveal melanoma | RBM15B, IGF2BP1, IGF2BP2, YTHDF3, YTHDF1 | Tissue | m6A regulators with prognostic value: RBM15B, IGF2BP1, IGF2BP2, YTHDF3, and YTHDF1. RBM15B is an independent prognostic factor and correlates with clinicopathologic characteristics. | [186] |
| Osteosarcoma | KIAA1429, HNRNPA2B1 METTL3, YTHDF3, METTL14, FTO, YTHDF2 | Tissue | Prognostic signatures. | [187] |
| Papillary thyroid carcinoma | HNRNPC, WTAP, RBM15, YTHDC2, YTHDC1, FTO, METTL14, METTL3, ALKBH5, KIAA1429, YTHDF1, ZC3H13 | Tissue | Prognostic signature RBM15, KIAA1429, FTO. | [188] |
| Endocrine system tumours | IGF2BP1, METTL14, RBMX, HNRNPC, IGF2BP3, HNRNPA2B1 ICBLL1, RBM15B, KIAA1429, WTAP | Tissue | Prognostic signatures. | [189] |
| Adrenocortical carcinoma | RBM15, ZC3H3, YTDHF1, YTDHF2, ALBH5, KIAA1429, YTHDC1, HNRNPC, WTAP, METTL3, FTO | Tissue | Independent prognostic risk signature: HNRNPC, RBM15, METTL14, and FTO. | [190] |
| Clear cell renal cell carcinoma | METTL3, METTL14 | Tissue | METTL3 and METTL14 are associated with prognosis and clinicopathological features. | [191] |
| Clear cell renal cell carcinoma | FTO, IGF2BP2, IGF2BP3, KIAA1429, YTHDC1, ZC3H13 | Tissue | m6A-related risk signature for prognosis. m6A regulators’ expression correlates with histological grade and staging. | [192] |
| Clear cell renal cell carcinoma | ALKBH5, FTO | Tissue | ALKBH5 and FTO decreased gene expression correlates with poor OS. | [193] |
| Clear cell renal cell carcinoma | METTL14 | Tissue | METTL14 probably methylates m6A in PTEN, leading to its expression change. METTL14 gene expression negatively correlates with the tumour stages and positively correlates with KIRC patients’ OS. | [194] |
| Bladder cancer | HNRNPA2B1 IGF2BP1, IGF2BP3, METTL3, YTHDF2, YTHDF1, FTO, ZC3H13, YTHDF3, YTHDC1, WTAP, METTL16, METTL14 | Tissue | Identified risk factors that correlate with advanced clinical stages: RBM15, HNRNPA2B1, HNRNPC, IGF2BP2, YTHDF1, and YTHDF2. | [195] |
| Bladder cancer | METTL3, WTAP, FTO, YTHDC1 | Tissue | Independent prognostic signature and predictor of clinicopathology. | [196] |
| m6A Regulators | GBM vs. NT | GBM vs. AST | GBM vs. LGG | High vs. Low Grade | Refs. |
|---|---|---|---|---|---|
| FTO | ↓ | ↓ | ↓ | ↓ | [100,105,107] |
| METTL3 | ↑ | ↓ | ↓ | [100,101,107,205] | |
| RBM15 | ↑ | [100,105,107] | |||
| RBM15B | ↑ | [107] | |||
| ZC3H13 | ↓ | ↓ | ↓ | [100,107] | |
| KIAA1429 (VIRMA) | ↓ | [107] | |||
| eIF3A | ↓ | ↓ | [101,202] | ||
| eIF3B | ↑ | ↑ | ↑ | [202] | |
| eIF3E | ↑ | [202] | |||
| eIF3I | ↑ | ↑ | ↑ | [202] | |
| eIF3K | ↑ | ↑ | ↑ | [202] | |
| eIF3L | ↓ | ↓ | [202] | ||
| eIF3M | ↑ | ↑ | ↑ | [202] | |
| FMR1 | ↓ | [101] | |||
| HNRPC | ↑ | ↓ | [100,107,213] | ||
| HNRNPA2B1 | ↑ | [107] | |||
| IGF2BP2 | ↑ | [101] | |||
| IGF2BP3 | ↑ | ↑ | ↑ | [101,212] | |
| RBMX | ↓ | [101] | |||
| YTHDF1 | ↑ | ↑ | [100,101,107,213] | ||
| YTHDF2 | ↑ | [100,107,213] | |||
| YTHDF3 | ↑ | ↓ | [101,107,214] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tűzesi, Á.; Hallal, S.; Satgunaseelan, L.; Buckland, M.E.; Alexander, K.L. Understanding the Epitranscriptome for Avant-Garde Brain Tumour Diagnostics. Cancers 2023, 15, 1232. https://doi.org/10.3390/cancers15041232
Tűzesi Á, Hallal S, Satgunaseelan L, Buckland ME, Alexander KL. Understanding the Epitranscriptome for Avant-Garde Brain Tumour Diagnostics. Cancers. 2023; 15(4):1232. https://doi.org/10.3390/cancers15041232
Chicago/Turabian StyleTűzesi, Ágota, Susannah Hallal, Laveniya Satgunaseelan, Michael E. Buckland, and Kimberley L. Alexander. 2023. "Understanding the Epitranscriptome for Avant-Garde Brain Tumour Diagnostics" Cancers 15, no. 4: 1232. https://doi.org/10.3390/cancers15041232
APA StyleTűzesi, Á., Hallal, S., Satgunaseelan, L., Buckland, M. E., & Alexander, K. L. (2023). Understanding the Epitranscriptome for Avant-Garde Brain Tumour Diagnostics. Cancers, 15(4), 1232. https://doi.org/10.3390/cancers15041232