
F-Box Protein 43, Stabilized by N6-Methyladenosine Methylation, Enhances Hepatocellular Carcinoma Cell Growth and Invasion via Promoting p53 Degradation in a Ubiquitin Conjugating Enzyme E2 C-Dependent Manner


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines, Small Interfering RNAs (siRNAs), and Plasmids
2.2. Tissue Samples
2.3. Online Analysis
2.4. IHC Assay
2.5. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.6. Methylated RNA Immunoprecipitation Quantitative Polymerase Chain Reaction (MeRIP-qPCT)
2.7. RNA Immunoprecipitation (RIP)
2.8. T7 Biotin Labeled RNA Synthesis and RNA Pulldown
2.9. RNA Stability Assays
2.10. Western Blot (WB)
2.11. Immunoprecipitation (IP)
2.12. Luciferase Reporter Assay
2.13. CCK-8 Assay
2.14. Plate Clone Formation Assay
2.15. Transwell Invasion Assay
2.16. Statistical Analysis
3. Results
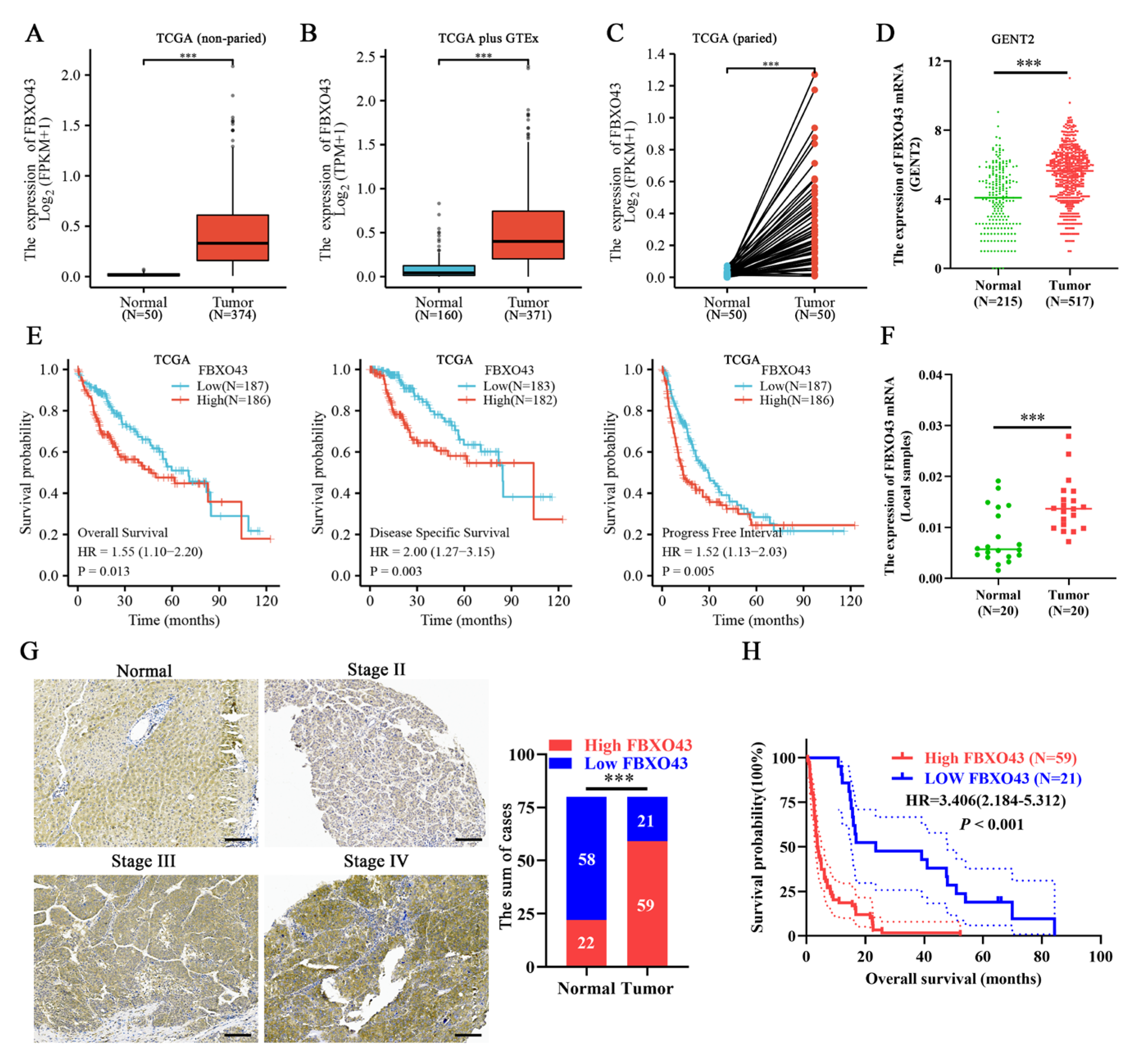
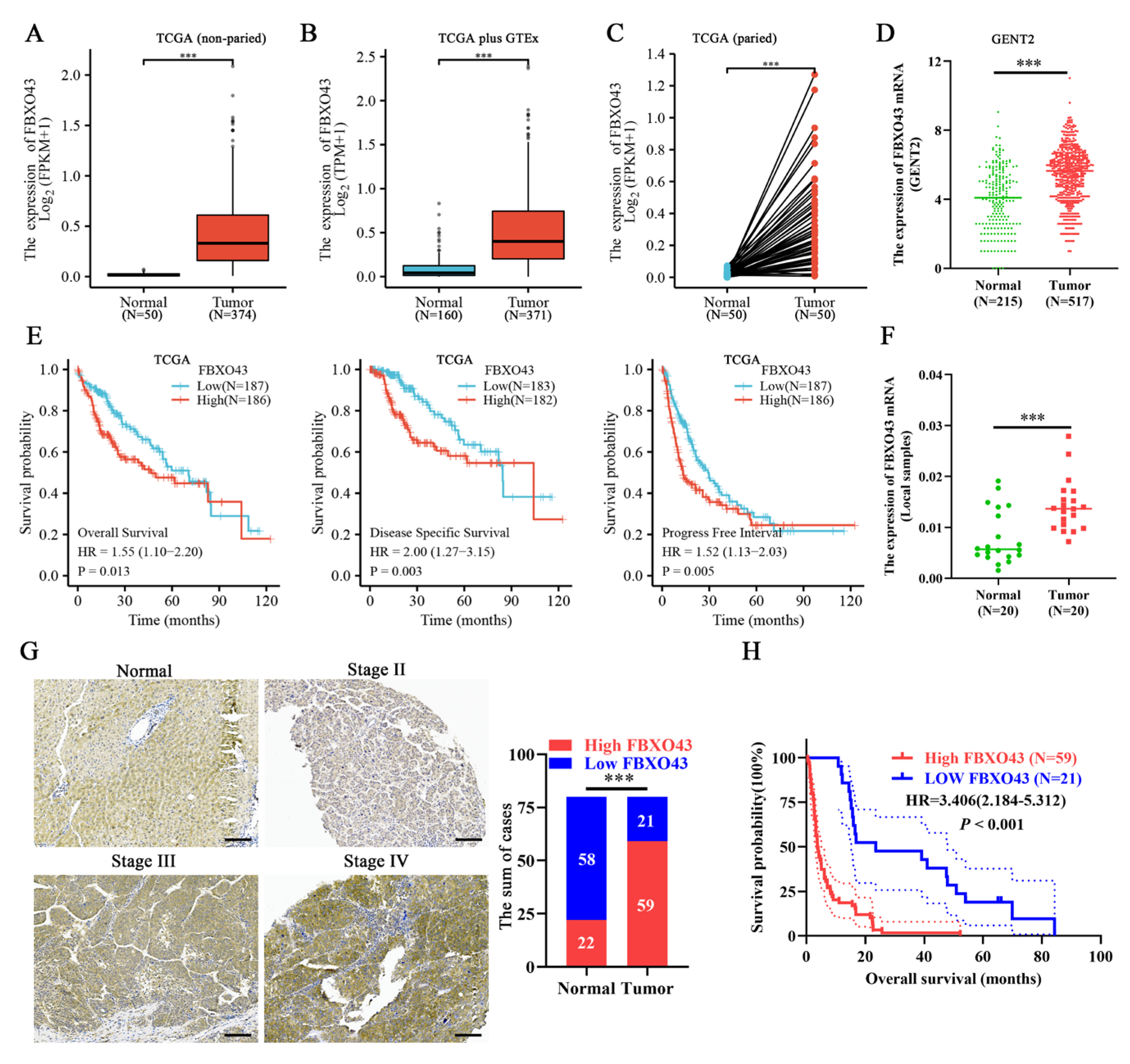
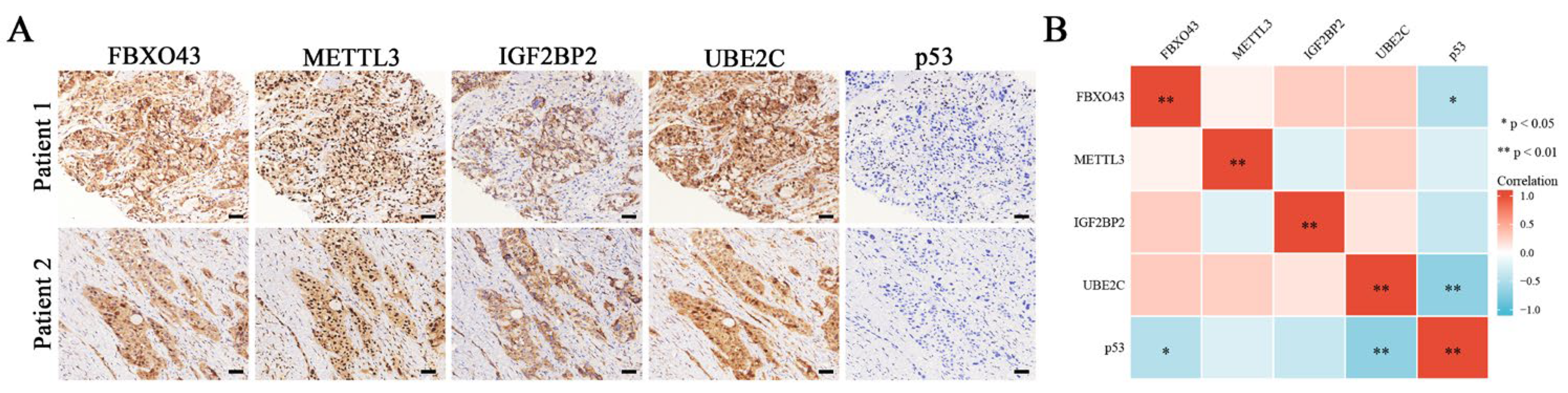
3.1. A High Level of FBXO43 Is Associated with Malignant Progression and Poor Prognosis in HCC
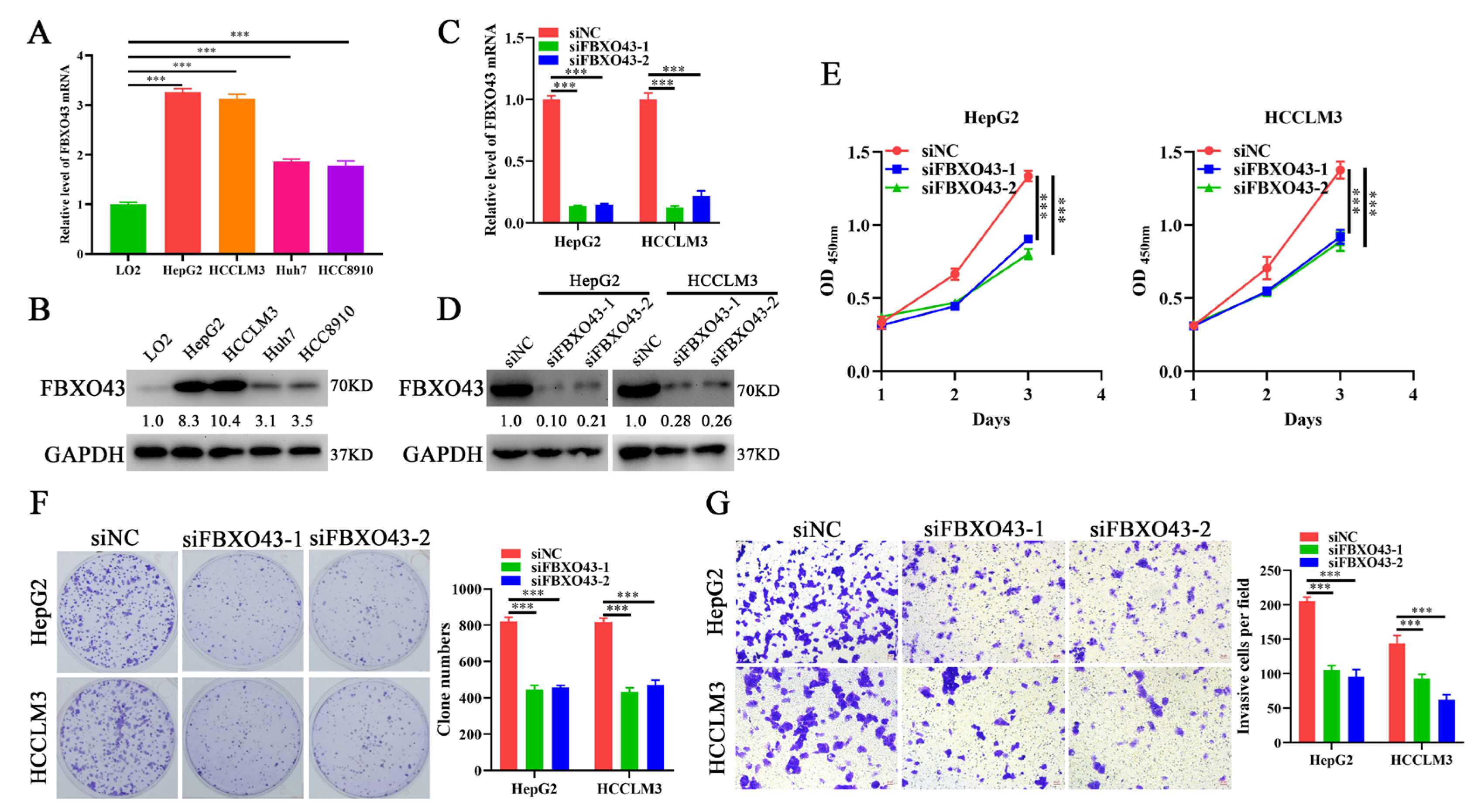
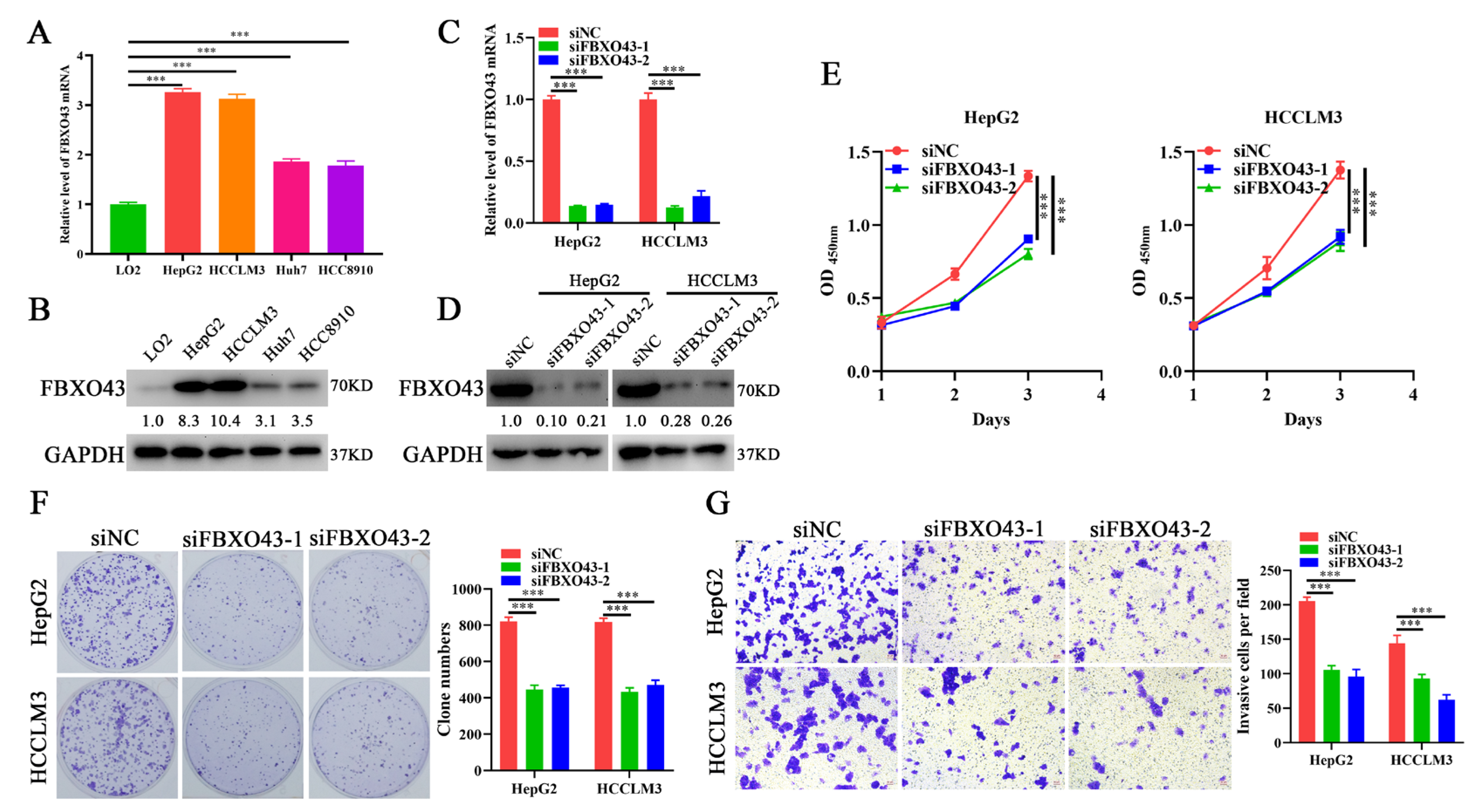
3.2. FBXO43 Knockdown Inhibits the Proliferation and Invasion of HCC Cells
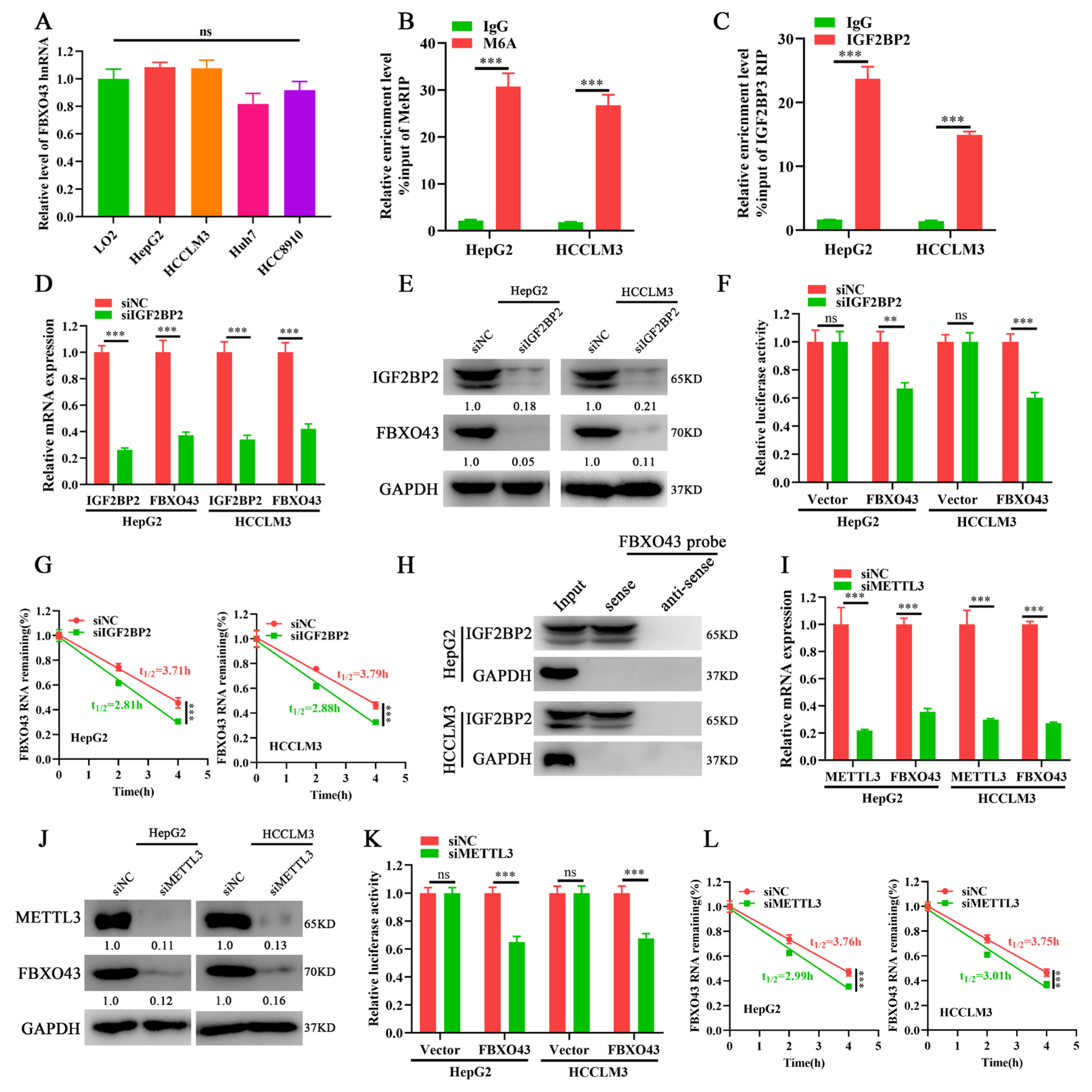
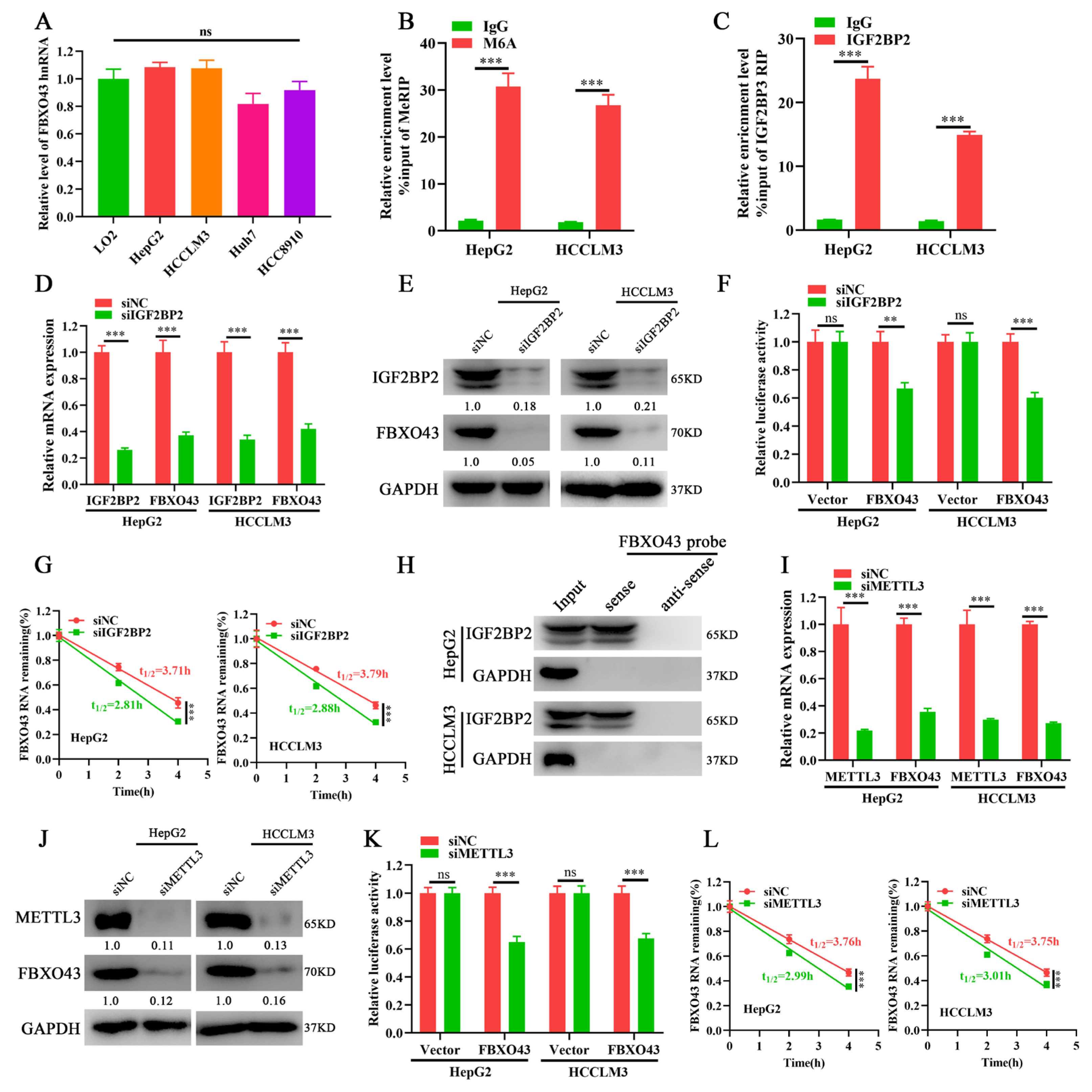
3.3. METTL3/IGF2BP2 Respectively Writes and Recognizes the m6A of FBXO43 mRNA and Enhances Its Stability in HCC
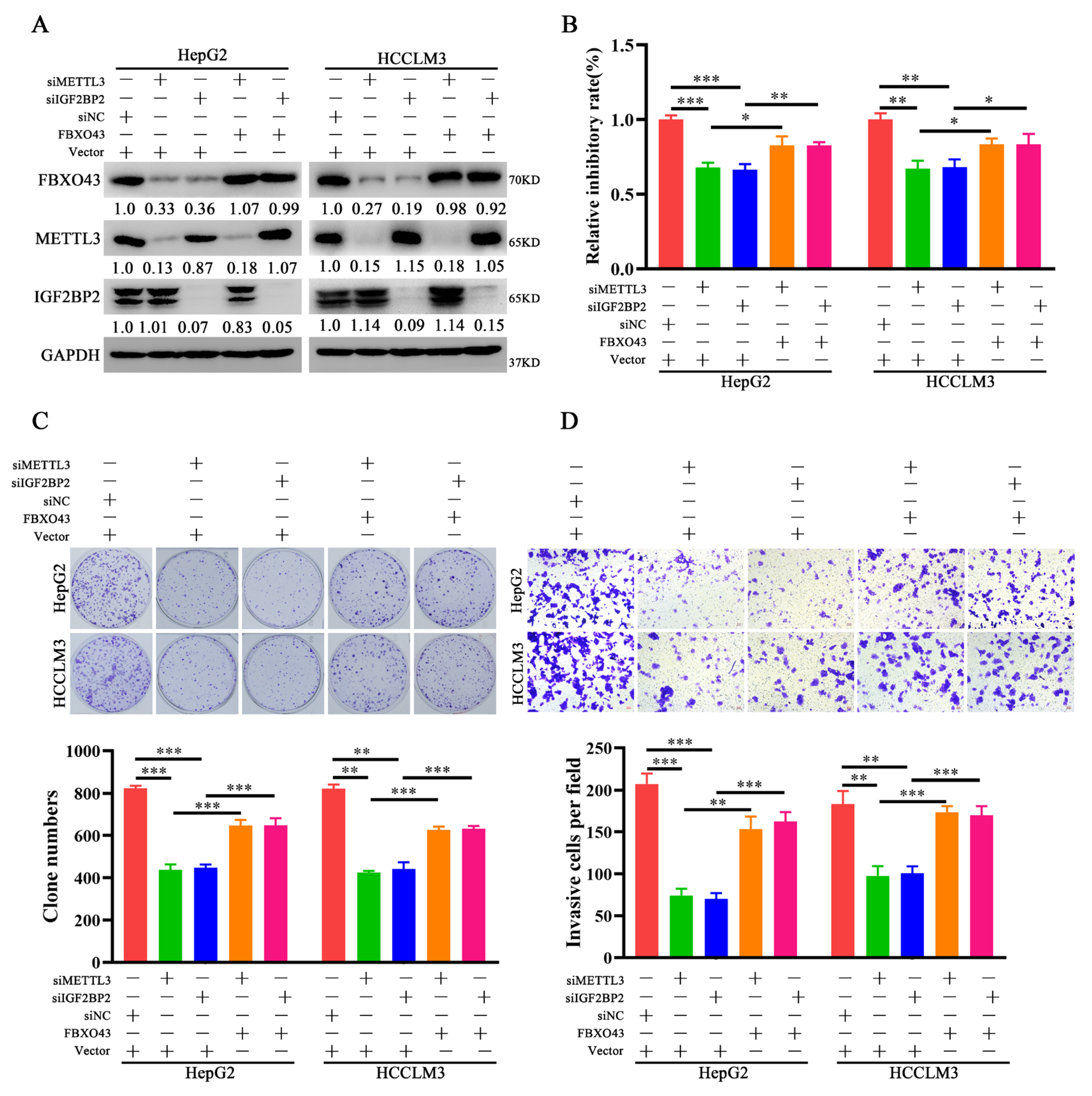
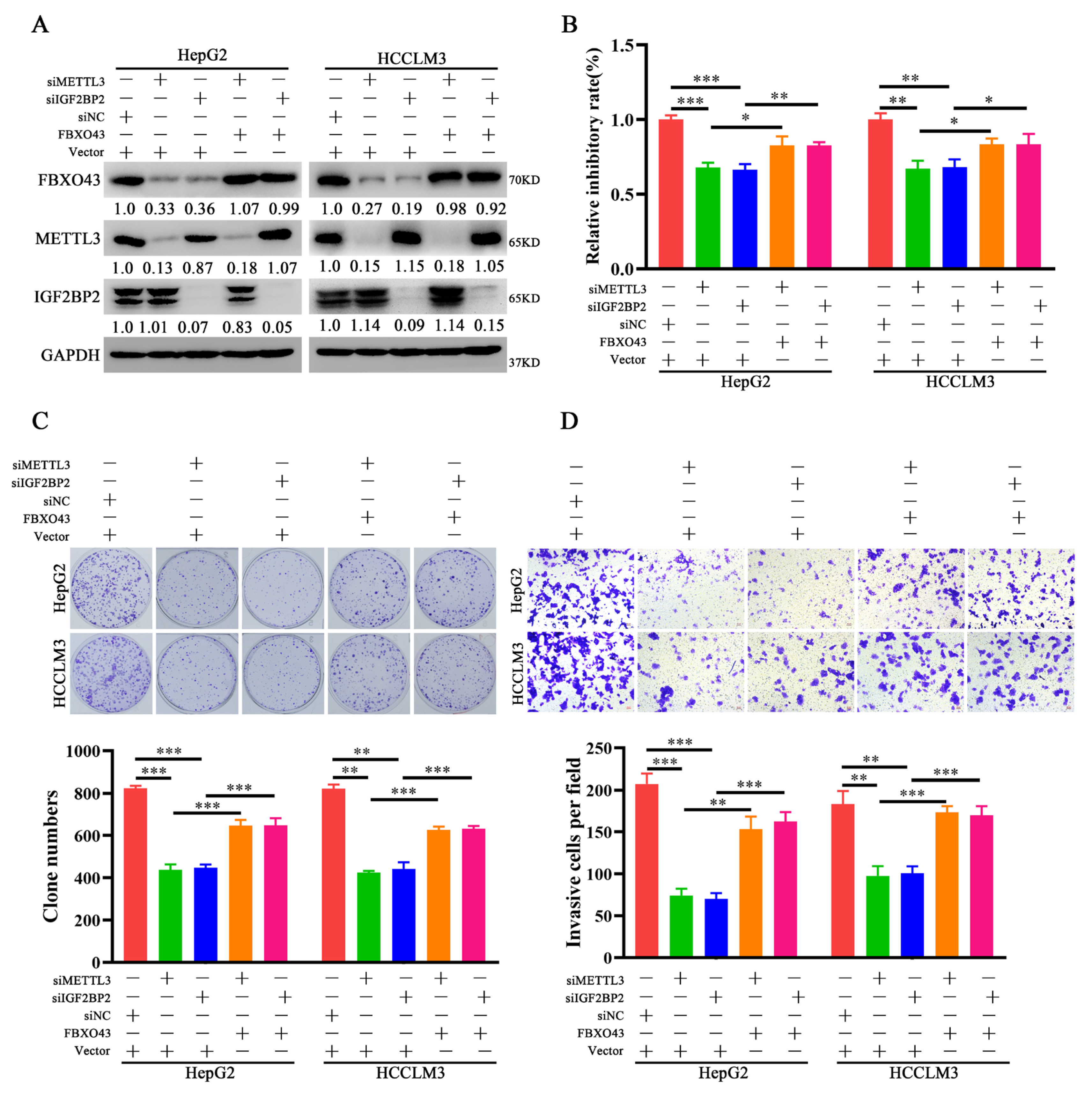
3.4. FBXO43 Is an Effector of the Oncogenic Role of METTL3 and IGF2BP2 in HCC
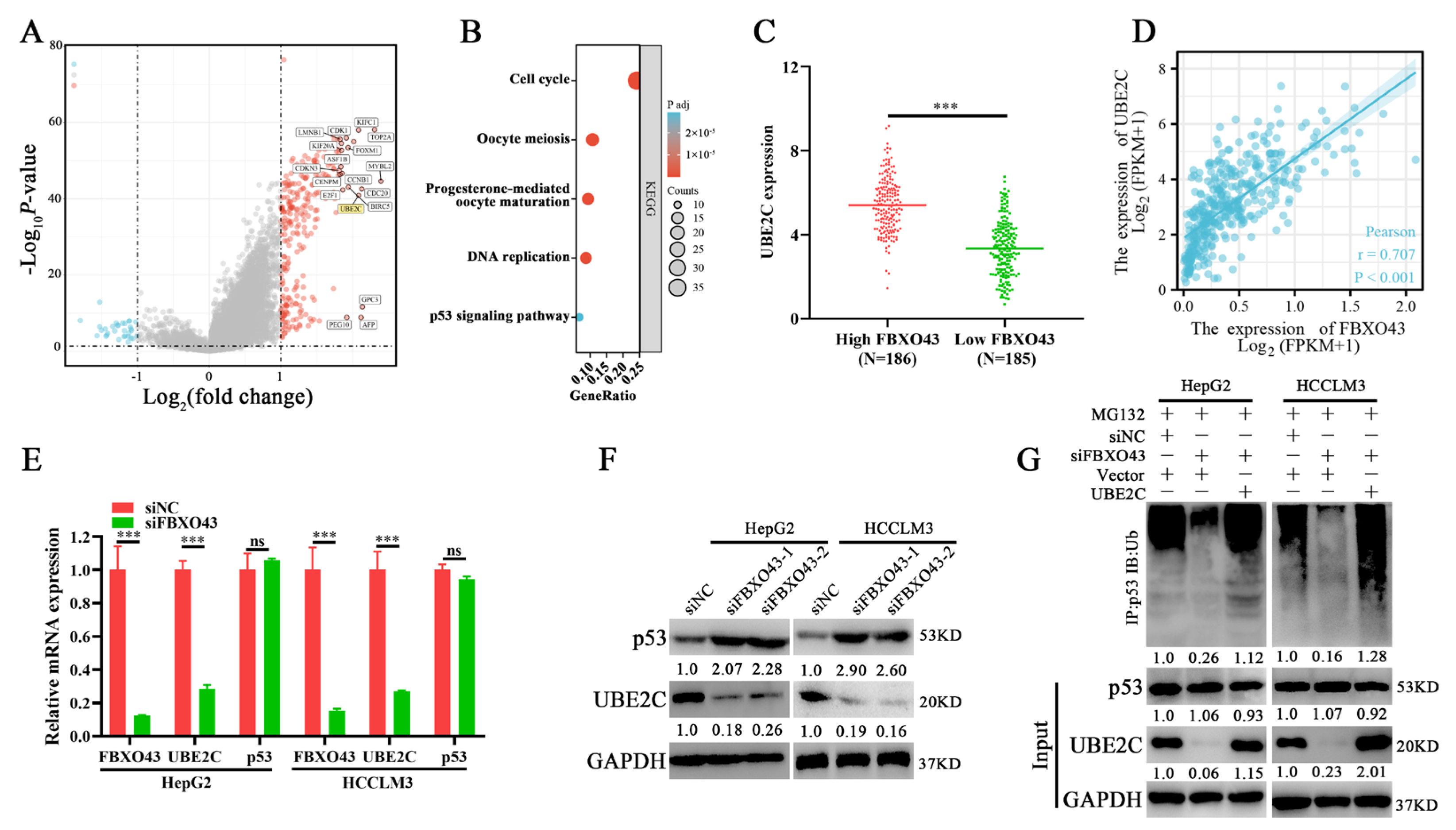
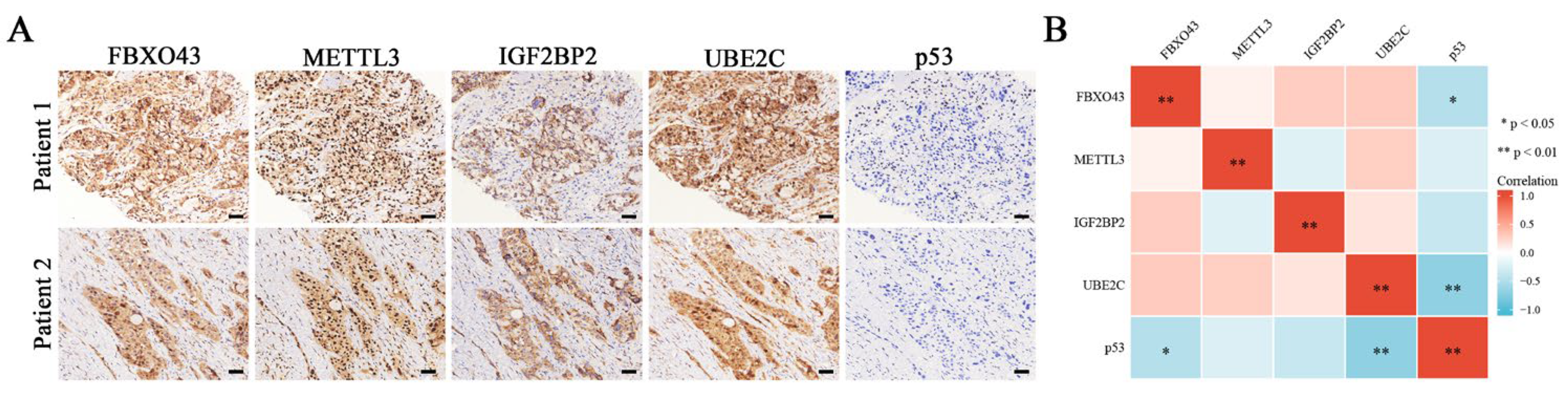
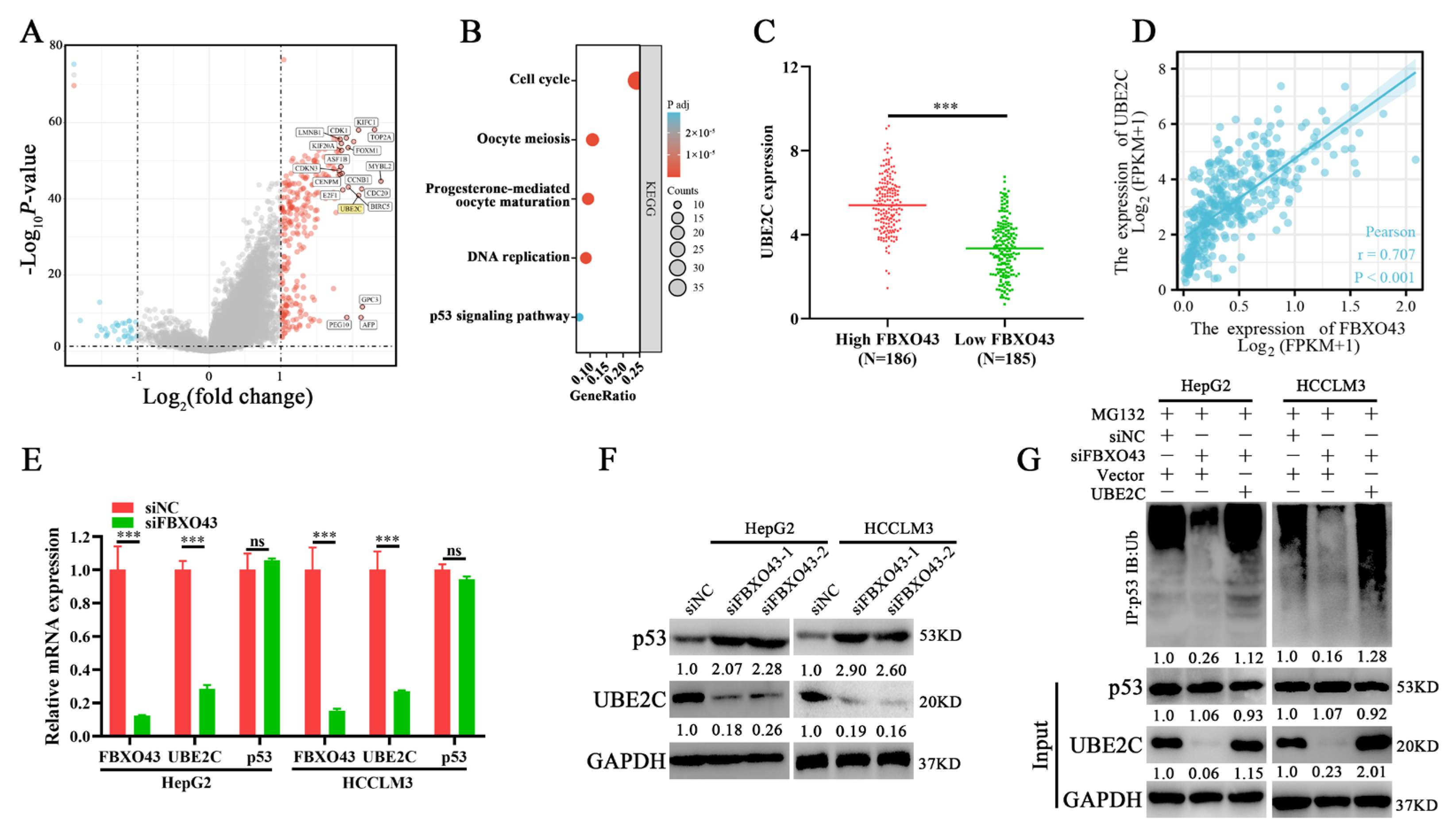
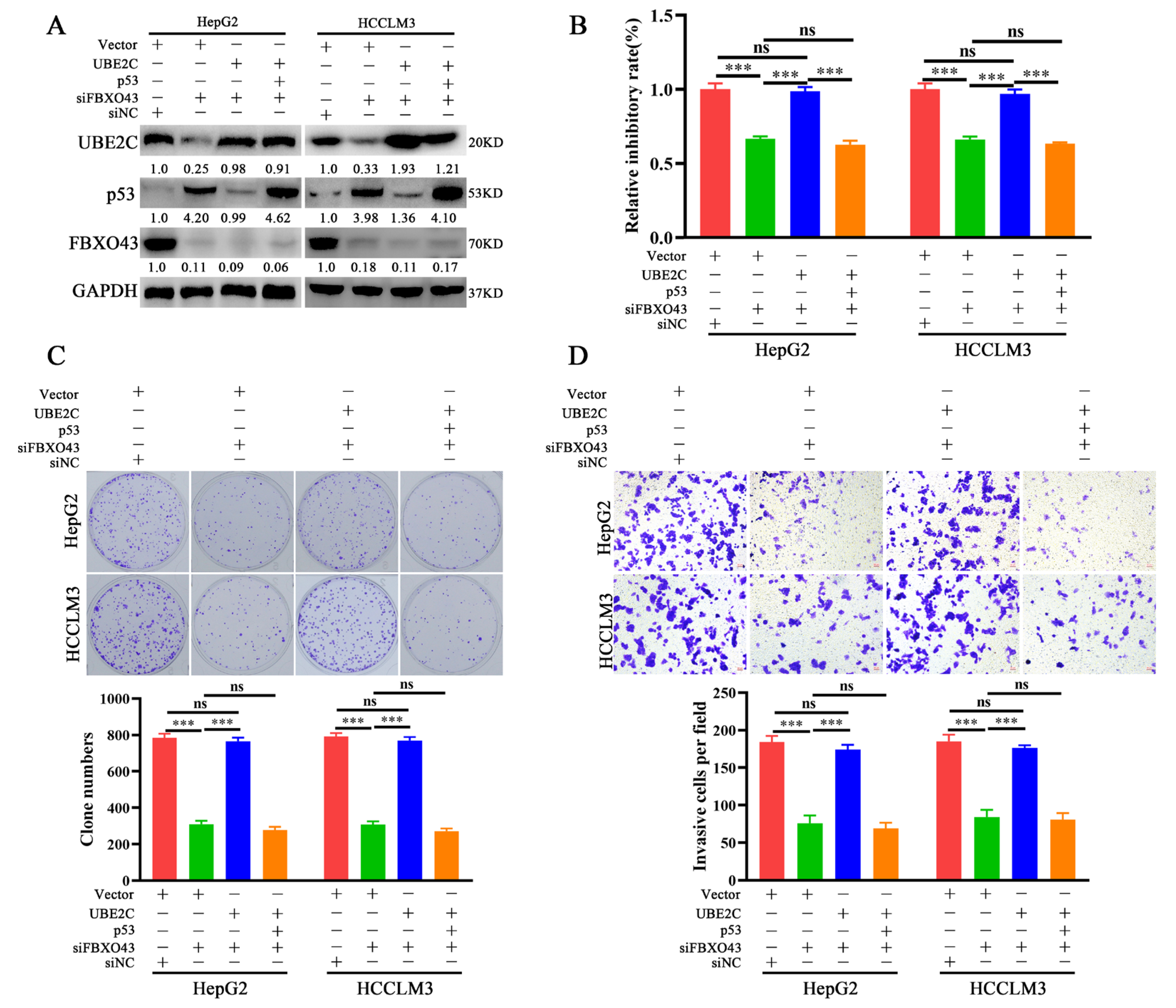
3.5. FBXO43 Promotes p53 Degradation by Maintaining UBE2C Expression in HCC
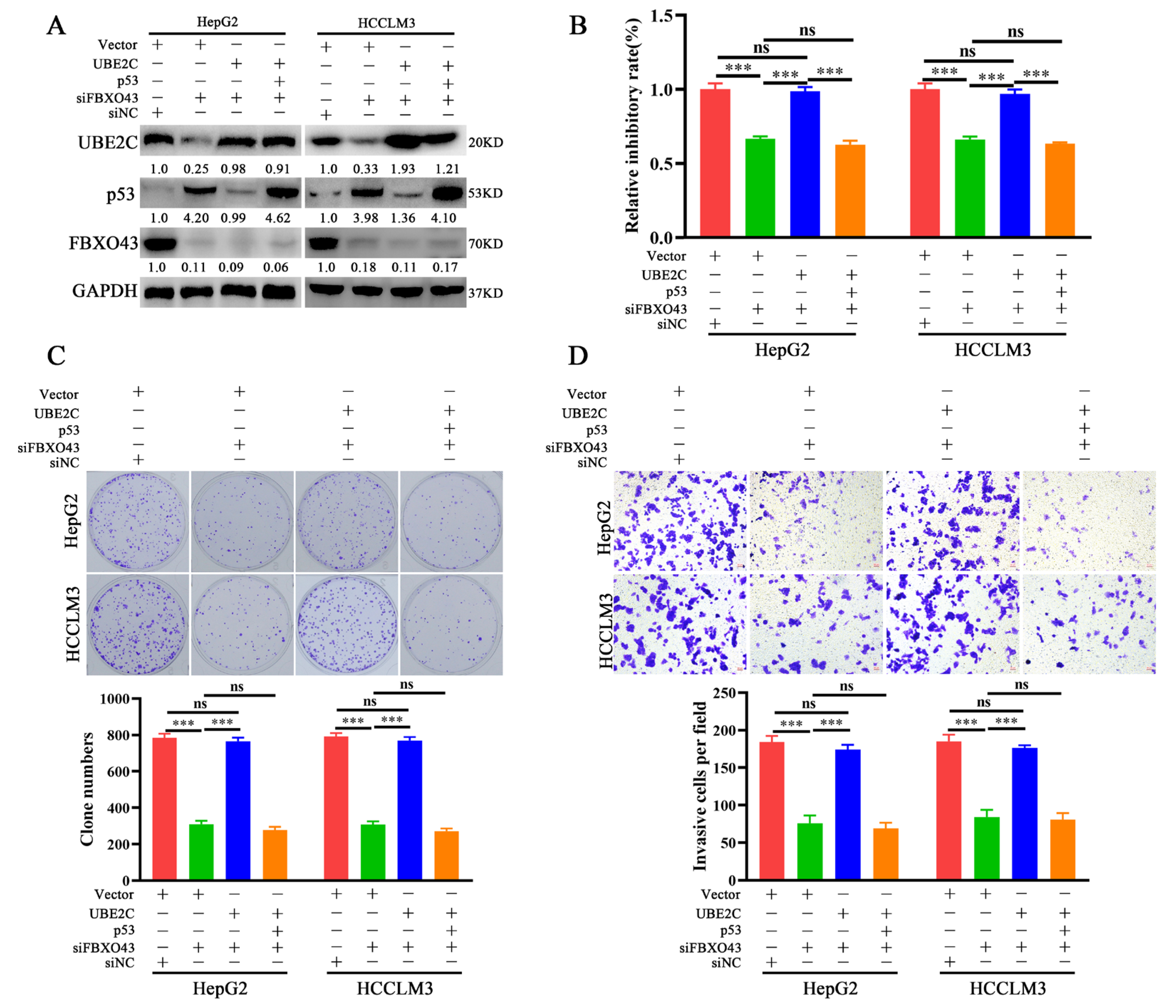
3.6. FBXO43 Exerts Oncogenic Roles by Regulating the UBE2C/p53 Axis in HCC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Villanueva, A. Hepatocellular Carcinoma. N. Engl. J. Med. 2019, 380, 1450–1462. [Google Scholar] [CrossRef]
- Tung, J.J.; Hansen, D.V.; Ban, K.H.; Loktev, A.V.; Summers, M.K.; Adler, J.R., 3rd; Jackson, P.K. A role for the anaphase-promoting complex inhibitor Emi2/XErp1, a homolog of early mitotic inhibitor 1, in cytostatic factor arrest of Xenopus eggs. Proc. Natl. Acad. Sci. USA 2005, 102, 4318–4323. [Google Scholar] [CrossRef] [PubMed]
- Tischer, T.; Hormanseder, E.; Mayer, T.U. The APC/C inhibitor XErp1/Emi2 is essential for Xenopus early embryonic divisions. Science 2012, 338, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chien, Y.H.; Ryan, A.; Kintner, C. Emi2 enables centriole amplification during multiciliated cell differentiation. Sci. Adv. 2022, 8, eabm7538. [Google Scholar] [CrossRef]
- Ma, R.; Zhu, K.; Yuan, D.; Gong, M.; Li, Y.; Li, K.; Meng, L. Downregulation of the FBXO43 gene inhibits tumor growth in human breast cancer by limiting its interaction with PCNA. J. Transl. Med. 2021, 19, 425. [Google Scholar] [CrossRef]
- Vadhan, A.; Wang, Y.Y.; Yuan, S.F.; Lee, Y.C.; Hu, S.C.; Huang, J.Y.; Ishikawa, T.; Hou, M.F. EMI2 expression as a poor prognostic factor in patients with breast cancer. Kaohsiung J. Med. Sci. 2020, 36, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Deng, B.; Zhu, M.; Wang, Y.; Yan, C.; Wang, T.; Liu, Y.; Li, G.; Ding, Y.; Jin, G. Integration of gwas and eQTL Analysis to Identify Risk Loci and Susceptibility Genes for Gastric Cancer. Front. Genet. 2020, 11, 679. [Google Scholar] [CrossRef]
- Xu, B.; Lv, W.; Li, X.; Zhang, L.; Lin, J. Prognostic genes of hepatocellular carcinoma based on gene coexpression network analysis. J. Cell. Biochem. 2019, 120, 11616–11623. [Google Scholar] [CrossRef]
- He, L.; Li, H.; Wu, A.; Peng, Y.; Shu, G.; Yin, G. Functions of N6-methyladenosine and its role in cancer. Mol. Cancer 2019, 18, 176. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, B.; Nie, Z.; Duan, L.; Xiong, Q.; Jin, Z.; Yang, C.; Chen, Y. The role of m6A modification in the biological functions and diseases. Signal Transduct. Target. Ther. 2021, 6, 74. [Google Scholar] [CrossRef]
- Zhou, T.; Cai, Z.; Ma, N.; Xie, W.; Gao, C.; Huang, M.; Bai, Y.; Ni, Y.; Tang, Y. A Novel Ten-Gene Signature Predicting Prognosis in Hepatocellular Carcinoma. Front. Cell Dev. Biol. 2020, 8, 629. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Yoon, B.H.; Kim, S.; Kim, S.Y. GENT2: An updated gene expression database for normal and tumor tissues. BMC Med. Genom. 2019, 12, 101. [Google Scholar] [CrossRef]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.; Yang, J.H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic. Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef]
- Menyhart, O.; Nagy, A.; Gyorffy, B. Determining consistent prognostic biomarkers of overall survival and vascular invasion in hepatocellular carcinoma. R. Soc. Open Sci. 2018, 5, 181006. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Hu, Z.; Liu, J.; Huang, W. FLOT2 upregulation promotes growth and invasion by interacting and stabilizing EphA2 in gliomas. Biochem. Biophys. Res. Commun. 2021, 548, 67–73. [Google Scholar] [CrossRef]
- Li, Z.X.; Zheng, Z.; Yang, P.Y.; Lin, L.; Zhou, G.Q.; Lv, J.W.; Zhang, L.L.; Chen, F.; Li, Y.Q.; Wu, C.F.; et al. WTAP-mediated m(6)A modification of lncRNA DIAPH1-AS1 enhances its stability to facilitate nasopharyngeal carcinoma growth and metastasis. Cell Death Differ. 2022, 29, 1137–1151. [Google Scholar] [CrossRef]
- Huang, W.; Zeng, C.; Hu, S.; Wang, L.; Liu, J. ATG3, a Target of miR-431-5p, Promotes Proliferation and Invasion of Colon Cancer via Promoting Autophagy. Cancer Manag. Res. 2019, 11, 10275–10285. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, P.; Li, Y.H.; Zhang, Z.; Cui, Q. SRAMP: Prediction of mammalian N6-methyladenosine (m6A) sites based on sequence-derived features. Nucleic. Acids Res. 2016, 44, e91. [Google Scholar] [CrossRef]
- Zhu, M.; Wu, M.; Bian, S.; Song, Q.; Xiao, M.; Huang, H.; You, L.; Zhang, J.; Zhang, J.; Cheng, C.; et al. DNA primase subunit 1 deteriorated progression of hepatocellular carcinoma by activating AKT/mTOR signaling and UBE2C-mediated P53 ubiquitination. Cell Biosci. 2021, 11, 42. [Google Scholar] [CrossRef]
- Liu, L.P.; Yang, M.; Peng, Q.Z.; Li, M.Y.; Zhang, Y.S.; Guo, Y.; Chen, Y.; Bao, S. UBE2T promotes hepatocellular carcinoma cell growth via ubiquitination of p53. Biochem. Biophys. Res. Commun. 2017, 493, 20–27. [Google Scholar] [CrossRef]
- Pan, Y.H.; Yang, M.; Liu, L.P.; Wu, D.C.; Li, M.Y.; Su, S.G. UBE2S enhances the ubiquitination of p53 and exerts oncogenic activities in hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2018, 503, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.P.; Schwank, J.; Staib, F.; Wang, X.W.; Harris, C.C. TP53 mutations and hepatocellular carcinoma: Insights into the etiology and pathogenesis of liver cancer. Oncogene 2007, 26, 2166–2176. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.K.; Midkiff, B.R.; Israelow, B.; Evans, M.J.; Lanford, R.E.; Walker, C.M.; Lemon, S.M.; McGivern, D.R.; Hepatitis, C. Virus Indirectly Disrupts DNA Damage-Induced p53 Responses by Activating Protein Kinase R. mBio 2017, 8, e00121-17. [Google Scholar] [CrossRef]
- Li, Q.G.; Xiao, T.; Zhu, W.; Yu, Z.Z.; Huang, X.P.; Yi, H.; Lu, S.S.; Tang, Y.Y.; Huang, W.; Xiao, Z.Q. HDAC7 promotes the oncogenicity of nasopharyngeal carcinoma cells by miR-4465-EphA2 signaling axis. Cell Death Dis. 2020, 11, 322. [Google Scholar] [CrossRef]
- Qu, N.; Bo, X.; Li, B.; Ma, L.; Wang, F.; Zheng, Q.; Xiao, X.; Huang, F.; Shi, Y.; Zhang, X. Role of N6-Methyladenosine (m(6)A) Methylation Regulators in Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 755206. [Google Scholar] [CrossRef]
- Wang, Y.F.; Ge, C.M.; Yin, H.Z.; Dai, Z.; Dong, J.P.; Ji, M.; Yang, F.M. Dysregulated N6-methyladenosine (m(6)A) processing in hepatocellular carcinoma. Ann. Hepatol. 2021, 25, 100538. [Google Scholar] [CrossRef]
- Chen, M.; Wei, L.; Law, C.T.; Tsang, F.H.; Shen, J.; Cheng, C.L.; Tsang, L.H.; Ho, D.W.; Chiu, D.; Lee, J.M.; et al. RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology 2018, 67, 2254–2270. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.Z.; Liu, Y.; Li, J.; Chen, M.T.; Huang, M.; Wang, F.; Yang, Q.S.; Yuan, J.H.; Sun, S.H. METTL16 promotes hepatocellular carcinoma progression through downregulating RAB11B-AS1 in an m(6)A-dependent manner. Cell Mol. Biol. Lett. 2022, 27, 41. [Google Scholar] [CrossRef]
- Pu, J.; Wang, J.; Qin, Z.; Wang, A.; Zhang, Y.; Wu, X.; Wu, Y.; Li, W.; Xu, Z.; Lu, Y.; et al. IGF2BP2 Promotes Liver Cancer Growth Through an m6A-FEN1-Dependent Mechanism. Front. Oncol. 2020, 10, 578816. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, J.; Dang, Z.; Su, Q.; Yang, J. Circ_0000775 promotes the migration, invasion and EMT of hepatic carcinoma cells by recruiting IGF2BP2 to stabilize CDC27. Pathol. Res. Pract. 2022, 235, 153908. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.; et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Swiatkowska, A.; Dutkiewicz, M.; Zydowicz-Machtel, P.; Szpotkowska, J.; Janecki, D.M.; Ciesiolka, J. Translational Control in p53 Expression: The Role of 5′-Terminal Region of p53 mRNA. Int. J. Mol. Sci. 2019, 20, 5382. [Google Scholar] [CrossRef]
- Chao, C.C. Mechanisms of p53 degradation. Clin. Chim. Acta 2015, 438, 139–147. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, W.; Yan, H.; Hu, Y.; He, Q.; Luo, P. Regulation of p53 stability as a therapeutic strategy for cancer. Biochem. Pharmacol. 2021, 185, 114407. [Google Scholar] [CrossRef] [PubMed]
- Wade, M.; Li, Y.C.; Wahl, G.M. MDM2, MDMX and p53 in oncogenesis and cancer therapy. Nat. Rev. Cancer 2013, 13, 83–96. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, C.; Wang, X.; Ly, P.; Belyi, V.; Xu-Monette, Z.Y.; Young, K.H.; Hu, W.; Feng, Z. E3 ubiquitin ligase TRIM32 negatively regulates tumor suppressor p53 to promote tumorigenesis. Cell Death Differ. 2014, 21, 1792–1804. [Google Scholar] [CrossRef]
- Li, X.L.; Niu, Z.; Sun, C.; Zhuo, S.; Yang, H.; Yang, X.; Liu, Y.; Yan, C.; Li, Z.; Cao, Q.; et al. Regulation of P53 signaling in breast cancer by the E3 ubiquitin ligase RNF187. Cell Death Dis. 2022, 13, 149. [Google Scholar] [CrossRef]
- Nicolau-Neto, P.; Palumbo, A.; De Martino, M.; Esposito, F.; de Almeida Simao, T.; Fusco, A.; Nasciutti, L.E.; Da Costa, N.M.; Pinto, L.R. UBE2C Is a Transcriptional Target of the Cell Cycle Regulator FOXM1. Genes 2018, 9, 188. [Google Scholar] [CrossRef]
- Li, L.; Li, Q. miR-543 impairs breast cancer cell phenotypes by targeting and suppressing ubiquitin-conjugating enzyme E2T (UBE2T). Bioengineered 2021, 12, 12394–12406. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, C.; Peng, D.; He, S.; Huang, C.; Qian, J.; Zhu, W.; Feng, N.; Gong, Y.; Li, X.; et al. MiR-182-5p inhibits the tumorigenesis of clear cell renal cell carcinoma by repressing UBE2T. Hum. Cell 2022, 35, 542–556. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | N | FBOX43 Expression | p Value | |
|---|---|---|---|---|
| High | Low | |||
| Age | ||||
| ≤45 | 38 | 28 | 10 | >0.9999 |
| >45 | 42 | 31 | 11 | |
| Sex | ||||
| Male | 75 | 54 | 21 | 0.3184 |
| Female | 5 | 5 | 0 | |
| AFP (ng/mL) | ||||
| ≤400 | 38 | 20 | 18 | <0.0001 |
| >400 | 42 | 39 | 3 | |
| T stage | ||||
| 1 | 18 | 9 | 9 | 0.0148 |
| 2 + 3 | 62 | 50 | 12 | |
| Hep-1 staining | ||||
| Positive | 61 | 44 | 17 | 0.7665 |
| Negative | 19 | 15 | 4 | |
| Tumor diameter | ||||
| ≤6cm | 37 | 26 | 11 | 0.6127 |
| >6cm | 43 | 33 | 10 | |
| WHO stage | ||||
| II | 11 | 5 | 6 | 0.0317 |
| III-IV | 69 | 54 | 15 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Zeng, C.; Liu, J.; Luo, H.; Huang, W. F-Box Protein 43, Stabilized by N6-Methyladenosine Methylation, Enhances Hepatocellular Carcinoma Cell Growth and Invasion via Promoting p53 Degradation in a Ubiquitin Conjugating Enzyme E2 C-Dependent Manner. Cancers 2023, 15, 957. https://doi.org/10.3390/cancers15030957
Zhou H, Zeng C, Liu J, Luo H, Huang W. F-Box Protein 43, Stabilized by N6-Methyladenosine Methylation, Enhances Hepatocellular Carcinoma Cell Growth and Invasion via Promoting p53 Degradation in a Ubiquitin Conjugating Enzyme E2 C-Dependent Manner. Cancers. 2023; 15(3):957. https://doi.org/10.3390/cancers15030957
Chicago/Turabian StyleZhou, Huijun, Chong Zeng, Jie Liu, Haijun Luo, and Wei Huang. 2023. "F-Box Protein 43, Stabilized by N6-Methyladenosine Methylation, Enhances Hepatocellular Carcinoma Cell Growth and Invasion via Promoting p53 Degradation in a Ubiquitin Conjugating Enzyme E2 C-Dependent Manner" Cancers 15, no. 3: 957. https://doi.org/10.3390/cancers15030957
APA StyleZhou, H., Zeng, C., Liu, J., Luo, H., & Huang, W. (2023). F-Box Protein 43, Stabilized by N6-Methyladenosine Methylation, Enhances Hepatocellular Carcinoma Cell Growth and Invasion via Promoting p53 Degradation in a Ubiquitin Conjugating Enzyme E2 C-Dependent Manner. Cancers, 15(3), 957. https://doi.org/10.3390/cancers15030957