



Exploiting the Stemness and Chemoresistance Transcriptome of Ewing Sarcoma to Identify Candidate Therapeutic Targets and Drug-Repurposing Candidates

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Western Blotting
2.3. Flow Cytometry
2.3.1. Cell Surface Expression of CD133
2.3.2. Co-Expression of ABCG1 and CD133
2.4. Self-Renewing Ability
2.5. Magnetically Activated Cell Sorting for the Enrichment of CD133-Positive Cells
2.6. RNA Expression of Markers of Pluripotency and Differentiation, the Wnt Signalling Pathway and ABC Transporter Proteins
2.7. Immunohistochemistry (IHC)
2.8. siRNA Knockdown of ABCG1
2.9. Apoptosis
2.10. Statistical Analyses
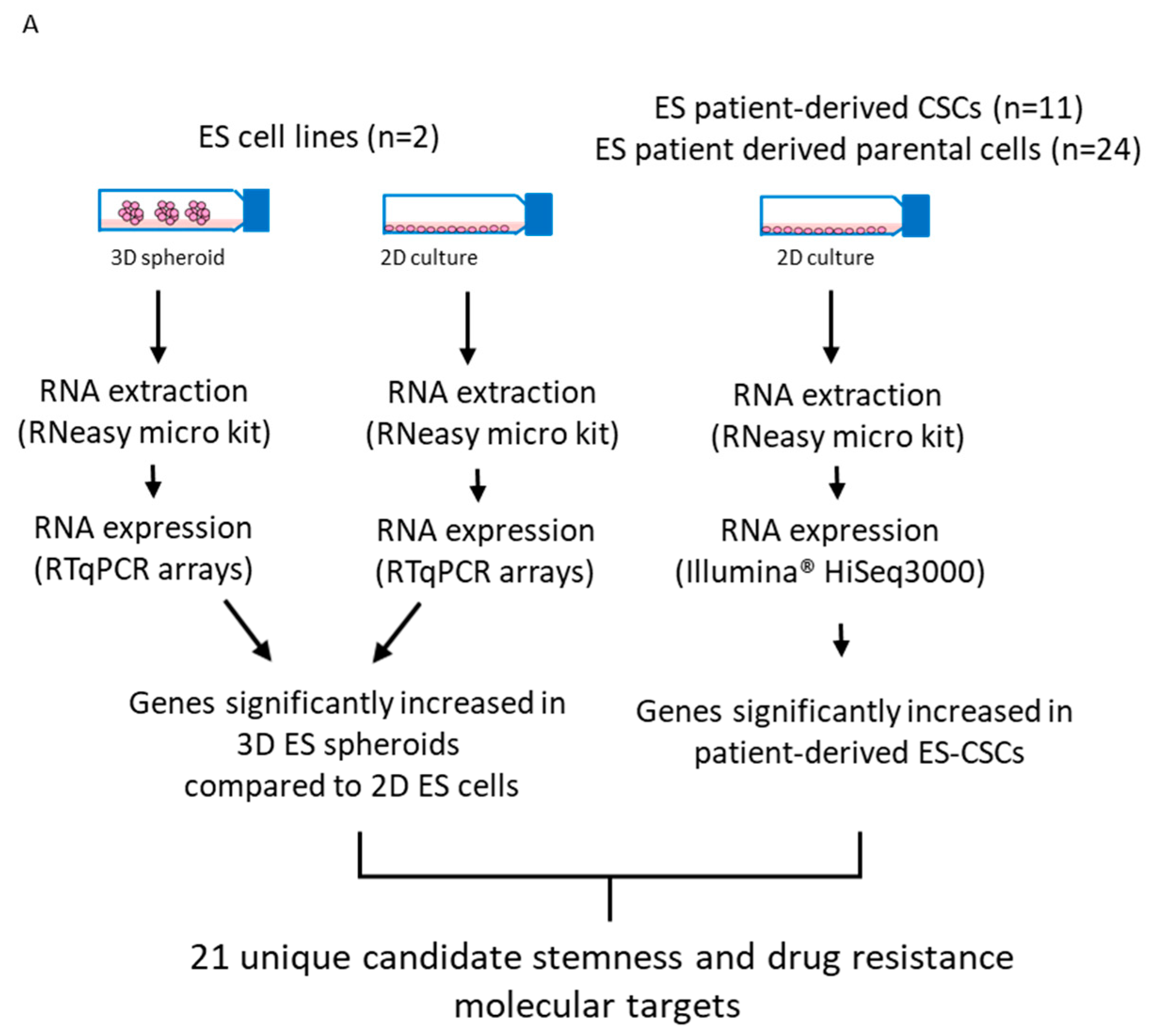
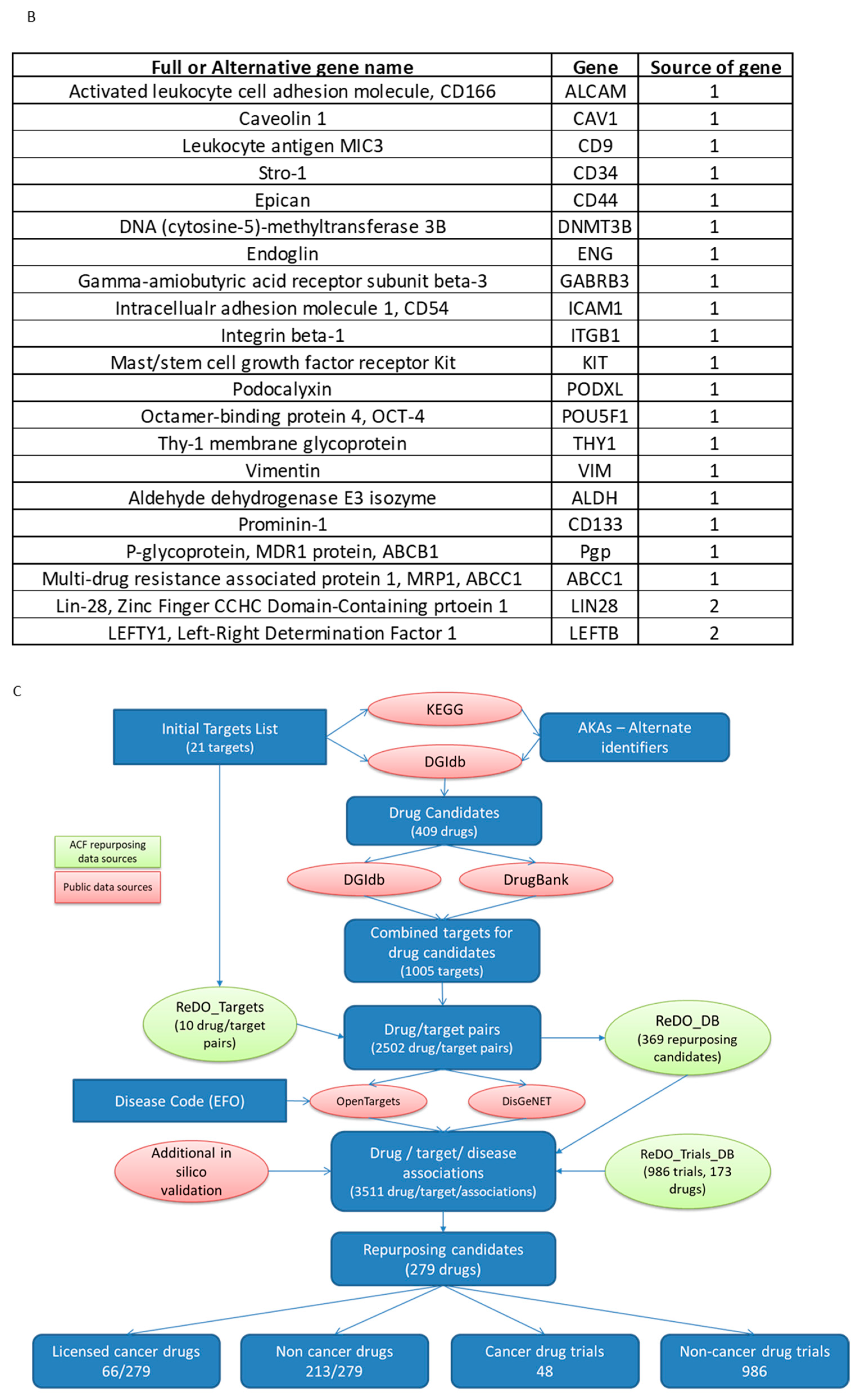
2.11. Identification of Candidate Drugs for Prioritised Molecular Targets
3. Results
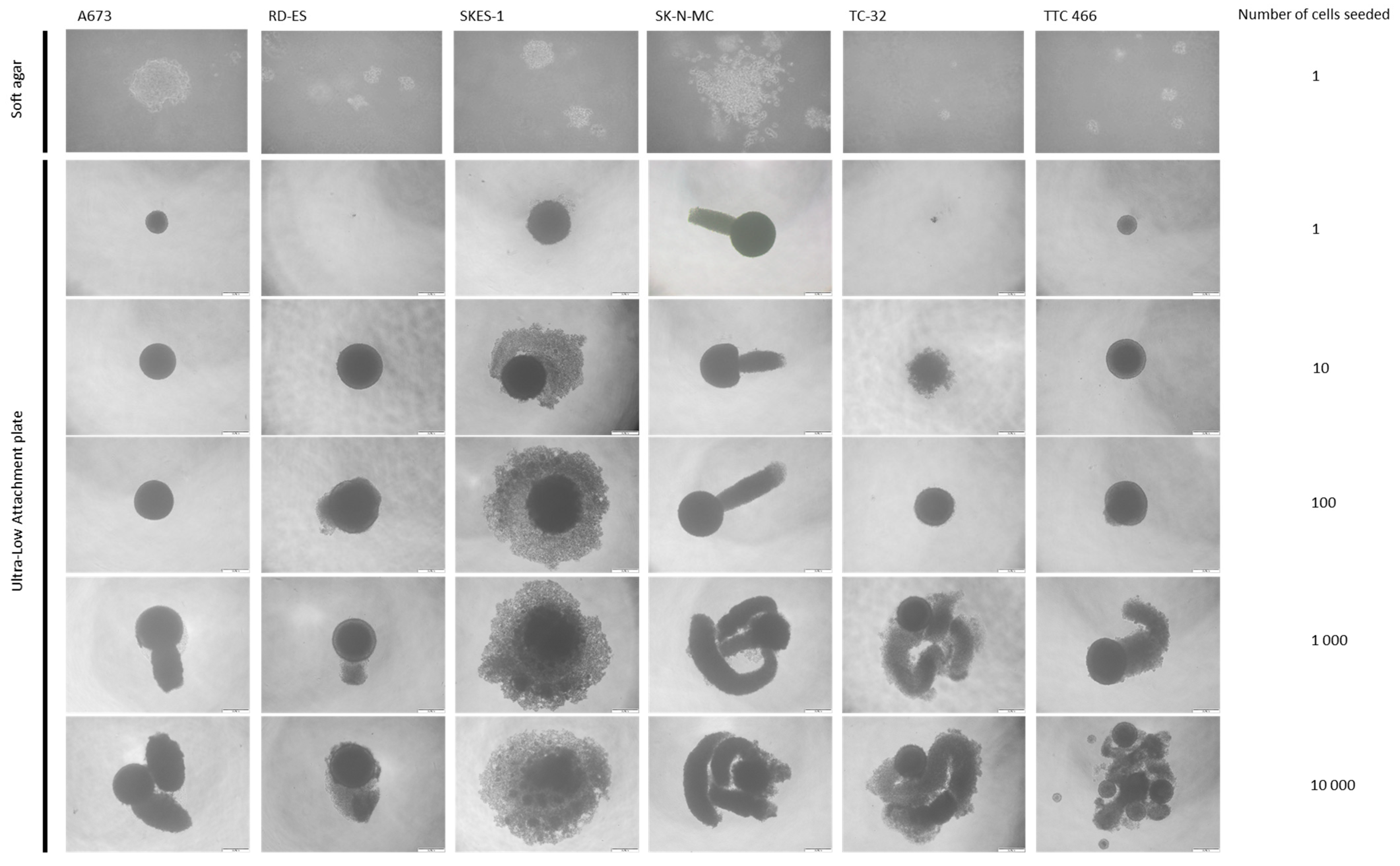
3.1. ES Cell Lines Produce Spheroids and Clones from a Single Cell
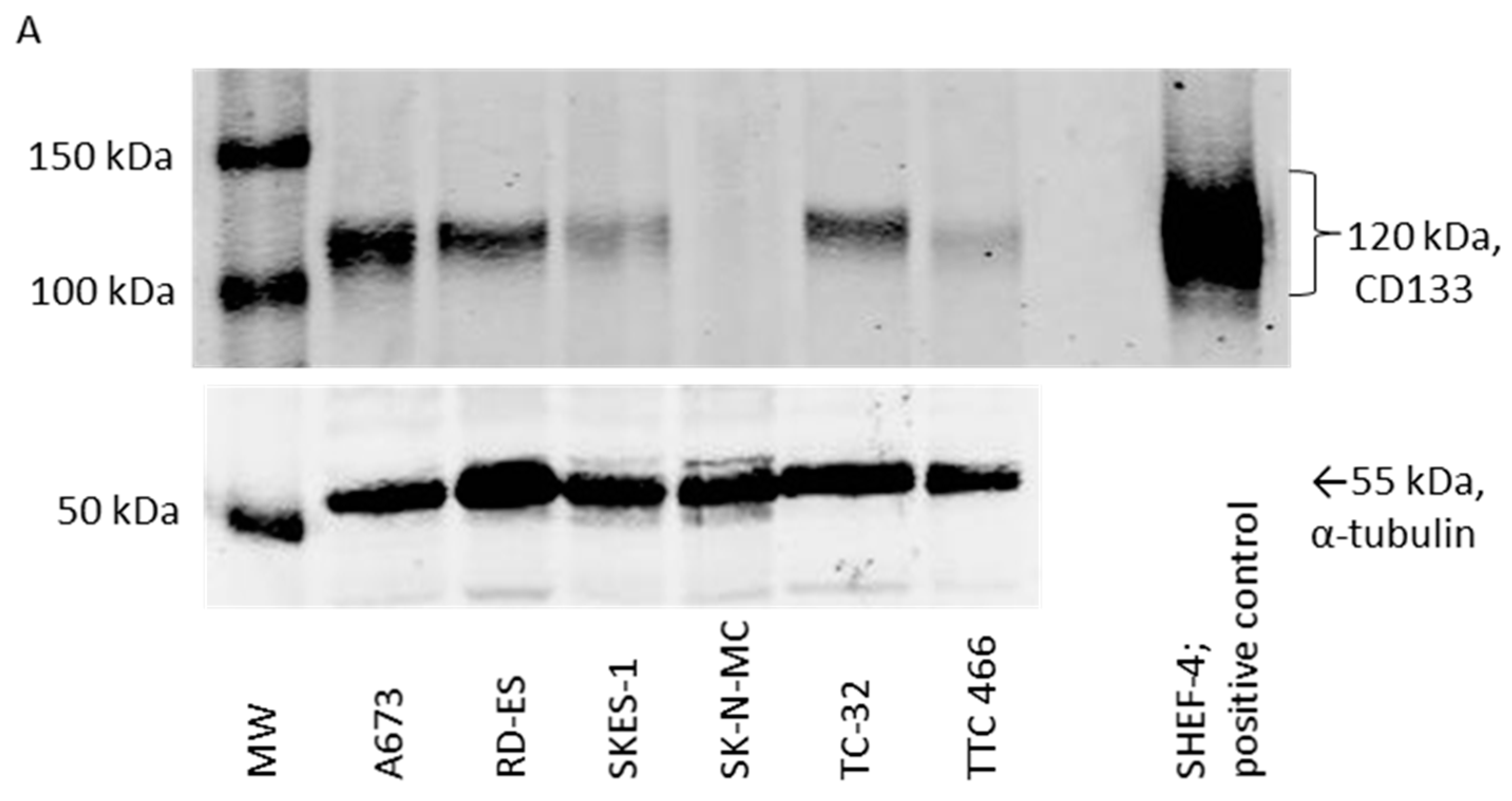
3.2. CD133 Identifies Some Self-Renewing Drug-Resistant ES-CSCs
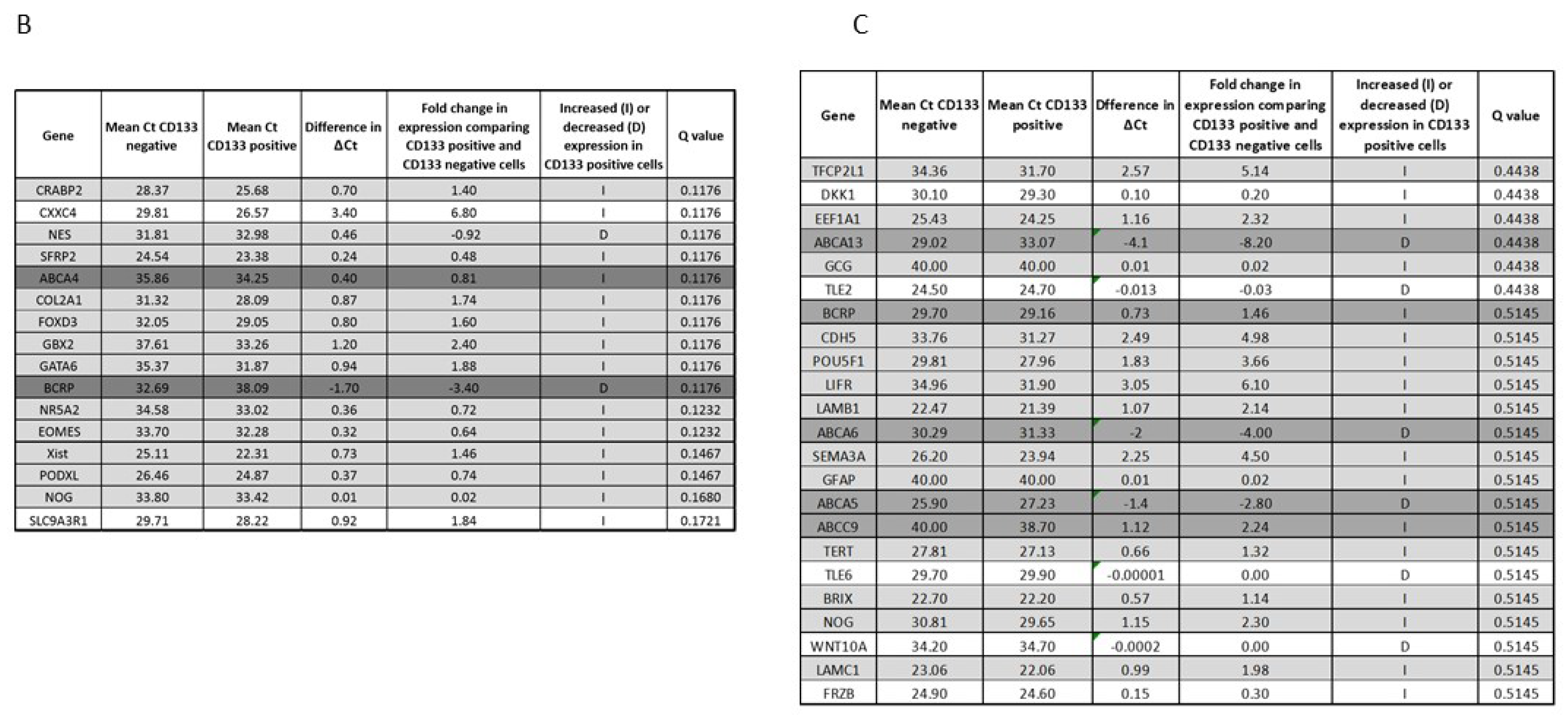
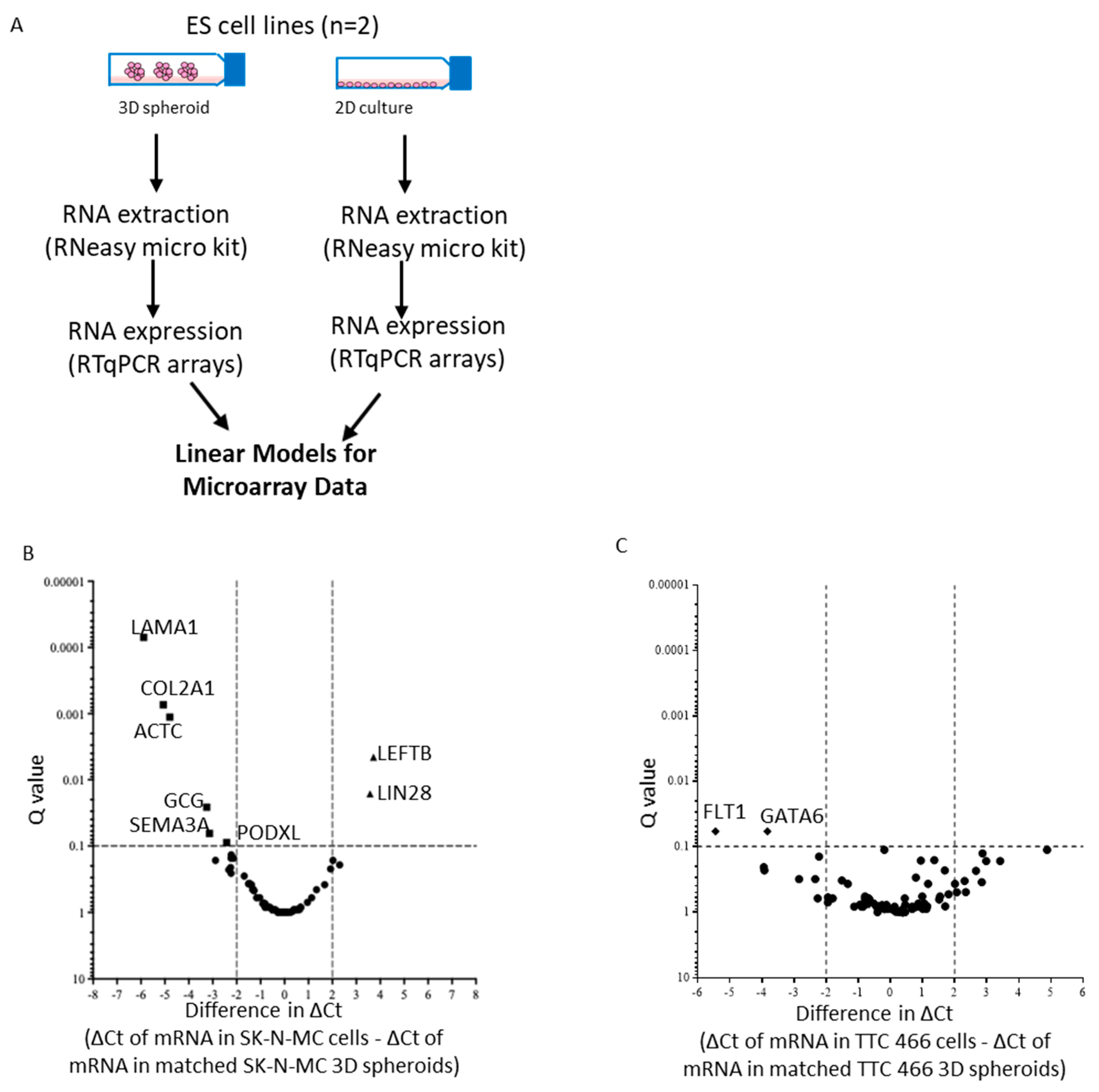
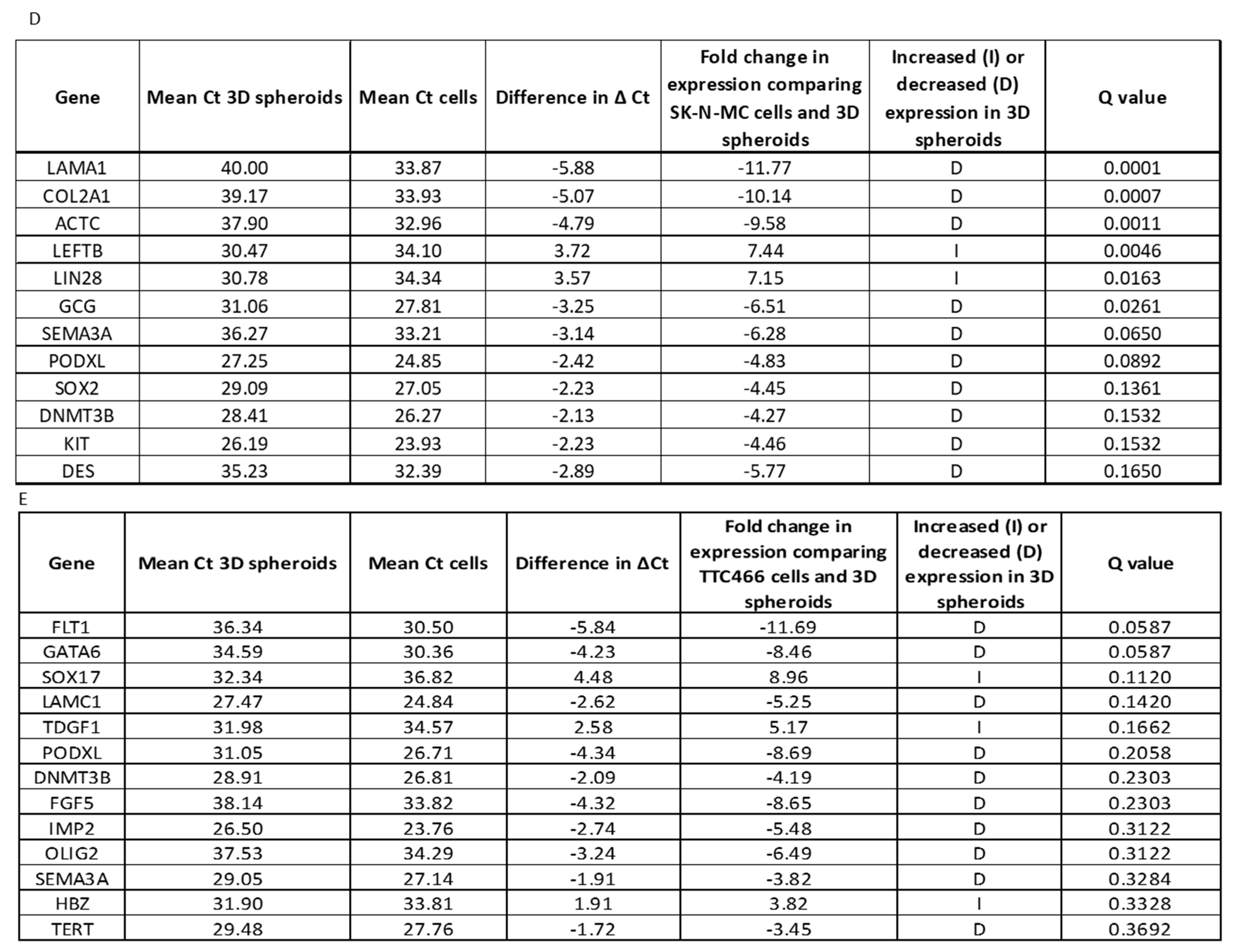
3.3. Gene Expression Profile of CD133 Low or Negative ES 3D Spheroids and 2D Cultures
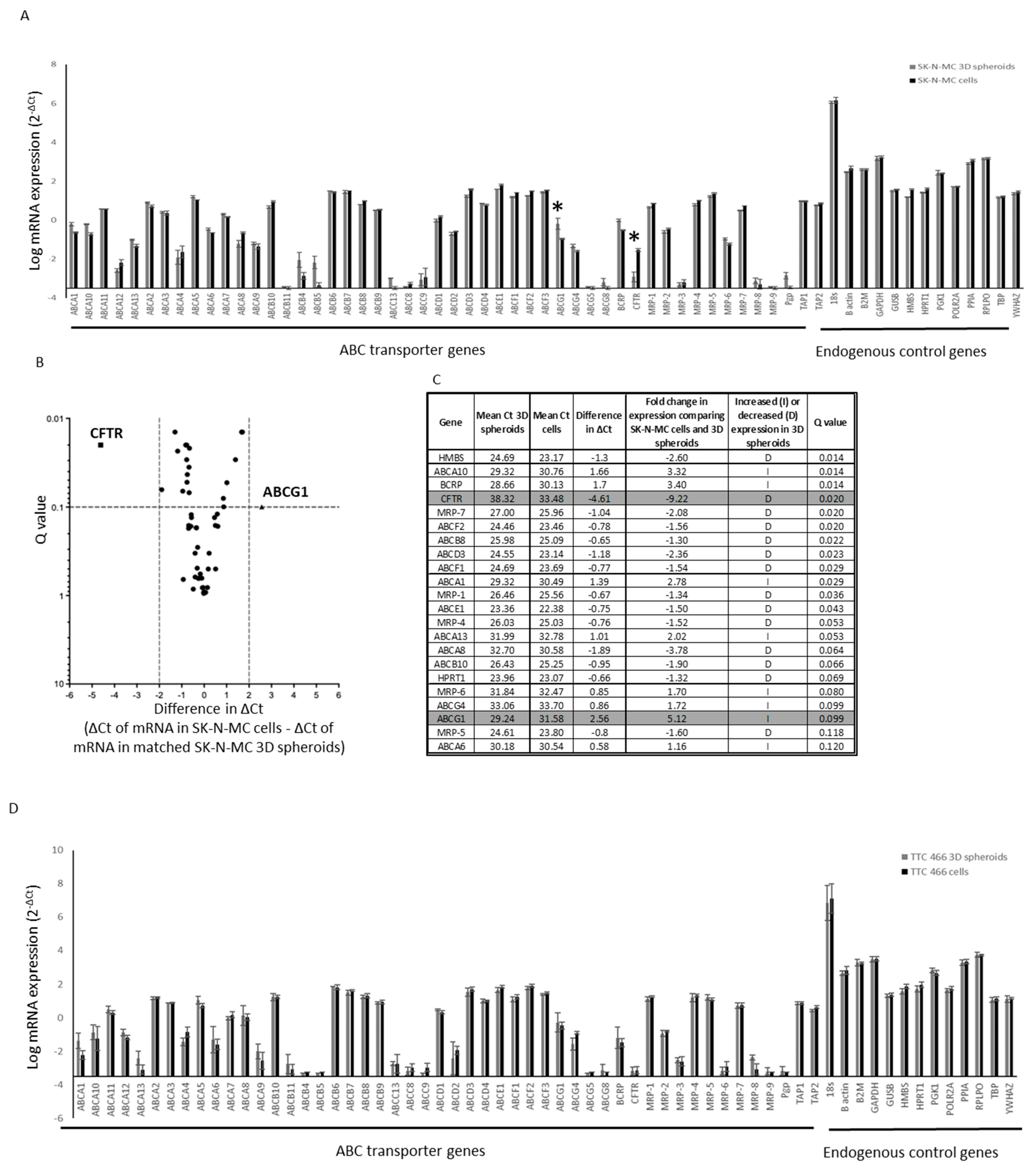
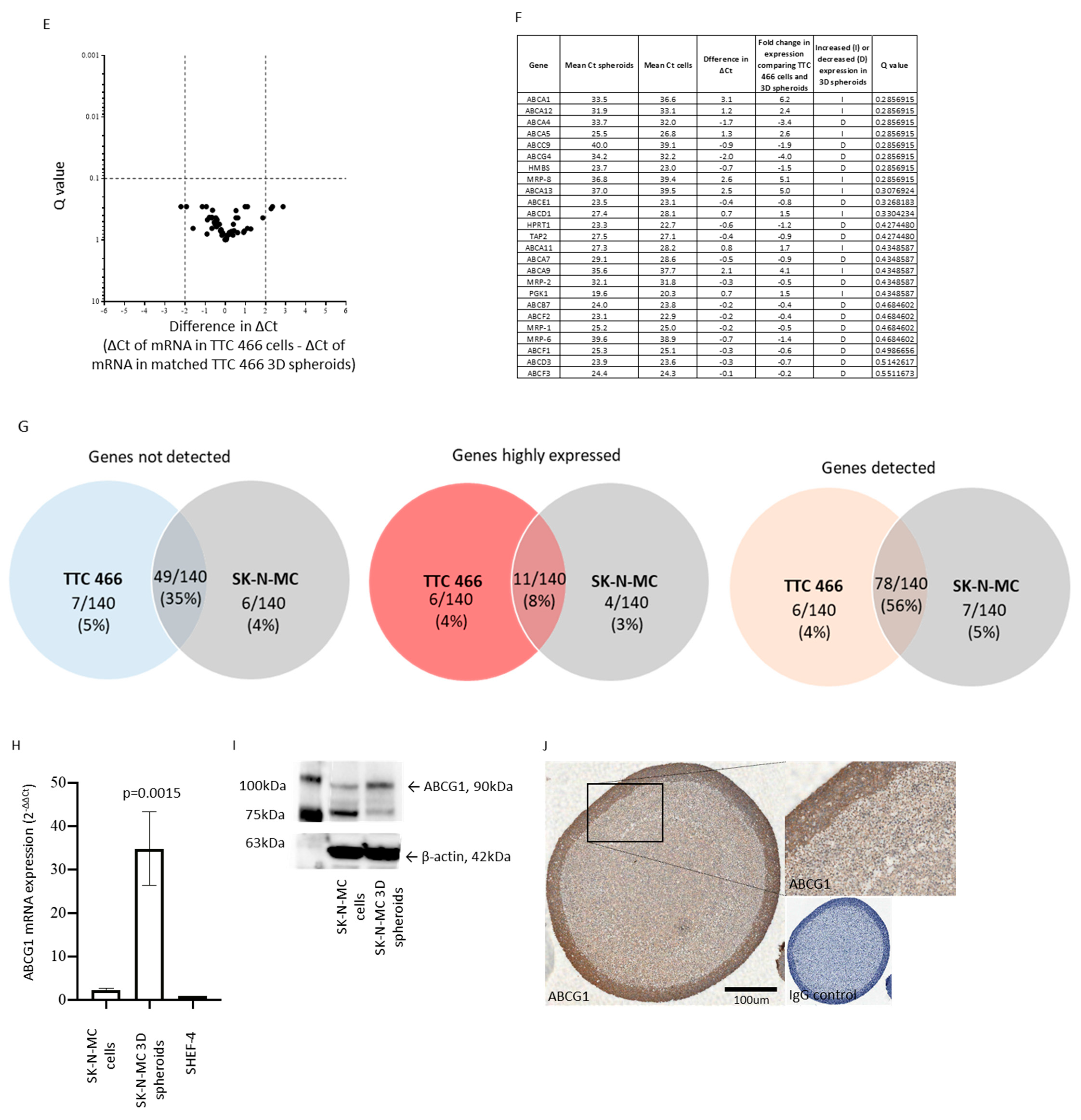
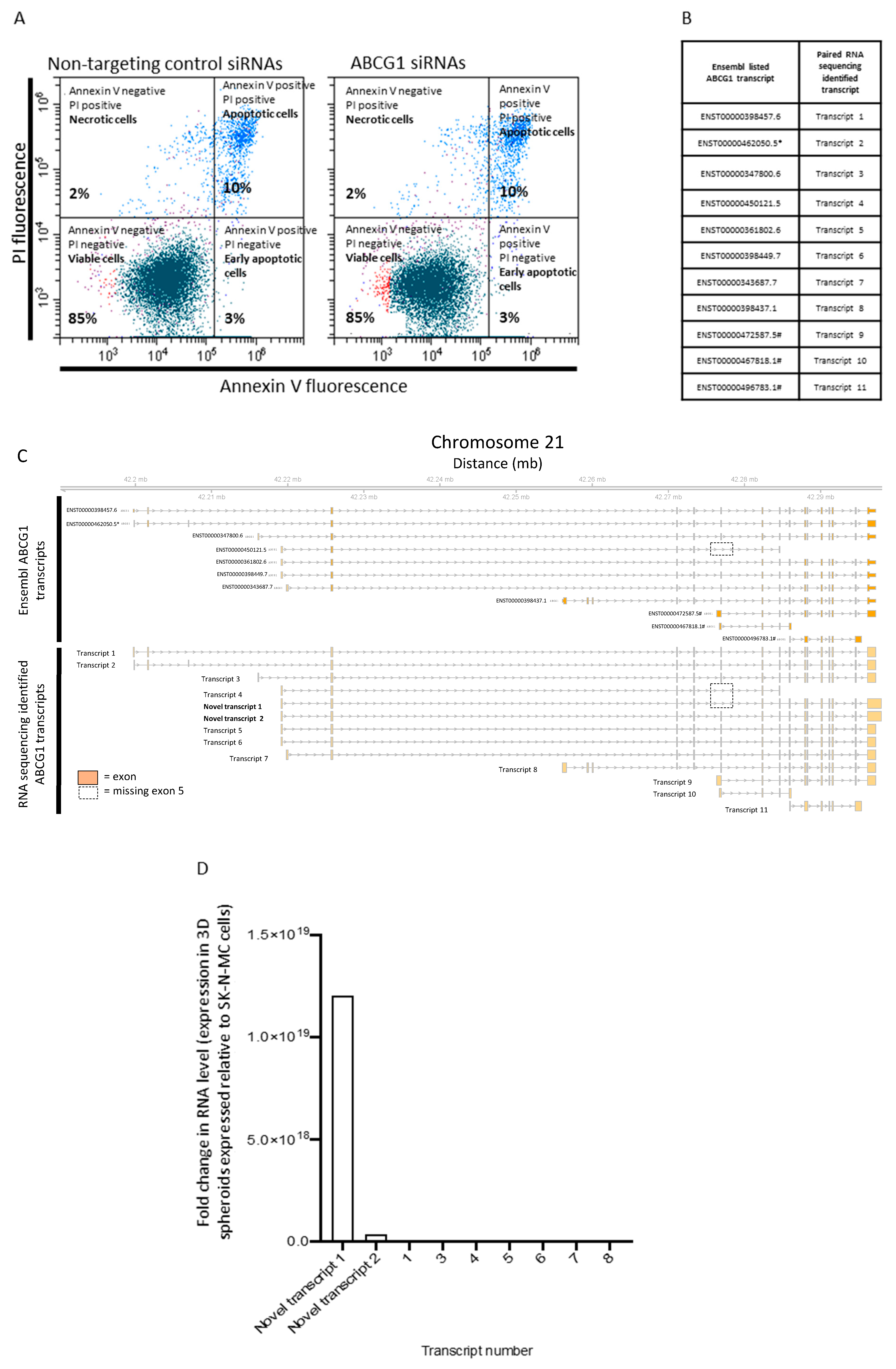
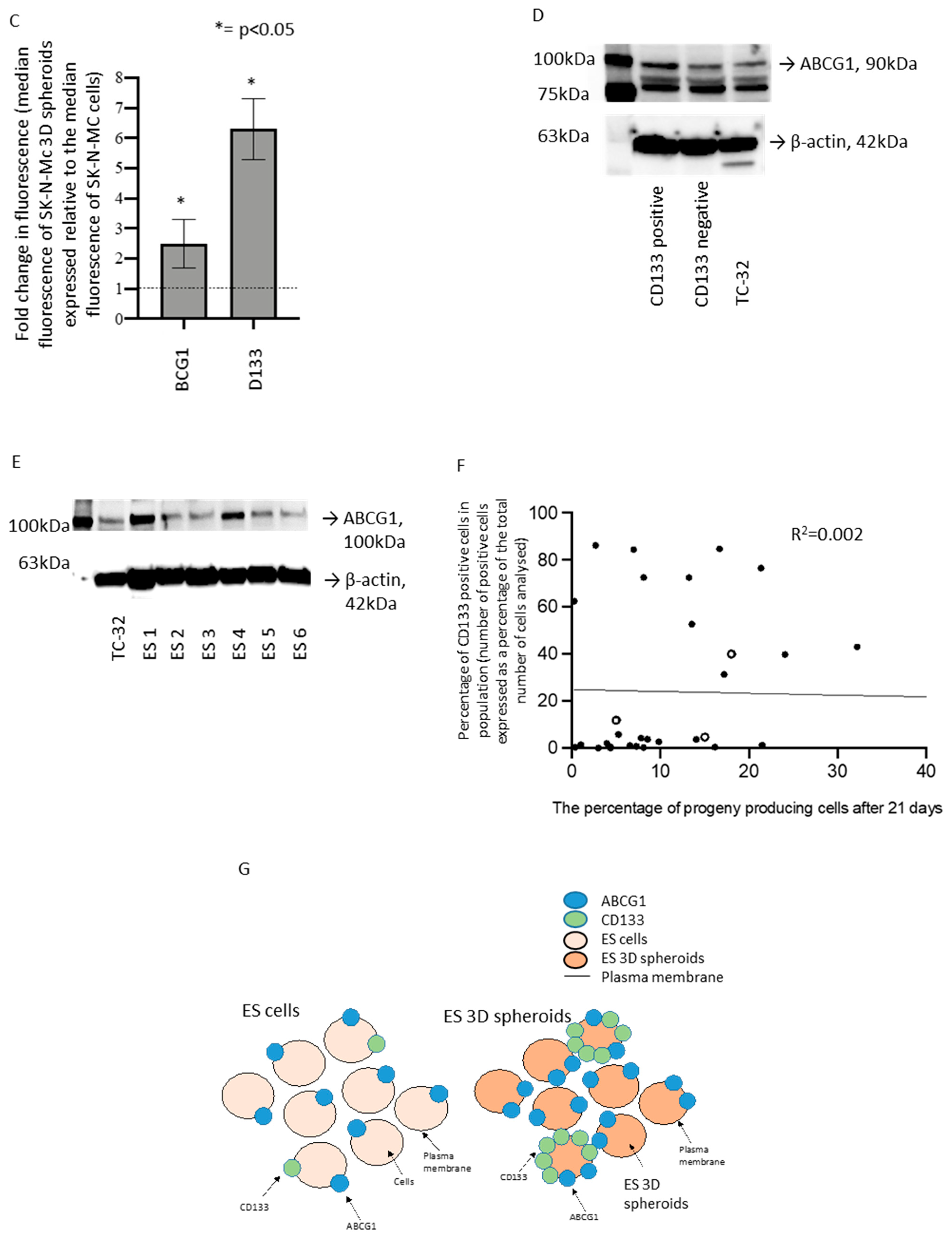
3.4. Functional Role and Characterisation of ABCG1 in ES-CSCs
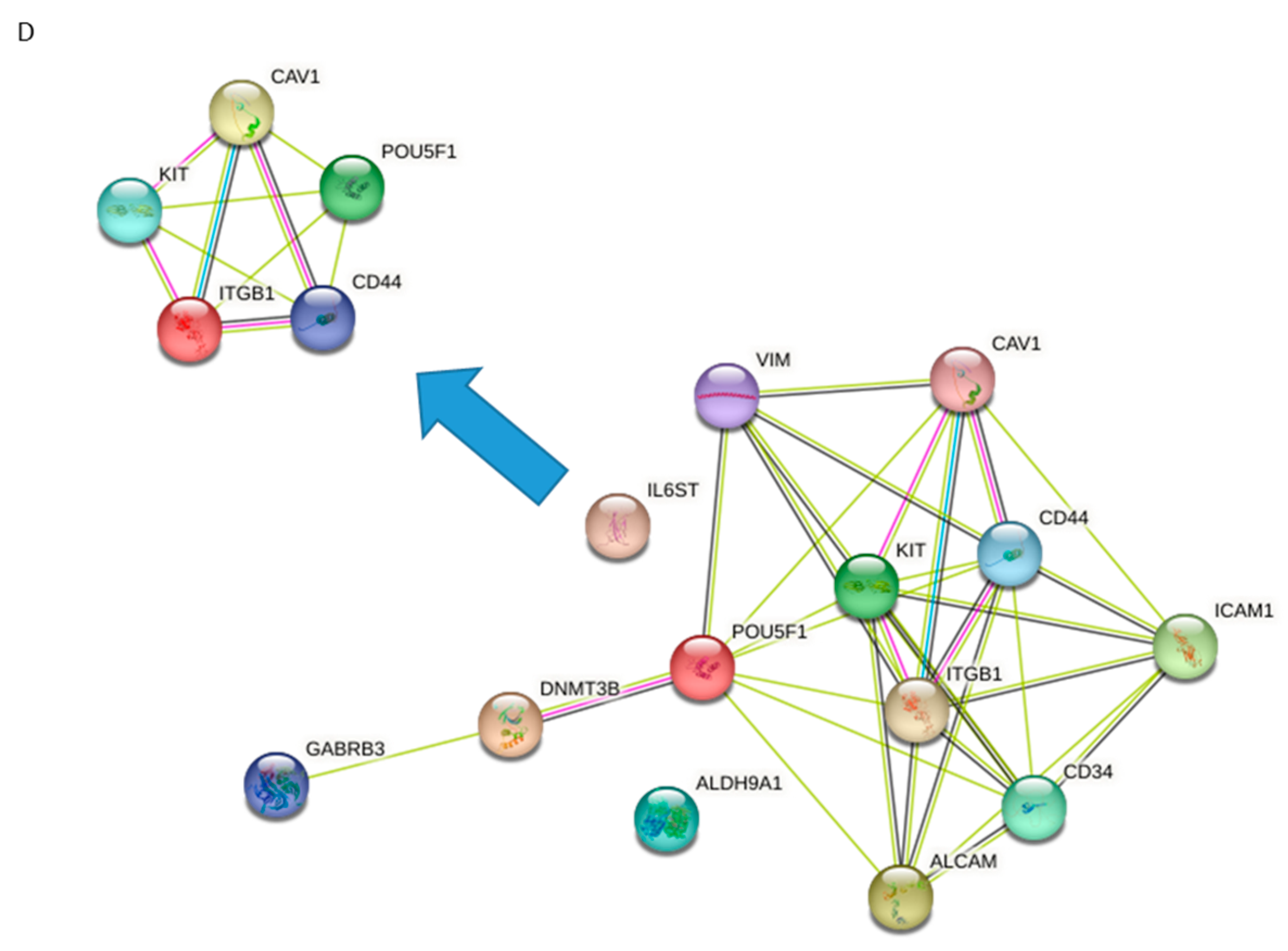
3.5. Identification of Candidate Drugs for ES-CSC Molecular Targets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balamuth, N.J.; Womer, R.B. Ewing’s sarcoma. Lancet Oncol. 2010, 11, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Cotterill, S.J.; Ahrens, S.; Paulussen, M.; Jurgens, H.F.; Voute, P.A.; Gadner, H.; Craft, A.W. Prognostic factors in Ewing’s tumor of bone: Analysis of 975 patients from the European Intergroup Cooperative Ewing’s Sarcoma Study Group. J. Clin. Oncol. 2000, 18, 3108–3114. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, S.; Hoffmann, C.; Jabar, S.; Braun-Munzinger, G.; Paulussen, M.; Dunst, J.; Rube, C.; Winkelmann, W.; Heinecke, A.; Gobel, U.; et al. Evaluation of prognostic factors in a tumor volume-adapted treatment strategy for localized Ewing sarcoma of bone: The CESS 86 experience. Cooperative Ewing Sarcoma Study. Med. Pediatr. Oncol. 1999, 32, 186–195. [Google Scholar] [CrossRef]
- Rodriguez-Galindo, C.; Navid, F.; Liu, T.; Billups, C.A.; Rao, B.N.; Krasin, M.J. Prognostic factors for local and distant control in Ewing sarcoma family of tumors. Ann. Oncol. 2007, 19, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Stahl, M.; Ranft, A.; Paulussen, M.; Bolling, T.; Vieth, V.; Bielack, S.; Gortitz, I.; Braun-Munzinger, G.; Hardes, J.; Jurgens, H.; et al. Risk of recurrence and survival after relapse in patients with Ewing sarcoma. Pediatr. Blood Cancer 2011, 57, 549–553. [Google Scholar] [CrossRef]
- Barker, L.M.; Pendergrass, T.W.; Sanders, J.E.; Hawkins, D.S. Survival after recurrence of Ewing’s sarcoma family of tumors. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2005, 23, 4354–4362. [Google Scholar] [CrossRef]
- Huang, M.; Lucas, K. Current therapeutic approaches in metastatic and recurrent ewing sarcoma. Sarcoma 2011, 2011, 863210. [Google Scholar] [CrossRef]
- Joo, K.M.; Kim, S.Y.; Jin, X.; Song, S.Y.; Kong, D.S.; Lee, J.I.; Jeon, J.W.; Kim, M.H.; Kang, B.G.; Jung, Y.; et al. Clinical and biological implications of CD133-positive and CD133-negative cells in glioblastomas. Lab. Investig. 2008, 88, 808–815. [Google Scholar] [CrossRef]
- Ogden, A.T.; Waziri, A.E.; Lochhead, R.A.; Fusco, D.; Lopez, K.; Ellis, J.A.; Kang, J.; Assanah, M.; McKhann, G.M.; Sisti, M.B.; et al. Identification of A2B5+CD133- tumor-initiating cells in adult human gliomas. Neurosurgery 2008, 62, 505–514. [Google Scholar] [CrossRef]
- Tirino, V.; Desiderio, V.; d’Aquino, R.; De Francesco, F.; Pirozzi, G.; Graziano, A.; Galderisi, U.; Cavaliere, C.; De Rosa, A.; Papaccio, G.; et al. Detection and characterization of CD133+ cancer stem cells in human solid tumours. PLoS ONE 2008, 3, e3469. [Google Scholar] [CrossRef]
- Walter, D.; Satheesha, S.; Albrecht, P.; Bornhauser, B.C.; D’Alessandro, V.; Oesch, S.M.; Rehrauer, H.; Leuschner, I.; Koscielniak, E.; Gengler, C.; et al. CD133 positive embryonal rhabdomyosarcoma stem-like cell population is enriched in rhabdospheres. PLoS ONE 2011, 6, e19506. [Google Scholar] [CrossRef]
- Wang, J.; Sakariassen, P.O.; Tsinkalovsky, O.; Immervoll, H.; Boe, S.O.; Svendsen, A.; Prestegarden, L.; Rosland, G.; Thorsen, F.; Stuhr, L.; et al. CD133 negative glioma cells form tumors in nude rats and give rise to CD133 positive cells. Int. J. Cancer 2008, 122, 761–768. [Google Scholar] [CrossRef]
- Cornaz-Buros, S.; Riggi, N.; DeVito, C.; Sarre, A.; Letovanec, I.; Provero, P.; Stamenkovic, I. Targeting cancer stem-like cells as an approach to defeating cellular heterogeneity in Ewing sarcoma. Cancer Res. 2014, 74, 6610–6622. [Google Scholar] [CrossRef]
- Suva, M.L.; Riggi, N.; Stehle, J.C.; Baumer, K.; Tercier, S.; Joseph, J.M.; Suva, D.; Clement, V.; Provero, P.; Cironi, L.; et al. Identification of cancer stem cells in Ewing’s sarcoma. Cancer Res. 2009, 69, 1776–1781. [Google Scholar] [CrossRef]
- Jiang, X.; Gwye, Y.; Russell, D.; Cao, C.; Douglas, D.; Hung, L.; Kovar, H.; Triche, T.J.; Lawlor, E.R. CD133 expression in chemo-resistant Ewing sarcoma cells. BMC Cancer 2010, 10, 116. [Google Scholar] [CrossRef]
- Fujii, H.; Honoki, K.; Tsujiuchi, T.; Kido, A.; Yoshitani, K.; Takakura, Y. Sphere-forming stem-like cell populations with drug resistance in human sarcoma cell lines. Int. J. Oncol. 2009, 34, 1381–1386. [Google Scholar]
- Roundhill, E.A.; Chicon-Bosch, M.; Jeys, L.; Parry, M.; Rankin, K.S.; Droop, A.; Burchill, S.A. RNA sequencing and functional studies of patient-derived cells reveal that neurexin-1 and regulators of this pathway are associated with poor outcomes in Ewing sarcoma. Cell Oncol. (Dordr) 2021, 44, 1065–1085. [Google Scholar] [CrossRef]
- Roundhill, E.A.; Jabri, S.; Burchill, S.A. ABCG1 and Pgp identify drug resistant, self-renewing osteosarcoma cells. Cancer Lett. 2019, 453, 142–157. [Google Scholar] [CrossRef]
- Charafe-Jauffret, E.; Ginestier, C.; Iovino, F.; Wicinski, J.; Cervera, N.; Finetti, P.; Hur, M.H.; Diebel, M.E.; Monville, F.; Dutcher, J.; et al. Breast cancer cell lines contain functional cancer stem cells with metastatic capacity and a distinct molecular signature. Cancer Res. 2009, 69, 1302–1313. [Google Scholar] [CrossRef]
- Suzuki, E.; Chiba, T.; Zen, Y.; Miyagi, S.; Tada, M.; Kanai, F.; Imazeki, F.; Miyazaki, M.; Iwama, A.; Yokosuka, O. Aldehyde dehydrogenase 1 is associated with recurrence-free survival but not stem cell-like properties in hepatocellular carcinoma. Hepatol. Res. 2012, 42, 1100–1111. [Google Scholar] [CrossRef]
- Bertolini, F.; Sukhatme, V.P.; Bouche, G. Drug repurposing in oncology--patient and health systems opportunities. Nat. Rev. Clin. Oncol. 2015, 12, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Pantziarka, P.; Verbaanderd, C.; Huys, I.; Bouche, G.; Meheus, L. Repurposing drugs in oncology: From candidate selection to clinical adoption. Semin. Cancer Biol. 2021, 68, 186–191. [Google Scholar] [CrossRef]
- Roundhill, E.A.; Burchill, S.A. Detection and characterisation of multi-drug resistance protein 1 (MRP-1) in human mitochondria. Br. J. Cancer 2012, 106, 1224–1233. [Google Scholar] [CrossRef] [PubMed]
- Aflatoonian, B.; Ruban, L.; Shamsuddin, S.; Baker, D.; Andrews, P.; Moore, H. Generation of Sheffield (Shef) human embryonic stem cell lines using a microdrop culture system. In Vitro Cell Dev. Biol. Anim. 2010, 46, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Skowron, M.; Berthold, F. Reliable transcript quantification by real-time reverse transcriptase-polymerase chain reaction in primary neuroblastoma using normalization to averaged expression levels of the control genes HPRT1 and SDHA. J. Mol. Diagn. 2005, 7, 89–96. [Google Scholar] [CrossRef]
- Lastowska, M.; Viprey, V.; Santibanez-Koref, M.; Wappler, I.; Peters, H.; Cullinane, C.; Roberts, P.; Hall, A.G.; Tweddle, D.A.; Pearson, A.D.; et al. Identification of candidate genes involved in neuroblastoma progression by combining genomic and expression microarrays with survival data. Oncogene 2007, 26, 7432–7444. [Google Scholar] [CrossRef]
- Roundhill, E.; Burchill, S. Membrane expression of MRP-1, but not MRP-1 splicing or Pgp expression, predicts survival in patients with ESFT. Br. J. Cancer 2013, 109, 195–206. [Google Scholar] [CrossRef]
- Kodani, M.; Yang, G.; Conklin, L.M.; Travis, T.C.; Whitney, C.G.; Anderson, L.J.; Schrag, S.J.; Taylor, T.H., Jr.; Beall, B.W.; Breiman, R.F.; et al. Application of TaqMan low-density arrays for simultaneous detection of multiple respiratory pathogens. J. Clin. Microbiol. 2011, 49, 2175–2182. [Google Scholar] [CrossRef]
- Demidenko, R.; Razanauskas, D.; Daniunaite, K.; Lazutka, J.R.; Jankevicius, F.; Jarmalaite, S. Frequent down-regulation of ABC transporter genes in prostate cancer. BMC Cancer 2015, 15, 683. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Brownhill, S.C.; Taylor, C.; Burchill, S.A. Chromosome 9p21 gene copy number and prognostic significance of p16 in ESFT. Br. J. Cancer 2007, 96, 1914–1923. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]
- Brownhill, S.; Cohen, D.; Burchill, S. Proliferation index: A continuous model to predict prognosis in patients with tumours of the Ewing’s sarcoma family. PLoS ONE 2014, 9, e104106. [Google Scholar] [CrossRef]
- Myatt, S.S.; Redfern, C.P.; Burchill, S.A. p38MAPK-Dependent sensitivity of Ewing’s sarcoma family of tumors to fenretinide-induced cell death. Clin. Cancer Res. 2005, 11, 3136–3148. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef]
- Freshour, S.L.; Kiwala, S.; Cotto, K.C.; Coffman, A.C.; McMichael, J.F.; Song, J.J.; Griffith, M.; Griffith, O.L.; Wagner, A.H. Integration of the Drug-Gene Interaction Database (DGIdb 4.0) with open crowdsource efforts. Nucleic Acids Res. 2021, 49, D1144–D1151. [Google Scholar] [CrossRef]
- Pantziarka, P.; Verbaanderd, C.; Sukhatme, V.; Rica Capistrano, I.; Crispino, S.; Gyawali, B.; Rooman, I.; Van Nuffel, A.M.; Meheus, L.; Sukhatme, V.P.; et al. ReDO_DB: The repurposing drugs in oncology database. Ecancermedicalscience 2018, 12, 886. [Google Scholar] [CrossRef]
- Ochoa, D.; Hercules, A.; Carmona, M.; Suveges, D.; Gonzalez-Uriarte, A.; Malangone, C.; Miranda, A.; Fumis, L.; Carvalho-Silva, D.; Spitzer, M.; et al. Open Targets Platform: Supporting systematic drug-target identification and prioritisation. Nucleic Acids Res. 2021, 49, D1302–D1310. [Google Scholar] [CrossRef]
- Pinero, J.; Ramirez-Anguita, J.M.; Sauch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef]
- Pantziarka, P.; Capistrano, I.R.; De Potter, A.; Vandeborne, L.; Bouche, G. An Open Access Database of Licensed Cancer Drugs. Front. Pharmacol. 2021, 12, 627574. [Google Scholar] [CrossRef]
- Pantziarka, P.; Vandeborne, L.; Bouche, G. A Database of Drug Repurposing Clinical Trials in Oncology. Front. Pharmacol. 2021, 12, 790952. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Kwon, J.; Huang, Y.; Tan, J.; Uhl, C.G.; He, R.; Zhou, C.; Liu, Y. Facile Tumor Spheroids Formation in Large Quantity with Controllable Size and High Uniformity. Sci. Rep. 2018, 8, 6837. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, S.; Lam, M.M.T.; Poon, T.C.W.; Sun, L.; Jiao, Y.; Wong, A.S.T.; Lee, L.T.O. Angiotensin II promotes ovarian cancer spheroid formation and metastasis by upregulation of lipid desaturation and suppression of endoplasmic reticulum stress. J. Exp. Clin. Cancer Res. 2019, 38, 116. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Sung, J.S.; Park, Y.S.; Chung, S.; Kim, Y.H. Isolation of spheroid-forming single cells from gastric cancer cell lines: Enrichment of cancer stem-like cells. Biotechniques 2018, 65, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Ivascu, A.; Kubbies, M. Rapid generation of single-tumor spheroids for high-throughput cell function and toxicity analysis. J. Biomol. Screen 2006, 11, 922–932. [Google Scholar] [CrossRef]
- Lee, C.; Lee, C.; Atakilit, A.; Siu, A.; Ramos, D.M. Differential spheroid formation by oral cancer cells. Anticancer Res. 2014, 34, 6945–6949. [Google Scholar]
- Ayob, A.Z.; Ramasamy, T.S. Cancer stem cells as key drivers of tumour progression. J. Biomed. Sci. 2018, 25, 20. [Google Scholar] [CrossRef]
- Ribeiro-Dantas, M.D.C.; Oliveira Imparato, D.; Dalmolin, M.G.S.; de Farias, C.B.; Brunetto, A.T.; da Cunha Jaeger, M.; Roesler, R.; Sinigaglia, M.; Siqueira Dalmolin, R.J. Reverse Engineering of Ewing Sarcoma Regulatory Network Uncovers PAX7 and RUNX3 as Master Regulators Associated with Good Prognosis. Cancers 2021, 13, 1860. [Google Scholar] [CrossRef]
- Cidre-Aranaz, F.; Li, J.; Holting, T.L.B.; Orth, M.F.; Imle, R.; Kutschmann, S.; Ammirati, G.; Ceranski, K.; Carreno-Gonzalez, M.J.; Kasan, M.; et al. Integrative gene network and functional analyses identify a prognostically relevant key regulator of metastasis in Ewing sarcoma. Mol. Cancer 2022, 21, 1. [Google Scholar] [CrossRef]
- Miller, H.E.; Gorthi, A.; Bassani, N.; Lawrence, L.A.; Iskra, B.S.; Bishop, A.J.R. Reconstruction of Ewing Sarcoma Developmental Context from Mass-Scale Transcriptomics Reveals Characteristics of EWSR1-FLI1 Permissibility. Cancers 2020, 12, 948. [Google Scholar] [CrossRef]
- Khoogar, R.; Li, F.; Chen, Y.; Ignatius, M.; Lawlor, E.R.; Kitagawa, K.; Huang, T.H.; Phelps, D.A.; Houghton, P.J. Single-cell RNA profiling identifies diverse cellular responses to EWSR1/FLI1 downregulation in Ewing sarcoma cells. Cell Oncol. 2022, 45, 19–40. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, T.G.; Alonso, M.; Avnet, S.; Banito, A.; Burdach, S.; Cidre-Aranaz, F.; Di Pompo, G.; Distel, M.; Dorado-Garcia, H.; Garcia-Castro, J.; et al. Sarcoma treatment in the era of molecular medicine. EMBO Mol. Med. 2020, 12, e11131. [Google Scholar] [CrossRef] [PubMed]
- Wilding, C.P.; Elms, M.L.; Judson, I.; Tan, A.C.; Jones, R.L.; Huang, P.H. The landscape of tyrosine kinase inhibitors in sarcomas: Looking beyond pazopanib. Expert Rev. Anticancer Ther. 2019, 19, 971–991. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Wojtowicz, K.; Savary, S.; Hamon, Y.; Trombik, T. Do ABC transporters regulate plasma membrane organization? Cell Mol. Biol. Lett. 2020, 25, 37. [Google Scholar] [CrossRef]
- Namba, Y.; Sogawa, C.; Okusha, Y.; Kawai, H.; Itagaki, M.; Ono, K.; Murakami, J.; Aoyama, E.; Ohyama, K.; Asaumi, J.I.; et al. Depletion of Lipid Efflux Pump ABCG1 Triggers the Intracellular Accumulation of Extracellular Vesicles and Reduces Aggregation and Tumorigenesis of Metastatic Cancer Cells. Front. Oncol. 2018, 8, 376. [Google Scholar] [CrossRef]
- Tian, C.; Huang, D.; Yu, Y.; Zhang, J.; Fang, Q.; Xie, C. ABCG1 as a potential oncogene in lung cancer. Exp. Ther. Med. 2017, 13, 3189–3194. [Google Scholar] [CrossRef]
- Yvan-Charvet, L.; Pagler, T.A.; Seimon, T.A.; Thorp, E.; Welch, C.L.; Witztum, J.L.; Tabas, I.; Tall, A.R. ABCA1 and ABCG1 protect against oxidative stress-induced macrophage apoptosis during efferocytosis. Circ. Res. 2010, 106, 1861–1869. [Google Scholar] [CrossRef]
- Xue, J.; Wei, J.; Dong, X.; Zhu, C.; Li, Y.; Song, A.; Liu, Z. ABCG1 deficiency promotes endothelial apoptosis by endoplasmic reticulum stress-dependent pathway. J. Physiol. Sci. 2013, 63, 435–444. [Google Scholar] [CrossRef]
- Apfelbaum, A.A.; Wrenn, E.D.; Lawlor, E.R. The importance of fusion protein activity in Ewing sarcoma and the cell intrinsic and extrinsic factors that regulate it: A review. Front. Oncol. 2022, 12, 1044707. [Google Scholar] [CrossRef]
- Elsnerova, K.; Mohelnikova-Duchonova, B.; Cerovska, E.; Ehrlichova, M.; Gut, I.; Rob, L.; Skapa, P.; Hruda, M.; Bartakova, A.; Bouda, J.; et al. Gene expression of membrane transporters: Importance for prognosis and progression of ovarian carcinoma. Oncol. Rep. 2016, 35, 2159–2170. [Google Scholar] [CrossRef]
- Litviakov, N.V.; Garbukov, E.; Slonimskaia, E.M.; Tsyganov, M.M.; Denisov, E.V.; Vtorushin, S.V.; Khristenko, K.; Zav’ialova, M.V.; Cherdyntseva, N.V. Correlation of metastasis-free survival in patients with breast cancer and changes in the direction of expression of multidrug resistance genes during neoadjuvant chemotherapy. Vopr. Onkol. 2013, 59, 334–340. [Google Scholar]
- Litviakov, N.V.; Cherdyntseva, N.V.; Tsyganov, M.M.; Slonimskaya, E.M.; Ibragimova, M.K.; Kazantseva, P.V.; Kzhyshkowska, J.; Choinzonov, E.L. Deletions of multidrug resistance gene loci in breast cancer leads to the down-regulation of its expression and predict tumor response to neoadjuvant chemotherapy. Oncotarget 2016, 7, 7829–7841. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.; Ready, N.E.; Su, L.; Wei, Y.; Christiani, D.C.; Wei, Q. Genetic variants in ABCG1 are associated with survival of nonsmall-cell lung cancer patients. Int. J. Cancer 2016, 138, 2592–2601. [Google Scholar] [CrossRef]
- Gu, H.M.; Li, G.; Gao, X.; Berthiaume, L.G.; Zhang, D.W. Characterization of palmitoylation of ATP binding cassette transporter G1: Effect on protein trafficking and function. Biochim. Biophys. Acta 2013, 1831, 1067–1078. [Google Scholar] [CrossRef]
- Gu, H.M.; Wang, F.Q.; Zhang, D.W. Caveolin-1 interacts with ATP binding cassette transporter G1 (ABCG1) and regulates ABCG1-mediated cholesterol efflux. Biochim. Biophys. Acta 2014, 1841, 847–858. [Google Scholar] [CrossRef]
- Riggi, N.; Suva, M.L.; Stamenkovic, I. The cancer stem cell paradigm in Ewing’s sarcoma: What can we learn about these rare cells from a rare tumor? Expert Rev. Anticancer Ther. 2011, 11, 143–145. [Google Scholar] [CrossRef]
- Glumac, P.M.; LeBeau, A.M. The role of CD133 in cancer: A concise review. Clin. Transl. Med. 2018, 7, 18. [Google Scholar] [CrossRef]
- Rappa, G.; Fodstad, O.; Lorico, A. The stem cell-associated antigen CD133 (Prominin-1) is a molecular therapeutic target for metastatic melanoma. Stem Cells 2008, 26, 3008–3017. [Google Scholar] [CrossRef]
- Danieau, G.; Morice, S.; Redini, F.; Verrecchia, F.; Royer, B.B. New Insights about the Wnt/beta-Catenin Signaling Pathway in Primary Bone Tumors and Their Microenvironment: A Promising Target to Develop Therapeutic Strategies? Int. J. Mol. Sci. 2019, 20, 3751. [Google Scholar] [CrossRef]
- Tirode, F.; Laud-Duval, K.; Prieur, A.; Delorme, B.; Charbord, P.; Delattre, O. Mesenchymal stem cell features of Ewing tumors. Cancer Cell 2007, 11, 421–429. [Google Scholar] [CrossRef]
- von Levetzow, C.; Jiang, X.; Gwye, Y.; von Levetzow, G.; Hung, L.; Cooper, A.; Hsu, J.H.; Lawlor, E.R. Modeling initiation of Ewing sarcoma in human neural crest cells. PLoS ONE 2011, 6, e19305. [Google Scholar] [CrossRef] [PubMed]
- Jawad, M.U.; Cheung, M.C.; Min, E.S.; Schneiderbauer, M.M.; Koniaris, L.G.; Scully, S.P. Ewing sarcoma demonstrates racial disparities in incidence-related and sex-related differences in outcome: An analysis of 1631 cases from the SEER database, 1973-2005. Cancer 2009, 115, 3526–3536. [Google Scholar] [CrossRef] [PubMed]
- Machiela, M.J.; Grunewald, T.G.P.; Surdez, D.; Reynaud, S.; Mirabeau, O.; Karlins, E.; Rubio, R.A.; Zaidi, S.; Grossetete-Lalami, S.; Ballet, S.; et al. Genome-wide association study identifies multiple new loci associated with Ewing sarcoma susceptibility. Nat. Commun. 2018, 9, 3184. [Google Scholar] [CrossRef] [PubMed]
- Randall, R.L.; Lessnick, S.L.; Jones, K.B.; Gouw, L.G.; Cummings, J.E.; Cannon-Albright, L.; Schiffman, J.D. Is There a Predisposition Gene for Ewing’s Sarcoma? J. Oncol. 2010, 2010, 397632. [Google Scholar] [CrossRef]
- Grunewald, T.G.P.; Cidre-Aranaz, F.; Surdez, D.; Tomazou, E.M.; de Alava, E.; Kovar, H.; Sorensen, P.H.; Delattre, O.; Dirksen, U. Ewing sarcoma. Nat. Rev. Dis. Prim. 2018, 4, 5. [Google Scholar] [CrossRef]
- Kumar, S.V.; Er, P.X.; Lawlor, K.T.; Motazedian, A.; Scurr, M.; Ghobrial, I.; Combes, A.N.; Zappia, L.; Oshlack, A.; Stanley, E.G.; et al. Kidney micro-organoids in suspension culture as a scalable source of human pluripotent stem cell-derived kidney cells. Development 2019, 146, dev172361. [Google Scholar] [CrossRef]
- Keller, F.; Rudiger, R.; MAthias, H. Towards optimized breast cancer 3D spheroid mono- and co-culture models for pharmacological research and screening. J. Cell. Biotechnol. 2019, 5, 89–101. [Google Scholar] [CrossRef]
- Wang, Y.J.; Herlyn, M. The emerging roles of Oct4 in tumor-initiating cells. Am. J. Physiol. Cell Physiol. 2015, 309, C709–C718. [Google Scholar] [CrossRef]
- Kim, J.; Kim, H.S.; Shim, J.J.; Lee, J.; Kim, A.Y.; Kim, J. Critical role of the fibroblast growth factor signalling pathway in Ewing’s sarcoma octamer-binding transcription factor 4-mediated cell proliferation and tumorigenesis. FEBS J. 2019, 286, 4443–4472. [Google Scholar] [CrossRef]
- Chua, V.; Chawla, S.P.; Zheng, K.; Kim, T.; Del Priore, G.; Kim, S. Phase II study of SM-88 in Ewing’s and other sarcomas. In Proceedings of the 2021 ASCO Annual Meeting I, Chicago, IL, USA, 28 May 2021. [Google Scholar]
- Chung, V.; Chawla, S.P.; Dong, H.; Kim, S.; Korn, R.L.; Lim, K.-H.; Noel, M.S.; Noonan, A.M.; Oberstein, P.E.; Ocean, A.J.; et al. Phase II/III study of SM-88 in patients with metastatic pancreatic cancer. In Proceedings of the 2021 Gastrointestinal Cancers Symposium, Online, 20 January 2021. [Google Scholar]
- Brennan, B.; Kirton, L.; Marec-Berard, P.; Martin-Broto, J.; Gelderblom, H.; Gaspar, N.; Strauss, S.; Sastre Urgelles, A.; Anderton, J.; Laurence, V.; et al. Comparison of two chemotherapy regimens in Ewing sarcoma (ES): Overall and subgroup results of the Euro Ewing 2012 randomized trial (EE2012). In Proceedings of the 2020 ASCO Annual Meeting I, Virtual, 25 May 2020; p. 11500. [Google Scholar]
- Serra, M.; Hattinger, C.M.; Pasello, M.; Casotti, C.; Fantoni, L.; Riganti, C.; Manara, M.C. Impact of ABC Transporters in Osteosarcoma and Ewing’s Sarcoma: Which Are Involved in Chemoresistance and Which Are Not? Cells 2021, 10, 2461. [Google Scholar] [CrossRef]
- Perri, T.; Fogel, M.; Mor, S.; Horev, G.; Meller, I.; Loven, D.; Issakov, J.; Kollender, Y.; Smirnov, A.; Zaizov, R.; et al. Effect of P-glycoprotein expression on outcome in the Ewing family of tumors. Pediatr. Hematol. Oncol. 2001, 18, 325–334. [Google Scholar] [CrossRef]
- Scotlandi, K.; Manara, M.C.; Hattinger, C.M.; Benini, S.; Perdichizzi, S.; Pasello, M.; Bacci, G.; Zanella, L.; Bertoni, F.; Picci, P.; et al. Prognostic and therapeutic relevance of HER2 expression in osteosarcoma and Ewing’s sarcoma. Eur. J. Cancer 2005, 41, 1349–1361. [Google Scholar] [CrossRef]
- Roessner, A.; Ueda, Y.; Bockhorn-Dworniczak, B.; Blasius, S.; Peters, A.; Wuisman, P.; Ritter, J.; Paulussen, M.; Jurgens, H.; Bocker, W. Prognostic implication of immunodetection of P glycoprotein in Ewing’s sarcoma. J. Cancer Res. Clin. Oncol. 1993, 119, 185–189. [Google Scholar] [CrossRef]
- Mancarella, C.; Pasello, M.; Ventura, S.; Grilli, A.; Calzolari, L.; Toracchio, L.; Lollini, P.L.; Donati, D.M.; Picci, P.; Ferrari, S.; et al. Insulin-Like Growth Factor 2 mRNA-Binding Protein 3 is a Novel Post-Transcriptional Regulator of Ewing Sarcoma Malignancy. Clin. Cancer Res. 2018, 24, 3704–3716. [Google Scholar] [CrossRef]
- Eisenmann, E.D.; Talebi, Z.; Sparreboom, A.; Baker, S.D. Boosting the oral bioavailability of anticancer drugs through intentional drug-drug interactions. Basic Clin. Pharmacol. Toxicol. 2022, 130 (Suppl. S1), 23–35. [Google Scholar] [CrossRef]
- Kang, Y. Analysis of cancer stem cell metastasis in xenograft animal models. Methods Mol. Biol. 2009, 568, 7–19. [Google Scholar] [CrossRef]
- Sampieri, K.; Fodde, R. Cancer stem cells and metastasis. Semin. Cancer Biol. 2012, 22, 187–193. [Google Scholar] [CrossRef]
- O’Callaghan, N.; Dhillon, V.; Thomas, P.; Fenech, M. A quantitative real-time PCR method for absolute telomere length. Biotechniques 2008, 44, 807–809. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Clone or Spheroid Formation (%) | |||||
|---|---|---|---|---|---|---|
| Clone Formation in Soft Agar from a Single Cell | Ultra-Low Attachment Plate | |||||
| 1 Cell | 10 Cells | 100 Cells | 1000 Cells | 10,000 Cells | ||
| A673 | 17 ±1 | 18 ± 5 | 88 ± 10 | 100 | 100 | 100 |
| RD-ES | 17 ±1 | 5 ± 1 | 45 ± 13 | 100 | 100 | 100 |
| SKES-1 | 29 ± 2 (p < 0.05) | 75 ± 7 (p < 0.05) | 98 ± 2 | 100 | 100 | 100 |
| SK-N-MC | 12 ± 1 | 64 ± 1 | 97 ± 3 | 100 | 100 | 100 |
| TC-32 | 7 ± 0.5 (p < 0.05) | 15 ± 3 | 93 ± 9 | 100 | 100 | 100 |
| TTC 466 | 13 ± 1 | 45 ± 2 | 96 ± 6 | 100 | 100 | 100 |
| Drug | Ewings Sarcoma Targets | Cancer Drug | BNFC approval | Off-Patent | Trial Count | Max Trial Phase | Trial Phase (Number of Trials in Phase) | Trial Status (Number of Trials in Status) | Cancer Type (Number of Trials in Cancer Type) | Trial Information for Phase 3 and Phase 4 trials (Trial Number, Phase, Status, Recruitment Status, Cancer Type) |
|---|---|---|---|---|---|---|---|---|---|---|
| Cabozantinib | KIT, RET, KDR, MET | Y | N | N | 6 | Phase 2 | Phase 1 (1), Phase 2 (5) | Recruiting (1), Active, not recruiting (3), Not yet recruiting (1), Other (1) | Multiple sarcomas, including Ewing’s (1), Mixed paediatric, including Ewing’s (3), Ewing’s or osteosarcoma (2) | NA |
| Dasatinib | KIT, MAPK14, LYN, HSPA8, EPHB4, CSK, BCR, BTK, PDGFRB, EPHA2, SRC, ABL1 | Y | Y | Y | 2 | Phase 2 | Phase 1/2 (1), Phase 2 (1) | Active, not recruiting (1), Completed (1) | Mixed paediatric, including Ewing’s (1), Multiple sarcomas, including Ewing’s (1) | NA |
| Decitabine | DNMT3B, DNMT1 | Y | N | Y | 1 | Phase 1 | Phase 1 (1) | Completed (1) | Mixed, including sarcomas (1) | NA |
| Erdafitinib | KIT, KDR, PDGFRB, PDGFRA, CSF1R, RET, FGFR1 | Y | N | N | 1 | Phase 2 | Phase 2 (1) | Recruiting (1) | Mixed paediatric, including Ewing’s (1) | NA |
| Imatinib | KIT, ABCB1, PDGFRB, ABL1, PDGFRA, CSF1R, NTRK1, RET, BCR | Y | Y | Y | 4 | Phase 2 | Phase 2 (4) | Completed (4) | Mixed, including sarcomas (1), Multiple sarcomas, including Ewing’s (1), Mixed paediatric, including Ewing’s (1), Ewing’s or DSRCT (1) | NI |
| Lenvatinib | KIT, RET, PDGFRA, FGFR1, FLT4, KDR, FLT1 | Y | N | N | 1 | Phase 1/2 | Phase 1/2 (1) | Active, not recruiting (1) | Mixed paediatric, including Ewing’s (1) | NA |
| Pazopanib | KIT, ITK, PDGFRB, PDGFRA, FLT4, KDR, FLT1 | Y | N | Y | 1 | Phase 2 | Phase 2 (1) | Completed (1) | Mixed paediatric, including Ewing’s (1) | NA |
| Regorafenib | KIT, RET, ABL1, BRAF, EPHA2, NTRK1, TEK, FGFR1, PDGFRB, PDGFRA, FLT4, KDR, FLT1 | Y | N | N | 5 | Phase 2 | Phase 1 (1), Phase 1/2 (1), Phase 2 (2), Other (1) | Recruiting (4), Active, not recruiting (1) | Multiple sarcomas, including Ewing’s (3), Ewing’s or osteosarcoma (1), Mixed paediatric, including Ewing’s (1) | NA |
| Sorafenib | KIT, ABCB1, FLT1, RET, FGFR1, PDGFRB, FLT3, KDR, FLT4, BRAF | Y | N | Unclear | 2 | Phase 2 | Phase 1 (1), Phase 2 (1) | Active, not recruiting (1), Completed (1) | Ewing’s or DSRCT (1), Mixed paediatric, including Ewing’s (1) | NA |
| Sunitinib | KIT, ABCB1, PDGFRA, CSF1R, FLT3, FLT4, KDR, FLT1, PDGFRB | Y | N | Y | 2 | Phase 2 | Phase 1/2 (1), Phase 2 (1) | Ongoing (1), Completed (1) | Multiple sarcomas, including Ewing’s (1), Mixed, including sarcomas (1) | NA |
| Temozolomide | ABCB1 | Y | Y | Y | 23 | Phase 3 | Phase 1 (10), Phase 1/2 (4), Phase 2 (8), Phase 3 (1) | Recruiting (8), Active, not recruiting (4), Completed (6), Not yet recruiting (2), Other (2), Terminated (1) | Mixed paediatric, including Ewing’s (6), Ewing’s (10), Mixed, including sarcomas (4), Ewing’s or DSRCT (1), Ewing’s or RMS (2) | NCT03495921 (Phase 3; Active, not recruiting)—Ewing sarcoma |
| Aliskiren | ABCB1 | N | N | Y | 1 | Phase 2 | Phase 2 (1) | Recruiting (1) | Multiple cancer types (1) | NA |
| Allopurinol | POU5F1 | N | Y | Y | 1 | Phase 1/2 | Phase 1/2 (1) | Not yet recruiting (1) | Lung (1) | NA |
| Atorvastatin | ABCB1, NR1I3, HDAC2, AHR, DPP4, HMGCR | N | Y | Y | 16 | Phase 3 | Phase 1 (4), Phase 2 (6), Phase 2/3 (1), Phase 3 (5) | Recruiting (13), Ongoing (1), Not yet recruiting (2) | Multiple cancer types, Leukaemia (1), GI (4), Breast (8), Urological (3) | NCT03024684 (Phase 3; Recruiting)—Hepatocellular carcinoma, NCT03819101 (Phase 3; Recruiting)—Prostate, NCT03971019 (Phase 3; Recruiting)—Breast, NCT04601116 (Phase 3; Recruiting)—Breast, NCT04026230 (Phase 3; Recruiting)—Prostate |
| Azithromycin | ABCC1 | N | Y | Y | 3 | Phase 2 | Phase 2 (3) | Ongoing (3) | Lymphoma (2), Breast (1) | NA |
| Carvedilol | ABCB1, HIF1A, GJA1, VEGFA | N | Y | Y | 2 | Phase 2 | Phase 1 (1), Phase 2 (1) | Active, not recruiting (1), Not yet recruiting (1) | Urological (1), CNS (1) | NA |
| Chlorpromazine | ABCB1 | N | N | Y | 3 | Phase 2 | Phase 1 (1), Phase 1/2 (1), Phase 2 (1) | Recruiting (2), Not yet recruiting (1) | GI (1), CNS (2) | NA |
| Citalopram | ABCB1 | N | Y | Y | 1 | Phase 3 | Phase 3 (1) | Ongoing (1) | CNS (1) | 2013-004705-59 (Phase 3; Ongoing)—Glioblastoma |
| Clarithromycin | ABCB1 | N | Y | Y | 18 | Phase 4 | Phase 1/2 (2), Phase 2 (10), Phase 3 (3), Phase 4 (1), Other (2) | Recruiting (8), Active, not recruiting (8), Not yet recruiting (1), Other (1) | Lymphoma (1), Other Haem-onc (15), Multiple cancer types (1), GI (1) | ChiCTR2100047608 (Phase 4; Recruiting)—Multiple Myeloma, NCT02575144 (Phase 3; Active, not recruiting)—Multiple Myeloma, NCT02516696 (Phase 3; Active, not recruiting)—Multiple Myeloma, NCT04287660 (Phase 3; Recruiting)—Multiple Myeloma |
| Clopidogrel | ABCB1 | N | N | Y | 1 | Phase 1 | Phase 1 (1) | Recruiting (1) | Head and Neck (1) | NA |
| Colchicine | ABCB1, TUBB | N | N | Y | 2 | Phase 2 | Phase 1 (1), Phase 2 (1) | Recruiting (2) | Urological, Multiple cancer types (1), GI (1) | NA |
| Cyclosporine | ABCC1, ABCB1 | N | N | Y | 2 | Phase 2 | Phase 1 (1), Phase 2 (1) | Recruiting (1), Active, not recruiting (1) | Leukaemia (1), Other Haem-onc (1) | NA |
| Deferoxamine | ABCB1 | N | N | Y | 4 | Phase 3 | Phase 1 (2), Phase 2 (1), Phase 3 (1) | Recruiting (3), Not yet recruiting (1) | Multiple cancer types (1), Breast (1), Leukaemia (1), GI (1) | IRCT20200313046756N2 (Phase 3; Pending)—Acute Myeloid Leukaemia |
| Digoxin | ABCB1 | N | Y | Y | 2 | Phase 2 | Phase 1 (1), Phase 2 (1) | Recruiting (2) | GI (1), Multiple cancer types (1) | NA |
| Disulfiram | ABCB1 | N | N | Y | 13 | Phase 2/3 | Phase 1 (4), Phase 1/2 (2), Phase 2 (6), Phase 2/3 (1) | Recruiting (7), Active, not recruiting (2), Ongoing (3), Not yet recruiting (1) | Multiple cancer types, GI (1), Soft-Tissue Sarcoma, Bone Sarcoma (1), CNS (3), Other Haem-onc (1), Urological (2), Multiple cancer types, Breast (1), GI (2), Breast (2) | NA |
| Doxycycline | ABCB1 | N | Y | Y | 10 | Phase 2 | Phase 1 (1), Phase 2 (9) | Recruiting (4), Active, not recruiting (4), Ongoing (1), Not yet recruiting (1) | Urological (1), Lymphoma (3), Multiple cancer types (1), Breast, Gynaecological (1), Breast (1), GI (2), Head and Neck (1) | NA |
| Fenofibrate | ABCB1, NR1I2 | N | Y | Y | 1 | Phase 2 | Phase 2 (1) | Recruiting (1) | CNS (1) | NA |
| Fostamatinib | * | N | N | N | 2 | Phase 1 | Phase 1 (2) | Recruiting (2) | Gynaecological (1), Other Haem-onc, Leukaemia (1) | NA |
| Indomethacin | ABCC1, PTGS2 | N | Y | Y | 5 | Phase 4 | Phase 1 (1), Phase 1/2 (1), Phase 2 (1), Phase 3 (1), Phase 4 (1) | Recruiting (2), Active, not recruiting (3) | Urological (2), Breast (1), Head and Neck (2) | ChiCTR2000038968 (Phase 4; Recruiting)—Prostate, NCT01265849 (Phase 3; Active, not recruiting)—Cancer of the oral cavity |
| Itraconazole | ABCB1, ERBB2 | N | Y | Y | 15 | Phase 3 | Phase 1 (4), Phase 1/2 (2), Phase 2 (7), Phase 3 (1), Other (1) | Recruiting (9), Active, not recruiting (2), Completed (1), Not yet recruiting (1), Other (1), Suspended (1) | Leukaemia, Other Haem-onc (1), Lung (2), Multiple cancer types, Lymphoma, Leukaemia (1), follow-up continuing (1), Multiple cancer types (2), GI (4), Urological (1), Gynaecological (2), Skin (1) | NCT03458221 (Phase 3; Not yet recruiting)—Ovarian |
| Ivermectin | ABCB1 | N | Y | Y | 2 | Phase 2 | Phase 2 (2) | Recruiting (1), Not yet recruiting (1) | Breast (1), Multiple cancer types (1) | NA |
| Ketoconazole | ABCB1, NR1I3, NR1I2, AR | N | Y | Y | 4 | Phase 2 | Phase 1 (2), Phase 2 (2) | Recruiting (1), Active, not recruiting (2), Ongoing (1) | Urological (2), Breast, CNS (1), CNS (1) | NA |
| Lansoprazole | ABCB1, MAPT | N | Y | Y | 3 | Phase 3 | Phase 2 (1), Phase 3 (2) | Recruiting (2), Active, not recruiting (1) | Breast (1), Lymphoma (1), GI (1) | NCT04874935 (Phase 3; Recruiting)—Breast, NCT03647072 (Phase 3; Recruiting)—Non-Hodgkin Lymphoma |
| Levetiracetam | ABCB1 | N | Y | Y | 2 | Phase 2 | Phase 2 (1), Other (1) | Not yet recruiting (2) | CNS (2) | NA |
| Losartan | ABCB1 | N | N | Y | 12 | Phase 3 | Phase 1 (4), Phase 2 (7), Phase 3 (1) | Recruiting (9), Active, not recruiting (1), Not yet recruiting (2) | Multiple cancer types (3), Bone Sarcoma (1), GI (7), Breast (1) | CTRI/2021/05/033482 (Phase 3; Not Yet Recruiting)—Pancreatic |
| Lovastatin | ABCB1, HDAC2, HMGCR | N | N | Y | 1 | Other | Other (1) | Not yet recruiting (1) | GI (1) | NA |
| Maprotiline | ABCB1 | N | N | Y | 1 | Phase 1 | Phase 1 (1) | Not yet recruiting (1) | CNS (1) | NA |
| Mefloquine | ABCB1 | N | Y | Y | 1 | Phase 1 | Phase 1 (1) | Active, not recruiting (1) | CNS (1) | NA |
| Miconazole | ABCB1, NR1I2 | N | Y | Y | 2 | Phase 2 | Phase 1 (1), Phase 2 (1) | Recruiting (2) | Multiple cancer types (2) | NA |
| Midazolam | GABRB3, ABCB1 | N | Y | Y | 2 | Phase 2 | Phase 2 (2) | Recruiting (1), Not yet recruiting (1) | Urological (2) | NA |
| Mifepristone | ABCB1, NR1I2, KLK3, NR3C1, PGR | N | N | Y | 2 | Phase 3 | Phase 2 (1), Phase 3 (1) | Active, not recruiting (1), Not yet recruiting (1) | Breast (2) | NCT05016349 (Phase 3; Not yet recruiting)—Breast |
| Miltefosine | ABCB1 | N | N | Y | 2 | Phase 2 | Phase 2 (2) | Recruiting (1), Active, not recruiting (1) | Breast (1), Multiple cancer types (1) | NA |
| Nelfinavir | ABCB1 | N | N | Y | 9 | Phase 3 | Phase 1 (2), Phase 1/2 (2), Phase 2 (3), Phase 3 (2) | Recruiting (5), Active, not recruiting (2), Ongoing (2) | Soft Tissue Sarcoma (1), Urological, Multiple cancer types (1), Gynaecological (3), Head and Neck (1), Other Haem-onc (1), GI (1), CNS (1) | NCT03256916 (Phase 3; Recruiting)—Advanced carcinoma of the cervix, CTRI/2017/08/009265 (Phase 3; Open to Recruitment)—Advanced carcinoma of the cervix |
| Nicardipine | ABCB1 | N | N | Y | 1 | Other | Other (1) | Not yet recruiting (1) | CNS (1) | NA |
| Omeprazole | ABCB1, AHR | N | Y | Y | 3 | Phase 2 | Phase 1 (2), Phase 2 (1) | Recruiting (2), Active, not recruiting (1) | Urological (1), Breast (1), GI (1) | NA |
| Pantoprazole | ABCB1 | N | N | Y | 2 | Phase 2 | Phase 1/2 (1), Phase 2 (1) | Active, not recruiting (1), Not yet recruiting (1) | Head and Neck (1), Urological (1) | NA |
| Phenytoin | POU5F1, SCN8A, SCN2A, SCN3A, NR1I2, SCN1A, SCN5A | N | Y | Y | 1 | Phase 2 | Phase 2 (1) | Recruiting (1) | Bone Sarcoma, Soft-Tissue Sarcoma (1) | NA |
| Pravastatin | ABCB1, HDAC2, HMGCR | N | N | Y | 2 | Phase 4 | Phase 2 (1), Phase 4 (1) | Active, not recruiting (1), Not yet recruiting (1) | Leukaemia (1), Breast (1) | ChiCTR2000034035 (Phase 4; Pending)—Breast |
| Propofol | GABRB3, ABCB1, SCN2A, SCN4A | N | Y | Y | 29 | Phase 4 | Phase 2 (1), Phase 3 (2), Phase 4 (7), Other (19) | Recruiting (17), Active, not recruiting (2), Ongoing (1), Not yet recruiting (9) | GI (6), Breast, GI (1), Multiple cancer types (7), Breast (5), Urological (3), Lung (6), CNS (1) | NCT01975064 (Phase 4; Recruiting)—Breast, colon, rectal, NCT05331911 (Phase 4; Not yet recruiting)—Liver, NCT04475705 (Phase 4; Recruiting)—Paediatric solid tumours, NCT05141877 (Phase 4; Not yet recruiting)—Brain tumour, NCT03034096 (Phase 4; Recruiting)—Adult cancer, NCT04513808 (Phase 3; Recruiting)—Oesophageal caner, ChiCTR2000040604 (Phase 4; Pending)—Non-small cell lung cancer, ACTRN12611000301965 (Phase 4; Not yet recruiting)—Breast, 2009-009114-40 (Phase 3; Ongoing)—Prostate |
| Propranolol | ABCB1, EGFR, ADRB3 | N | N | Y | 19 | Phase 3 | Phase 1 (3), Phase 1/2 (1), Phase 2 (13), Phase 2/3 (1), Phase 3 (1) | Recruiting (10), Active, not recruiting (1), Not yet recruiting (7), Other (1) | GI (8), Urological (2), Soft Tissue Sarcoma (2), Gynaecological (1), Skin (3), Other (1), Multiple cancer types (2) | CTRI/2019/11/021924 (Phase 3; Not Yet Recruiting)—Ovarian |
| Rifampicin | ABCB1, NR1I2 | N | Y | Y | 1 | Phase 1 | Phase 1 (1) | Recruiting (1) | Multiple cancer types, Lymphoma, Leukaemia (1) | NA |
| Ritonavir | ABCC1, ABCB1, NR1I2 | N | Y | Y | 1 | Phase 1 | Phase 1 (1) | Recruiting (1) | Leukaemia, Lymphoma (1) | NA |
| Sertraline | ABCB1, SLC29A4, SLC6A3 | N | Y | Y | 1 | Phase 1 | Phase 1 (1) | Recruiting (1) | Leukaemia (1) | NA |
| Simvastatin | ABCB1, HDAC2, HMGCR | N | Y | Y | 18 | Phase 4 | Phase 1 (4), Phase 2 (11), Phase 3 (1), Phase 4 (2) | Recruiting (7), Active, not recruiting (3), Ongoing (3), Not yet recruiting (4), Suspended (1) | Breast (5), Other Haem-onc (1), Lung (3), Gynaecological (1), GI (6), Multiple cancer types (2) | ChiCTR2000034035 (Phase 4; Pending)—Breast, 2010-018491-24 (Phase 4; Ongoing)—Adults with bone metastasis, NCT03971019 (Phase 3; Recruiting)—Breast |
| Sirolimus | ABCB1, MTOR | N | Y | Y | 28 | Phase 4 | Phase 1 (7), Phase 1/2 (4), Phase 2 (13), Phase 3 (2), Phase 4 (1), Other (1) | Recruiting (17), Active, not recruiting (7), Ongoing (3), Not yet recruiting (1) | Lung (2), Multiple cancer types, Lung (1), Multiple cancer types (6), Other (2), Soft-Tissue Sarcoma (3), Urological, Multiple cancer types (1), Bone Sarcoma, Soft Tissue Sarcoma (1), Gynaecological (1), Leukaemia (3), Endocrine (1), GI (3), CNS (1), Bone Sarcoma (2), Breast (1) | NCT04775173 (Phase 3; Recruiting)—Haemangioendothelioma, ChiCTR1900021896 (Phase 4; Recruiting)—Liver, NCT04736589 (Phase 3; Not yet recruiting)—Breast |
| Valganciclovir | ABCB1 | N | Y | Y | 5 | Phase 2 | Phase 1/2 (2), Phase 2 (2), Other (1) | Recruiting (4), Not yet recruiting (1) | GI (1), Lymphoma (1), Head and Neck (1), CNS (1), Multiple cancer types (1) | NA |
| Verapamil | ABCC1, ABCB1 | N | N | Y | 1 | Phase 1 | Phase 1 (1) | Active, not recruiting (1) | Lymphoma (1) | NA |
| Warfarin | ABCB1, AXL, NR1I2 | N | N | Y | 1 | Phase 1 | Phase 1 (1) | Recruiting (1) | GI (1) | NA |
| Zidovudine | ABCC1, ABCB1, TERT | N | Y | Y | 1 | Phase 2 | Phase 2 (1) | Recruiting (1) | Leukaemia (1) | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roundhill, E.A.; Pantziarka, P.; Liddle, D.E.; Shaw, L.A.; Albadrani, G.; Burchill, S.A. Exploiting the Stemness and Chemoresistance Transcriptome of Ewing Sarcoma to Identify Candidate Therapeutic Targets and Drug-Repurposing Candidates. Cancers 2023, 15, 769. https://doi.org/10.3390/cancers15030769
Roundhill EA, Pantziarka P, Liddle DE, Shaw LA, Albadrani G, Burchill SA. Exploiting the Stemness and Chemoresistance Transcriptome of Ewing Sarcoma to Identify Candidate Therapeutic Targets and Drug-Repurposing Candidates. Cancers. 2023; 15(3):769. https://doi.org/10.3390/cancers15030769
Chicago/Turabian StyleRoundhill, Elizabeth Ann, Pan Pantziarka, Danielle E. Liddle, Lucy A. Shaw, Ghadeer Albadrani, and Susan Ann Burchill. 2023. "Exploiting the Stemness and Chemoresistance Transcriptome of Ewing Sarcoma to Identify Candidate Therapeutic Targets and Drug-Repurposing Candidates" Cancers 15, no. 3: 769. https://doi.org/10.3390/cancers15030769
APA StyleRoundhill, E. A., Pantziarka, P., Liddle, D. E., Shaw, L. A., Albadrani, G., & Burchill, S. A. (2023). Exploiting the Stemness and Chemoresistance Transcriptome of Ewing Sarcoma to Identify Candidate Therapeutic Targets and Drug-Repurposing Candidates. Cancers, 15(3), 769. https://doi.org/10.3390/cancers15030769