
Integrins Can Act as Suppressors of Ras-Mediated Oncogenesis in the Drosophila Wing Disc Epithelium
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Strains
2.2. Immunocytochemistry and Imaging
2.3. Quantification of Fluorescence Intensity
3. Results
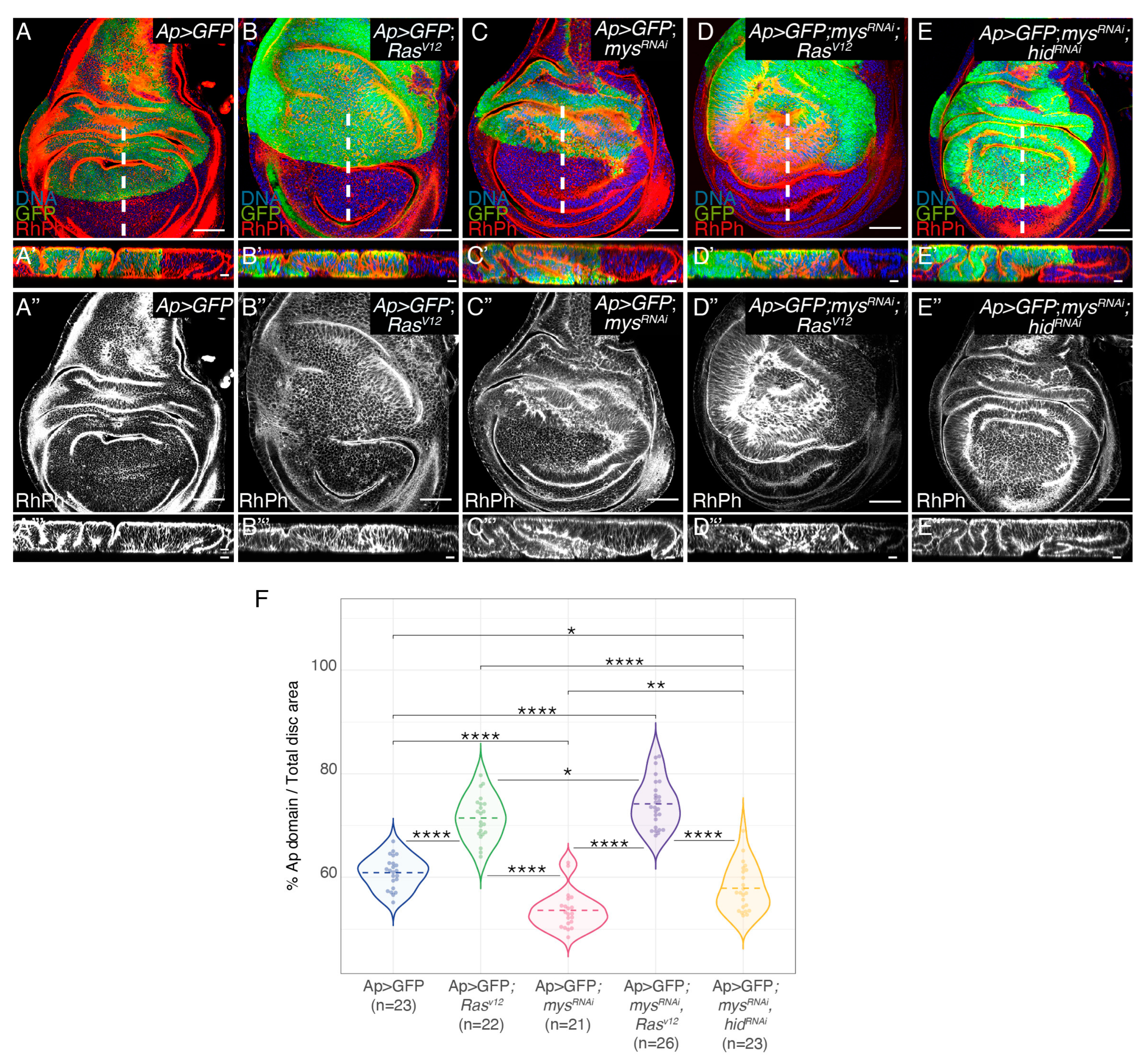
3.1. Integrin Knockdown Enhances RasV12 Hyperplastic Phenotype
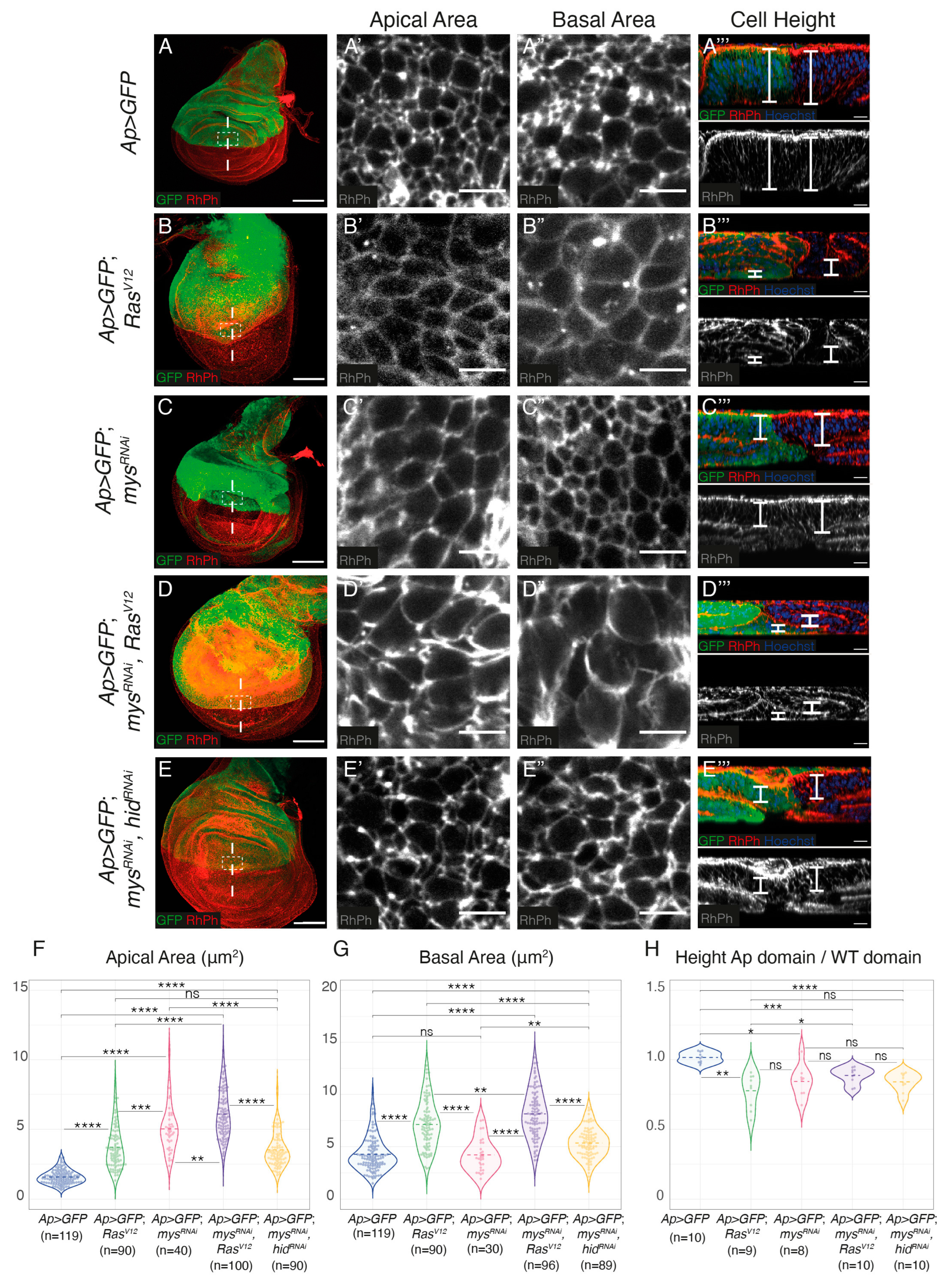
3.2. Integrin Knockdown Enhances RasV12-Associated Cell Shape Changes and Growth
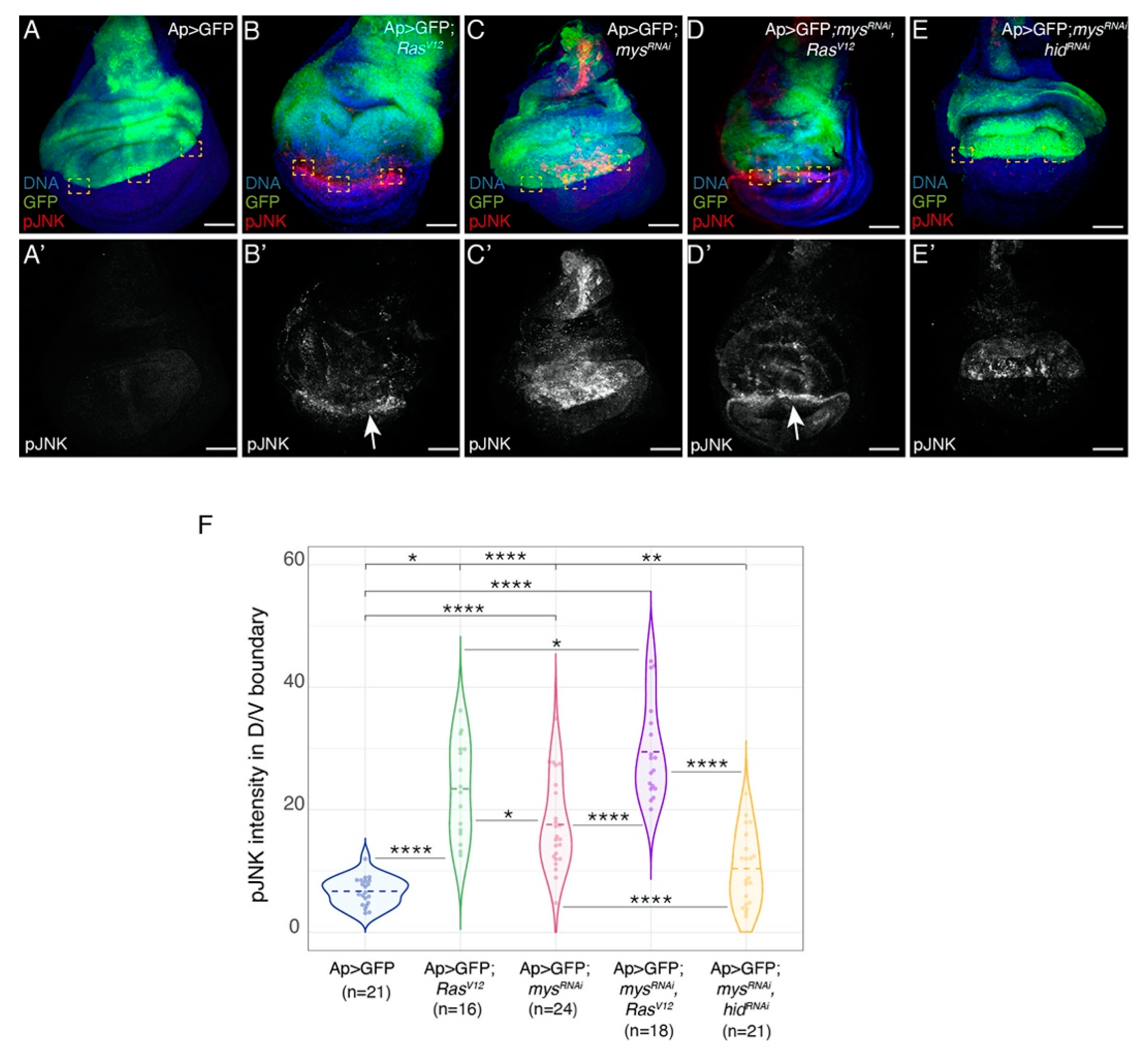
3.3. Integrins Restrict the Ability of RasV12 Tumour Cells to Stimulate Apoptosis in Neighbouring Wild-Type Cells
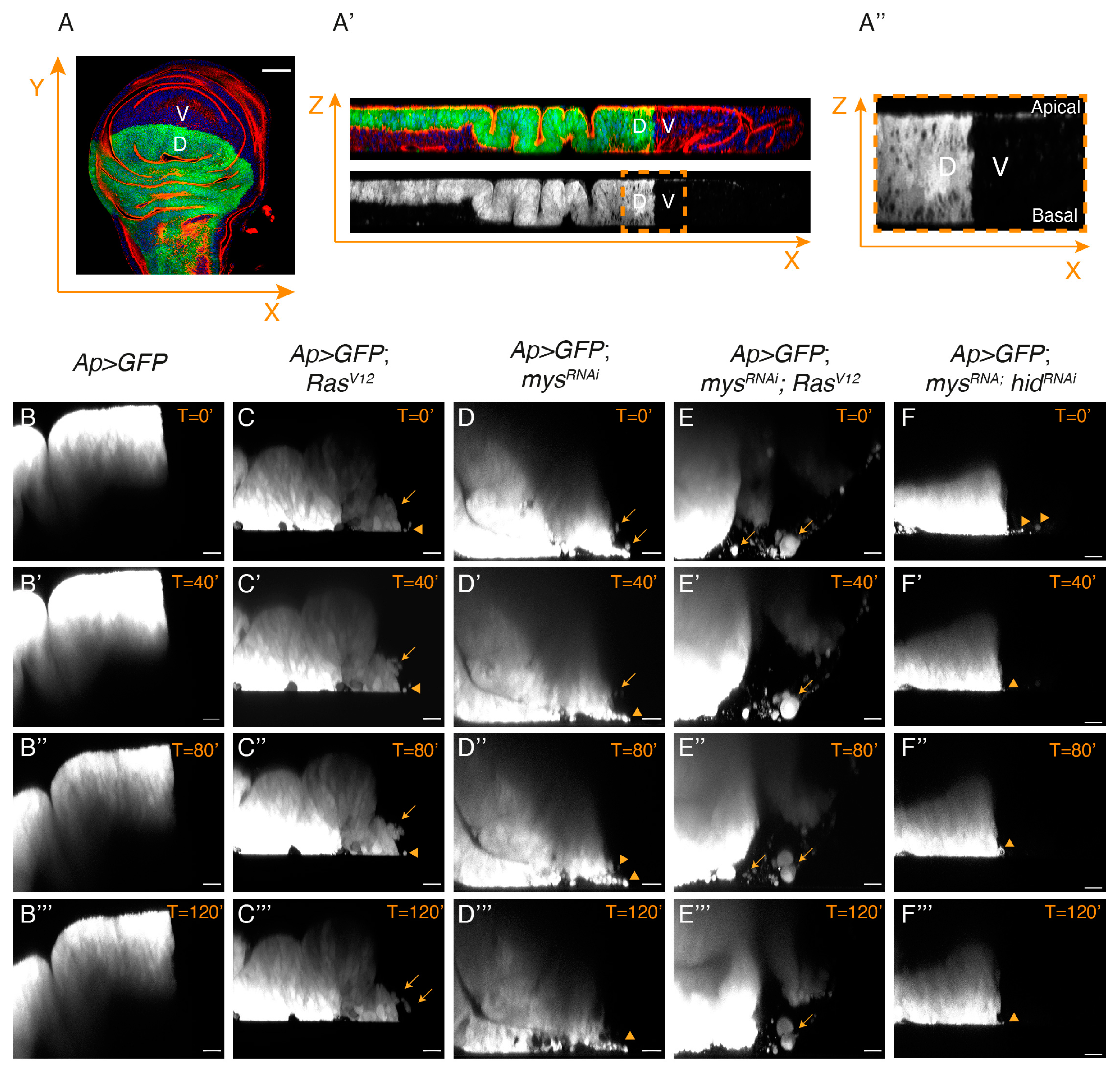
3.4. Integrins Limit the Ability of Epithelial RasV12 Tumour Cells to Leave the Epithelium and Move
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- Hamidi, H.; Ivaska, J. Every step of the way: Integrins in cancer progression and metastasis. Nat. Rev. Cancer 2018, 18, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119, 3901–3903. [Google Scholar] [CrossRef]
- Cagnet, S.; Faraldo, M.M.; Kreft, M.; Sonnenberg, A.; Raymond, K.; Glukhova, M.A. Signaling events mediated by a3b1 integrin are essential for mammary tumorigenesis. Oncogene 2014, 33, 4286–4295. [Google Scholar] [CrossRef]
- White, D.E.; Kurpios, N.A.; Zuo, D.; Hassell, J.A.; Blaess, S.; Mueller, U.; Muller, W.J. Targeted disruption of beta1-integrin in a transgenic mouse model of human breast cancer reveals an essential role in mammary tumor induction. Cancer Cell 2004, 6, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, N.E.; Zhang, Z.; Madamanchi, A.; Boyd, K.L.; O’Rear, L.D.; Nashabi, A.; Li, Z.; Dupont, W.D.; Zijlstra, A.; Zutter, M.M. The alpha(2)beta(1) integrin is a metastasis suppressor in mouse models and human cancer. J. Clin. Investig. 2011, 121, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Truong, H.H.; Xiong, J.; Ghotra, V.P.; Nirmala, E.; Haazen, L.; Le Devedec, S.E.; Balcioglu, H.E.; He, S.; Snaar-Jagalska, B.E.; Vreugdenhil, E.; et al. beta1 integrin inhibition elicits a prometastatic switch through the TGFbeta-miR-200-ZEB network in E-cadherin-positive triple-negative breast cancer. Sci. Signal. 2014, 7, ra15. [Google Scholar] [CrossRef]
- Brown, N.; Gregory, S.L.; Martin-Bermudo, M.D. Integrins as mediators of morphogenesis in Drosophila. Dev. Biol. 2000, 223, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Brower, D.L. Platelets with wings:the maturation of Drosophila integrin biology. Curr. Opin. Cell Biol. 2003, 15, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Bokel, C.; Brown, N.H. Integrins in Development: Moving on, respoding to, and sticking to the extracellular matrix. Dev. Cell 2002, 3, 311–321. [Google Scholar] [CrossRef]
- Fernández-Miñán, A.; Martín-Bermudo, M.D.; González-Reyes, A. Integrin signaling regulates spindle orientation in Drosophila to preserve the follicular-epithelium monolayer. Curr. Biol. 2007, 17, 683–688. [Google Scholar] [CrossRef]
- Lovegrove, H.E.; Bergstralh, D.T.; St Johnston, D. The role of integrins in Drosophila egg chamber morphogenesis. Development 2019, 146, dev182774. [Google Scholar] [CrossRef] [PubMed]
- Santa-Cruz Mateos, C.; Valencia-Exposito, A.; Palacios, I.M.; Martin-Bermudo, M.D. Integrins regulate epithelial cell shape by controlling the architecture and mechanical properties of basal actomyosin networks. PLoS Genet. 2020, 16, e1008717. [Google Scholar] [CrossRef]
- Valencia-Exposito, A.; Gomez-Lamarca, M.J.; Widmann, T.J.; Martin-Bermudo, M.D. Integrins Cooperate With the EGFR/Ras Pathway to Preserve Epithelia Survival and Architecture in Development and Oncogenesis. Front. Cell Dev. Biol. 2022, 10, 892691. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, B.K.; Irvine, K.D. The wing imaginal disc. Genetics 2022, 220, iyac020. [Google Scholar] [CrossRef] [PubMed]
- Brower, D.L.; Jaffe, S.M. Requirements for integrins during Drosophila wing development. Nature 1989, 342, 285–287. [Google Scholar] [CrossRef]
- Wilcox, M.; DiAntonio, A.; Leptin, M. The function of PS integrins in Drosophila wing morphogenesis. Development 1989, 107, 891–897. [Google Scholar] [CrossRef]
- Dominguez-Gimenez, P.; Brown, N.H.; Martin-Bermudo, M.D. Integrin-ECM interactions regulate the changes in cell shape driving the morphogenesis of the Drosophila wing epithelium. J. Cell Sci. 2007, 120, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C. Drosophila melanogaster: A model and a tool to investigate malignancy and identify new therapeutics. Nat. Rev. Cancer 2013, 13, 172–183. [Google Scholar] [CrossRef]
- Mirzoyan, Z.; Sollazzo, M.; Allocca, M.; Valenza, A.M.; Grifoni, D.; Bellosta, P. Drosophila melanogaster: A Model Organism to Study Cancer. Front. Genet. 2019, 10, 51. [Google Scholar] [CrossRef]
- Forbes, S.A.; Beare, D.; Gunasekaran, P.; Leung, K.; Bindal, N.; Boutselakis, H.; Ding, M.; Bamford, S.; Cole, C.; Ward, S.; et al. COSMIC: Exploring the world’s knowledge of somatic mutations in human cancer. Nucleic Acids Res. 2015, 43, D805–D811. [Google Scholar] [CrossRef]
- Lee, T.; Feig, L.; Montell, D.J. Two distinct roles for Ras in a developmentally regulated cell migration. Development 1996, 122, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Villa-Fombuena, G.; Lobo-Pecellin, M.; Marin-Menguiano, M.; Rojas-Rios, P.; Gonzalez-Reyes, A. Live imaging of the Drosophila ovarian niche shows spectrosome and centrosome dynamics during asymmetric germline stem cell division. Development 2021, 148, dev199716. [Google Scholar] [CrossRef]
- Prober, D.A.; Edgar, B.A. Ras1 promotes cellular growth in the Drosophila wing. Cell 2000, 100, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Soler Beatty, J.; Molnar, C.; Luque, C.M.; de Celis, J.F.; Martin-Bermudo, M.D. EGFRAP encodes a new negative regulator of the EGFR acting in both normal and oncogenic EGFR/Ras-driven tissue morphogenesis. PLoS Genet. 2021, 17, e1009738. [Google Scholar] [CrossRef]
- Genevet, A.; Polesello, C.; Blight, K.; Robertson, F.; Collinson, L.M.; Pichaud, F.; Tapon, N. The Hippo pathway regulates apical-domain size independently of its growth-control function. J. Cell Sci. 2009, 122, 2360–2370. [Google Scholar] [CrossRef] [PubMed]
- Brumby, A.M.; Richardson, H.E. scribble mutants cooperate with oncogenic Ras or Notch to cause neoplastic overgrowth in Drosophila. EMBO J. 2003, 22, 5769–5779. [Google Scholar] [CrossRef]
- Lee, J.L.; Streuli, C.H. Integrins and epithelial cell polarity. J. Cell Sci. 2014, 127, 3217–3225. [Google Scholar] [CrossRef] [PubMed]
- Goulas, S.; Conder, R.; Knoblich, J.A. The Par complex and integrins direct asymmetric cell division in adult intestinal stem cells. Cell Stem Cell 2012, 11, 529–540. [Google Scholar] [CrossRef]
- Dillard, C.; Reis, J.G.T.; Rusten, T.E. Ras(V12); scrib(-/-) Tumors: A Cooperative Oncogenesis Model Fueled by Tumor/Host Interactions. Int. J. Mol. Sci. 2021, 22, 8873. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, Y.; Louani, A.; Dragu, A.; Sanchez-Sanchez, B.J.; Serna-Morales, E.; Yolland, L.; Gyoergy, A.; Vizcay, G.; Fleck, R.A.; Heddleston, J.M.; et al. A Moving Source of Matrix Components Is Essential for De Novo Basement Membrane Formation. Curr. Biol. 2017, 27, 3526–3534.e3524. [Google Scholar] [CrossRef]
- Karim, F.D.; Rubin, G.M. Ectopic expression of activated Ras1 induces hyperplastic growth and increased cell death in Drosophila imaginal tissues. Development 1998, 125, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Murcia, L.; Clemente-Ruiz, M.; Pierre-Elies, P.; Royou, A.; Milan, M. Selective Killing of RAS-Malignant Tissues by Exploiting Oncogene-Induced DNA Damage. Cell Rep. 2019, 28, 119–131.e114. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Ohsawa, S.; Igaki, T. Mitochondrial defects trigger proliferation of neighbouring cells via a senescence-associated secretory phenotype in Drosophila. Nat. Commun. 2014, 5, 5264. [Google Scholar] [CrossRef]
- Chabu, C.; Xu, T. Oncogenic Ras stimulates Eiger/TNF exocytosis to promote growth. Development 2014, 141, 4729–4739. [Google Scholar] [CrossRef]
- Perez-Garijo, A.; Fuchs, Y.; Steller, H. Apoptotic cells can induce non-autonomous apoptosis through the TNF pathway. Elife 2013, 2, e01004. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E.; Valon, L.; Levillayer, F.; Levayer, R. Competition for Space Induces Cell Elimination through Compaction-Driven ERK Downregulation. Curr. Biol. 2019, 29, 23–34.e28. [Google Scholar] [CrossRef]
- Ramovs, V.; Te Molder, L.; Sonnenberg, A. The opposing roles of laminin-binding integrins in cancer. Matrix Biol. 2017, 57–58, 213–243. [Google Scholar] [CrossRef]
- Su, C.Y.; Li, J.Q.; Zhang, L.L.; Wang, H.; Wang, F.H.; Tao, Y.W.; Wang, Y.Q.; Guo, Q.R.; Li, J.J.; Liu, Y.; et al. The Biological Functions and Clinical Applications of Integrins in Cancers. Front. Pharmacol. 2020, 11, 579068. [Google Scholar] [CrossRef]
- Li, Z.; Zheng, W.; Wang, H.; Cheng, Y.; Fang, Y.; Wu, F.; Sun, G.; Sun, G.; Lv, C.; Hui, B. Application of Animal Models in Cancer Research: Recent Progress and Future Prospects. Cancer Manag. Res. 2021, 13, 2455–2475. [Google Scholar] [CrossRef] [PubMed]
- Huck, L.; Pontier, S.M.; Zuo, D.M.; Muller, W.J. beta1-integrin is dispensable for the induction of ErbB2 mammary tumors but plays a critical role in the metastatic phase of tumor progression. Proc. Natl. Acad. Sci. USA 2010, 107, 15559–15564. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Barlow, B.; O’Rear, L.; Jarvis, B.; Li, Z.; Dickeson, K.; Dupont, W.; Zutter, M. Loss of the alpha2beta1 integrin alters human papilloma virus-induced squamous carcinoma progression in vivo and in vitro. PLoS ONE 2011, 6, e26858. [Google Scholar] [CrossRef]
- Kren, A.; Baeriswyl, V.; Lehembre, F.; Wunderlin, C.; Strittmatter, K.; Antoniadis, H.; Fassler, R.; Cavallaro, U.; Christofori, G. Increased tumor cell dissemination and cellular senescence in the absence of beta1-integrin function. EMBO J. 2007, 26, 2832–2842. [Google Scholar] [CrossRef] [PubMed]
- Moran-Jones, K.; Ledger, A.; Naylor, M.J. beta1 integrin deletion enhances progression of prostate cancer in the TRAMP mouse model. Sci. Rep. 2012, 2, 526. [Google Scholar] [CrossRef]
- Du, Q.; Wang, W.; Liu, T.; Shang, C.; Huang, J.; Liao, Y.; Qin, S.; Chen, Y.; Liu, P.; Liu, J.; et al. High Expression of Integrin alpha3 Predicts Poor Prognosis and Promotes Tumor Metastasis and Angiogenesis by Activating the c-Src/Extracellular Signal-Regulated Protein Kinase/Focal Adhesion Kinase Signaling Pathway in Cervical Cancer. Front. Oncol. 2020, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Stoletov, K.; Kato, H.; Zardouzian, E.; Kelber, J.; Yang, J.; Shattil, S.; Klemke, R. Visualizing extravasation dynamics of metastatic tumor cells. J. Cell Sci. 2010, 123, 2332–2341. [Google Scholar] [CrossRef]
- Tanaka, S.; Senda, N.; Iida, A.; Sehara-Fujisawa, A.; Ishii, T.; Sato, F.; Toi, M.; Itou, J. In silico analysis-based identification of the target residue of integrin alpha6 for metastasis inhibition of basal-like breast cancer. Genes Cells 2019, 24, 596–607. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Zhang, X.; Ren, J.; Pang, Z.; Wang, C.; Xu, N.; Xi, R. Integrin signaling is required for maintenance and proliferation of intestinal stem cells in Drosophila. Dev. Biol. 2013, 377, 177–187. [Google Scholar] [CrossRef]
- Bajpai, A.; Quazi, T.A.; Tang, H.W.; Manzar, N.; Singh, V.; Thakur, A.; Ateeq, B.; Perrimon, N.; Sinha, P. A Drosophila model of oral peptide therapeutics for adult intestinal stem cell tumors. Dis. Model. Mech. 2020, 13, dmm044420. [Google Scholar] [CrossRef]
- Saxena, A.; Denholm, B.; Bunt, S.; Bischoff, M.; VijayRaghavan, K.; Skaer, H. Epidermal growth factor signalling controls myosin II planar polarity to orchestrate convergent extension movements during Drosophila tubulogenesis. PLoS Biol. 2014, 12, e1002013. [Google Scholar] [CrossRef]
- Tamada, M.; Zallen, J.A. Square Cell Packing in the Drosophila Embryo through Spatiotemporally Regulated EGF Receptor Signaling. Dev. Cell 2015, 35, 151–161. [Google Scholar] [CrossRef]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Bianconi, D.; Unseld, M.; Prager, G.W. Integrins in the Spotlight of Cancer. Int. J. Mol. Sci. 2016, 17, 2037. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, M.; Vaughen, J.; Igaki, T. Non-autonomous overgrowth by oncogenic niche cells: Cellular cooperation and competition in tumorigenesis. Cancer Sci. 2015, 106, 1651–1658. [Google Scholar] [CrossRef]
- Avraamides, C.J.; Garmy-Susini, B.; Varner, J.A. Integrins in angiogenesis and lymphangiogenesis. Nat. Rev. Cancer 2008, 8, 604–617. [Google Scholar] [CrossRef]
- Zutter, M.M.; Santoro, S.A.; Staatz, W.D.; Tsung, Y.L. Re-expression of the alpha 2 beta 1 integrin abrogates the malignant phenotype of breast carcinoma cells. Proc. Natl. Acad. Sci. USA 1995, 92, 7411–7415. [Google Scholar] [CrossRef] [PubMed]
- Varzavand, A.; Hacker, W.; Ma, D.; Gibson-Corley, K.; Hawayek, M.; Tayh, O.J.; Brown, J.A.; Henry, M.D.; Stipp, C.S. alpha3beta1 Integrin Suppresses Prostate Cancer Metastasis via Regulation of the Hippo Pathway. Cancer Res. 2016, 76, 6577–6587. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lewis, W.; Page-McCaw, A. Basement membrane mechanics shape development: Lessons from the fly. Matrix Biol. 2019, 75–76, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Xiao, Y.; Lin, H.P.; Yang, P.; Humphries, B.; Gao, T.; Yang, C. Integrin alpha9 depletion promotes beta-catenin degradation to suppress triple-negative breast cancer tumor growth and metastasis. Int. J. Cancer 2019, 145, 2767–2780. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Xing, X.; Cai, L.B.; Zhu, L.; Yang, X.M.; Wang, Y.H.; Yang, Q.; Nie, H.Z.; Zhang, Z.G.; Li, J.; et al. Integrin alpha9 Suppresses Hepatocellular Carcinoma Metastasis by Rho GTPase Signaling. J. Immunol. Res. 2018, 2018, 4602570. [Google Scholar] [CrossRef] [PubMed]
- Hegerfeldt, Y.; Tusch, M.; Brocker, E.B.; Friedl, P. Collective cell movement in primary melanoma explants: Plasticity of cell-cell interaction, beta1-integrin function, and migration strategies. Cancer Res. 2002, 62, 2125–2130. [Google Scholar] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Abarca Millán, A.; Martín-Bermudo, M.D. Integrins Can Act as Suppressors of Ras-Mediated Oncogenesis in the Drosophila Wing Disc Epithelium. Cancers 2023, 15, 5432. https://doi.org/10.3390/cancers15225432
Martínez-Abarca Millán A, Martín-Bermudo MD. Integrins Can Act as Suppressors of Ras-Mediated Oncogenesis in the Drosophila Wing Disc Epithelium. Cancers. 2023; 15(22):5432. https://doi.org/10.3390/cancers15225432
Chicago/Turabian StyleMartínez-Abarca Millán, Ana, and María D. Martín-Bermudo. 2023. "Integrins Can Act as Suppressors of Ras-Mediated Oncogenesis in the Drosophila Wing Disc Epithelium" Cancers 15, no. 22: 5432. https://doi.org/10.3390/cancers15225432
APA StyleMartínez-Abarca Millán, A., & Martín-Bermudo, M. D. (2023). Integrins Can Act as Suppressors of Ras-Mediated Oncogenesis in the Drosophila Wing Disc Epithelium. Cancers, 15(22), 5432. https://doi.org/10.3390/cancers15225432