

Characterization of an Mtbp Hypomorphic Allele in a Diethylnitrosamine-Induced Liver Carcinogenesis Model
 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
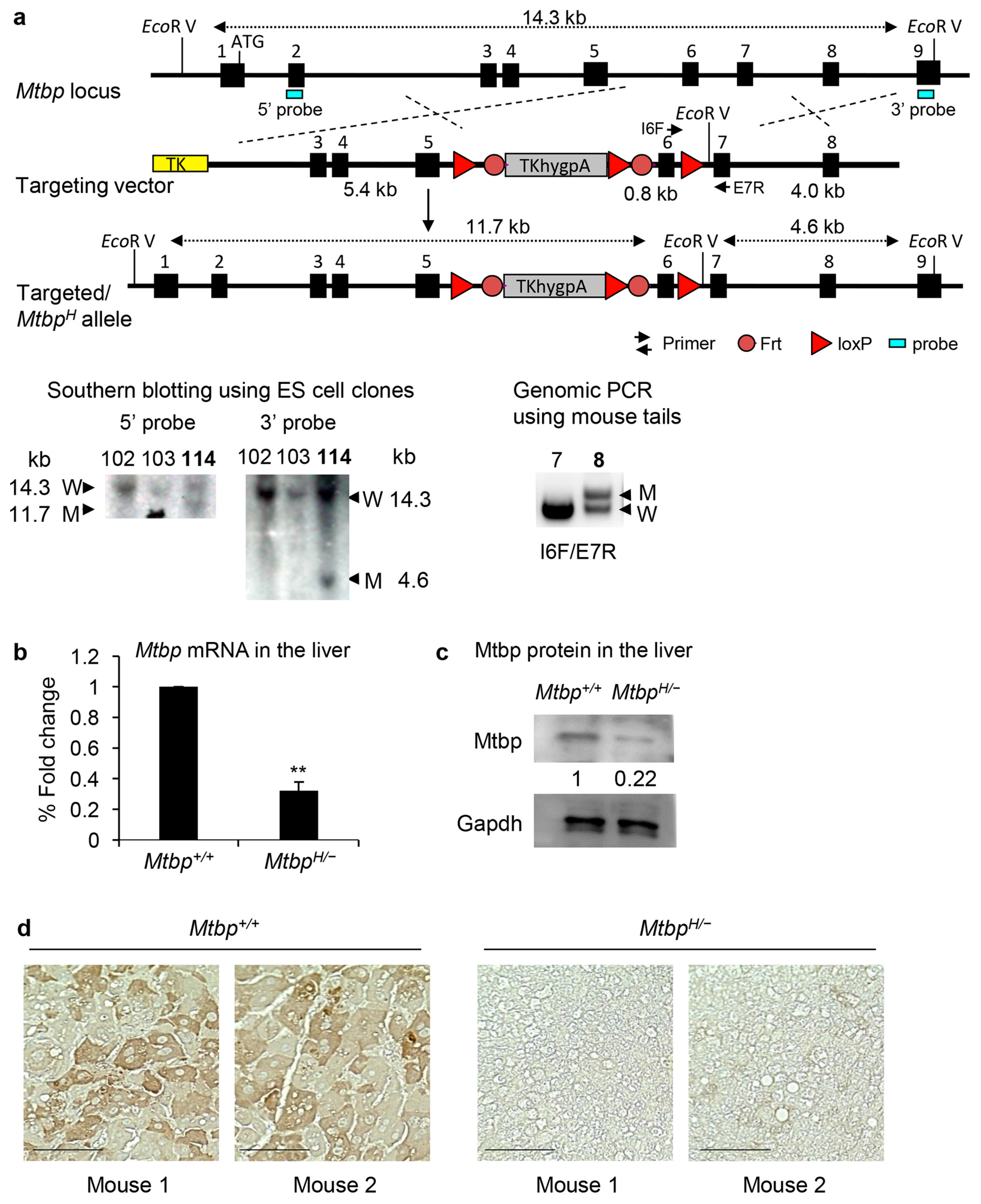
2.1. Generation of Mtbp Hypomorphic Mice
2.2. Genotyping
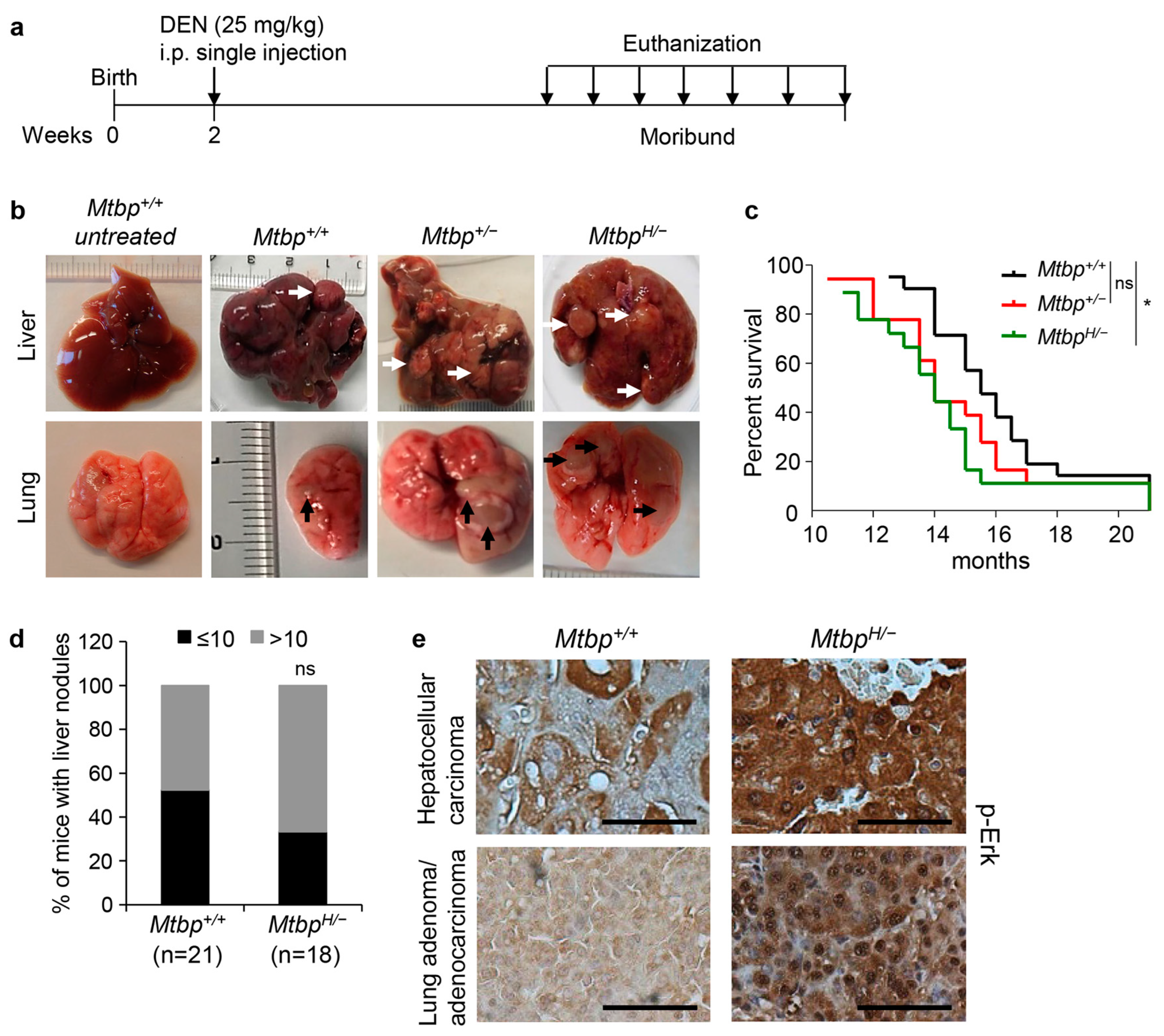
2.3. DEN-Induced Liver Carcinogenesis
2.4. Antibodies
2.5. Immunofluorescence
2.6. Quantitative Reverse Transcriptase PCR (qRT-PCR)
2.7. 3T3 Assay
2.8. Transwell Migration Assay
2.9. Immunohistochemistry (IHC)
2.10. Statistical Analysis
3. Results
3.1. Generation of Mtbp Hypomorphic (MtbpH/−) Mice
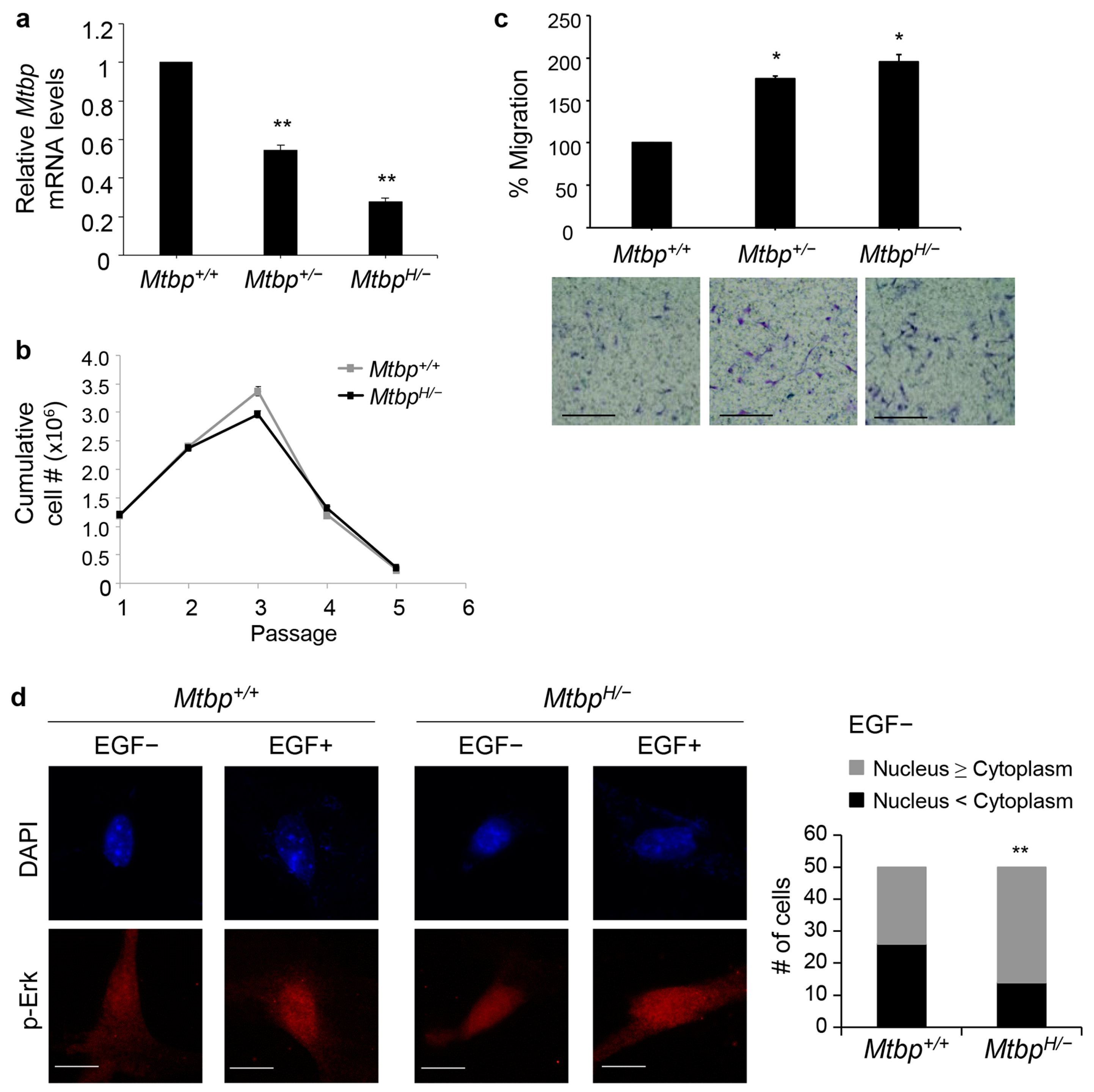
3.2. Increased Cell Migration of MtbpH/− MEFs
3.3. Reduced Mtbp Enhances Liver Carcinogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boyd, M.T.; Vlatkovic, N.; Haines, D.S. A novel cellular protein (MTBP) binds to MDM2 and induces a G1 arrest that is suppressed by MDM2. J. Biol. Chem. 2000, 275, 31883–31890. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, N.; Adhikari, A.S.; Iyer, S.V.; Hekmatdoost, K.; Welch, D.R.; Iwakuma, T. MTBP suppresses cell migration and filopodia formation by inhibiting ACTN4. Oncogene 2013, 32, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Iwakuma, T.; Agarwal, N. MDM2 binding protein, a novel metastasis suppressor. Cancer Metastasis Rev. 2012, 31, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Odvody, J.; Vincent, T.; Arrate, M.P.; Grieb, B.; Wang, S.; Garriga, J.; Lozano, G.; Iwakuma, T.; Haines, D.S.; Eischen, C.M. A deficiency in Mdm2 binding protein inhibits Myc-induced B-cell proliferation and lymphomagenesis. Oncogene 2010, 29, 3287–3296. [Google Scholar] [CrossRef] [PubMed]
- Boos, D.; Yekezare, M.; Diffley, J.F. Identification of a heteromeric complex that promotes DNA replication origin firing in human cells. Science 2013, 340, 981–984. [Google Scholar] [CrossRef] [PubMed]
- Grieb, B.C.; Chen, X.; Eischen, C.M. MTBP is overexpressed in triple-negative breast cancer and contributes to its growth and survival. Mol. Cancer Res. 2014, 12, 1216–1224. [Google Scholar] [CrossRef]
- Agarwal, N.; Tochigi, Y.; Adhikari, A.S.; Cui, S.; Cui, Y.; Iwakuma, T. MTBP plays a crucial role in mitotic progression and chromosome segregation. Cell Death Differ. 2011, 18, 1208–1219. [Google Scholar] [CrossRef][Green Version]
- Kohler, K.; Sanchez-Pulido, L.; Hofer, V.; Marko, A.; Ponting, C.P.; Snijders, A.P.; Feederle, R.; Schepers, A.; Boos, D. The Cdk8/19-cyclin C transcription regulator functions in genome replication through metazoan Sld7. PLoS Biol. 2019, 17, e2006767. [Google Scholar] [CrossRef] [PubMed]
- Vlatkovic, N.; El-Fert, A.; Devling, T.; Ray-Sinha, A.; Gore, D.M.; Rubbi, C.P.; Dodson, A.; Jones, A.S.; Helliwell, T.R.; Jones, T.M.; et al. Loss of MTBP expression is associated with reduced survival in a biomarker-defined subset of patients with squamous cell carcinoma of the head and neck. Cancer 2011, 117, 2939–2950. [Google Scholar] [CrossRef]
- Iwakuma, T.; Tochigi, Y.; Van Pelt, C.S.; Caldwell, L.C.; Terzian, T.; Parant, J.M.; Chau, G.P.; Koch, J.G.; Eischen, C.M.; Lozano, G. Mtbp haploinsufficiency in mice increases tumor metastasis. Oncogene 2008, 27, 1813–1820. [Google Scholar] [CrossRef][Green Version]
- Bi, Q.; Ranjan, A.; Fan, R.; Agarwal, N.; Welch, D.R.; Weinman, S.A.; Ding, J.; Iwakuma, T. MTBP inhibits migration and metastasis of hepatocellular carcinoma. Clin. Exp. Metastasis 2015, 32, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Iyer, S.V.; Ward, C.; Link, T.; Diaz, F.J.; Dhar, A.; Tawfik, O.W.; Weinman, S.A.; Azuma, Y.; Izumi, T.; et al. MTBP inhibits the Erk1/2-Elk-1 signaling in hepatocellular carcinoma. Oncotarget 2018, 9, 21429–21443. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, Z.; Jin, J.; Long, Z.; Liu, X.; Cai, H.; Zhou, Y.; Huang, H.; Wang, Y. MDM2 binding protein as a predictor of metastasis and a novel prognostic biomarker in patients with gastric cancer. Oncol. Lett. 2017, 14, 6409–6416. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, Y.; Sun, Y.; Zhao, X.; Sun, X.; Gong, T.; Liang, Z.; Ma, Y.; Zhang, X. Genome-wide analysis of lncRNAs, miRNAs, and mRNAs forming a prognostic scoring system in esophageal squamous cell carcinoma. PeerJ 2020, 8, e8368. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Tang, J.; Guo, Z.; Dai, Y.; Nie, J.; Hu, W.; Liu, N.; Ye, C.; Li, S.; Pei, H.; et al. Long Non-Coding RNA CRYBG3 Promotes Lung Cancer Metastasis via Activating the eEF1A1/MDM2/MTBP Axis. Int. J. Mol. Sci. 2021, 22, 3211. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Chen, M.; Yang, J.; Yang, C.; Lu, X.; Tian, H.; Liu, C. MTBP regulates migration and invasion of prostate cancer cells in vitro. Nan Fang Yi Ke Da Xue Xue Bao J. South. Med. Univ. 2019, 39, 6–12. [Google Scholar] [CrossRef]
- Pan, B.; Han, H.; Wu, L.; Xiong, Y.; Zhang, J.; Dong, B.; Yang, Y.; Chen, J. MTBP promotes migration and invasion by regulation of ZEB2-mediated epithelial-mesenchymal transition in lung cancer cells. Onco Targets Ther. 2018, 11, 6741–6756. [Google Scholar] [CrossRef]
- Wang, H.; Chu, F.; Zhijie, L.; Bi, Q.; Lixin, L.; Zhuang, Y.; Xiaofeng, Z.; Niu, X.; Zhang, D.; Xi, H.; et al. MTBP enhances the activation of transcription factor ETS-1 and promotes the proliferation of hepatocellular carcinoma cells. Front. Oncol. 2022, 12, 985082. [Google Scholar] [CrossRef]
- Shayimu, P.; Yusufu, A.; Rehemutula, A.; Redati, D.; Jiapaer, R.; Tuerdi, R. MTBP promoted the proliferation, migration and invasion of colon cancer cells by activating the expression of ZEB2. Anim. Cells Syst. 2021, 25, 152–160. [Google Scholar] [CrossRef]
- Grieb, B.C.; Boyd, K.; Mitra, R.; Eischen, C.M. Haploinsufficiency of the Myc regulator Mtbp extends survival and delays tumor development in aging mice. Aging 2016, 8, 2590–2602. [Google Scholar] [CrossRef]
- van Deursen, J.; Ruitenbeek, W.; Heerschap, A.; Jap, P.; ter Laak, H.; Wieringa, B. Creatine kinase (CK) in skeletal muscle energy metabolism: A study of mouse mutants with graded reduction in muscle CK expression. Proc. Natl. Acad. Sci. USA 1994, 91, 9091–9095. [Google Scholar] [CrossRef] [PubMed]
- Kalitsis, P.; Fowler, K.J.; Griffiths, B.; Earle, E.; Chow, C.W.; Jamsen, K.; Choo, K.H. Increased chromosome instability but not cancer predisposition in haploinsufficient Bub3 mice. Genes Chromosomes Cancer 2005, 44, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, T.; Fang, Y.; Xie, S.; Huang, X.; Mahmood, R.; Ramaswamy, G.; Sakamoto, K.M.; Darzynkiewicz, Z.; Xu, M.; et al. BUBR1 deficiency results in abnormal megakaryopoiesis. Blood 2004, 103, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Jeganathan, K.; Malureanu, L.; Baker, D.J.; Abraham, S.C.; van Deursen, J.M. Bub1 mediates cell death in response to chromosome missegregation and acts to suppress spontaneous tumorigenesis. J. Cell Biol. 2007, 179, 255–267. [Google Scholar] [CrossRef]
- Dawlaty, M.M.; van Deursen, J.M. Gene targeting methods for studying nuclear transport factors in mice. Methods 2006, 39, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Shawlot, W.; Deng, J.M.; Fohn, L.E.; Behringer, R.R. Restricted beta-galactosidase expression of a hygromycin-lacZ gene targeted to the beta-actin locus and embryonic lethality of beta-actin mutant mice. Transgenic Res. 1998, 7, 95–103. [Google Scholar] [CrossRef]
- Vesselinovitch, S.D.; Mihailovich, N. Kinetics of diethylnitrosamine hepatocarcinogenesis in the infant mouse. Cancer Res. 1983, 43, 4253–4259. [Google Scholar] [PubMed]
- Mervai, Z.; Egedi, K.; Kovalszky, I.; Baghy, K. Diethylnitrosamine induces lung adenocarcinoma in FVB/N mouse. BMC Cancer 2018, 18, 157. [Google Scholar] [CrossRef]
- Lantinga-van Leeuwen, I.S.; Dauwerse, J.G.; Baelde, H.J.; Leonhard, W.N.; van de Wal, A.; Ward, C.J.; Verbeek, S.; Deruiter, M.C.; Breuning, M.H.; de Heer, E.; et al. Lowering of Pkd1 expression is sufficient to cause polycystic kidney disease. Hum. Mol. Genet. 2004, 13, 3069–3077. [Google Scholar] [CrossRef]
- Mendrysa, S.M.; O’Leary, K.A.; McElwee, M.K.; Michalowski, J.; Eisenman, R.N.; Powell, D.A.; Perry, M.E. Tumor suppression and normal aging in mice with constitutively high p53 activity. Genes Dev. 2006, 20, 16–21. [Google Scholar] [CrossRef]
- Hartman, T.K.; Wengenack, T.M.; Poduslo, J.F.; van Deursen, J.M. Mutant mice with small amounts of BubR1 display accelerated age-related gliosis. Neurobiol. Aging 2007, 28, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Zonderland, G.; Vanzo, R.; Gadi, S.A.; Martin-Doncel, E.; Coscia, F.; Mund, A.; Lerdrup, M.; Benada, J.; Boos, D.; Toledo, L. The TRESLIN-MTBP complex couples completion of DNA replication with S/G2 transition. Mol. Cell 2022, 82, 3350–3365.e7. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, L.; Jiang, Y.; Hu, T.; Zhang, D.; Qiao, Q.; Wang, R.; Wang, M.; Han, S. MTBP regulates cell survival and therapeutic sensitivity in TP53 wildtype glioblastomas. Theranostics 2019, 9, 6019–6030. [Google Scholar] [CrossRef] [PubMed]
- Brady, M.; Vlatkovic, N.; Boyd, M.T. Regulation of p53 and MDM2 activity by MTBP. Mol. Cell Biol. 2005, 25, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Grieb, B.C.; Gramling, M.W.; Arrate, M.P.; Chen, X.; Beauparlant, S.L.; Haines, D.S.; Xiao, H.; Eischen, C.M. Oncogenic protein MTBP interacts with MYC to promote tumorigenesis. Cancer Res. 2014, 74, 3591–3602. [Google Scholar] [CrossRef]
- Grieb, B.C.; Eischen, C.M. MTBP and MYC: A Dynamic Duo in Proliferation, Cancer, and Aging. Biology 2022, 11, 881. [Google Scholar] [CrossRef]
- Metzger, J.; Nolte, A.; Uhde, A.K.; Hewicker-Trautwein, M.; Distl, O. Whole genome sequencing identifies missense mutation in MTBP in Shar-Pei affected with Autoinflammatory Disease (SPAID). BMC Genom. 2017, 18, 348. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genotype | Gender (Male: M, Female: F) | # of Mice | # of Mice with Lung Nodules | # of Mice with Metastatic Nodules |
|---|---|---|---|---|
| Mtbp+/+ | M | 12 | 5 | 0 |
| F | 9 | 3 | 0 | |
| Total | 21 | 8 (38.1%) | 0 | |
| MtbpH/− | M | 10 | 8 | 1 |
| F | 8 | 4 | 0 | |
| Total | 18 | 12 (66.7%) | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranjan, A.; Thoenen, E.A.; Kaida, A.; Wood, S.; Van Dyke, T.; Iwakuma, T. Characterization of an Mtbp Hypomorphic Allele in a Diethylnitrosamine-Induced Liver Carcinogenesis Model. Cancers 2023, 15, 4596. https://doi.org/10.3390/cancers15184596
Ranjan A, Thoenen EA, Kaida A, Wood S, Van Dyke T, Iwakuma T. Characterization of an Mtbp Hypomorphic Allele in a Diethylnitrosamine-Induced Liver Carcinogenesis Model. Cancers. 2023; 15(18):4596. https://doi.org/10.3390/cancers15184596
Chicago/Turabian StyleRanjan, Atul, Elizabeth A. Thoenen, Atsushi Kaida, Stephanie Wood, Terry Van Dyke, and Tomoo Iwakuma. 2023. "Characterization of an Mtbp Hypomorphic Allele in a Diethylnitrosamine-Induced Liver Carcinogenesis Model" Cancers 15, no. 18: 4596. https://doi.org/10.3390/cancers15184596
APA StyleRanjan, A., Thoenen, E. A., Kaida, A., Wood, S., Van Dyke, T., & Iwakuma, T. (2023). Characterization of an Mtbp Hypomorphic Allele in a Diethylnitrosamine-Induced Liver Carcinogenesis Model. Cancers, 15(18), 4596. https://doi.org/10.3390/cancers15184596