
Alternative Lengthening of Telomeres Is Rare in Canine Histiocytic Sarcoma

 ,
,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Spontaneous Histiocytic Sarcoma Tissue Samples and Clinical Data
2.2. Control Tissue Samples and Cell Lines
2.3. DNA Extraction and Quantification
2.4. Radiolabel C-Circle Assay and Telomeric Content
2.5. Telomere-Specific Fluorescence In Situ Hybridization (FISH)
2.6. Statistical Analysis
3. Results
3.1. Patient Characteristics and Clinical Data
3.2. ALT Activity in Canine Histiocytic Sarcoma Tissue and Controls
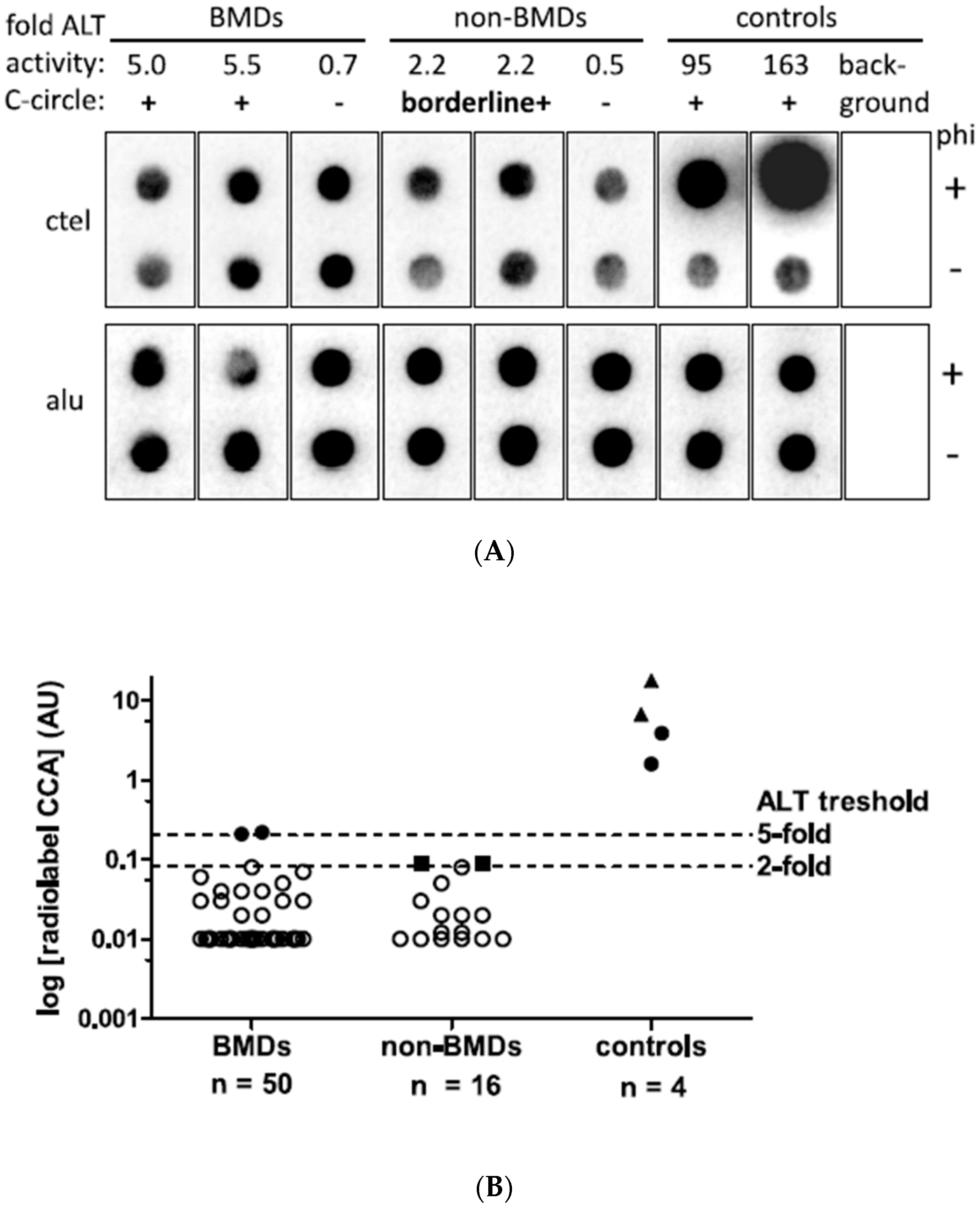
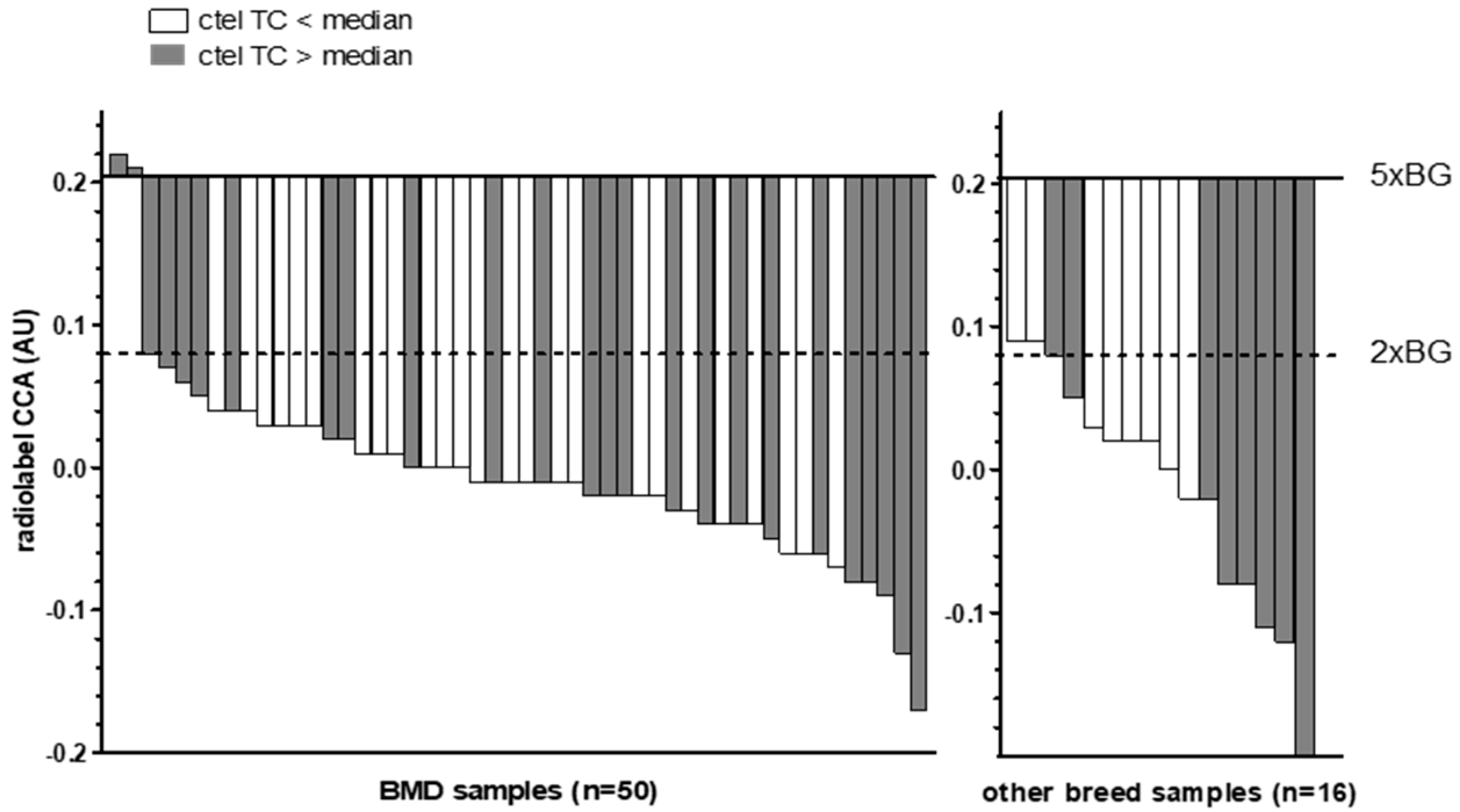
3.3. Telomeric Content in Canine Histiocytic Sarcoma Tissue and Controls
3.4. Evaluation of CCA Results Using Ultrabright and Giant Intranuclear Telomere Foci Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres Shorten during Ageing of Human Fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Nasir, L. Telomeres and Telomerase: Biological and Clinical Importance in Dogs. Vet. J. 2008, 175, 155–163. [Google Scholar] [CrossRef]
- Shay, J.W.; Bacchetti, S. A Survey of Telomerase Activity in Human Cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Zhang, J.-M.; Zou, L. Alternative Lengthening of Telomeres: From Molecular Mechanisms to Therapeutic Outlooks. Cell Biosci. 2020, 10, 30. [Google Scholar] [CrossRef]
- Sohn, E.J.; Goralsky, J.A.; Shay, J.W.; Min, J. The Molecular Mechanisms and Therapeutic Prospects of Alternative Lengthening of Telomeres (ALT). Cancers 2023, 15, 1945. [Google Scholar] [CrossRef]
- Gao, J.; Pickett, H.A. Targeting Telomeres: Advances in Telomere Maintenance Mechanism-Specific Cancer Therapies. Nat. Rev. Cancer 2022, 22, 515–532. [Google Scholar] [CrossRef]
- Hu, Y.; Shi, G.; Zhang, L.; Li, F.; Jiang, Y.; Jiang, S.; Ma, W.; Zhao, Y.; Songyang, Z.; Huang, J. Switch Telomerase to ALT Mechanism by Inducing Telomeric DNA Damages and Dysfunction of ATRX and DAXX. Sci. Rep. 2016, 6, 32280. [Google Scholar] [CrossRef]
- Gaspar, T.B.; Sá, A.; Lopes, J.M.; Sobrinho-Simões, M.; Soares, P.; Vinagre, J. Telomere Maintenance Mechanisms in Cancer. Genes 2018, 9, 241. [Google Scholar] [CrossRef]
- Claude, E.; Decottignies, A. Telomere Maintenance Mechanisms in Cancer: Telomerase, ALT or Lack Thereof. Curr. Opin. Genet. Dev. 2020, 60, 1–8. [Google Scholar] [CrossRef]
- Kreilmeier, T.; Sampl, S.; Deloria, A.J.; Walter, I.; Reifinger, M.; Hauck, M.; Borst, L.B.; Holzmann, K.; Kleiter, M. Alternative Lengthening of Telomeres Does Exist in Various Canine Sarcomas. Mol. Carcinog. 2017, 56, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Bicanova, L.; Kreilmeier-Berger, T.; Reifinger, M.; Holzmann, K.; Kleiter, M. Prevalence and Potentially Prognostic Value of C-Circles Associated with Alternative Lengthening of Telomeres in Canine Appendicular Osteosarcoma. Vet. Comp. Oncol. 2021, 19, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Henson, J.D.; Lau, L.M.; Koch, S.; Martin La Rotta, N.; Dagg, R.A.; Reddel, R.R. The C-Circle Assay for Alternative-Lengthening-of-Telomeres Activity. Methods 2017, 114, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Heaphy, C.M.; Subhawong, A.P.; Hong, S.-M.; Goggins, M.G.; Montgomery, E.A.; Gabrielson, E.; Netto, G.J.; Epstein, J.I.; Lotan, T.L.; Westra, W.H.; et al. Prevalence of the Alternative Lengthening of Telomeres Telomere Maintenance Mechanism in Human Cancer Subtypes. Am. J. Pathol. 2011, 179, 1608–1615. [Google Scholar] [CrossRef]
- Amorim, J.; Santos, G.; Vinagre, J.; Soares, P. The Role of ATRX in the Alternative Lengthening of Telomeres (ALT) Phenotype. Genes 2016, 7, 66. [Google Scholar] [CrossRef]
- Yasir, S.; Thompson, S.; Chen, Z.E.; Knudson, R.; Knutson, D.; Kloft-Nelson, S.; Graham, R.P.; Jain, D.; Simon, S.M.; Wu, T.-T.; et al. Alternative Lengthening of Telomeres in Primary Hepatic Neoplasms. Hum. Pathol. 2023, 131, 79–86. [Google Scholar] [CrossRef]
- Heaphy, C.M.; Zarinshenas, R.; Barber, J.R.; Davis, C.; Brosnan-Cashman, J.A.; De Marzo, A.M.; Joshu, C.E.; Platz, E.A.; Meeker, A.K. A High-Throughput Approach Measures Cell Type-Specific Telomere Lengths in Fixed Archival Tissues from Patient Cohorts for Research on Prognosis. medRxiv 2022. [Google Scholar] [CrossRef]
- da Costa, C.; Egeler, R.; Hoogeboom, M.; Szuhai, K.; Forsyth, R.; Niesters, M.; de Krijger, R.; Tazi, A.; Hogendoorn, P.; Annels, N. Differences in Telomerase Expression by the CD1a+ Cells in Langerhans Cell Histiocytosis Reflect the Diverse Clinical Presentation of the Disease. J. Pathol. 2007, 212, 188–197. [Google Scholar] [CrossRef]
- Dalia, S.; Shao, H.; Sagatys, E.; Cualing, H.; Sokol, L. Dendritic Cell and Histiocytic Neoplasms: Biology, Diagnosis, and Treatment. Cancer Control 2014, 21, 290–300. [Google Scholar] [CrossRef]
- Gounder, M.; Desai, V.; Kuk, D.; Agaram, N.; Arcila, M.; Durham, B.; Keohan, M.L.; Dickson, M.A.; D’Angelo, S.P.; Shukla, N.; et al. Impact of Surgery, Radiation and Systemic Therapy on the Outcomes of Patients with Dendritic Cell and Histiocytic Sarcomas. Eur. J. Cancer 2015, 51, 2413–2422. [Google Scholar] [CrossRef]
- Vos, J.A.; Abbondanzo, S.L.; Barekman, C.L.; Andriko, J.W.; Miettinen, M.; Aguilera, N.S. Histiocytic Sarcoma: A Study of Five Cases Including the Histiocyte Marker CD163. Mod. Pathol. 2005, 18, 693–704. [Google Scholar] [CrossRef]
- Craig, L.E.; Julian, M.E.; Ferracone, J.D. The Diagnosis and Prognosis of Synovial Tumors in Dogs: 35 Cases. Vet. Pathol. 2002, 39, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Dobson, J.; Hoather, T.; McKinley, T.J.; Wood, J.L.N. Mortality in a Cohort of Flat-Coated Retrievers in the UK. Vet. Comp. Oncol. 2009, 7, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Rosin, A.; Moore, P.; Dubielzig, R. Malignant Histiocytosis in Bernese Mountain Dogs. J. Am. Vet. Med. Assoc. 1986, 188, 1041–1045. [Google Scholar]
- Moore, P.F. A Review of Histiocytic Diseases of Dogs and Cats. Vet. Pathol. 2014, 51, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Abadie, J.; Hedan, B.; Cadieu, E.; De Brito, C.; Devauchelle, P.; Bourgain, C.; Parker, H.G.; Vaysse, A.; Margaritte-Jeannin, P.; Galibert, F.; et al. Epidemiology, Pathology, and Genetics of Histiocytic Sarcoma in the Bernese Mountain Dog Breed. J. Hered. 2009, 100, S19–S27. [Google Scholar] [CrossRef]
- Ruple, A.; Morley, P.S. Risk Factors Associated with Development of Histiocytic Sarcoma in Bernese Mountain Dogs. J. Vet. Intern. Med. 2016, 30, 1197–1203. [Google Scholar] [CrossRef]
- Dobson, J.M.; Samuel, S.; Milstein, H.; Rogers, K.; Wood, J.L.N. Canine Neoplasia in the UK: Estimates of Incidence Rates from a Population of Insured Dogs. J. Small Anim. Pract. 2002, 43, 240–246. [Google Scholar] [CrossRef]
- Fant, P.; Caldin, M.; Furlanello, T.; De Lorenzi, D.; Bertolini, G.; Bettini, G.; Morini, M.; Masserdotti, C. Primary Gastric Histiocytic Sarcoma in a Dog—A Case Report. J. Vet. Med. Ser. A 2004, 51, 358–362. [Google Scholar] [CrossRef]
- Hayden, D.W.; Waters, D.J.; Burke, B.A.; Manivel, J.C. Disseminated Malignant Histiocytosis in a Golden Retriever: Clinicopathologic, Ultrastructural, and Immunohistochemical Findings. Vet. Pathol. 1993, 30, 256–264. [Google Scholar] [CrossRef]
- Naranjo, C.; Dubielzig, R.R.; Friedrichs, K.R. Canine Ocular Histiocytic Sarcoma. Vet. Ophthalmol. 2007, 10, 179–185. [Google Scholar] [CrossRef]
- Skorupski, K.A.; Clifford, C.A.; Paoloni, M.C.; Lara-Garcia, A.; Barber, L.; Kent, M.S.; LeBlanc, A.K.; Sabhlok, A.; Mauldin, E.A.; Shofer, F.S.; et al. CCNU for the Treatment of Dogs with Histiocytic Sarcoma. J. Vet. Intern. Med. 2007, 21, 121–126. [Google Scholar] [CrossRef]
- Shortman, K.; Liu, Y.-J. Mouse and Human Dendritic Cell Subtypes. Nat. Rev. Immunol. 2002, 2, 151–161. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Dagg, R.; Zhang, Y.; Lee, J.H.Y.; Lu, R.; Martin La Rotta, N.; Sampl, S.; Korkut-Demirbaş, M.; Holzmann, K.; Lau, L.M.S.; et al. The C-Circle Biomarker Is Secreted by Alternative-Lengthening-of-Telomeres Positive Cancer Cells inside Exosomes and Provides a Blood-Based Diagnostic for ALT Activity. Cancers 2021, 13, 5369. [Google Scholar] [CrossRef]
- Cesare, A.J.; Heaphy, C.M.; O’Sullivan, R.J. Visualization of Telomere Integrity and Function In Vitro and In Vivo Using Immunofluorescence Techniques. Curr. Protoc. Cytom. 2015, 73, 12–40. [Google Scholar] [CrossRef]
- MacKenzie, D.; Watters, A.K.; To, J.T.; Young, M.W.; Muratori, J.; Wilkoff, M.H.; Abraham, R.G.; Plummer, M.M.; Zhang, D. ALT Positivity in Human Cancers: Prevalence and Clinical Insights. Cancers 2021, 13, 2384. [Google Scholar] [CrossRef]
- Henson, J.D.; Hannay, J.A.; McCarthy, S.W.; Royds, J.A.; Yeager, T.R.; Robinson, R.A.; Wharton, S.B.; Jellinek, D.A.; Arbuckle, S.M.; Yoo, J.; et al. A Robust Assay for Alternative Lengthening of Telomeres in Tumors Shows the Significance of Alternative Lengthening of Telomeres in Sarcomas and Astrocytomas. Clin. Cancer Res. 2005, 11, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Ulaner, G.A.; Huang, H.-Y.; Otero, J.; Zhao, Z.; Ben-Porat, L.; Satagopan, J.M.; Gorlick, R.; Meyers, P.; Healey, J.H.; Huvos, A.G.; et al. Absence of a Telomere Maintenance Mechanism as a Favorable Prognostic Factor in Patients with Osteosarcoma. Cancer Res. 2003, 63, 1759–1763. [Google Scholar] [PubMed]
- Matsuo, T.; Shimose, S.; Kubo, T.; Fujimori, J.; Yasunaga, Y.; Ochi, M. Telomeres and Telomerase in Sarcomas. Anticancer. Res. 2009, 29, 3833–3836. [Google Scholar] [PubMed]
- Shay, J.W.; Reddel, R.R.; Wright, W.E. Cancer and Telomeres—An ALTernative to Telomerase. Science 2012, 336, 1388–1390. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters | All (n) | BMDs (n) | Non-BMDs (n) |
|---|---|---|---|
| Breeds | |||
| mixed breeds | 3 | 0 | 3 |
| purebred | 60 | 47 | 13 |
| Sex | |||
| male/neutered * | 28/13 | 19/10 | 9/3 |
| female/spayed * | 30/11 | 25/7 | 5/4 |
| unknown | 5 | 3 | 2 |
| Age categories | |||
| mean age (years) | 8.1 | 7.6 | 9.5 |
| <5 years | 6 | 5 | 1 |
| 5–10 years | 46 | 36 | 10 |
| >10 years | 8 | 4 | 4 |
| unknown | 3 | 2 | 1 |
| Tumor site | |||
| skin | 29 | 19 | 10 |
| disseminated | 8 | 5 | 3 |
| spleen only | 11 | 9 | 2 |
| CNS | 1 | 0 | 1 |
| lymph nodes | 2 | 2 | 0 |
| liver | 2 | 2 | 0 |
| other | 2 | 2 | 0 |
| unknown | 8 | 8 | 0 |
| total number of cases | 63 | 47 | 16 |
| AU to BG Values of CCA | Cells Quantified | UBITF Cells | GITF Cells | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Case ID | Species/ Breed | Diagnosis | <2 | 2–5 | >5 | # | # | % | # | % | ALT | TC | Reference |
| 86638 | BMD | Histiocytic sarcoma | + | 10,153 | 2506 | 24.68 | 2478 | 24.41 | + | 2.62 | this study | ||
| 23671 | BMD | histiocytic sarcoma | + | 14,532 | 2385 | 16.41 | 922 | 6.34 | + | 3.00 | this study | ||
| 5332 | non-BMD | histiocytic sarcoma | + | 12,023 | 21 | 0.17 | 1219 | 10.14 | − | 1.50 | this study | ||
| 5249 | non-BMD | histiocytic sarcoma | + | 19,953 | 3 | 0.02 | 402 | 2.01 | − | 1.57 | this study | ||
| 89-2 | non-BMD | histiocytic sarcoma | + | 13,400 | 1534 | 11.45 | 2817 | 21.02 | + | 1.82 | [11] | ||
| 5682 | non-BMD | soft tissue sarcoma | + | 5969 | 5 | 0.08 | 168 | 2.81 | − | 0.99 | [11] | ||
| 6455 | non-BMD | hemangiosarcoma | + | 12,975 | 6090 | 46.94 | 62 | 0.48 | + | 3.33 | [11] | ||
| YTBO | human | astrocytoma | + | 3283 | 292 | 8.89 | 263 | 8.01 | + | 1.13 | [34] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kreilmeier-Berger, T.; Aupperle-Lellbach, H.; Reifinger, M.; Hörstke, N.V.; Holzmann, K.; Kleiter, M. Alternative Lengthening of Telomeres Is Rare in Canine Histiocytic Sarcoma. Cancers 2023, 15, 4214. https://doi.org/10.3390/cancers15174214
Kreilmeier-Berger T, Aupperle-Lellbach H, Reifinger M, Hörstke NV, Holzmann K, Kleiter M. Alternative Lengthening of Telomeres Is Rare in Canine Histiocytic Sarcoma. Cancers. 2023; 15(17):4214. https://doi.org/10.3390/cancers15174214
Chicago/Turabian StyleKreilmeier-Berger, Theresa, Heike Aupperle-Lellbach, Martin Reifinger, Nicolai Valentin Hörstke, Klaus Holzmann, and Miriam Kleiter. 2023. "Alternative Lengthening of Telomeres Is Rare in Canine Histiocytic Sarcoma" Cancers 15, no. 17: 4214. https://doi.org/10.3390/cancers15174214
APA StyleKreilmeier-Berger, T., Aupperle-Lellbach, H., Reifinger, M., Hörstke, N. V., Holzmann, K., & Kleiter, M. (2023). Alternative Lengthening of Telomeres Is Rare in Canine Histiocytic Sarcoma. Cancers, 15(17), 4214. https://doi.org/10.3390/cancers15174214