
Whole Exome Sequencing of Thymoma Patients Exhibiting Exceptional Responses to Pemetrexed Monotherapy
 , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Tissue Collection
2.2. Assessment and Tumor Response
2.3. Definition of Exceptional Response to Pemetrexed
2.4. DNA Extraction
2.5. Sequencing Analysis
2.6. Copy Number Variation (CNV) Analysis
2.7. Statistical Analysis
3. Results
3.1. Patient Characteristics and Sample Collection
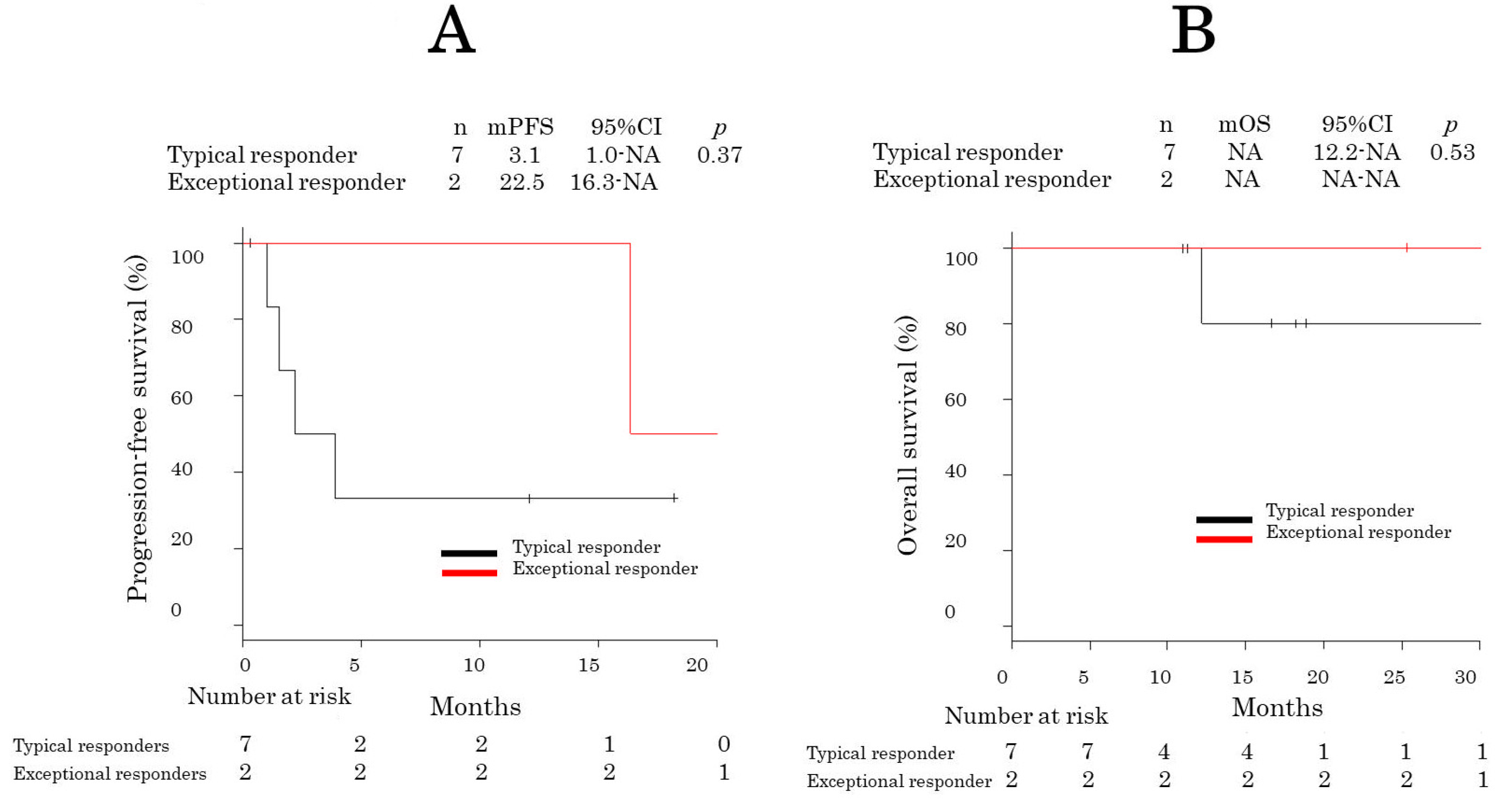
3.2. Response and Survival Analysis
3.3. Whole-Exome Sequencing
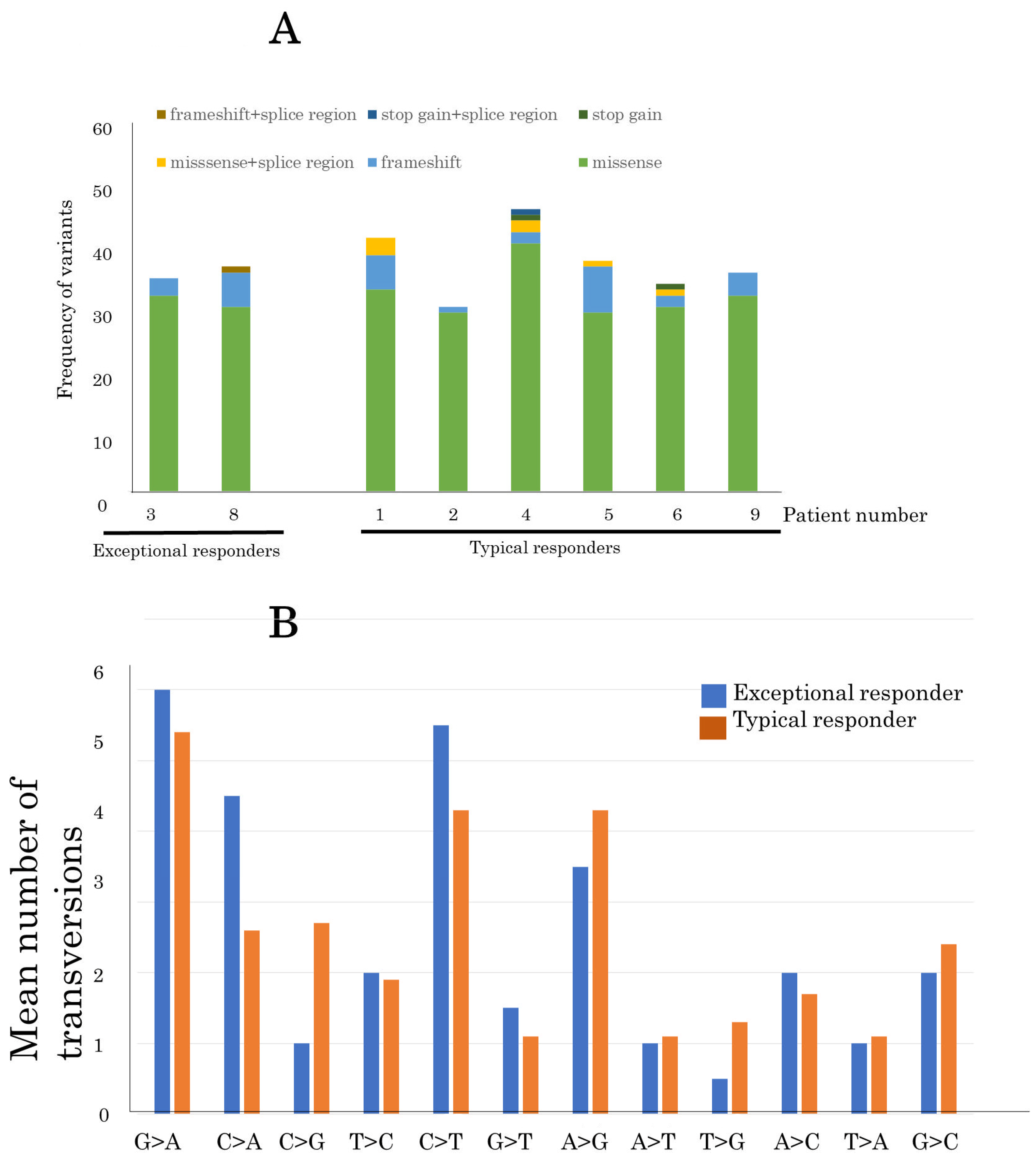
3.4. Identification of SNVs and Indels
3.5. Identification of CNVs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Travis, W.D.; Brambilla, E.; Burke, A.P.; Marx, A.; Nicholson, A.G. Pathology and Genetics: Tumors of the Lung, Pleura, Thymus and Heart, 4th ed.; IRCA Press: Gallarate, Italy, 2015; Volume 7. [Google Scholar]
- Hirai, F.; Toyozawa, R.; Nosaki, K.; Seto, T. Are Anthracycline-Based Regimens Truly Indicated To Be the Standard Chemotherapy Regimen for Thymic Carcinoma? J. Thorac. Oncol. 2016, 11, 115–121. [Google Scholar] [CrossRef]
- Loehrer, P.J.; Chen, M.; Kim, K.; Aisner, S.C.; Einhorn, L.H.; Livingston, R.; Johnson, D. Cisplatin, doxorubicin, and cyclophosphamide plus thoracic radiation therapy for limited-stage unresectable thymoma: An intergroup trial. J. Clin. Oncol. 1997, 15, 3093–3099. [Google Scholar] [CrossRef]
- Villela, L.R.; Stanford, B.L.; Shah, S.R. Pemetrexed, a novel antifolate therapeutic alternative for cancer chemotherapy. Pharmacotherapy 2006, 26, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Gbolahan, O.B.; Porter, R.F.; Salter, J.T.; Yiannoutsos, C.; Burns, M.; Chiorean, E.G.; Loehrer, P.J. A Phase II Study of Pemetrexed in Patients with Recurrent Thymoma and Thymic Carcinoma. J. Thorac. Oncol. 2018, 13, 1940–1948. [Google Scholar] [CrossRef]
- Lim, S.M.; Park, H.S.; Kim, S.; Ali, S.M.; Greenbowe, J.R.; Yang, I.S.; Kwon, N.J.; Lee, J.L.; Ryu, M.H.; Ahn, J.H.; et al. Next-generation sequencing reveals somatic mutations that confer exceptional response to everolimus. Oncotarget 2016, 7, 10547–10556. [Google Scholar] [CrossRef]
- Wagle, N.; Grabiner, B.C.; Van Allen, E.M.; Hodis, E.; Jacobus, S.; Supko, J.G.; Stewart, M.; Choueiri, T.K.; Gandhi, L.; Cleary, J.M.; et al. Activating mTOR mutations in a patient with an extraordinary response on a phase I trial of everolimus and pazopanib. Cancer Discov. 2014, 4, 546–553. [Google Scholar] [CrossRef]
- Krejci, J.; Pesek, M.; Grossmann, P.; Krejci, M.; Ricar, J.; Benesova, L.; Minarik, M. Extraordinary response to erlotinib therapy in a patient with lung adenocarcinoma exhibiting KRAS mutation and EGFR amplification. Cancer Genom. Proteom. 2011, 8, 135–138. [Google Scholar]
- Lim, S.M.; Kim, E.; Jung, K.H.; Kim, S.; Koo, J.S.; Kim, S.I.; Park, S.; Park, H.S.; Park, B.W.; Cho, Y.U.; et al. Genomic landscape of extraordinary responses in metastatic breast cancer. Commun. Biol. 2021, 4, 449. [Google Scholar] [CrossRef]
- Gray, H.J.; Chatterjee, P.; Rosati, R.; Appleyard, L.R.; Durenberger, G.J.; Diaz, R.L.; Swan, H.A.; Peretti, D.; Pollastro, M.; Ainge, T.; et al. Extraordinary clinical response to ibrutinib in low-grade ovarian cancer guided by organoid drug testing. NPJ Precis. Oncol. 2023, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Jiang, Z.; Liu, J.; Haverty, P.M.; Guan, Y.; Stinson, J.; Yue, P.; Zhang, Y.; Pant, K.P.; Bhatt, D.; et al. The mutation spectrum revealed by paired genome sequences from a lung cancer patient. Nature 2010, 465, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.J.; Ding, L.; Shen, D.; Luo, J.; Suman, V.J.; Wallis, J.W.; Van Tine, B.A.; Hoog, J.; Goiffon, R.J.; Goldstein, T.C.; et al. Whole-genome analysis informs breast cancer response to aromatase inhibition. Nature 2012, 486, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Network, C.G.A. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Petrini, I.; Meltzer, P.S.; Kim, I.K.; Lucchi, M.; Park, K.S.; Fontanini, G.; Gao, J.; Zucali, P.A.; Calabrese, F.; Favaretto, A.; et al. A specific missense mutation in GTF2I occurs at high frequency in thymic epithelial tumors. Nat. Genet. 2014, 46, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Lei, Y.; Wu, X.; Huang, Y.; Rao, H.; Zhang, Y.; Wang, F. GTF2I mutation frequently occurs in more indolent thymic epithelial tumors and predicts better prognosis. Lung Cancer 2017, 110, 48–52. [Google Scholar] [CrossRef]
- Belani, R.; Oliveira, G.; Erikson, G.A.; Ra, S.; Schechter, M.S.; Lee, J.K.; Shipman, W.J.; Haaser, S.M.; Torkamani, A. ASXL1 and DNMT3A mutation in a cytogenetically normal B3 thymoma. Oncogenesis 2014, 3, e111. [Google Scholar] [CrossRef] [PubMed]
- Girard, N.; Shen, R.; Guo, T.; Zakowski, M.F.; Heguy, A.; Riely, G.J.; Huang, J.; Lau, C.; Lash, A.E.; Ladanyi, M.; et al. Comprehensive genomic analysis reveals clinically relevant molecular distinctions between thymic carcinomas and thymomas. Clin. Cancer Res. 2009, 15, 6790–6799. [Google Scholar] [CrossRef]
- Girard, N.; Basse, C.; Schrock, A.; Ramkissoon, S.; Killian, K.; Ross, J.S. Comprehensive Genomic Profiling of 274 Thymic Epithelial Tumors Unveils Oncogenic Pathways and Predictive Biomarkers. Oncologist 2022, 27, 919–929. [Google Scholar] [CrossRef]
- Inoue, M.; Marx, A.; Zettl, A.; Ströbel, P.; Müller-Hermelink, H.K.; Starostik, P. Chromosome 6 suffers frequent and multiple aberrations in thymoma. Am. J. Pathol. 2002, 161, 1507–1513. [Google Scholar] [CrossRef]
- Zettl, A.; Ströbel, P.; Wagner, K.; Katzenberger, T.; Ott, G.; Rosenwald, A.; Peters, K.; Krein, A.; Semik, M.; Müller-Hermelink, H.K.; et al. Recurrent genetic aberrations in thymoma and thymic carcinoma. Am. J. Pathol. 2000, 157, 257–266. [Google Scholar] [CrossRef]
- Qian, X.; Song, Z. Efficacy of pemetrexed-based regimen in relapsed advanced thymic epithelial tumors and its association with thymidylate synthetase level. OncoTargets Ther. 2016, 9, 4527–4531. [Google Scholar] [CrossRef]
- Bai, Y.; Ma, X.; Zhang, Y.H. Systematic review and meta-analysis of the predictive power of MTHFR polymorphisms for pemetrexed drug efficacy and toxicity in non-small cell lung cancer patients. J. Chemother. 2021, 34, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Visser, S.; Hou, J.; Bezemer, K.; de Vogel, L.L.; Hegmans, J.P.J.J.; Stricker, B.H.; Philipsen, S.; Aerts, J.G.J.V. Prediction of response to pemetrexed in non-small-cell lung cancer with immunohistochemical phenotyping based on gene expression profiles. BMC Cancer 2019, 19, 440. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Oguri, T.; Kunii, E.; Sone, K.; Uemura, T.; Takakuwa, O.; Maeno, K.; Kanemitsu, Y.; Ohkubo, H.; Takemura, M.; et al. A folylpoly-γ-glutamate synthase single nucleotide polymorphism associated with response to pemetrexed treatment combined with platinum for non-small cell lung cancer. Lung Cancer 2016, 102, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Shan, F.; Liu, Y.L.; Wang, Q.; Shi, Y.L. Thymidylate synthase predicts poor response to pemetrexed chemotherapy in patients with advanced breast cancer. Oncol. Lett. 2018, 16, 3274–3280. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.T.; Chien, P.J.; Chen, S.H.; Sheu, G.T.; Jan, M.S.; Wang, B.Y.; Chang, W.W. BMI1-Mediated Pemetrexed Resistance in Non-Small Cell Lung Cancer Cells Is Associated with Increased SP1 Activation and Cancer Stemness. Cancers 2020, 12, 2069. [Google Scholar] [CrossRef]
- Qin, Y.; Sekine, I.; Hanazono, M.; Morinaga, T.; Fan, M.; Takiguchi, Y.; Tada, Y.; Shingyoji, M.; Yamaguchi, N.; Tagawa, M. AMPK activation induced in pemetrexed-treated cells is associated with development of drug resistance independently of target enzyme expression. Mol. Oncol. 2019, 13, 1419–1432. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Z.; Gao, S.; Wu, K.; Bai, F.; Zhang, Q.; Wang, H.; Ye, Q.; Xu, F.; Sun, H.; et al. Correction to: Human drug efflux transporter ABCC5 confers acquired resistance to pemetrexed in breast cancer. Cancer Cell. Int. 2021, 21, 183. [Google Scholar] [CrossRef]
- Eisenhauer, E.A.; Therasse, P.; Bogaerts, J.; Schwartz, L.H.; Sargent, D.; Ford, R.; Dancey, J.; Arbuck, S.; Gwyther, S.; Mooney, M.; et al. New response evaluation criteria in solid tumours: Revised RECIST guideline (version 1.1). Eur. J. Cancer 2009, 45, 228–247. [Google Scholar] [CrossRef]
- Talevich, E.; Shain, A.H.; Botton, T.; Bastian, B.C. CNVkit: Genome-Wide Copy Number Detection and Visualization from Targeted DNA Sequencing. PLoS Comput. Biol. 2016, 12, e1004873. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.C.; Couturier, D.L.; de Santiago, I.; Sauer, C.M.; Vias, M.; Angelova, M.; Sanders, D.; Piskorz, A.; Hall, J.; Hosking, K.; et al. Clonal somatic copy number altered driver events inform drug sensitivity in high-grade serous ovarian cancer. Nat. Commun. 2022, 13, 6360. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Han, X.; Zheng, G.; Yang, Y.; Li, Y.; Zhang, E.; Yang, L.; Dong, M.; He, D.; He, J.; et al. The adverse impact of a gain in chromosome 1q on the prognosis of multiple myeloma treated with bortezomib-based regimens: A retrospective single-center study in China. Front. Oncol. 2022, 12, 1084683. [Google Scholar] [CrossRef]
- Petrini, I.; Rajan, A.; Pham, T.; Voeller, D.; Davis, S.; Gao, J.; Wang, Y.; Giaccone, G. Whole genome and transcriptome sequencing of a B3 thymoma. PLoS ONE 2013, 8, e60572. [Google Scholar] [CrossRef] [PubMed]
- Petrini, I.; Meltzer, P.S.; Zucali, P.A.; Luo, J.; Lee, C.; Santoro, A.; Lee, H.S.; Killian, K.J.; Wang, Y.; Tsokos, M.; et al. Copy number aberrations of BCL2 and CDKN2A/B identified by array-CGH in thymic epithelial tumors. Cell. Death Dis. 2012, 3, e351. [Google Scholar] [CrossRef]
- Radovich, M.; Pickering, C.R.; Felau, I.; Ha, G.; Zhang, H.; Jo, H.; Hoadley, K.A.; Anur, P.; Zhang, J.; McLellan, M.; et al. The Integrated Genomic Landscape of Thymic Epithelial Tumors. Cancer Cell. 2018, 33, 244–258.210. [Google Scholar] [CrossRef]
- Xu, J.F.; Kang, Q.; Ma, X.Y.; Pan, Y.M.; Yang, L.; Jin, P.; Wang, X.; Li, C.G.; Chen, X.C.; Wu, C.; et al. A Novel Method to Detect Early Colorectal Cancer Based on Chromosome Copy Number Variation in Plasma. Cell. Physiol. Biochem. 2018, 45, 1444–1454. [Google Scholar] [CrossRef]
- Yu, D.; Liu, Z.; Su, C.; Han, Y.; Duan, X.; Zhang, R.; Liu, X.; Yang, Y.; Xu, S. Copy number variation in plasma as a tool for lung cancer prediction using Extreme Gradient Boosting (XGBoost) classifier. Thorac. Cancer 2020, 11, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.M.; Barwick, B.G.; Joseph, N.; Heffner, L.T.; Hofmeister, C.C.; Bernal, L.; Dhodapkar, M.V.; Gupta, V.A.; Jaye, D.L.; Wu, J.; et al. Gain of Chromosome 1q is associated with early progression in multiple myeloma patients treated with lenalidomide, bortezomib, and dexamethasone. Blood Cancer J. 2019, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Rajeshwari, M.; Sharma, M.C.; Kakkar, A.; Nambirajan, A.; Suri, V.; Sarkar, C.; Singh, M.; Saran, R.K.; Gupta, R.K. Evaluation of chromosome 1q gain in intracranial ependymomas. J. Neurooncol. 2016, 127, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Gratias, E.J.; Jennings, L.J.; Anderson, J.R.; Dome, J.S.; Grundy, P.; Perlman, E.J. Gain of 1q is associated with inferior event-free and overall survival in patients with favorable histology Wilms tumor: A report from the Children’s Oncology Group. Cancer 2013, 119, 3887–3894. [Google Scholar] [CrossRef] [PubMed]
- Robb, T.J.; Ward, Z.; Houseman, P.; Woodhouse, B.; Patel, R.; Fitzgerald, S.; Tsai, P.; Lawrence, B.; Parker, K.; Print, C.G.; et al. Chromosomal Aberrations Accumulate during Metastasis of Virus-Negative Merkel Cell Carcinoma. J. Investig. Dermatol. 2023, 143, 1168–1177.e1162. [Google Scholar] [CrossRef]
- McEvoy, C.R.; Morley, A.A.; Firgaira, F.A. Evidence for whole chromosome 6 loss and duplication of the remaining chromosome in acute lymphoblastic leukemia. Genes. Chromosomes Cancer 2003, 37, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Persson, F.; Fehr, A.; Sundelin, K.; Schulte, B.; Löning, T.; Stenman, G. Studies of genomic imbalances and the MYB-NFIB gene fusion in polymorphous low-grade adenocarcinoma of the head and neck. Int. J. Oncol. 2012, 40, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Pedersen-Bjergaard, J.; Pedersen, M.; Roulston, D.; Philip, P. Different genetic pathways in leukemogenesis for patients presenting with therapy-related myelodysplasia and therapy-related acute myeloid leukemia. Blood 1995, 86, 3542–3552. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, K.; Wang, X.; Liu, Y.; Guo, D.; Bian, Z.; Su, L.; Liu, K.; Gu, X.; Guo, X.; et al. Multiple-level copy number variations in cell-free DNA for prognostic prediction of HCC with radical treatments. Cancer Sci. 2021, 112, 4772–4784. [Google Scholar] [CrossRef]
- Chiang, J.; Moreira, D.C.; Li, X.; Furtado, L.V. Prognostic significance of chromosome arm 1q gain and methylation class in molecularly defined diffuse leptomeningeal glioneuronal tumor. Acta Neuropathol. 2022, 144, 1185–1187. [Google Scholar] [CrossRef]
- Aalto, Y.; Eriksson, L.; Seregard, S.; Larsson, O.; Knuutila, S. Concomitant loss of chromosome 3 and whole arm losses and gains of chromosome 1, 6, or 8 in metastasizing primary uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2001, 42, 313–317. [Google Scholar]
- Thomson, J.P.; Hollis, R.L.; van Baal, J.; Ilenkovan, N.; Churchman, M.; van de Vijver, K.; Dijk, F.; Meynert, A.M.; Bartos, C.; Rye, T.; et al. Whole exome sequencing of low grade serous ovarian carcinoma identifies genomic events associated with clinical outcome. Gynecol. Oncol. 2023, 174, 157–166. [Google Scholar] [CrossRef]
- Hattori, N.; Hirose, Y.; Sasaki, H.; Nakae, S.; Hayashi, S.; Ohba, S.; Adachi, K.; Hayashi, T.; Nishiyama, Y.; Hasegawa, M.; et al. World Health Organization grade II-III astrocytomas consist of genetically distinct tumor lineages. Cancer Sci. 2016, 107, 1159–1164. [Google Scholar] [CrossRef]
- Takeuchi, I.; Yanagi, K.; Takada, S.; Uchiyama, T.; Igarashi, A.; Motomura, K.; Hayashi, Y.; Nagano, N.; Matsuoka, R.; Sugiyama, H.; et al. STAT6 gain-of-function variant exacerbates multiple allergic symptoms. J. Allergy Clin. Immunol. 2023, 151, 1402–1409.e1406. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiang, W.; Yang, N.; Zhang, Y. Concurrent ERBB2 missense mutations D769Y and D742 N Are Novel Acquired Mechanism of Gefitinib Resistance but Responds to Gefitinib plus Pyrotinib. Lung Cancer 2020, 144, 90–91. [Google Scholar] [CrossRef] [PubMed]
- Cousin, M.A.; Zimmermann, M.T.; Mathison, A.J.; Blackburn, P.R.; Boczek, N.J.; Oliver, G.R.; Lomberk, G.A.; Urrutia, R.A.; Deyle, D.R.; Klee, E.W. Functional validation reveals the novel missense V419L variant in. Cold Spring Harb. Mol. Case Stud. 2017, 3, a001727. [Google Scholar] [CrossRef] [PubMed]
- Güemes, M.; Martín-Rivada, Á.; Ortiz-Cabrera, N.V.; Martos-Moreno, G.; Pozo-Román, J.; Argente, J. LZTR1: Genotype Expansion in Noonan Syndrome. Horm. Res. Paediatr. 2019, 92, 269–275. [Google Scholar] [CrossRef]
- Verrijn Stuart, A.A.; Ozisik, G.; de Vroede, M.A.; Giltay, J.C.; Sinke, R.J.; Peterson, T.J.; Harris, R.M.; Weiss, J.; Jameson, J.L. An amino-terminal DAX1 (NROB1) missense mutation associated with isolated mineralocorticoid deficiency. J. Clin. Endocrinol. Metab. 2007, 92, 755–761. [Google Scholar] [CrossRef]
- Puffenberger, E.G.; Jinks, R.N.; Wang, H.; Xin, B.; Fiorentini, C.; Sherman, E.A.; Degrazio, D.; Shaw, C.; Sougnez, C.; Cibulskis, K.; et al. A homozygous missense mutation in HERC2 associated with global developmental delay and autism spectrum disorder. Hum. Mutat. 2012, 33, 1639–1646. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Zhang, H.; Su, Y.; Wen, Z.; Zhu, Z.; Chen, G.; Peng, L.; Du, C.; Xie, H.; Li, H.; et al. Identification of two novel PCDHA9 mutations associated with Hirschsprung’s disease. Gene 2018, 658, 96–104. [Google Scholar] [CrossRef]
- Mouden, C.; de Tayrac, M.; Dubourg, C.; Rose, S.; Carré, W.; Hamdi-Rozé, H.; Babron, M.C.; Akloul, L.; Héron-Longe, B.; Odent, S.; et al. Homozygous STIL mutation causes holoprosencephaly and microcephaly in two siblings. PLoS ONE 2015, 10, e0117418. [Google Scholar] [CrossRef]
- Kowalik, A.; Karpinski, P.; Markiewicz, A.; Orlowska-Heitzman, J.; Romanowska-Dixon, B.; Donizy, P.; Hoang, M.P. Molecular profiling of primary uveal melanoma: Results of a Polish cohort. Melanoma Res. 2023, 33, 104–115. [Google Scholar] [CrossRef]
- Bilusic, M.; Girardi, D.; Zhou, Y.; Jung, K.; Pei, J.; Slifker, M.; Chen, Q.; Meerzaman, D.; Alpaugh, K.; Young, D.; et al. Molecular Profiling of Exceptional Responders to Cancer Therapy. Oncologist 2021, 26, 186–195. [Google Scholar] [CrossRef]
- Zaman, T.; Helbig, K.L.; Clatot, J.; Thompson, C.H.; Kang, S.K.; Stouffs, K.; Jansen, A.E.; Verstraete, L.; Jacquinet, A.; Parrini, E.; et al. SCN3A-Related Neurodevelopmental Disorder: A Spectrum of Epilepsy and Brain Malformation. Ann. Neurol. 2020, 88, 348–362. [Google Scholar] [CrossRef] [PubMed]
- Anuraga, G.; Tang, W.C.; Phan, N.N.; Ta, H.D.K.; Liu, Y.H.; Wu, Y.F.; Lee, K.H.; Wang, C.Y. Comprehensive Analysis of Prognostic and Genetic Signatures for General Transcription Factor III (GTF3) in Clinical Colorectal Cancer Patients Using Bioinformatics Approaches. Curr. Issues Mol. Biol. 2021, 43, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Hamanoue, S.; Goto, H.; Saito, T.; Nagai, J.I.; Masuno, M.; Umeda, Y.; Kurosawa, K. Diamond-Blackfan anemia caused by chromosome 1p22 deletion encompassing. Hum. Genome Var. 2019, 6, 36. [Google Scholar] [CrossRef]
- Wheeler, D.A.; Takebe, N.; Hinoue, T.; Hoadley, K.A.; Cardenas, M.F.; Hamilton, A.M.; Laird, P.W.; Wang, L.; Johnson, A.; Dewal, N.; et al. Molecular Features of Cancers Exhibiting Exceptional Responses to Treatment. Cancer Cell. 2021, 39, 38–53.e37. [Google Scholar] [CrossRef] [PubMed]
- Górska, A.A.; Sandmann, C.; Riechert, E.; Hofmann, C.; Malovrh, E.; Varma, E.; Kmietczyk, V.; Ölschläger, J.; Jürgensen, L.; Kamuf-Schenk, V.; et al. Muscle-specific Cand2 is translationally upregulated by mTORC1 and promotes adverse cardiac remodeling. EMBO Rep. 2021, 22, e52170. [Google Scholar] [CrossRef]
- Dong, S.S.; Yao, S.; Chen, Y.X.; Guo, Y.; Zhang, Y.J.; Niu, H.M.; Hao, R.H.; Shen, H.; Tian, Q.; Deng, H.W.; et al. Detecting epistasis within chromatin regulatory circuitry reveals CAND2 as a novel susceptibility gene for obesity. Int. J. Obes. 2019, 43, 450–456. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Gender | Age | WHO-Subtypes | Masaoka-Stage | PS | Line of Therapy | Previous Chemotherapy | Paraneoplastic Syndrome | Response | Eceptional Response | Mean Depth of Target Lesion (×) | PFS, Months | OS, Months |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | M | 61 | B1 | IVa | 1 | 2 | CBDCA+PTX | non | SD | no | 161.7 | 3.9 | 12.1 |
| 2 | F | 43 | B3 | IVa | 1 | 2 | ADOC | non | SD | no | 179.3 | NR | NR |
| 3 | F | 51 | B3 | III | 1 | 3 | CBDCA+PTX, CODE | MG | PR | yes | 96.5 | 28.6 | NR |
| 4 | F | 66 | B3 | IVb | 0 | 2 | ADOC | non | SD | no | 147.8 | NR | NR |
| 5 | F | 54 | B2+B3 | III | 1 | 2 | CODE | PRCA | SD | no | 177.6 | NR | NR |
| 6 | F | 36 | B2+B3 | IVb | 1 | 2 | CODE | non | PD | no | 189.1 | 2.2 | NR |
| 7 | F | 77 | B2 | IVa | 1 | 3 | AMR, CODE | non | SD | no | 39.7 | 1.0 | NR |
| 8 | F | 51 | B3 | IVa | 1 | 2 | CODE | MG | PR | yes | 205.2 | 16.3 | NR |
| 9 | F | 45 | B2 | III | 0 | 1 | non | non | PD | no | 183.7 | 1.5 | NR |
| Patient No | Gene Name | Position | Nucleotide | AF | Effect |
|---|---|---|---|---|---|
| 1 | STAT6 | chr12:57,102,481 | c.1321G>A | 0.3783784 | missense |
| 1 | CFAP54 | chr12: 96,489,820 | c.211C>T | 0.4841629 | missense |
| 1 | ERBBB2 | chr17:39,723,566 | c.2114C>T | 0.3850932 | missense |
| 1 | BIRC6 | chr2:32,431,088 | c.3246C>A | 0.50 | missense and splice region |
| 1 | TGFBR2 | chr3:30,623,201 | c.97G>A | 0.3109244 | missense and splice region |
| 2 | SACS | chr13:23,336,092 | c.7784C>T | 0.4642857 | missense |
| 2 | UBR5 | chr8:102,286,377 | c.5198G>A | 0.4926829 | missense |
| 3 | HNF1A | chr12:120,993,565 | c.572G>A | 0.6333333 | missense |
| 3 | LZTR1 | chr22:20,988,851 | c.572C>T | 0.5785441 | missense |
| 4 | TDRD6 | chr6:46,688,924 | c.796C>A | 0.3931624 | missense |
| 4 | UTRN | chr6:144,782,110 | c.8821A>G | 0.2698413 | missense |
| 4 | PKHD1L1 | chr8:109,401,516 | c.1301C>A | 0.4382022 | missense |
| 5 | PAM | chr5:102,974,297 | c.1344G>C | 0.4698795 | missense |
| 5 | NROB1 | chrX:30,308,879 | c.485C>T | 0.4477825 | missense |
| 6 | DOCK2 | chr5:169,759,723 | c.2395A>C | 0.75 | missense |
| 6 | ANK1 | chr8:169,759,723 | c.2473G>A | 0.3671875 | missense |
| 8 | HERC2 | chr15:28,255,910 | c.2833G>A | 0.4848485 | missense |
| 8 | ATP2C2 | chr16:84,422,514 | c.749C>T | 0.4411765 | missense |
| 8 | AXIN2 | chr17:65,549,596 | c.880G>C | 0.5213033 | missense |
| 8 | PCDHA9 | chr5:140,850,275 | c.1780G>A | 0.6423611 | missense |
| 9 | STIL | chr1:47,280,717 | c.1741T>G | 0.6149068 | missense |
| 9 | ZFXH2 | chr16:72,796,380 | c.6302T>C | 0.3972603 | missense |
| 9 | LYL1 | chr19:13,099,622 | c.540C>G | 0.4915254 | missense |
| 9 | FBXW7 | chr4:152,326,028 | c.1622C>G | 0.4647887 | missense |
| Patient No | CHROM | POS | REF | ALT | AF | Effect | Gene_Name | Deletorius | HGVS.p |
|---|---|---|---|---|---|---|---|---|---|
| 1 | chr11 | 44,919,272 | C | CCTGG | 0.4245077 | frameshift_variant | TSPAN18 | p.Asp132fs | |
| 1 | chr16 | 27,470,158 | C | CTT | 0.5069124 | frameshift_variant | GTF3C1 | yes | p.Gln1589fs |
| 1 | chr17 | 50,528,794 | TCA | T | 0.16 | frameshift_variant | MYCBPAP | p.Lys880fs | |
| 1 | chr3 | 157,437,716 | AG | A | 0.5151515 | frameshift_variant | PTX3 | p.Arg112fs | |
| 1 | chr7 | 34,658,494 | GA | G | 0.4679803 | frameshift_variant | NPSR1 | p.Thr29fs | |
| 1 | chr8 | 141,163,405 | A | AG | 0.4418605 | frameshift_variant | DENND3 | p.Asp397fs | |
| 2 | chr22 | 23,838,973 | TC | T | 0.52 | frameshift_variant | DERL3 | p.Gly5fs | |
| 3 | chr22 | 23,838,973 | TC | T | 0.4519231 | frameshift_variant | DERL3 | p.Gly5fs | |
| 3 | chr3 | 196,707,700 | TGG | T | 0.15 | frameshift_variant | CEP19 | p.Ala118fs | |
| 3 | chr6 | 117,365,113 | GTA | G | 0.1025641 | frameshift_variant | ROS1 | p.Thr1022fs | |
| 4 | chr12 | 113,163,221 | CCT | C | 0.5619835 | frameshift_variant | DDX54 | p.Arg664fs | |
| 4 | chr12 | 121,954,299 | GA | G | 0.4193548 | frameshift_variant | WDR66 | p.Glu502fs | |
| 5 | chr10 | 99,930,218 | C | CTATATATA | 0.425 | frameshift_variant | DNMBP | p.Glu182fs | |
| 5 | chr12 | 13,090,671 | C | CT | 0.4492188 | frameshift_variant | GSG1 | p.Gly66fs | |
| 5 | chr16 | 29,797,007 | A | ATCCC | 0.4829396 | frameshift_variant | KIF22 | p.Pro64fs | |
| 5 | chr17 | 41,084,543 | A | AT | 0.130137 | frameshift_variant | KRTAP4-7 | p.Ser113fs | |
| 5 | chr17 | 46,549,134 | ATC | A | 0.2313725 | frameshift_variant | LRRC37A2 | p.Leu1333fs | |
| 5 | chr2 | 29,920,063 | G | GC | 0.4986702 | frameshift_variant | ALK | p.Arg200fs | |
| 5 | chr2 | 165,127,882 | A | AATAAT | 0.4411765 | frameshift_variant | SCN3A | yes | p.Cys1048fs |
| 5 | chr9 | 137,232,083 | T | TC | 0.4404762 | frameshift_variant | SLC34A3 | p.Ala34fs | |
| 6 | chr11 | 68,033,138 | T | TC | 0.4482759 | frameshift_variant | NDUFS8 | p.Arg77fs | |
| 6 | chr7 | 2,250,932 | CG | C | 0.4650206 | frameshift_variant | NUDT1 | p.Tyr159fs | |
| 8 | chr1 | 92,833,642 | CAG | C | 0.3971631 | frameshift_variant | RPL5 | yes | p.Asp59fs |
| 8 | chr12 | 55,470,067 | TACAATCTGTA | T | 0.4090909 | frameshift_variant | OR6C70 | p.Leu21fs | |
| 8 | chr15 | 79,456,702 | GAAAA | G | 0.1346154 | frameshift_variant | KIAA1024 | p.Lys186fs | |
| 8 | chr15 | 79,456,706 | A | ACCCC | 0.1390728 | frameshift_variant | KIAA1024 | p.Asn187fs | |
| 8 | chr19 | 43,613,237 | C | CAAAG | 0.4780876 | frameshift_variant | SRRM5 | p.Asp376fs | |
| 8 | chr19 | 54,574,302 | GC | G | 0.1104294 | frameshift_variant& | LILRA2 | p.His25fs | |
| splice_region_variant | |||||||||
| 8 | chr3 | 12,815,222 | TC | T | 0.88 | frameshift_variant | CAND2 | p.Ser364fs | |
| 9 | chr1 | 53,082,128 | A | AG | 0.4193548 | frameshift_variant | PODN | p.Glu652fs | |
| 9 | chr11 | 66,366,223 | C | CAGCCAGATCTGGG | 0.3613861 | frameshift_variant | SLC29A2 | p.Thr293fs | |
| 9 | chr19 | 53,807,589 | C | CT | 0.4126214 | frameshift_variant | NLRP12 | p.Glu718fs | |
| 9 | chr7 | 100,578,215 | GC | G | 0.5545455 | frameshift_variant | LRCH4 | p.Leu298fs |
| CHR | Patient 1 | Patient 2 | Patient 4 | Patient 5 | Patient 6 | Patient 8 | Patient 9 |
|---|---|---|---|---|---|---|---|
| 1p | |||||||
| 1q | gain | gain | gain | gain | |||
| 2p | loss | ||||||
| 2q | |||||||
| 3p | loss | ||||||
| 3q | gain | ||||||
| 4p | |||||||
| 4q | |||||||
| 5p | gain | gain | gain | ||||
| 5q | gain | ||||||
| 6p | loss | loss | |||||
| 6q | loss | loss | |||||
| 7p | gain | gain | |||||
| 7q | gain | gain | |||||
| 8p | |||||||
| 8q | |||||||
| 9p | gain | ||||||
| 9q | |||||||
| 10p | |||||||
| 10q | |||||||
| 11p | |||||||
| 11q | |||||||
| 12p | |||||||
| 12q | |||||||
| 13p | loss | ||||||
| 13q | loss | ||||||
| 14p | gain | ||||||
| 14q | gain | ||||||
| 15p | |||||||
| 15q | |||||||
| 16p | |||||||
| 16q | |||||||
| 17p | |||||||
| 17q | |||||||
| 18p | |||||||
| 18q | |||||||
| 19p | |||||||
| 19q | |||||||
| 20p | |||||||
| 20q | |||||||
| 21p | |||||||
| 21q | |||||||
| 22p | |||||||
| 22q |
| Chr | Gain | Loss | ||||
|---|---|---|---|---|---|---|
| p Arm (%) | q Arm (%) | Whole Arms (%) | p Arm (%) | q Arm (%) | Whole Arms (%) | |
| 1 | 0 (0) | 4 (57.1) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 2 | 0 (0) | 0 (0) | 0 (0) | 1 (14.3) | 0 (0) | 0 (0) |
| 3 | 0 (0) | 1 (14.3) | 0 (0) | 1 (14.3) | 0 (0) | 0 (0) |
| 4 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 5 | 3 (42.9) | 1 (14.3) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 6 | 0 (0) | 0 (0) | 0 (0) | 2 (28.6) | 2 (28.6) | 0 (0) |
| 7 | 2 (28.6) | 2 (28.6) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 8 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 9 | 1 (14.3) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 10 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 11 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 12 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 13 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (14.3) |
| 14 | 0 (0) | 0 (0) | 1 (14.3) | 0 (0) | 0 (0) | 0 (0) |
| 15 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 16 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 17 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 18 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 19 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 20 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 21 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 22 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, T.; Goto, Y.; Horie, M.; Masuda, K.; Shinno, Y.; Matsumoto, Y.; Okuma, Y.; Yoshida, T.; Horinouchi, H.; Motoi, N.; et al. Whole Exome Sequencing of Thymoma Patients Exhibiting Exceptional Responses to Pemetrexed Monotherapy. Cancers 2023, 15, 4018. https://doi.org/10.3390/cancers15164018
Tanaka T, Goto Y, Horie M, Masuda K, Shinno Y, Matsumoto Y, Okuma Y, Yoshida T, Horinouchi H, Motoi N, et al. Whole Exome Sequencing of Thymoma Patients Exhibiting Exceptional Responses to Pemetrexed Monotherapy. Cancers. 2023; 15(16):4018. https://doi.org/10.3390/cancers15164018
Chicago/Turabian StyleTanaka, Tomohiro, Yasushi Goto, Masafumi Horie, Ken Masuda, Yuki Shinno, Yuji Matsumoto, Yusuke Okuma, Tatsuya Yoshida, Hidehito Horinouchi, Noriko Motoi, and et al. 2023. "Whole Exome Sequencing of Thymoma Patients Exhibiting Exceptional Responses to Pemetrexed Monotherapy" Cancers 15, no. 16: 4018. https://doi.org/10.3390/cancers15164018
APA StyleTanaka, T., Goto, Y., Horie, M., Masuda, K., Shinno, Y., Matsumoto, Y., Okuma, Y., Yoshida, T., Horinouchi, H., Motoi, N., Yatabe, Y., Watanabe, S., Yamamoto, N., & Ohe, Y. (2023). Whole Exome Sequencing of Thymoma Patients Exhibiting Exceptional Responses to Pemetrexed Monotherapy. Cancers, 15(16), 4018. https://doi.org/10.3390/cancers15164018