

Role of Intracellular Drug Disposition in the Response of Acute Myeloid Leukemia to Cytarabine and Idarubicin Induction Chemotherapy

 , , , ,
, , , ,  and
and 
Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Population and Elegibility
2.2. Cell Lines
2.3. Cell Viability Assay
2.4. Determination of Gene Expression
2.5. Functional Studies
2.6. Statistical Analysis
3. Results
3.1. Characteristics of the Study Population
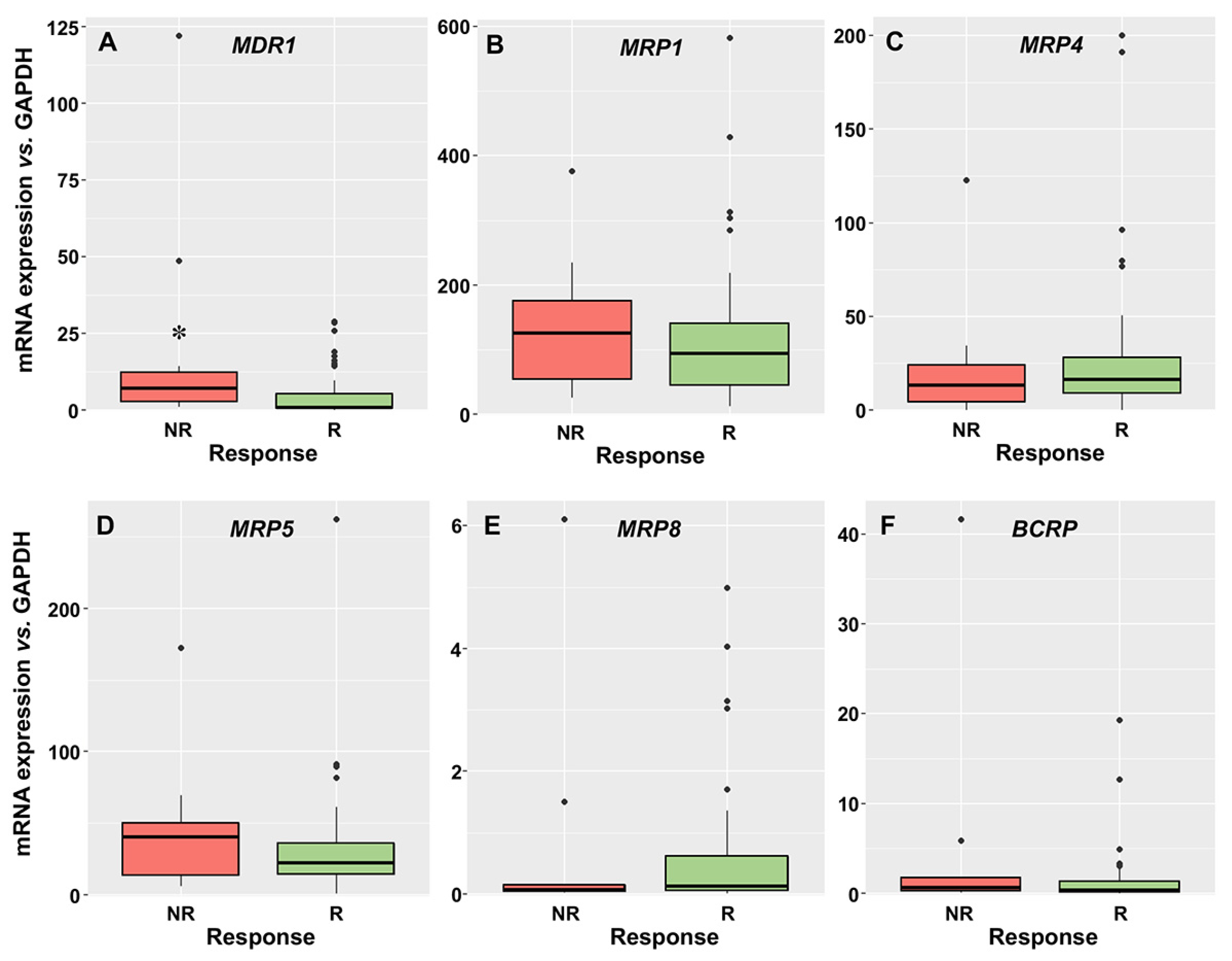
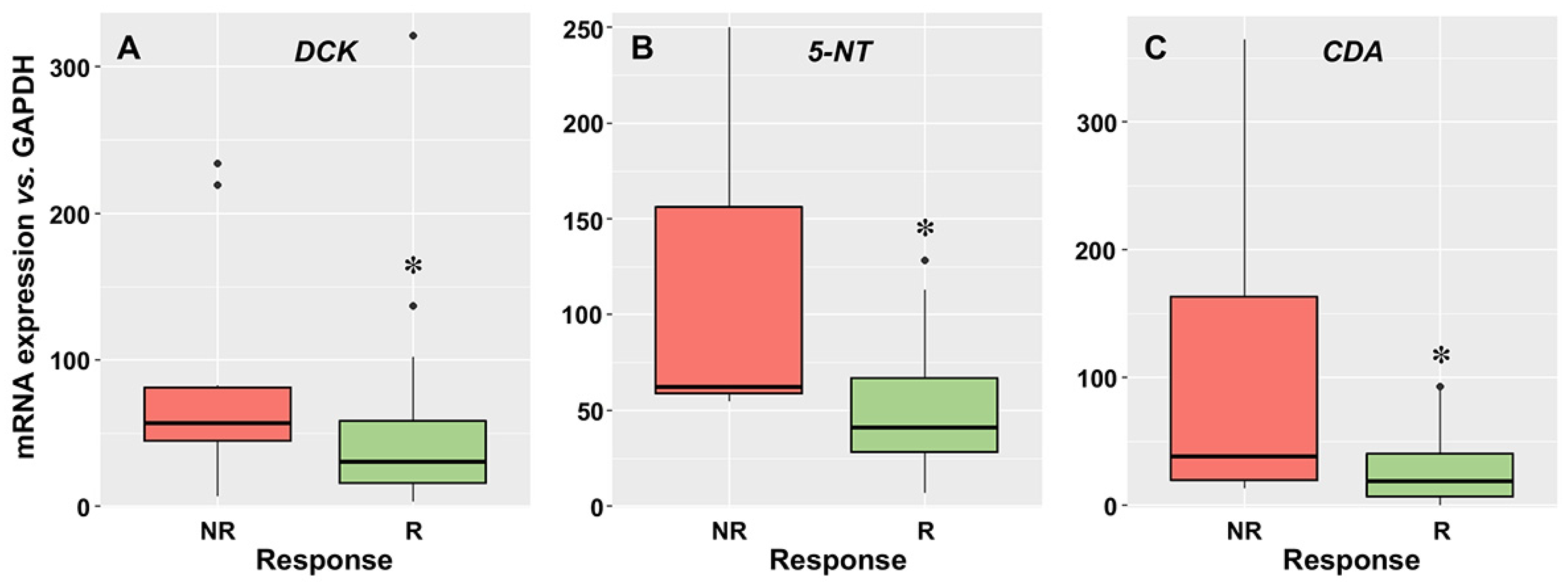
3.2. Relationship between the Expression of Drug Transporters and Enzymes and the Response to Induction Therapy
3.3. Sensitivity of Leukemic Cells to Cytarabine and Idarubicin
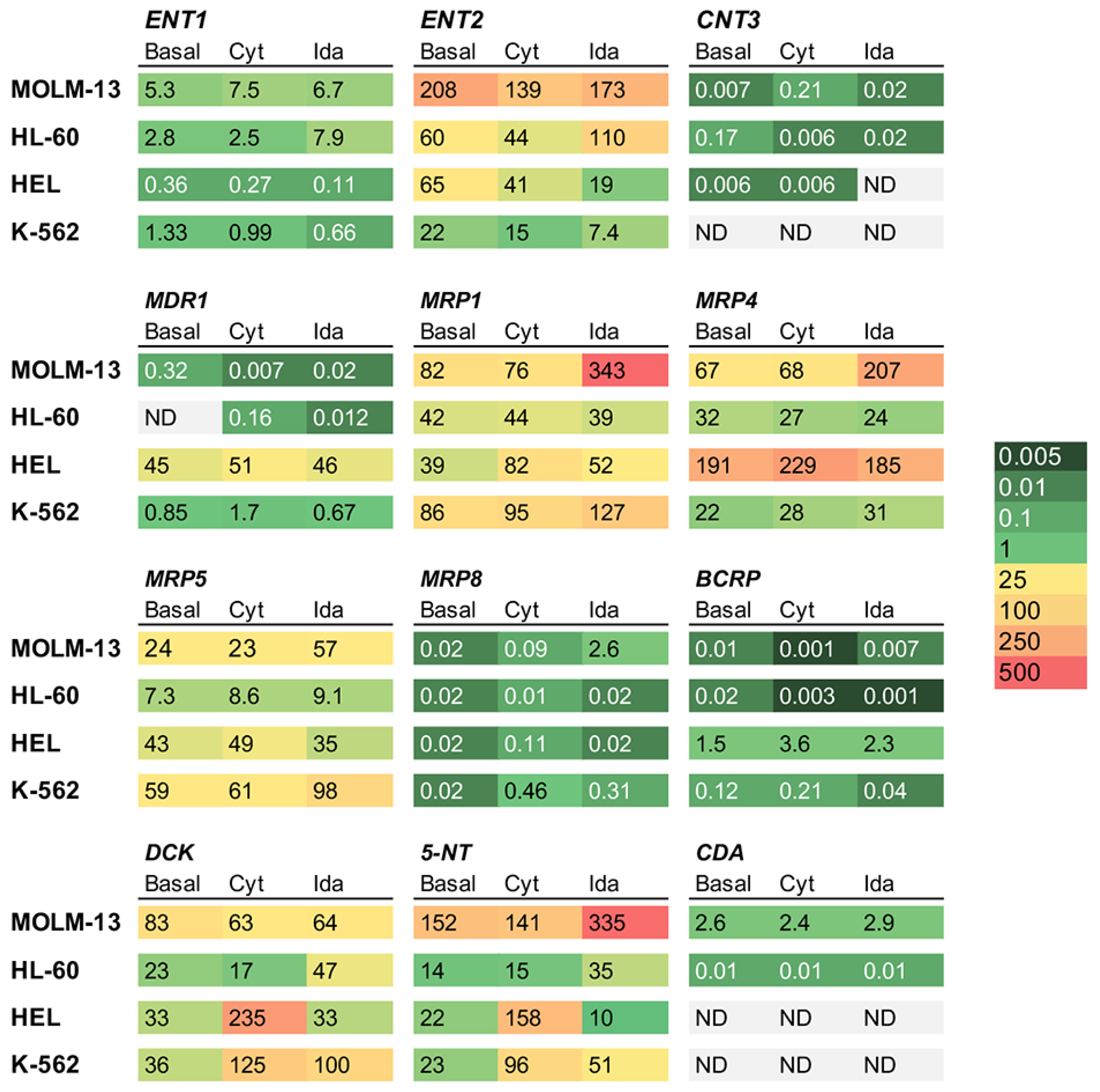
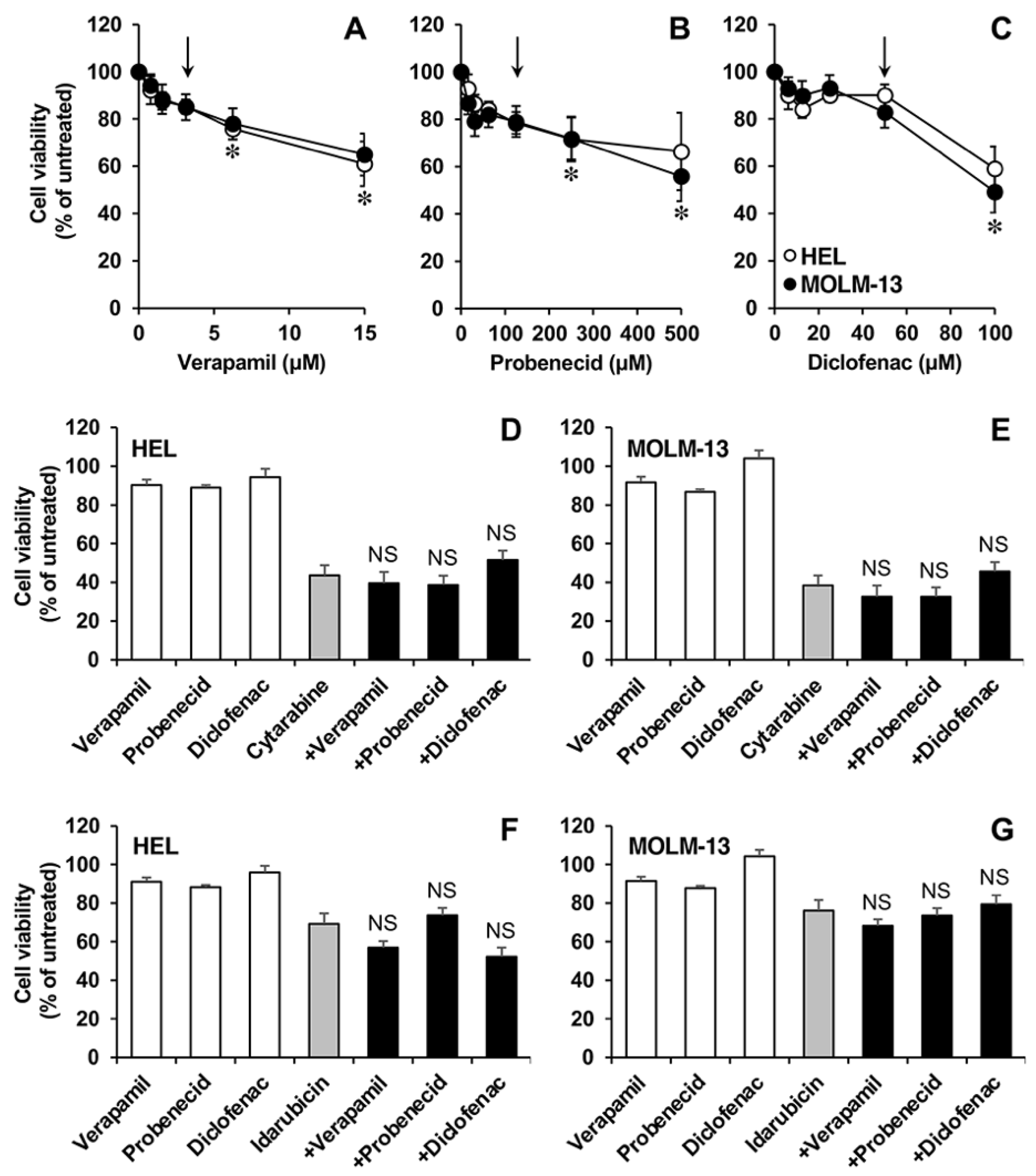
3.4. Effect of Drug Exposure of Transporters and Enzymes in AML Cell Lines
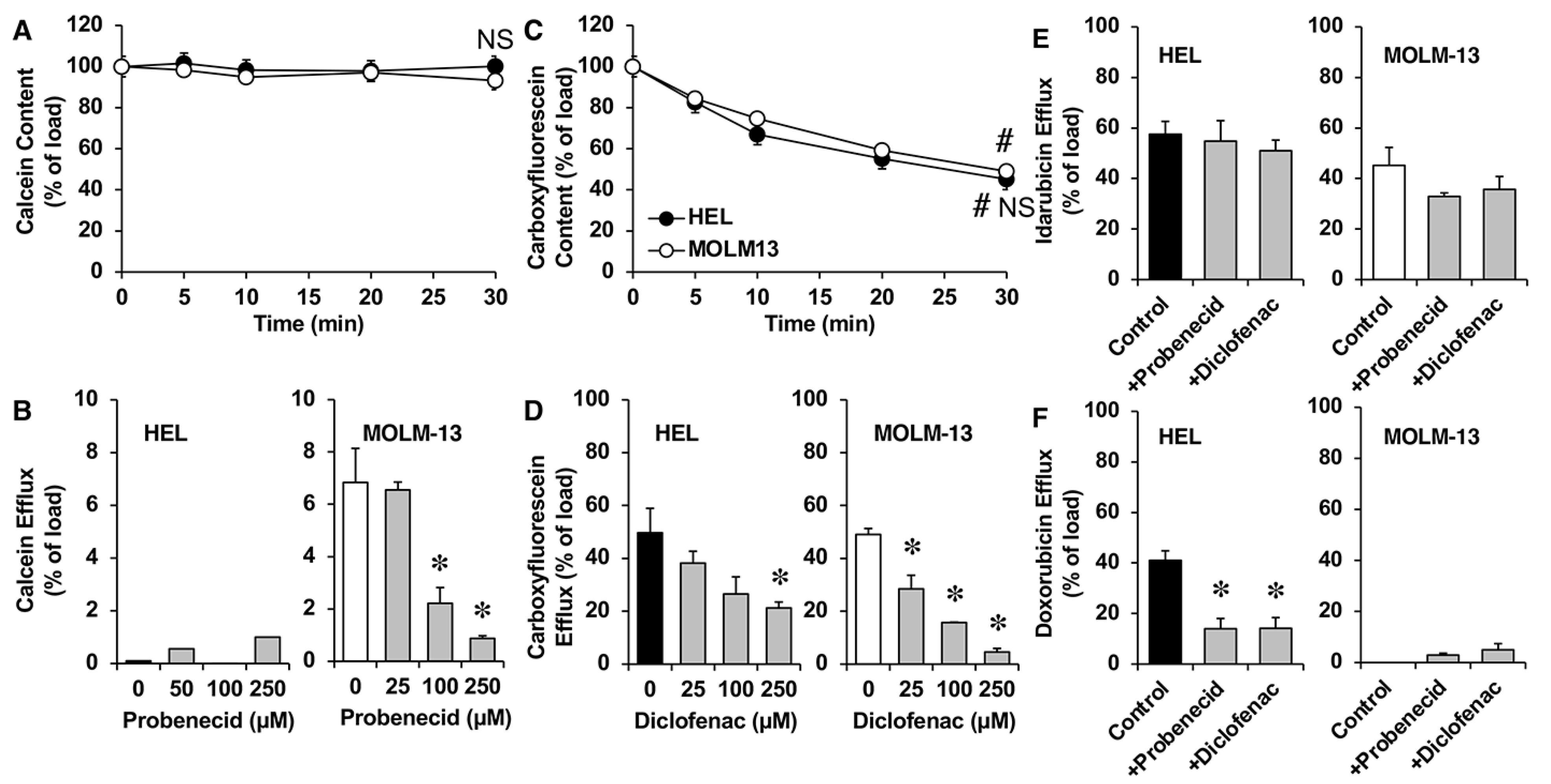
3.5. Functional Studies in AML Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marin, J.J.; Briz, O.; Rodriguez-Macias, G.; Diez-Martin, J.L.; Macias, R.I. Role of drug transport and metabolism in the chemoresistance of acute myeloid leukemia. Blood Rev. 2016, 30, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Thol, F.; Schlenk, R.F.; Heuser, M.; Ganser, A. How I treat refractory and early relapsed acute myeloid leukemia. Blood 2015, 126, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Marin, J.J.; Monte, M.J.; Blazquez, A.G.; Macias, R.I.; Serrano, M.A.; Briz, O. The role of reduced intracellular concentrations of active drugs in the lack of response to anticancer chemotherapy. Acta Pharmacol. Sin. 2014, 35, 1–10. [Google Scholar] [CrossRef]
- Abraham, A.; Varatharajan, S.; Karathedath, S.; Philip, C.; Lakshmi, K.M.; Jayavelu, A.K.; Mohanan, E.; Janet, N.B.; Srivastava, V.M.; Shaji, R.V.; et al. RNA expression of genes involved in cytarabine metabolism and transport predicts cytarabine response in acute myeloid leukemia. Pharmacogenomics 2015, 16, 877–890. [Google Scholar] [CrossRef]
- Adema, A.D.; Floor, K.; Smid, K.; Honeywell, R.J.; Scheffer, G.L.; Jansen, G.; Peters, G.J. Overexpression of MRP4 (ABCC4) and MRP5 (ABCC5) confer resistance to the nucleoside analogs cytarabine and troxacitabine, but not gemcitabine. Springerplus 2014, 3, 732. [Google Scholar] [CrossRef]
- Guo, Y.; Kock, K.; Ritter, C.A.; Chen, Z.S.; Grube, M.; Jedlitschky, G.; Illmer, T.; Ayres, M.; Beck, J.F.; Siegmund, W.; et al. Expression of ABCC-type nucleotide exporters in blasts of adult acute myeloid leukemia: Relation to long-term survival. Clin. Cancer Res. 2009, 15, 1762–1769. [Google Scholar] [CrossRef]
- Grant, S. Ara-C: Cellular and molecular pharmacology. Adv. Cancer Res. 1998, 72, 197–233. [Google Scholar] [CrossRef]
- Wu, B.; Mao, Z.J.; Wang, Z.; Wu, P.; Huang, H.; Zhao, W.; Zhang, L.; Zhang, Z.; Yin, H.; Gale, R.P.; et al. Deoxycytidine Kinase (DCK) Mutations in Human Acute Myeloid Leukemia Resistant to Cytarabine. Acta Haematol. 2021, 144, 534–541. [Google Scholar] [CrossRef]
- Fanciullino, R.; Farnault, L.; Donnette, M.; Imbs, D.C.; Roche, C.; Venton, G.; Berda-Haddad, Y.; Ivanov, V.; Ciccolini, J.; Ouafik, L.; et al. CDA as a predictive marker for life-threatening toxicities in patients with AML treated with cytarabine. Blood Adv. 2018, 2, 462–469. [Google Scholar] [CrossRef]
- Yamauchi, T.; Negoro, E.; Kishi, S.; Takagi, K.; Yoshida, A.; Urasaki, Y.; Iwasaki, H.; Ueda, T. Intracellular cytarabine triphosphate production correlates to deoxycytidine kinase/cytosolic 5'-nucleotidase II expression ratio in primary acute myeloid leukemia cells. Biochem. Pharmacol. 2009, 77, 1780–1786. [Google Scholar] [CrossRef]
- Hubeek, I.; Peters, G.J.; Broekhuizen, R.; Zwaan, C.M.; Kaaijk, P.; van Wering, E.S.; Gibson, B.E.; Creutzig, U.; Janka-Schaub, G.E.; den Boer, M.L.; et al. In vitro sensitivity and cross-resistance to deoxynucleoside analogs in childhood acute leukemia. Haematologica 2006, 91, 17–23. [Google Scholar] [PubMed]
- Regev, R.; Yeheskely-Hayon, D.; Katzir, H.; Eytan, G.D. Transport of anthracyclines and mitoxantrone across membranes by a flip-flop mechanism. Biochem. Pharmacol. 2005, 70, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Karim, H.; Bogason, A.; Bhuiyan, H.; Fotoohi, A.K.; Lafolie, P.; Vitols, S. Comparison of uptake mechanisms for anthracyclines in human leukemic cells. Curr. Drug Deliv. 2013, 10, 404–412. [Google Scholar] [CrossRef]
- Andreev, E.; Brosseau, N.; Carmona, E.; Mes-Masson, A.M.; Ramotar, D. The human organic cation transporter OCT1 mediates high affinity uptake of the anticancer drug daunorubicin. Sci. Rep. 2016, 6, 20508. [Google Scholar] [CrossRef]
- Leith, C.P.; Kopecky, K.J.; Godwin, J.; McConnell, T.; Slovak, M.L.; Chen, I.M.; Head, D.R.; Appelbaum, F.R.; Willman, C.L. Acute myeloid leukemia in the elderly: Assessment of multidrug resistance (MDR1) and cytogenetics distinguishes biologic subgroups with remarkably distinct responses to standard chemotherapy. A Southwest Oncology Group study. Blood 1997, 89, 3323–3329. [Google Scholar] [CrossRef]
- Megias-Vericat, J.E.; Rojas, L.; Herrero, M.J.; Boso, V.; Montesinos, P.; Moscardo, F.; Poveda, J.L.; Sanz, M.A.; Alino, S.F. Influence of ABCB1 polymorphisms upon the effectiveness of standard treatment for acute myeloid leukemia: A systematic review and meta-analysis of observational studies. Pharm. J. 2015, 15, 109–118. [Google Scholar] [CrossRef]
- Broxterman, H.J.; Sonneveld, P.; van Putten, W.J.; Lankelma, J.; Eekman, C.A.; Ossenkoppele, G.J.; Pinedo, H.M.; Lowenberg, B.; Schuurhuis, G.J. P-glycoprotein in primary acute myeloid leukemia and treatment outcome of idarubicin/cytosine arabinoside-based induction therapy. Leukemia 2000, 14, 1018–1024. [Google Scholar] [CrossRef]
- Pallis, M.; Hills, R.; White, P.; Grundy, M.; Russell, N.; Burnett, A. Analysis of the interaction of induction regimens with p-glycoprotein expression in patients with acute myeloid leukaemia: Results from the MRC AML15 trial. Blood Cancer J. 2011, 1, e23. [Google Scholar] [CrossRef]
- Schaich, M.; Soucek, S.; Thiede, C.; Ehninger, G.; Illmer, T.; Group, S.A.S. MDR1 and MRP1 gene expression are independent predictors for treatment outcome in adult acute myeloid leukaemia. Br. J. Haematol. 2005, 128, 324–332. [Google Scholar] [CrossRef]
- Wojnowski, L.; Kulle, B.; Schirmer, M.; Schluter, G.; Schmidt, A.; Rosenberger, A.; Vonhof, S.; Bickeboller, H.; Toliat, M.R.; Suk, E.K.; et al. NAD(P)H oxidase and multidrug resistance protein genetic polymorphisms are associated with doxorubicin-induced cardiotoxicity. Circulation 2005, 112, 3754–3762. [Google Scholar] [CrossRef]
- Benderra, Z.; Faussat, A.M.; Sayada, L.; Perrot, J.Y.; Chaoui, D.; Marie, J.P.; Legrand, O. Breast cancer resistance protein and P-glycoprotein in 149 adult acute myeloid leukemias. Clin. Cancer Res. 2004, 10, 7896–7902. [Google Scholar] [CrossRef] [PubMed]
- Damiani, D.; Tiribelli, M.; Calistri, E.; Geromin, A.; Chiarvesio, A.; Michelutti, A.; Cavallin, M.; Fanin, R. The prognostic value of P-glycoprotein (ABCB) and breast cancer resistance protein (ABCG2) in adults with de novo acute myeloid leukemia with normal karyotype. Haematologica 2006, 91, 825–828. [Google Scholar] [PubMed]
- Megias-Vericat, J.E.; Montesinos, P.; Herrero, M.J.; Moscardo, F.; Boso, V.; Rojas, L.; Martinez-Cuadron, D.; Hervas, D.; Boluda, B.; Garcia-Robles, A.; et al. Impact of ABC single nucleotide polymorphisms upon the efficacy and toxicity of induction chemotherapy in acute myeloid leukemia. Leuk. Lymphoma 2017, 58, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Megias-Vericat, J.E.; Martinez-Cuadron, D.; Herrero, M.J.; Alino, S.F.; Poveda, J.L.; Sanz, M.A.; Montesinos, P. Pharmacogenetics of Metabolic Genes of Anthracyclines in Acute Myeloid Leukemia. Curr. Drug Metab. 2018, 19, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Cheson, B.D.; Bennett, J.M.; Kopecky, K.J.; Buchner, T.; Willman, C.L.; Estey, E.H.; Schiffer, C.A.; Doehner, H.; Tallman, M.S.; Lister, T.A.; et al. Revised recommendations of the International Working Group for Diagnosis, Standardization of Response Criteria, Treatment Outcomes, and Reporting Standards for Therapeutic Trials in Acute Myeloid Leukemia. J. Clin. Oncol. 2003, 21, 4642–4649. [Google Scholar] [CrossRef]
- Macias, R.I.R.; Sanchez-Martin, A.; Rodriguez-Macias, G.; Sanchez-Abarca, L.I.; Lozano, E.; Herraez, E.; Odero, M.D.; Diez-Martin, J.L.; Marin, J.J.G.; Briz, O. Role of drug transporters in the sensitivity of acute myeloid leukemia to sorafenib. Oncotarget 2018, 9, 28474–28485. [Google Scholar] [CrossRef]
- Dohner, H.; Estey, E.; Grimwade, D.; Amadori, S.; Appelbaum, F.R.; Buchner, T.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Larson, R.A.; et al. Diagnosis and management of AML in adults: 2017 ELN recommendations from an international expert panel. Blood 2017, 129, 424–447. [Google Scholar] [CrossRef]
- Pisco, A.O.; Jackson, D.A.; Huang, S. Reduced Intracellular Drug Accumulation in Drug-Resistant Leukemia Cells is Not Only Solely Due to MDR-Mediated Efflux but also to Decreased Uptake. Front. Oncol. 2014, 4, 306. [Google Scholar] [CrossRef]
- Su, M.; Chang, Y.T.; Hernandez, D.; Jones, R.J.; Ghiaur, G. Regulation of drug metabolizing enzymes in the leukaemic bone marrow microenvironment. J. Cell. Mol. Med. 2019, 23, 4111–4117. [Google Scholar] [CrossRef]
- Jakobsen, I.; Sundkvist, M.; Bjorn, N.; Green, H.; Lotfi, K. Early changes in gene expression profiles in AML patients during induction chemotherapy. BMC Genomics 2022, 23, 752. [Google Scholar] [CrossRef]
- Pastor-Anglada, M.; Perez-Torras, S. Emerging Roles of Nucleoside Transporters. Front. Pharmacol. 2018, 9, 606. [Google Scholar] [CrossRef]
- Wood, P.; Burgess, R.; MacGregor, A.; Yin, J.A. P-glycoprotein expression on acute myeloid leukaemia blast cells at diagnosis predicts response to chemotherapy and survival. Br. J. Haematol. 1994, 87, 509–514. [Google Scholar] [CrossRef]
- Doxani, C.; Voulgarelis, M.; Zintzaras, E. MDR1 mRNA expression and MDR1 gene variants as predictors of response to chemotherapy in patients with acute myeloid leukaemia: A meta-analysis. Biomarkers 2013, 18, 425–435. [Google Scholar] [CrossRef]
- Shi, P.; Zha, J.; Guo, X.; Chen, F.; Fan, Z.; Huang, F.; Meng, F.; Liu, X.; Feng, R.; Xu, B. Idarubicin is superior to daunorubicin in remission induction of de novo acute myeloid leukemia patients with high MDR1 expression. Pharmacogenomics 2013, 14, 17–23. [Google Scholar] [CrossRef]
- Kurata, M.; Rathe, S.K.; Bailey, N.J.; Aumann, N.K.; Jones, J.M.; Veldhuijzen, G.W.; Moriarity, B.S.; Largaespada, D.A. Using genome-wide CRISPR library screening with library resistant DCK to find new sources of Ara-C drug resistance in AML. Sci. Rep. 2016, 6, 36199. [Google Scholar] [CrossRef]
- Mahfouz, R.Z.; Jankowska, A.; Ebrahem, Q.; Gu, X.; Visconte, V.; Tabarroki, A.; Terse, P.; Covey, J.; Chan, K.; Ling, Y.; et al. Increased CDA expression/activity in males contributes to decreased cytidine analog half-life and likely contributes to worse outcomes with 5-azacytidine or decitabine therapy. Clin. Cancer Res. 2013, 19, 938–948. [Google Scholar] [CrossRef]
- Schneider, C.; Oellerich, T.; Baldauf, H.M.; Schwarz, S.M.; Thomas, D.; Flick, R.; Bohnenberger, H.; Kaderali, L.; Stegmann, L.; Cremer, A.; et al. SAMHD1 is a biomarker for cytarabine response and a therapeutic target in acute myeloid leukemia. Nat. Med. 2017, 23, 250–255. [Google Scholar] [CrossRef]
- Lohri, A.; van Hille, B.; Bacchi, M.; Fopp, M.; Joncourt, F.; Reuter, J.; Cerny, T.; Fey, M.F.; Herrmann, R. Five putative drug resistance parameters (MDR1/P-glycoprotein, MDR-associated protein, glutathione-S-transferase, bcl-2 and topoisomerase IIalpha) in 57 newly diagnosed acute myeloid leukaemias. Swiss Group for Clinical Cancer Research (SAKK). Eur. J. Haematol. 1997, 59, 206–215. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein/Gen | Primer Forward (5′-3′) | Primer Reverse (5′-3′) | Acccess Number |
|---|---|---|---|
| ENT1 (SLC29A1) | CCATCGATCTGGAGCCCGT | TGTCATGGTGATGGTGTTCTCGGT | NM_001078177 |
| ENT2 (SLC29A2) | CCGCCATCCCGTACTTCCA | TGTTGAAGTTGAAGGCATCCTCGG | NM_001532 |
| CNT1 (SLC28A1) | GCTCTGCACTGGGCTCTCT | TTGAGAAACCTCCTCAGCTTTGGC | NM_004213 |
| CNT2 (SLC28A2) | GGGCTGGAGCTCATGGAAAAAGAA | CCACCGACTCCTCCTCTGGTAA | NM_004212.3 |
| CNT3 (SLC28A3) | GGTTCTGGCTGAAGTGGGTGAT | ACATTATGAGCCCACCGAAGGACA | NM_022127 |
| MDR1 (ABCB1) | GCGCGAGGTCGGAATGGAT | CCATGGATGATGGCAGCCAAAGTT | NM_000927 |
| BCRP (ABCG2) | CCCAGGCCTCTATAGCTCAGATCATT | CACGGCTGAAACACTGCTGAAACA | NM_004827 |
| MRP1 (ABCC1) | CCGCTCTGGGACTGGAATGT | GTGTCATCTGAATGTAGCCTCGGT | NM_004996 |
| MRP4 (ABCC4) | TGCAAGGGTTCTGGGATAAAGA | CTTTGGCACTTTCCTCAATTAACG | NM_005845 |
| MRP5 (ABCC5) | GTTCAGGAGAACTCGACCGTTGG | TTTGGAAGTAGTCCGGATGGGCTT | NM_005688 |
| MRP8 (ABCC8) | CGGTCTCCTTTATTCTCCCACA | AGCCTCTTAAACTGGCTGATGAAGT | NM_032583 |
| CDA (CDA) | GCTATCGCCAGTGACATGCAAGA | AGTTGGTGCCAAACTCTCTCATGACT | NM_001785 |
| DCK (DCK) | GGGAACATCGCTGCAGGGAA | ACAGGTTCAGGAACCACTTCCCA | NM_000788 |
| 5-NT (NT5C2) | AGAAGCCTATCATCGGGTGTTTGTGAA | CATACTCTGGGGACTTGTACACAGCAA | NM_012229 |
| GAPDH (GAPDH) | TGAGCCCGCAGCCTCC | TACGACCAAATCCGTTGACTCC | NM_002046 |
| Pumps | Fluorescent Substrates | Inhibitors |
|---|---|---|
| MDR1 | 3,3′-Diethyloxacarbocyanine iodide (DiOC2(3)) | Verapamil |
| MRP1-2 | Calcein | Probenecid |
| MRP3-5 | Carboxyfluorescein | Probenecid, Diclofenac |
| BCRP | Mitoxantrone | Fumitremorgin C (FTC) |
| Variable | Responders (n = 54) | Non-Responders (n = 13) |
|---|---|---|
| Age, yr | ||
| Median (Range) | 53 (27–75) | 59 (44–63) |
| Female gender, n (%) | 28 (51.8) | 7 (53.8) |
| WBC count per µL | ||
| Median (Range) | 25,100 (900–234,900) | 43,800 (2000–141,000) |
| Platelet count per µL | ||
| Median (Range) | 67,500 (6000–321,000) | 74,000 (25,000–244,000) |
| Bone marrow blasts, % | ||
| Median (Range) | 73 (20–95) | 58 (22–96) |
| Cytogenetic risk | ||
| Favorable, n (%) | 1 (1.9) | 0 (0) |
| Intermediate, n (%) | 35 (64.8) | 10 (76.9) |
| Adverse, n (%) | 18 (33.3) | 3 (23.1) |
| Molecular risk | ||
| NPM1+/FLT3-ITD−, n (%) | 10 (18.5) | 3 (23.1) |
| NPM1+/FLT3-ITD+, n (%) | 8 (14.8) | 3 (23.1) |
| NPM1−/FLT3-ITD−, n (%) | 31 (57.4) | 6 (46.1) |
| NPM1−/FLT3-ITD+, n (%) | 5 (9.3) | 1 (7.7) |
| Cytarabine (nM) | Idarubicin (nM) | |
|---|---|---|
| MOLM-13 | 68.3± 4.2 | 3.5 ± 0.7 |
| HL-60 | 24.4 ± 1.1 | 7.5 ± 0.8 |
| HEL | 60.2 ± 4.6 | 17.2 ± 1.0 |
| K-562 | 5700 ± 624 | 162 ± 21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Macías, G.; Briz, O.; Cives-Losada, C.; Chillón, M.C.; Martínez-Laperche, C.; Martínez-Arranz, I.; Buño, I.; González-Díaz, M.; Díez-Martín, J.L.; Marin, J.J.G.; et al. Role of Intracellular Drug Disposition in the Response of Acute Myeloid Leukemia to Cytarabine and Idarubicin Induction Chemotherapy. Cancers 2023, 15, 3145. https://doi.org/10.3390/cancers15123145
Rodríguez-Macías G, Briz O, Cives-Losada C, Chillón MC, Martínez-Laperche C, Martínez-Arranz I, Buño I, González-Díaz M, Díez-Martín JL, Marin JJG, et al. Role of Intracellular Drug Disposition in the Response of Acute Myeloid Leukemia to Cytarabine and Idarubicin Induction Chemotherapy. Cancers. 2023; 15(12):3145. https://doi.org/10.3390/cancers15123145
Chicago/Turabian StyleRodríguez-Macías, Gabriela, Oscar Briz, Candela Cives-Losada, María C. Chillón, Carolina Martínez-Laperche, Ibon Martínez-Arranz, Ismael Buño, Marcos González-Díaz, José L. Díez-Martín, Jose J. G. Marin, and et al. 2023. "Role of Intracellular Drug Disposition in the Response of Acute Myeloid Leukemia to Cytarabine and Idarubicin Induction Chemotherapy" Cancers 15, no. 12: 3145. https://doi.org/10.3390/cancers15123145
APA StyleRodríguez-Macías, G., Briz, O., Cives-Losada, C., Chillón, M. C., Martínez-Laperche, C., Martínez-Arranz, I., Buño, I., González-Díaz, M., Díez-Martín, J. L., Marin, J. J. G., & Macias, R. I. R. (2023). Role of Intracellular Drug Disposition in the Response of Acute Myeloid Leukemia to Cytarabine and Idarubicin Induction Chemotherapy. Cancers, 15(12), 3145. https://doi.org/10.3390/cancers15123145