
Re-Evaluating the Role of PTHrP in Breast Cancer


{kind=link}
{kind=link}
{kind=link}
Abstract
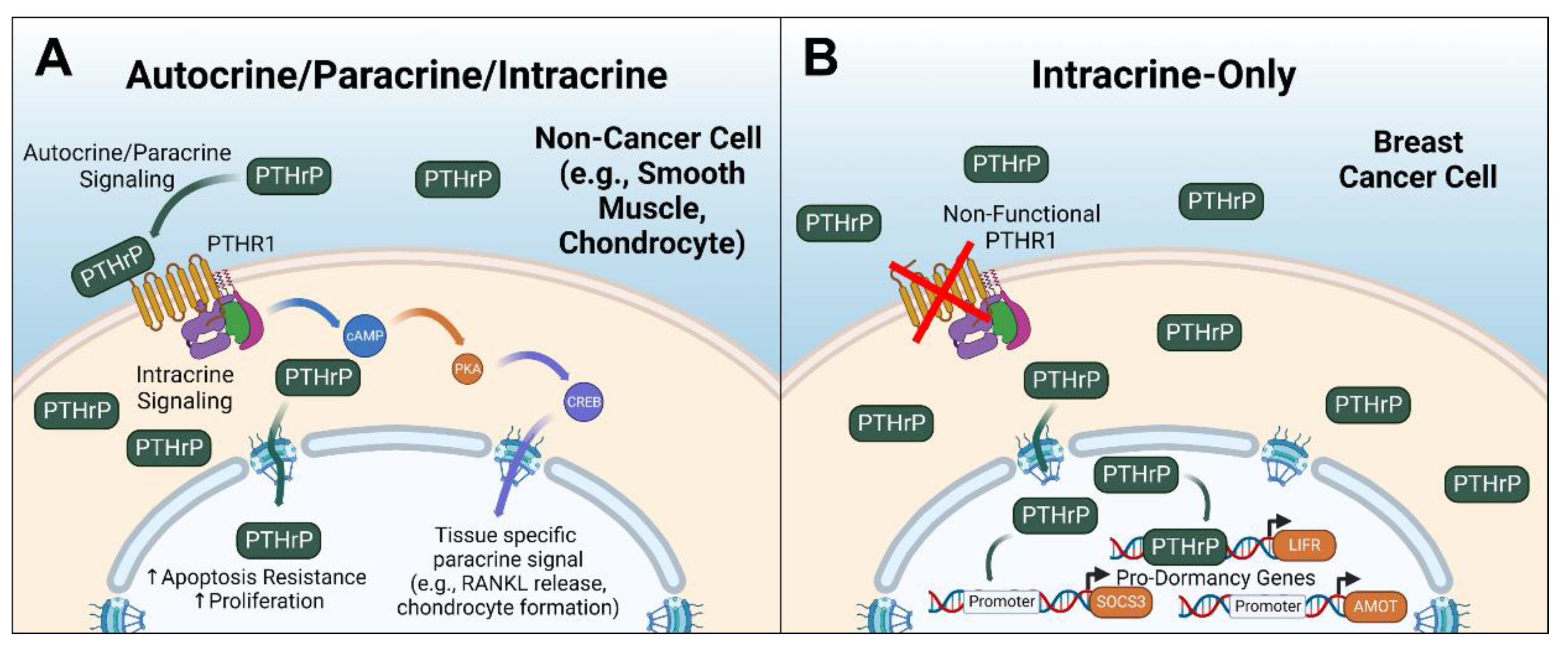
Simple Summary
Abstract
1. Introduction
1.1. A Brief History of Parathyroid Hormone-Related Protein (PTHrP) and the Receptor Binding Model
1.2. Physiologic Functions of PTHrP
1.3. PTHrP and the Vicious Cycle of Bone Destruction
2. Clinical Significance of PTHrP in Breast Cancer: Complications in the Paracrine Model
2.1. PTHrP as a Prognostic Factor
2.2. Inhibiting PTHrP in an Oncology Setting
3. Intracrine PTHrP in Cancer: Mounting Evidence
4. Structural Insights: Reconciling the Intracrine and Paracrine Models
4.1. Nuclear Localization of PTHrP
4.2. The C-Terminal Region of PTHrP
5. Proteolytic Processing of PTHrP: Re-Evaluating the Model in the Context of Breast Cancer
6. Clinical Administration of PTH/PTHrP Analogs in Cancer Patients: Beneficial or Detrimental?
7. Generating a New Model: Unanswered Questions and Future Directions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PTHrP | Parathyroid-hormone-related protein |
| PTH | Parathyroid hormone |
| PTHR1 | Parathyroid hormone receptor 1 |
| cAMP | Cyclic adenosine monophosphate |
| PLC | Phospholipase C |
| Ca2+ | Calcium |
| PKC | Protein kinase C |
| PTHLH | Parathyroid-hormone-like hormone |
| RANK | Receptor activator of nuclear factor kappa–B |
| RANKL | Receptor activator of nuclear factor kappa–B ligand |
| TGF-β | Transforming growth factor beta |
| BMP | Bone morphogenetic protein |
| IGF | Insulin-like growth factor |
| ER | Estrogen receptor |
| MMP | Matrix metalloproteinase |
| LIFR | Leukemia inhibitory factor receptor |
| NLS | Nuclear localization signal |
| VEGFR2 | Vascular endothelial growth factor receptor 2 |
References
- Pioszak, A.A.; Parker, N.R.; Gardella, T.J.; Xu, H.E. Structural basis for parathyroid hormone-related protein binding to the parathyroid hormone receptor and design of conformation-selective peptides. J. Biol. Chem. 2009, 284, 28382–28391. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.J.; Johnson, R.W. Multiple actions of parathyroid hormone-related protein in breast cancer bone metastasis. Br. J. Pharmacol. 2021, 178, 1923–1935. [Google Scholar] [CrossRef] [PubMed]
- Cheloha, R.W.; Gellman, S.H.; Vilardaga, J.P.; Gardella, T.J. PTH receptor-1 signalling-mechanistic insights and therapeutic prospects. Nat. Rev. Endocrinol. 2015, 11, 712–724. [Google Scholar] [CrossRef] [PubMed]
- Albright, F. Case 39061. N. Engl. J. Med. 1953, 248, 248–254. [Google Scholar] [CrossRef]
- Omenn, G.S.; Roth, S.I.; Baker, W.H. Hyperparathyroidism associated with malignant tumors of nonparathyroid origin. Cancer 1969, 24, 1004–1012. [Google Scholar] [CrossRef]
- Burtis, W.J.; Wu, T.; Bunch, C.; Wysolmerski, J.J.; Insogna, K.L.; Weir, E.C.; Broadus, A.E.; Stewart, A.F. Identification of a novel 17,000-dalton parathyroid hormone-like adenylate cyclase-stimulating protein from a tumor associated with humoral hypercalcemia of malignancy. J. Biol. Chem. 1987, 262, 7151–7156. [Google Scholar] [CrossRef]
- Karaplis, A.C.; Luz, A.; Glowacki, J.; Bronson, R.T.; Tybulewicz, V.L.; Kronenberg, H.M.; Mulligan, R.C. Lethal skeletal dysplasia from targeted disruption of the parathyroid hormone-related peptide gene. Genes Dev. 1994, 8, 277–289. [Google Scholar] [CrossRef]
- Weir, E.C.; Philbrick, W.M.; Amling, M.; Neff, L.A.; Baron, R.; Broadus, A.E. Targeted overexpression of parathyroid hormone-related peptide in chondrocytes causes chondrodysplasia and delayed endochondral bone formation. Proc. Natl. Acad. Sci. USA 1996, 93, 10240–10245. [Google Scholar] [CrossRef]
- Miao, D.; He, B.; Jiang, Y.; Kobayashi, T.; Sorocéanu, M.A.; Zhao, J.; Su, H.; Tong, X.; Amizuka, N.; Gupta, A.; et al. Osteoblast-derived PTHrP is a potent endogenous bone anabolic agent that modifies the therapeutic efficacy of administered PTH 1-34. J. Clin. Investig. 2005, 115, 2402–2411. [Google Scholar] [CrossRef]
- VanHouten, J.N.; Dann, P.; Stewart, A.F.; Watson, C.J.; Pollak, M.; Karaplis, A.C.; Wysolmerski, J.J. Mammary-specific deletion of parathyroid hormone–related protein preserves bone mass during lactation. J. Clin. Investig. 2003, 112, 1429–1436. [Google Scholar] [CrossRef]
- Qing, H.; Ardeshirpour, L.; Divieti Pajevic, P.; Dusevich, V.; Jähn, K.; Kato, S.; Wysolmerski, J.; Bonewald, L.F. Demonstration of osteocytic perilacunar/canalicular remodeling in mice during lactation. J. Bone Miner. Res. 2012, 27, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.M.; Sebag, M.; Rhim, J.S.; Kremer, R.; Goltzman, D. Antisense-mediated inhibition of parathyroid hormone-related peptide production in a keratinocyte cell line impedes differentiation. Mol. Endocrinol. 1994, 8, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Massfelder, T.; Dann, P.; Wu, T.L.; Vasavada, R.; Helwig, J.-J.; Stewart, A.F. Opposing mitogenic and anti-mitogenic actions of parathyroid hormone-related protein in vascular smooth muscle cells: A critical role for nuclear targeting. Proc. Natl. Acad. Sci. USA 1997, 94, 13630–13635. [Google Scholar] [CrossRef] [PubMed]
- Sicari, B.M.; Troxell, R.; Salim, F.; Tanwir, M.; Takane, K.K.; Fiaschi-Taesch, N. c-myc and skp2 coordinate p27 degradation, vascular smooth muscle proliferation, and neointima formation induced by the parathyroid hormone-related protein. Endocrinology 2012, 153, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Eilon, G.; Mundy, G.R. Direct resorption of bone by human breast cancer cells in vitro. Nature 1978, 276, 726–728. [Google Scholar] [CrossRef]
- Pfeilschifter, J.; Mundy, G.R. Modulation of type beta transforming growth factor activity in bone cultures by osteotropic hormones. Proc. Natl. Acad. Sci. USA 1987, 84, 2024–2028. [Google Scholar] [CrossRef]
- Yin, J.J.; Selander, K.; Chirgwin, J.M.; Dallas, M.; Grubbs, B.G.; Wieser, R.; Massagué, J.; Mundy, G.R.; Guise, T.A. TGF-beta signaling blockade inhibits PTHrP secretion by breast cancer cells and bone metastases development. J. Clin. Investig. 1999, 103, 197–206. [Google Scholar] [CrossRef]
- Sterling, J.A.; Oyajobi, B.O.; Grubbs, B.; Padalecki, S.S.; Munoz, S.A.; Gupta, A.; Story, B.; Zhao, M.; Mundy, G.R. The hedgehog signaling molecule Gli2 induces parathyroid hormone-related peptide expression and osteolysis in metastatic human breast cancer cells. Cancer Res. 2006, 66, 7548–7553. [Google Scholar] [CrossRef]
- Kozlow, W.; Guise, T.A. Breast Cancer Metastasis to Bone: Mechanisms of Osteolysis and Implications for Therapy. J. Mammary Gland. Biol. Neoplasia 2005, 10, 169–180. [Google Scholar] [CrossRef]
- Edwards, C.M.; Johnson, R.W. From Good to Bad: The Opposing Effects of PTHrP on Tumor Growth, Dormancy, and Metastasis Throughout Cancer Progression. Front. Oncol. 2021, 11, 644303. [Google Scholar] [CrossRef]
- Southby, J.; Kissin, M.W.; Danks, J.A.; Hayman, J.A.; Moseley, J.M.; Henderson, M.A.; Bennett, R.C.; Martin, T.J. Immunohistochemical Localization of Parathyroid Hormone-related Protein in Human Breast Cancer. Cancer Res. 1990, 50, 7710–7716. [Google Scholar]
- Yoshida, A.; Nakamura, Y.; Shimizu, A.; Harada, M.; Kameda, Y.; Nagano, A.; Inaba, M.; Asaga, T. Significance of the parathyroid hormone-related protein expression in breast carcinoma. Breast Cancer 2000, 7, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Powell, G.J.; Southby, J.; Danks, J.A.; Stillwell, R.G.; Hayman, J.A.; Henderson, M.A.; Bennett, R.C.; Martin, T.J. Localization of parathyroid hormone-related protein in breast cancer metastases: Increased incidence in bone compared with other sites. Cancer Res. 1991, 51, 3059–3061. [Google Scholar]
- Henderson, M.A.; Danks, J.A.; Moseley, J.M.; Slavin, J.L.; Harris, T.L.; McKinlay, M.R.; Hopper, J.L.; Martin, T.J. Parathyroid Hormone-Related Protein Production by Breast Cancers, Improved Survival, and Reduced Bone Metastases. J. Natl. Cancer Inst. 2001, 93, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.H.; Utama, F.E.; Sato, T.; Peck, A.R.; Langenheim, J.F.; Udhane, S.S.; Sun, Y.; Liu, C.; Girondo, M.A.; Kovatich, A.J.; et al. Loss of Nuclear Localized Parathyroid Hormone-Related Protein in Primary Breast Cancer Predicts Poor Clinical Outcome and Correlates with Suppressed Stat5 Signaling. Clin. Cancer Res. 2018, 24, 6355–6366. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.J.; Gigola, G.; Zwenger, A.; Carriquiriborde, M.; Gentil, F.; Gentili, C. Potential therapeutic targets for growth arrest of colorectal cancer cells exposed to PTHrP. Mol. Cell Endocrinol. 2018, 478, 32–44. [Google Scholar] [CrossRef]
- Huang, D.C.; Yang, X.F.; Ochietti, B.; Fadhil, I.; Camirand, A.; Kremer, R. Parathyroid Hormone-Related Protein: Potential Therapeutic Target for Melanoma Invasion and Metastasis. Endocrinology 2014, 155, 3739–3749. [Google Scholar] [CrossRef]
- Gallwitz, W.E.; Guise, T.A.; Mundy, G.R. Guanosine nucleotides inhibit different syndromes of PTHrP excess caused by human cancers in vivo. J. Clin. Investig. 2002, 110, 1559–1572. [Google Scholar] [CrossRef]
- Li, J.; Karaplis, A.C.; Huang, D.C.; Siegel, P.M.; Camirand, A.; Yang, X.F.; Muller, W.J.; Kremer, R. PTHrP drives breast tumor initiation, progression, and metastasis in mice and is a potential therapy target. J. Clin. Investig. 2011, 121, 4655–4669. [Google Scholar] [CrossRef]
- Saito, H.; Tsunenari, T.; Onuma, E.; Sato, K.; Ogata, E.; Yamada-Okabe, H. Humanized monoclonal antibody against parathyroid hormone-related protein suppresses osteolytic bone metastasis of human breast cancer cells derived from MDA-MB-231. Anticancer Res. 2005, 25, 3817–3823. [Google Scholar]
- Mayhew, V.; Omokehinde, T.; Johnson, R.W. Tumor dormancy in bone. Cancer Rep. 2020, 3, e1156. [Google Scholar] [CrossRef] [PubMed]
- Reagan, P.; Pani, A.; Rosner, M.H. Approach to diagnosis and treatment of hypercalcemia in a patient with malignancy. Am. J. Kidney Dis. 2014, 63, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.; Finkelstein, D.M.; Barrios, C.; Martin, M.; Iwata, H.; Hegg, R.; Glaspy, J.; Periañez, A.M.; Tonkin, K.; Deleu, I. Adjuvant denosumab in early breast cancer (D-CARE): An international, multicentre, randomised, controlled, phase 3 trial. Lancet Oncol. 2020, 21, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Gnant, M.; Pfeiler, G.; Dubsky, P.C.; Hubalek, M.; Greil, R.; Jakesz, R.; Wette, V.; Balic, M.; Haslbauer, F.; Melbinger, E. Adjuvant denosumab in breast cancer (ABCSG-18): A multicentre, randomised, double-blind, placebo-controlled trial. Lancet 2015, 386, 433–443. [Google Scholar] [CrossRef]
- Gnant, M.; Pfeiler, G.; Steger, G.G.; Egle, D.; Greil, R.; Fitzal, F.; Wette, V.; Balic, M.; Haslbauer, F.; Melbinger-Zeinitzer, E.; et al. Adjuvant denosumab in postmenopausal patients with hormone receptor-positive breast cancer (ABCSG-18): Disease-free survival results from a randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Oncol. 2019, 20, 339–351. [Google Scholar] [CrossRef]
- Lee, A.V.; Oesterreich, S.; Davidson, N.E. MCF-7 Cells—Changing the Course of Breast Cancer Research and Care for 45 Years. J. Natl. Cancer Inst. 2015, 107, djv073. [Google Scholar] [CrossRef]
- Martin, T.; Findlay, D.; MacIntyre, I.; Eisman, J.; Michelangeli, V.; Moseley, J.; Partridge, N. Calcitonin receptors in a cloned human breast cancer cell line (MCF 7). Biochem. Biophys. Res. Commun. 1980, 96, 150–156. [Google Scholar] [CrossRef]
- Johnson, R.W.; Sun, Y.; Ho, P.W.; Chan, A.S.; Johnson, J.A.; Pavlos, N.J.; Sims, N.A.; Martin, T.J. Parathyroid hormone-related protein negatively regulates tumor cell dormancy genes in a PTHR1/cyclic AMP-independent manner. Front. Endocrinol. 2018, 9, 241. [Google Scholar] [CrossRef]
- Johnson, R.W.; Finger, E.C.; Olcina, M.M.; Vilalta, M.; Aguilera, T.; Miao, Y.; Merkel, A.R.; Johnson, J.R.; Sterling, J.A.; Wu, J.Y. Induction of LIFR confers a dormancy phenotype in breast cancer cells disseminated to the bone marrow. Nat. Cell Biol. 2016, 18, 1078–1089. [Google Scholar] [CrossRef]
- Edwards, C.M.; Clements, M.E.; Vecchi, L.A., 3rd; Johnson, J.A.; Johnson, R.W. HDAC inhibitors stimulate LIFR when it is repressed by hypoxia or PTHrP in breast cancer. J. Bone Oncol. 2021, 31, 100407. [Google Scholar] [CrossRef]
- Grinman, D.Y.; Boras-Granic, K.; Takyar, F.M.; Dann, P.; Hens, J.R.; Marmol, C.; Lee, J.; Choi, J.; Chodosh, L.A.; Sola, M.E.G.; et al. PTHrP induces STAT5 activation, secretory differentiation and accelerates mammary tumor development. Breast Cancer Res. 2022, 24, 30. [Google Scholar] [CrossRef] [PubMed]
- Guise, T.A.; Yin, J.J.; Thomas, R.J.; Dallas, M.; Cui, Y.; Gillespie, M.T. Parathyroid hormone-related protein (PTHrP)-(1-139) isoform is efficiently secreted in vitro and enhances breast cancer metastasis to bone in vivo. Bone 2002, 30, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.J. Parathyroid Hormone-Related Protein, Its Regulation of Cartilage and Bone Development, and Role in Treating Bone Diseases. Physiol. Rev. 2016, 96, 831–871. [Google Scholar] [CrossRef] [PubMed]
- Mangin, M.; Webb, A.C.; Dreyer, B.E.; Posillico, J.T.; Ikeda, K.; Weir, E.C.; Stewart, A.F.; Bander, N.H.; Milstone, L.; Barton, D.E.; et al. Identification of a cDNA encoding a parathyroid hormone-like peptide from a human tumor associated with humoral hypercalcemia of malignancy. Proc. Natl. Acad. Sci. USA 1988, 85, 597–601. [Google Scholar] [CrossRef]
- Hastings, R.H.; Asirvatham, A.; Quintana, R.; Sandoval, R.; Dutta, R.; Burton, D.W.; Deftos, L.J. Parathyroid hormone-related protein-(38–64) regulates lung cell proliferation after silica injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 283, L12–L21. [Google Scholar] [CrossRef]
- Wu, T.L.; Vasavada, R.C.; Yang, K.; Massfelder, T.; Ganz, M.; Abbas, S.K.; Care, A.D.; Stewart, A.F. Structural and physiologic characterization of the mid-region secretory species of parathyroid hormone-related protein. J. Biol. Chem. 1996, 271, 24371–24381. [Google Scholar] [CrossRef]
- Fenton, A.J.; Kemp, B.E.; Hammonds, R.G., Jr.; Mitchelhill, K.; Moseley, J.M.; Martin, T.J.; Nicholson, G.C. A potent inhibitor of osteoclastic bone resorption within a highly conserved pentapeptide region of parathyroid hormone-related protein; PTHrP[107-111]. Endocrinology 1991, 129, 3424–3426. [Google Scholar] [CrossRef]
- Lam, M.H.; Briggs, L.J.; Hu, W.; Martin, T.J.; Gillespie, M.T.; Jans, D.A. Importin beta recognizes parathyroid hormone-related protein with high affinity and mediates its nuclear import in the absence of importin alpha. J. Biol. Chem. 1999, 274, 7391–7398. [Google Scholar] [CrossRef]
- Kumari, R.; Robertson, J.F.; Watson, S.A. Nuclear targeting of a midregion PTHrP fragment is necessary for stimulating growth in breast cancer cells. Int. J. Cancer 2006, 119, 49–59. [Google Scholar] [CrossRef]
- Aarts, M.M.; Rix, A.; Guo, J.; Bringhurst, R.; Henderson, J.E. The nucleolar targeting signal (NTS) of parathyroid hormone related protein mediates endocytosis and nucleolar translocation. J. Bone Miner. Res. 1999, 14, 1493–1503. [Google Scholar] [CrossRef]
- Henderson, J.E.; Amizuka, N.; Warshawsky, H.; Biasotto, D.; Lanske, B.M.; Goltzman, D.; Karaplis, A.C. Nucleolar localization of parathyroid hormone-related peptide enhances survival of chondrocytes under conditions that promote apoptotic cell death. Mol. Cell Biol. 1995, 15, 4064–4075. [Google Scholar] [CrossRef] [PubMed]
- Massfelder, T.; Fiaschi-Taesch, N.; Stewart, A.F.; Helwig, J.J. Parathyroid hormone-related peptide—A smooth muscle tone and proliferation regulatory protein. Curr. Opin. Nephrol. Hypertens. 1998, 7, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, R.M.; Kemp, B.E.; Barden, J.A. Structure study of osteostatin PTHrP[Thr107](107-139). Biochim. Biophys. Acta 1999, 1432, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Portal-Núñez, S.; Ardura, J.A.; Lozano, D.; Martínez de Toda, I.; De la Fuente, M.; Herrero-Beaumont, G.; Largo, R.; Esbrit, P. Parathyroid hormone-related protein exhibits antioxidant features in osteoblastic cells through its N-terminal and osteostatin domains. Bone Joint Res. 2018, 7, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Alonso, V.; de Gortázar, A.R.; Ardura, J.A.; Andrade-Zapata, I.; Alvarez-Arroyo, M.V.; Esbrit, P. Parathyroid hormone-related protein (107–139) increases human osteoblastic cell survival by activation of vascular endothelial growth factor receptor-2. J. Cell Physiol. 2008, 217, 717–727. [Google Scholar] [CrossRef]
- de Gortázar, A.R.; Alonso, V.; Alvarez-Arroyo, M.V.; Esbrit, P. Transient exposure to PTHrP (107–139) exerts anabolic effects through vascular endothelial growth factor receptor 2 in human osteoblastic cells in vitro. Calcif. Tissue Int. 2006, 79, 360–369. [Google Scholar] [CrossRef]
- Valín, A.; García-Ocaña, A.; De Miguel, F.; Sarasa, J.L.; Esbrit, P. Antiproliferative effect of the C-terminal fragments of parathyroid hormone-related protein, PTHrP-(107–111) and (107–139), on osteoblastic osteosarcoma cells. J. Cell Physiol. 1997, 170, 209–215. [Google Scholar] [CrossRef]
- Liu, B.; Goltzman, D.; Rabbani, S.A. Processing of pro-PTHRP by the prohormone convertase, furin: Effect on biological activity. Am. J. Physiol. 1995, 268, E832–E838. [Google Scholar] [CrossRef]
- Hook, V.Y.; Burton, D.; Yasothornsrikul, S.; Hastings, R.H.; Deftos, L.J. Proteolysis of ProPTHrP (1–141) by “prohormone thiol protease” at multibasic residues generates PTHrP-related peptides: Implications for PTHrP peptide production in lung cancer cells. Biochem. Biophys. Res. Commun. 2001, 285, 932–938. [Google Scholar] [CrossRef]
- Ruchon, A.F.; Marcinkiewicz, M.; Ellefsen, K.; Basak, A.; Aubin, J.; Crine, P.; Boileau, G. Cellular localization of neprilysin in mouse bone tissue and putative role in hydrolysis of osteogenic peptides. J. Bone Miner. Res. 2000, 15, 1266–1274. [Google Scholar] [CrossRef]
- Iwamura, M.; Hellman, J.; Cockett, A.T.; Lilja, H.; Gershagen, S. Alteration of the hormonal bioactivity of parathyroid hormone-related protein (PTHrP) as a result of limited proteolysis by prostate-specific antigen. Urology 1996, 48, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Kamalakar, A.; Washam, C.L.; Akel, N.S.; Allen, B.J.; Williams, D.K.; Swain, F.L.; Leitzel, K.; Lipton, A.; Gaddy, D.; Suva, L.J. PTHrP (12–48) Modulates the Bone Marrow Microenvironment and Suppresses Human Osteoclast Differentiation and Lifespan. J. Bone Miner. Res. 2017, 32, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Oda, K.; Ikeda, M.; Tsuji, E.; Sohda, M.; Takami, N.; Misumi, Y.; Ikehara, Y. Sequence requirements for proteolytic cleavage of precursors with paired basic amino acids. Biochem. Biophys. Res. Commun. 1991, 179, 1181–1186. [Google Scholar] [CrossRef] [PubMed]
- López, J.J.; Siegfried, G.; Cantonero, C.; Soulet, F.; Descarpentrie, J.; Smani, T.; Badiola, I.; Pernot, S.; Evrard, S.; Rosado, J.A.; et al. Furin Prodomain ppFurin Enhances Ca(2+) Entry Through Orai and TRPC6 Channels’ Activation in Breast Cancer Cells. Cancers 2021, 13, 1670. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Khatib, A.M.; Creemers, J.W.M. Loss of Proprotein Convertase Furin in Mammary Gland Impairs proIGF1R and proIR Processing and Suppresses Tumorigenesis in Triple Negative Breast Cancer. Cancers 2020, 12, 2686. [Google Scholar] [CrossRef] [PubMed]
- Suehiro, M.; Murakami, M.; Fukuchi, M. Circulating forms of immunoreactive parathyroid hormone-related protein for identifying patients with humoral hypercalcemia of malignancy: A comparative study with C-terminal(109–141)- and N-terminal(1–86)-region-specific PTHrP radioassay. Ann. Nucl. Med. 1994, 8, 231–237. [Google Scholar] [CrossRef]
- Washam, C.L.; Byrum, S.D.; Leitzel, K.; Ali, S.M.; Tackett, A.J.; Gaddy, D.; Sundermann, S.E.; Lipton, A.; Suva, L.J. Identification of PTHrP (12–48) as a Plasma Biomarker Associated with Breast Cancer Bone MetastasisPTHrP (12–48) and Bone Metastasis. Cancer Epidemiol. Biomark. Prev. 2013, 22, 972–983. [Google Scholar] [CrossRef]
- Leder, B.Z.; O’Dea, L.S.L.; Zanchetta, J.R.; Kumar, P.; Banks, K.; McKay, K.; Lyttle, C.R.; Hattersley, G. Effects of Abaloparatide, a Human Parathyroid Hormone-Related Peptide Analog, on Bone Mineral Density in Postmenopausal Women with Osteoporosis. J. Clin. Endocrinol. Metab. 2015, 100, 697–706. [Google Scholar] [CrossRef]
- Waqas, K.; Lima Ferreira, J.; Tsourdi, E.; Body, J.J.; Hadji, P.; Zillikens, M.C. Updated guidance on the management of cancer treatment-induced bone loss (CTIBL) in pre- and postmenopausal women with early-stage breast cancer. J. Bone Oncol. 2021, 28, 100355. [Google Scholar] [CrossRef]
- Swami, S.; Johnson, J.; Bettinson, L.A.; Kimura, T.; Zhu, H.; Albertelli, M.A.; Johnson, R.W.; Wu, J.Y. Prevention of breast cancer skeletal metastases with parathyroid hormone. JCI Insight 2017, 2, e90874. [Google Scholar] [CrossRef]
- Brown, H.K.; Allocca, G.; Ottewell, P.D.; Wang, N.; Brown, N.J.; Croucher, P.I.; Eaton, C.L.; Holen, I. Parathyroid Hormone (PTH) Increases Skeletal Tumour Growth and Alters Tumour Distribution in an In Vivo Model of Breast Cancer. Int. J. Mol. Sci. 2018, 19, 2920. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.D.; Hattersley, G.; Riis, B.J.; Williams, G.C.; Lau, E.; Russo, L.A.; Alexandersen, P.; Zerbini, C.A.; Hu, M.Y.; Harris, A.G.; et al. Effect of Abaloparatide vs. Placebo on New Vertebral Fractures in Postmenopausal Women with Osteoporosis: A Randomized Clinical Trial. JAMA 2016, 316, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Neer, R.M.; Arnaud, C.D.; Zanchetta, J.R.; Prince, R.; Gaich, G.A.; Reginster, J.Y.; Hodsman, A.B.; Eriksen, E.F.; Ish-Shalom, S.; Genant, H.K.; et al. Effect of parathyroid hormone (1–34) on fractures and bone mineral density in postmenopausal women with osteoporosis. N. Engl. J. Med. 2001, 344, 1434–1441. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kane, J.F.; Johnson, R.W. Re-Evaluating the Role of PTHrP in Breast Cancer. Cancers 2023, 15, 2670. https://doi.org/10.3390/cancers15102670
Kane JF, Johnson RW. Re-Evaluating the Role of PTHrP in Breast Cancer. Cancers. 2023; 15(10):2670. https://doi.org/10.3390/cancers15102670
Chicago/Turabian StyleKane, Jeremy F., and Rachelle W. Johnson. 2023. "Re-Evaluating the Role of PTHrP in Breast Cancer" Cancers 15, no. 10: 2670. https://doi.org/10.3390/cancers15102670
APA StyleKane, J. F., & Johnson, R. W. (2023). Re-Evaluating the Role of PTHrP in Breast Cancer. Cancers, 15(10), 2670. https://doi.org/10.3390/cancers15102670