
MiR-150-5p Overexpression in Triple-Negative Breast Cancer Contributes to the In Vitro Aggressiveness of This Breast Cancer Subtype

 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Samples
2.2. Breast Cell Lines
2.3. RNA Isolation
2.4. Real-Time Quantitative PCR (RT-qPCR)
2.5. Reverse Transfection Assays
2.6. Cell Proliferation Assay
2.7. Clonogenic Assay
2.8. Wound Healing Assay
2.9. Cytotoxic Assays
2.10. Western Blotting
2.11. The Cancer Genome Atlas Data Processing and Analysis
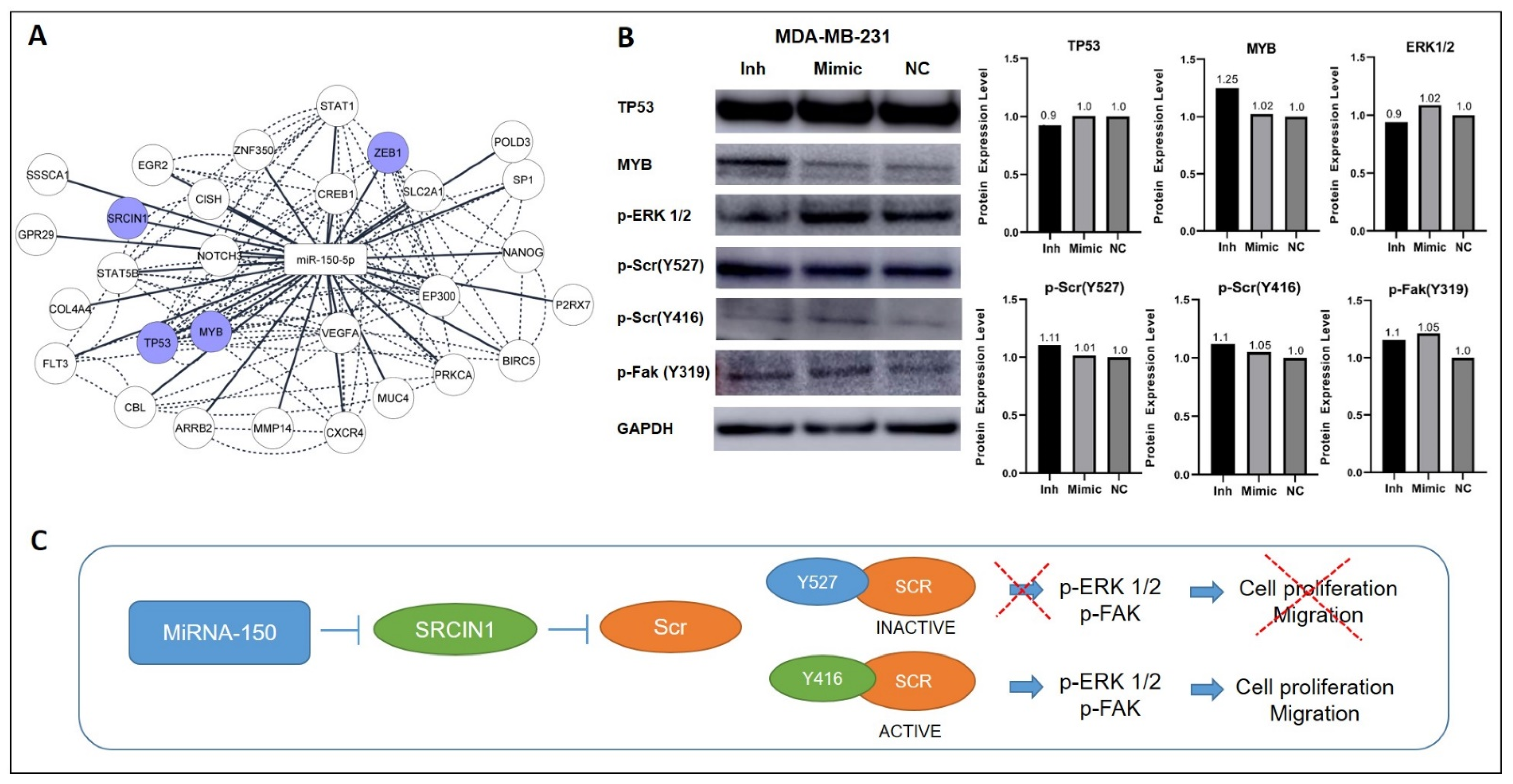
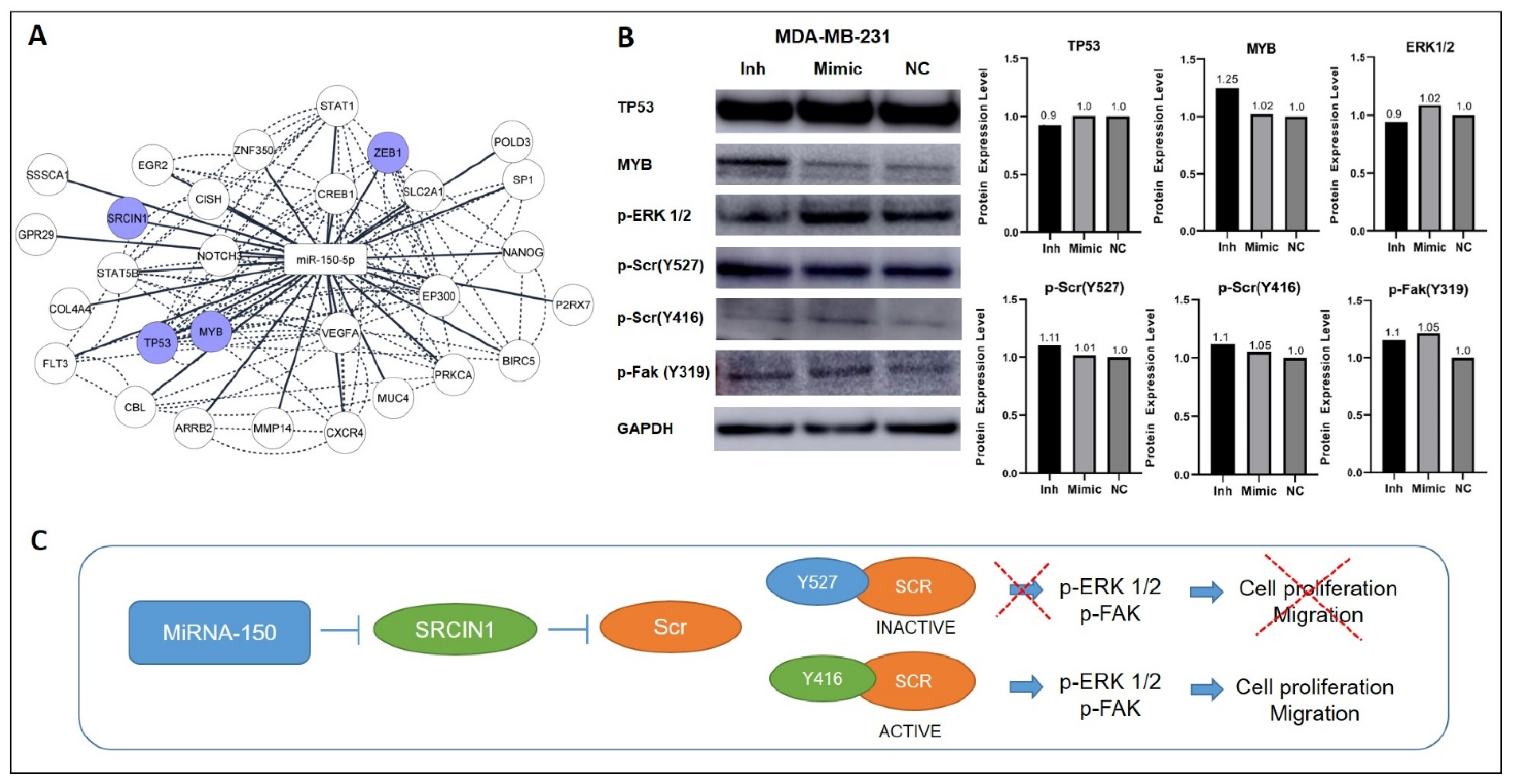
2.12. Network of miR-150-5p and Functionally Validated Target mRNAs
2.13. Statistical Analysis
3. Results
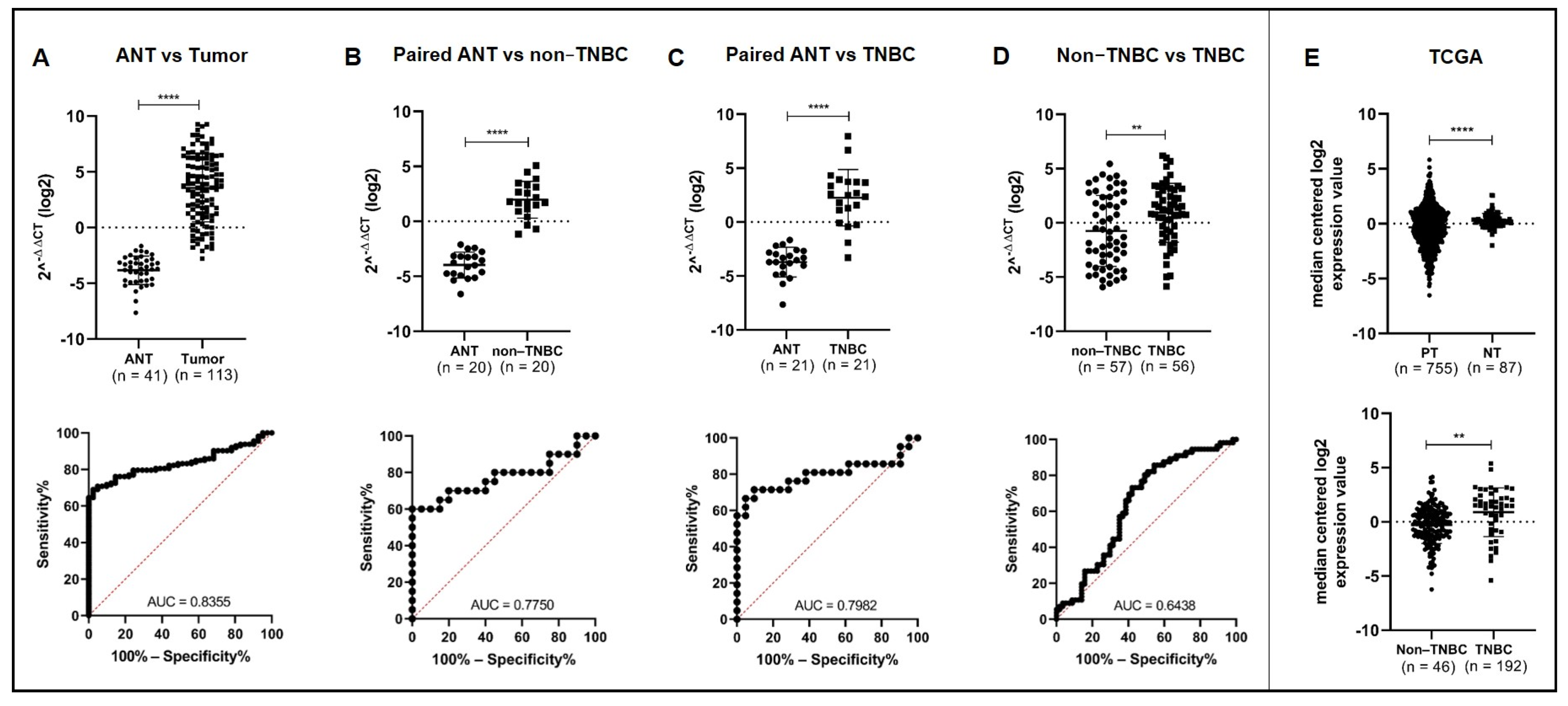
3.1. MiR-150-5p Is Significantly Upregulated in Tumor Breast Tissues and in TNBC
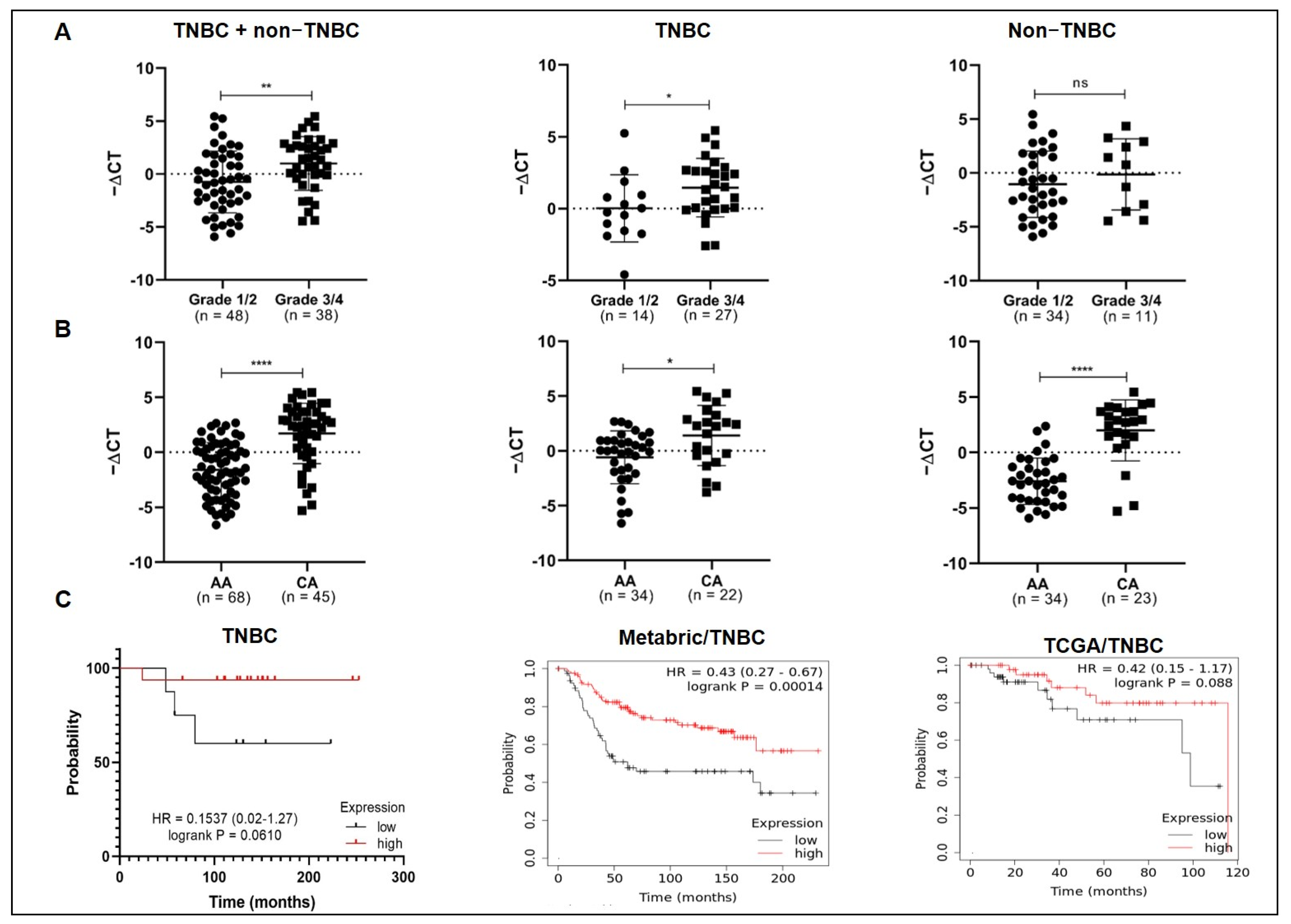
3.2. MiR-150-5p Expression Levels Are Associated with Tumor Grade, Ethnicity, and Overall Survival in TNBC
3.3. MiR-150-5p Is Upregulated in TNBC Cell Lines Compared with Non-TNBC and Non-Tumor Breast Cell Lines
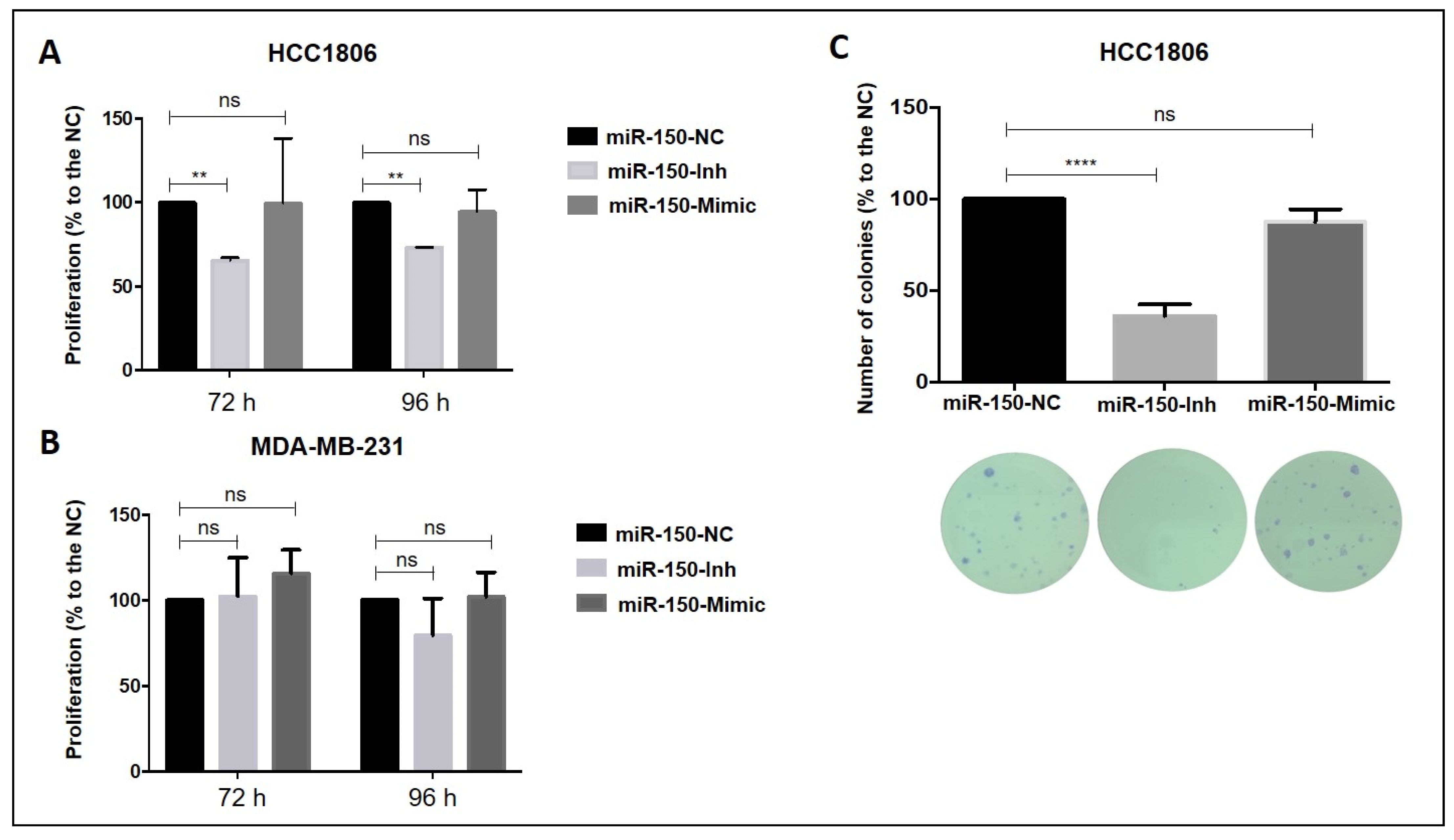
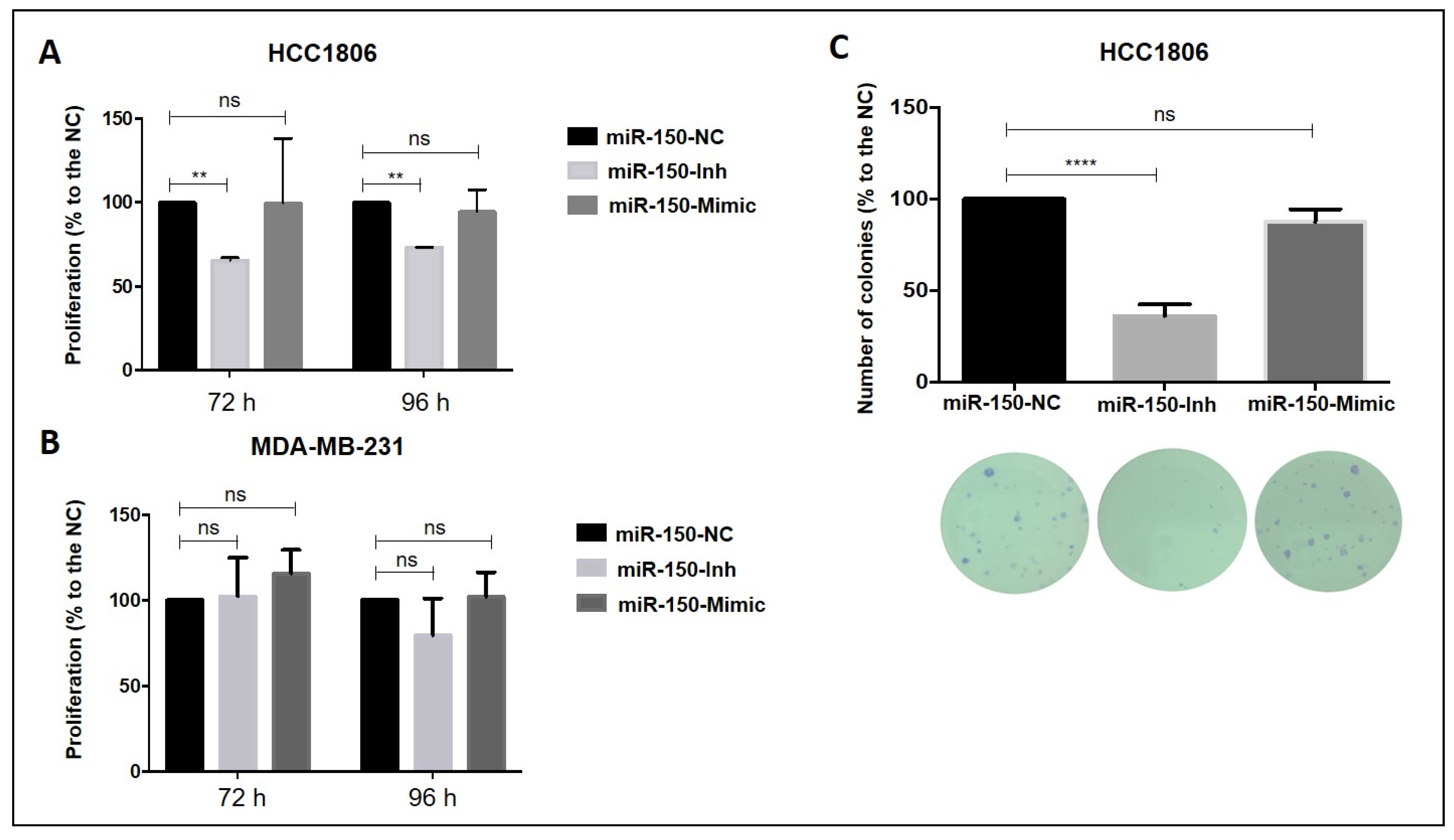
3.4. Inhibition of MiR-150-5p Expression Reduces Cell Proliferation and Clonogenic Growth in TNBC Cells
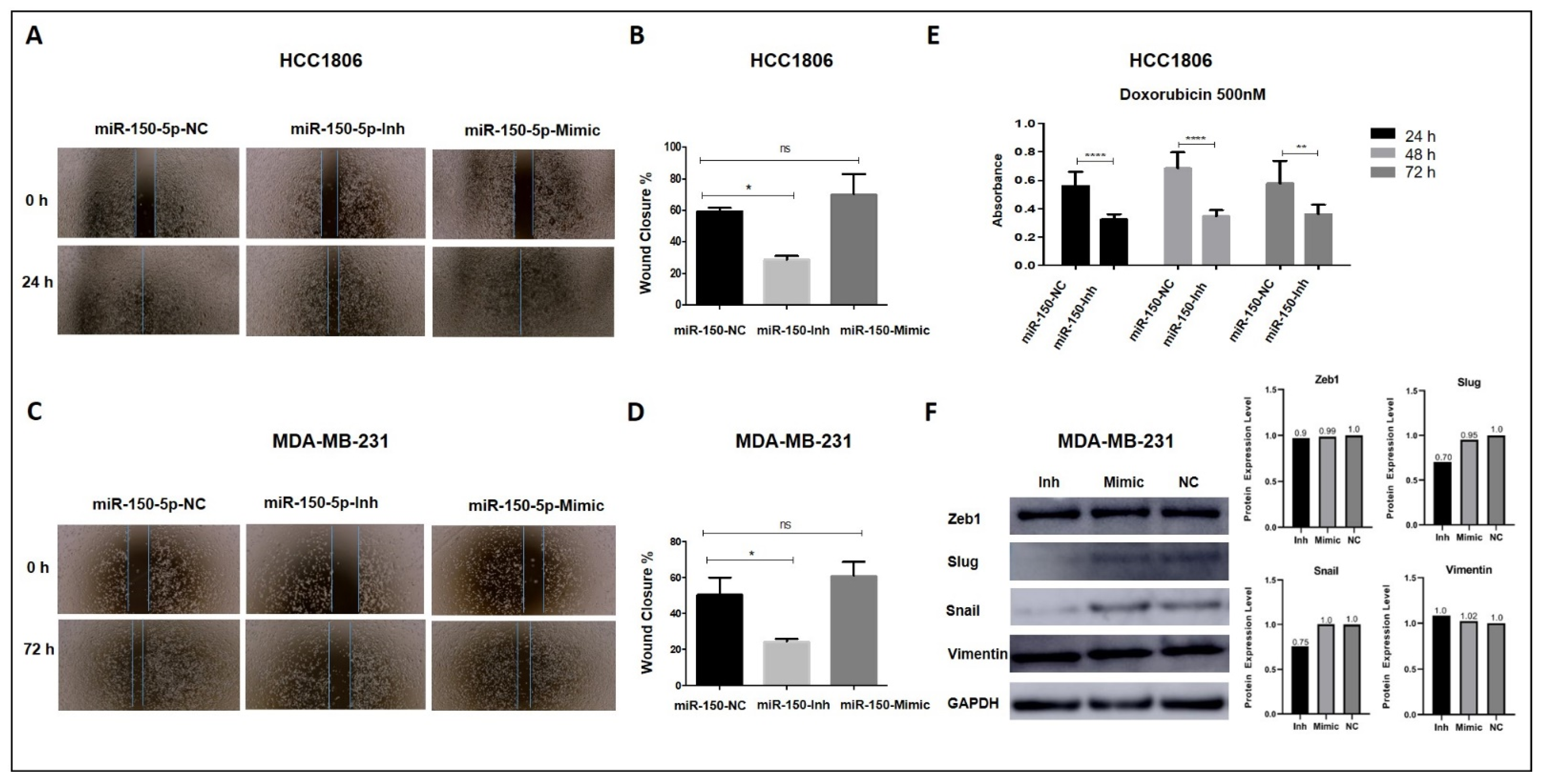
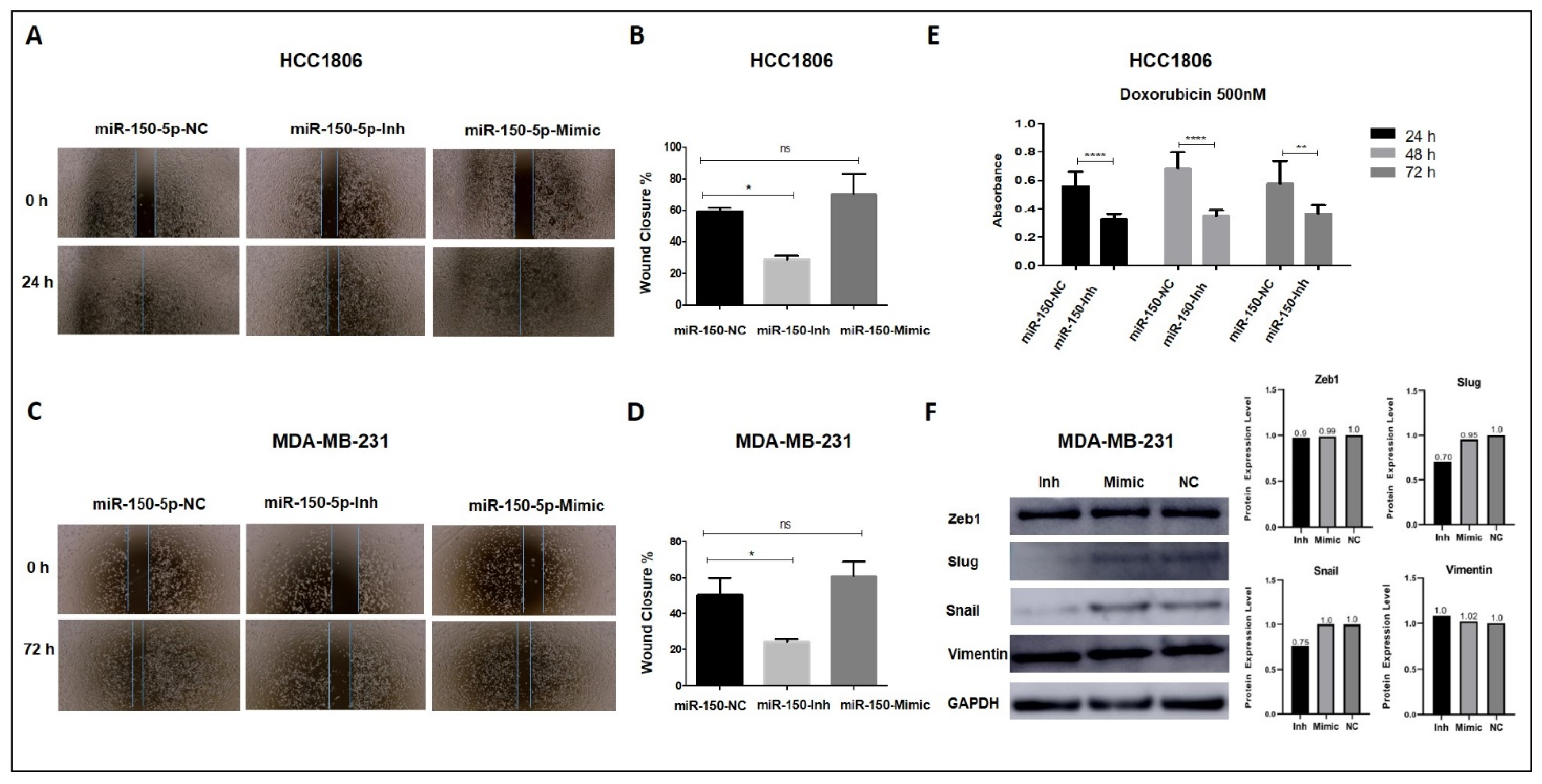
3.5. Inhibition of MiR-150-5p Expression Reduces Cell Migration and Resistance to Doxorubicin in TNBC Cells
3.6. Inhibition of MiR-150-5p Expression Reduces the Expression of MYB and Members of the ERK1/2 Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, R.; Gianni, L. Hallmarks of triple negative breast cancer emerging at last? Cell Res. 2014, 24, 904–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Roosbroeck, K.; Calin, G.A. Cancer Hallmarks and MicroRNAs: The Therapeutic Connection. Adv. Cancer Res. 2017, 135, 119–149. [Google Scholar] [PubMed]
- Ibragimova, M.K.; Tsyganov, M.M.; Litviakov, N.V. Molecular-genetic portrait of breast cancer with triple negative phenotype. Cancers 2021, 13, 5348. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C.M. The role of microRNAs in human cancer. Signal. Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [Green Version]
- Syeda, Z.A.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory mechanism of microrna expression in cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef] [Green Version]
- Dragomir, M.P.; Knutsen, E.; Calin, G.A. Classical and noncanonical functions of miRNAs in cancers. Trends Genet. 2021, 38, 379–394. [Google Scholar] [CrossRef]
- Arghiani, N.; Shah, K. Modulating microRNAs in cancer: Next-generation therapies. Cancer Biol. Med. 2021, 19, 289–304. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—Challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- He, Y.; Jiang, X.; Chen, J. The role of miR-150 in normal and malignant hematopoiesis. Oncogene 2014, 33, 3887–3893. [Google Scholar] [CrossRef]
- Yokobori, T.; Suzuki, S.; Tanaka, N.; Inose, T.; Sohda, M.; Sano, A.; Sakai, M.; Nakajima, M.; Miyazaki, T.; Kato, H.; et al. MiR-150 is associated with poor prognosis in esophageal squamous cell carcinoma via targeting the EMT inducer ZEB1. Cancer Sci. 2013, 104, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yang, Y.; Zhang, P.; Wang, F.; Ma, Y.; Qin, H.; Wang, Y. miR-150 functions as a tumour suppressor in human colorectal cancer by targeting c-Myb. J. Cell. Mol. Med. 2014, 18, 2125–2134. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-H.; Chen, J.; Zhao, F.; Zhang, B.-R.; Yu, H.-S.; Jin, H.-Y.; Dai, J.-H. MiR-150-5p suppresses colorectal cancer cell migration and invasion through targeting MUC4. Asian. Pac. J. Cancer Prev. 2014, 15, 6269–6273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as Biomarkers and Diagnostics. J. Cell. Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, F.; Lin, B.; Luo, H.; Liu, M.; Wu, J.; Li, C.; Li, R.; Zhang, X.; Zhou, K.; et al. MiR-150 inhibits proliferation and tu-morigenicity via retarding G1/S phase transition in nasopharyngeal carcinoma. Int. J. Oncol. 2017, 50, 1097–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zeng, K.; Xu, M.; Hu, X.; Liu, X.; Xu, T.; He, B.; Pan, Y.; Sun, H.; Wang, S. SP1-induced lncRNA-ZFAS1 contributes to colorectal cancer progression via the miR-150-5p/VEGFA axis. Cell Death Dis. 2018, 9, 982. [Google Scholar] [CrossRef] [PubMed]
- Oboshi, W.; Hayashi, K.; Takeuchi, H.; Ikeda, K.; Yamaguchi, Y.; Kimura, A.; Nakamura, T.; Yukimasa, N. MicroRNA-150 suppresses p27Kip1 expression and promotes cell proliferation in HeLa human cervical cancer cells. Oncol. Lett. 2020, 20, 210. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, X.; Huang, J.; Xu, Z.; Xing, C.; Yang, J.; Zhou, X. MicroRNA hsa-miR-150-5p inhibits nasopharyngeal carcino-genesis by suppressing PYCR1 (pyrroline-5-carboxylate reductase 1). Bioengineered 2021, 12, 9766–9778. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, X.; Wan, M.; Zeng, X.; Luo, J.; Xu, Y.; Ji, L.; Zhang, J.A.; Fan, P.; Zhong, J.; et al. FoxP3-miR-150-5p/3p suppresses ovarian tumorigenesis via an IGF1R/IRS1 pathway feedback loop. Cell Death Dis. 2021, 12, 275. [Google Scholar] [CrossRef]
- Cao, M.; Hou, D.; Liang, H.; Gong, F.; Wang, Y.; Yan, X.; Jiang, X.; Wang, C.; Zhang, J.; Zen, K.; et al. MiR-150 promotes the proliferation and migration of lung cancer cells by targeting SRC kinase signalling inhibitor 1. Eur. J. Cancer 2014, 50, 1013–1024. [Google Scholar] [CrossRef]
- Xiao, G.; Wang, P.; Zheng, X.; Liu, D.; Sun, X. FAM83A-AS1 promotes lung adenocarcinoma cell migration and invasion by targeting miR-150-5p and modifying MMP14. Cell Cycle 2019, 18, 2972–2985. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Li, W.; Li, J.; Zhang, Y.; Zhang, X.; Xu, Y.; Hu, Y.; Li, Q.; Sun, Q.; Ma, Z. Higher expression of miR-150-5p promotes tumorigenesis by suppressing LKB1 in non-small cell lung cancer. Pathol. Res. Pract. 2020, 216, 153145. [Google Scholar] [CrossRef] [PubMed]
- Quan, X.; Chen, D.; Li, M.; Chen, X.; Huang, M. MicroRNA-150-5p and SRC kinase signaling inhibitor 1 involvement in the pathological development of gastric cancer. Exp. Ther. Med. 2019, 18, 2667–2674. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Jin, H.; Yang, Z.; Luo, G.; Lu, Y.; Li, K.; Ren, G.; Su, T.; Pan, Y.; Feng, B.; et al. MiR-150 promotes gastric cancer pro-liferation by negatively regulating the pro-apoptotic gene EGR2. Biochem. Biophys. Res. Commun. 2010, 392, 340–345. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, P.; Cao, L.; Li, F.; Duan, S.; Yuan, G.; Xiao, L.; Guo, L.; Yin, H.; Xie, D.; et al. Long intergenic non-coding rna 01121 promotes breast cancer cell proliferation, migration, and invasion via the mir-150-5p/ HMGA2 axis. Cancer Manag. Res. 2019, 11, 10859–10870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; Wu, D.; Zhang, Z.; Li, S. Regulatory effect of the MAFG-AS1/miR-150-5p/MYB axis on the proliferation and migration of breast cancer cells. Int. J. Oncol. 2021, 58, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Cascione, L.; Gasparini, P.; Lovat, F.; Carasi, S.; Pulvirenti, A.; Ferro, A.; Alder, H.; He, G.; Vecchione, A.; Croce, C.M.; et al. Integrated MicroRNA and mRNA Signatures Associated with Survival in Triple Negative Breast Cancer. PLoS ONE 2013, 8, e55910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Q.; Guo, Z.; Qian, H. Role of microRNA-150-5p/SRCIN1 axis in the progression of breast cancer. Exp. Ther. Med. 2019, 17, 2221–2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.; Xu, P.; Wang, H.; Niu, Z.; Zhu, D.; Lin, Q.; Tang, L.; Ren, L. MicroRNA-150 suppresses triple-negative breast cancer metastasis through targeting HMGA2. Onco. Targets. Ther. 2018, 11, 2319–2332. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Chen, Y.; Wu, W.; Ouyang, N.; Chen, J.; Li, H.; Liu, X.; Su, F.; Lin, L.; Yao, Y. MiR-150 promotes human breast cancer growth and malignant behavior by targeting the pro-apoptotic purinergic P2X7 receptor. PLoS ONE 2013, 8, e80707. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Qiu, N.; Xia, H.; Liang, H.; Li, H.; Ao, X. Long non-coding RNA FOXD2-AS1/miR-150-5p/PFN2 axis regulates breast cancer malignancy and tumorigenesis. Int. J. Oncol. 2019, 54, 1043–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Liu, Y.; Du, Y.; Cheng, T.; Xia, W. Long non-coding RNA BLACAT1 promotes breast cancer cell proliferation and metastasis by miR-150-5p/CCR2. Cell Biosci. 2019, 9, 14. [Google Scholar] [CrossRef]
- Wolff, A.C.; Hammond, M.E.H.; Schwartz, J.N.; Hagerty, K.L.; Allred, D.C.; Cote, R.J.; Dowsett, M.; Fitzgibbons, P.L.; Hanna, W.M.; Langer, A.; et al. American Society of Clinical Oncology/College of American Pathologists guideline recommendations for human epidermal growth factor receptor 2 testing in breast cancer. J. Clin. Oncol. 2007, 25, 118–145. [Google Scholar] [CrossRef] [Green Version]
- Hammond, M.E.H.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer (unabridged version). Arch. Pathol. Lab. Med. 2010, 134, e48–e72. [Google Scholar] [CrossRef]
- International Cell Line Authentication Committee (ICLAC) Guidelines. Available online: https://iclac.org/resources/human-cell-line-authentication/ (accessed on 1 June 2018).
- Torresan, C.; Oliveira, M.M.C.; Pereira, S.R.F.; Ribeiro, E.M.S.F.; Marian, C.; Gusev, Y.; Lima, R.S.; Urban, C.A.; Berg, P.E.; Haddad, B.R.; et al. Increased copy number of the DLX4 homeobox gene in breast axillary lymph node metastasis. Cancer Genet. 2014, 207, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.F.; Cólus, I.M.S.; Fonseca, A.S.; Antunes, V.C.; Kumar, D.; Cavalli, L.R. MiR-182-5p Modulates Prostate Cancer Aggressive Phenotypes by Targeting EMT Associated Pathways. Biomolecules 2022, 12, 187. [Google Scholar] [CrossRef] [PubMed]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; Van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research Techniques Made Simple: Analysis of Collective Cell Migration Using the Wound Healing Assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef] [Green Version]
- Matsiaka, O.M.; Penington, C.J.; Baker, R.E.; Simpson, M.J. Discrete and Continuum Approximations for Collective Cell Mi-gration in a Scratch Assay with Cell Size Dynamics. Bull. Math. Biol. 2018, 80, 738–757. [Google Scholar] [CrossRef] [PubMed]
- Varankar, S.S.; Bapat, S.A. Migratory metrics of wound healing: A Quantification Approach for in vitro Scratch Assays. Front. Oncol. 2018, 8, 633. [Google Scholar] [CrossRef] [PubMed]
- Somchai, P.; Phongkitkarun, K.; Kueanjinda, P.; Jamnongsong, S.; Vaeteewoottacharn, K.; Luvira, V.; Okada, S.; Jirawatnotai, S.; Sampattavanich, S. Novel Analytical Platform For Robust Identification of Cell Migration Inhibitors. Sci. Rep. 2020, 10, 931. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- GDAC Fire Browse TCGA Portal. Available online: http://firebrowse.org/ (accessed on 17 February 2022).
- Kaplan Meier Plotter. Available online: http://kmplot.com/analysis/ (accessed on 1 February 2022).
- miRTargetLink 2.0. Available online: https://ccb-compute.cs.uni-saarland.de/mirtargetlink2 (accessed on 16 February 2022).
- STRING Tool. Available online: https://string-db.org (accessed on 16 February 2022).
- Shanon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikoswski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Lü, L.; Mao, X.; Shi, P.; He, B.; Xu, K.; Zhang, S.; Wang, J. MicroRNAs in the prognosis of triple-negative breast cancer. Medicine 2017, 96, e7085. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, D.; Braun, M.; Kordek, R.; Sadej, R.; Romanska, H. MicroRNAs in regulation of triple-negative breast cancer pro-gression. J. Cancer Res. Clin. Oncol. 2018, 144, 1401–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Gu, H.; Xiong, X.; Ao, H.; Cao, J.; Lin, W.; Yu, M.; Lin, J.; Cui, Q. MicroRNAs Involved in Carcinogenesis, Prognosis, Therapeutic Resistance and Applications in Human Triple-Negative Breast Cancer. Cells 2019, 8, 1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balkrishna, A.; Mittal, R.; Arya, V. Potential Role of miRNA in Metastatic Cascade of Triple-Negative Breast Cancer. Curr. Cancer Drug Targets 2021, 21, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Riaz, M.; van Jaarsveld, M.T.M.; Hollestelle, A.; Prager-van der Smissen, W.J.C.; Heine, A.A.J.; Boersma, A.W.M.; Liu, J.; Helmijr, J.; Ozturk, B.; Smid, M.; et al. MiRNA expression profiling of 51 human breast cancer cell lines reveals subtype and driver mutation-specific miRNAs. Breast Cancer Res. 2013, 15, R33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blenkiron, C.; Goldstein, L.D.; Thorne, N.P.; Spiteri, I.; Chin, S.F.; Dunning, M.J.; Barbosa-Morais, N.L.; Teschendorff, A.E.; Green, A.R.; Ellis, I.O.; et al. MicroRNA expression profiling of human breast cancer identifies new markers of tumor subtype. Genome Biol. 2007, 8, R214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Søkilde, R.; Persson, H.; Ehinger, A.; Pirona, A.C.; Fernö, M.; Hegardt, C.; Larsson, C.; Loman, N.; Malmberg, M.; Rydén, L.; et al. Refinement of breast cancer molecular classification by miRNA expression profiles. BMC Genom. 2019, 20, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, V.; Davey, M.G.; Annuk, H.; Miller, N.; Dwyer, R.M.; Lowery, A.; Kerin, M.J. Micrornas in molecular classification and pathogenesis of breast tumors. Cancers 2021, 13, 5332. [Google Scholar] [CrossRef] [PubMed]
- Sugita, B.; Gill, M.; Mahajan, A.; Duttargi, A.; Kirolikar, S.; Almeida, R.; Regis, K.; Oluwasanmi, O.L.; Marchi, F.; Marian, C.; et al. Differentially expressed miRNAs in triple negative breast cancer between African-American and non-Hispanic white women. Oncotarget 2016, 7, 79274–79291. [Google Scholar] [CrossRef] [Green Version]
- Sugita, B.M.; Pereira, S.R.; De Almeida, R.C.; Gill, M.; Mahajan, A.; Duttargi, A.; Kirolikar, S.; Fadda, P.; De Lima, R.S.; Urban, C.A.; et al. Integrated copy number and miRNA expression analysis in triple negative breast cancer of Latin American patients. Oncotarget 2019, 10, 6184–6203. [Google Scholar] [CrossRef] [Green Version]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, U.; Lai, X.; Winter, F.; Wolkenhauer, O.; Vera, J.; Gupta, S.K. Cooperative gene regulation by microRNA pairs and their identification using a computational workflow. Nucleic Acids Res. 2014, 42, 7539–7552. [Google Scholar] [CrossRef] [Green Version]
- Elango, R.; Alsaleh, K.A.; Vishnubalaji, R.; Manikandan, M.; Ali, A.M.; Abd El-Aziz, N.; Altheyab, A.; Al-Rikabi, A.; Alfayez, M.; Aldahmash, A.; et al. MicroRNA Expression Profiling on Paired Primary and Lymph Node Metastatic Breast Cancer Revealed Distinct microRNA Profile Associated With LNM. Front. Oncol. 2020, 10, 756. [Google Scholar] [CrossRef] [PubMed]
- Bushel, P.R.; McGovern, R.; Liu, L.; Hofmann, O.; Huda, A.; Lu, J.; Hide, W.; Lin, X. Population differences in tran-script-regulator expression quantitative trait loci. PLoS ONE 2012, 7, e34286. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Graham, K.; Shen, J.; Campbell, L.E.S.; Singh, P.; Zirpoli, G.; Roberts, M.; Ciupak, G.; Davis, W.; Hwang, H.; et al. Genetic variants in microRNAs and breast cancer risk in African American and European American women. Breast Cancer Res. Treat. 2013, 141, 447–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volk-Draper, L.D.; Rajput, S.; Hall, K.L.; Wilber, A.; Ran, S. Novel model for basaloid triple-negative breast cancer: Behavior in vivo and response to therapy. Neoplasia 2012, 14, 926–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrell, J.C.; Pfefferle, A.D.; Zalles, N.; Prat, A.; Fan, C.; Khramtsov, A.; Olopade, O.I.; Troester, M.A.; Dudley, A.C.; Perou, C.M. Endothelial-like properties of claudin-low breast cancer cells promote tumor vascular permeability and metastasis. Clin. Exp. Metastasis 2014, 31, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Luan, P.B.; Jia, X.Z.; Yao, J. MiR-769-5p functions as an oncogene by down-regulating RYBP expression in gastric cancer. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 6699–6706. [Google Scholar] [CrossRef]
- Lee, D. miR-769-5p is associated with prostate cancer recurrence and modulates proliferation and apoptosis of cancer cells. Exp. Ther. Med. 2021, 21, 335. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, B.; Duan, A.; Shen, K.; Zhang, Q.; Tang, X.; Wei, Y.; Tang, J.; Zhang, S. Exosomal transfer of miR-769-5p promotes osteosarcoma proliferation and metastasis by targeting DUSP16. Cancer Cell Int. 2021, 21, 541. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Newbury, P.A.; Glicksberg, B.S.; Zeng, W.Z.D.; Paithankar, S.; Andrechek, E.R.; Chen, B. Evaluating cell lines as models for metastatic breast cancer through integrative analysis of genomic data. Nat. Commun. 2019, 10, 2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, J.; Zhang, Y.; Deng, M.; Ye, R.; Zhao, S.; Wang, Y.; Li, J.; Zhao, Z. MicroRNA control of epithelial-mesenchymal transition in cancer stem cells. Int. J. Cancer 2014, 135, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Li, Y.; Wang, J.; Wen, Z.; Lai, M.; Zhang, H. Molecular mechanisms of microRNAs in regulating epitheli-al-mesenchymal transitions in human cancers. Cancer Lett. 2016, 371, 301–313. [Google Scholar] [CrossRef]
- Lachat, C.; Peixoto, P.; Hervouet, E. Epithelial to mesenchymal transition history: From embryonic development to cancers. Biomolecules 2021, 11, 782. [Google Scholar] [CrossRef] [PubMed]
- Derynck, R.; Turley, S.J.; Akhurst, R.J. TGFβ biology in cancer progression and immunotherapy. Nat. Rev. Clin. Oncol. 2021, 18, 9–34. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Han, S. miR-150-5p promotes the proliferation and epithelial-mesenchymal transition of cervical carcinoma cells via targeting SRCIN1. Pathol.-Res. Pract. 2019, 215, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.N.; Burton, L.J.; Henderson, V.; Randle, D.D.; Morton, D.J.; Smith, B.A.; Taliaferro-Smith, L.; Nagappan, P.; Yates, C.; Zayzafoon, M.; et al. Snail promotes epithelial mesenchymal transition in breast cancer cells in part via activation of nuclear ERK2. PLoS ONE 2014, 9, e104987. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TNBC | Non-TNBC | p-Value | |
|---|---|---|---|
| Age | |||
| ≤53 | 28 | 34 | p > 0.9999 |
| 53 | 19 | 23 | |
| Tumor Size | |||
| ≤2 cm | 33 | 27 | p = 0.6758 |
| >2 cm | 18 | 19 | |
| Histology | |||
| Ductal | 56 | 5 | p = 0.0129 |
| Lobular | 0 | 7 | |
| Grade | |||
| 1/2 | 0 | 29 | p = 0.0135 |
| 2/3 | 46 | 19 | |
| Lymph node | |||
| Positive | 17 | 22 | p = 0.5152 |
| Negative | 24 | 22 | |
| Metastasis | |||
| Positive | 3 | 8 | p = 0.2177 |
| Negative | 41 | 44 | |
| Race | |||
| AA | 34 | 34 | p > 0.9999 |
| CA | 22 | 23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugita, B.M.; Rodriguez, Y.; Fonseca, A.S.; Nunes Souza, E.; Kallakury, B.; Cavalli, I.J.; Ribeiro, E.M.S.F.; Aneja, R.; Cavalli, L.R. MiR-150-5p Overexpression in Triple-Negative Breast Cancer Contributes to the In Vitro Aggressiveness of This Breast Cancer Subtype. Cancers 2022, 14, 2156. https://doi.org/10.3390/cancers14092156
Sugita BM, Rodriguez Y, Fonseca AS, Nunes Souza E, Kallakury B, Cavalli IJ, Ribeiro EMSF, Aneja R, Cavalli LR. MiR-150-5p Overexpression in Triple-Negative Breast Cancer Contributes to the In Vitro Aggressiveness of This Breast Cancer Subtype. Cancers. 2022; 14(9):2156. https://doi.org/10.3390/cancers14092156
Chicago/Turabian StyleSugita, Bruna M., Yara Rodriguez, Aline S. Fonseca, Emanuelle Nunes Souza, Bhaskar Kallakury, Iglenir J. Cavalli, Enilze M. S. F. Ribeiro, Ritu Aneja, and Luciane R. Cavalli. 2022. "MiR-150-5p Overexpression in Triple-Negative Breast Cancer Contributes to the In Vitro Aggressiveness of This Breast Cancer Subtype" Cancers 14, no. 9: 2156. https://doi.org/10.3390/cancers14092156
APA StyleSugita, B. M., Rodriguez, Y., Fonseca, A. S., Nunes Souza, E., Kallakury, B., Cavalli, I. J., Ribeiro, E. M. S. F., Aneja, R., & Cavalli, L. R. (2022). MiR-150-5p Overexpression in Triple-Negative Breast Cancer Contributes to the In Vitro Aggressiveness of This Breast Cancer Subtype. Cancers, 14(9), 2156. https://doi.org/10.3390/cancers14092156