
Tissue miRNA Combinations for the Differential Diagnosis of Adrenocortical Carcinoma and Adenoma Established by Artificial Intelligence

 , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Collection and Ethics Approval
2.2. Literature Search
2.3. Sample Processing and RNA Isolation
2.4. Analysis of the miRNA Panel Expression by Real-Time RT-qPCR
2.5. Statistical Analysis
3. Results
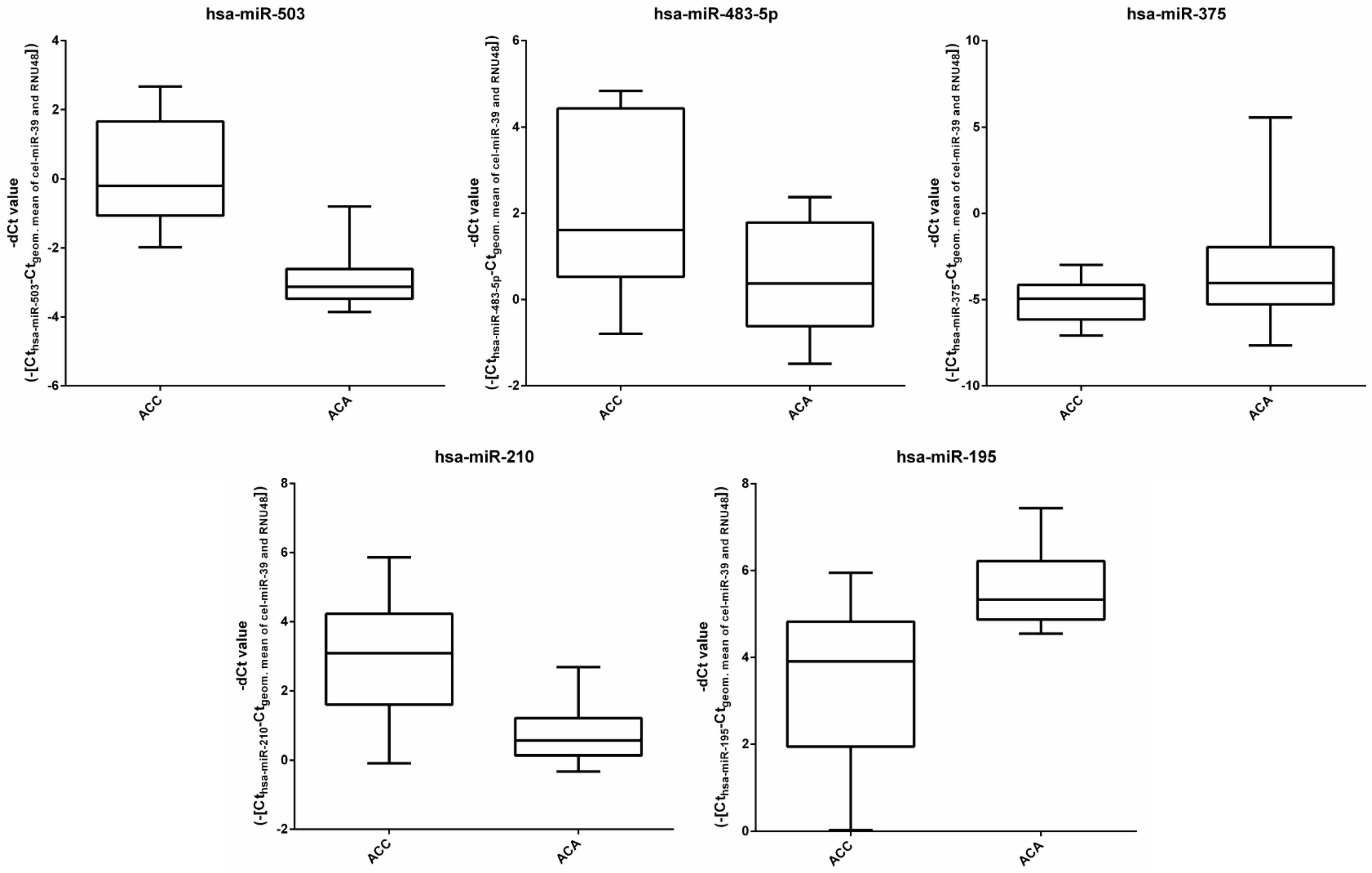
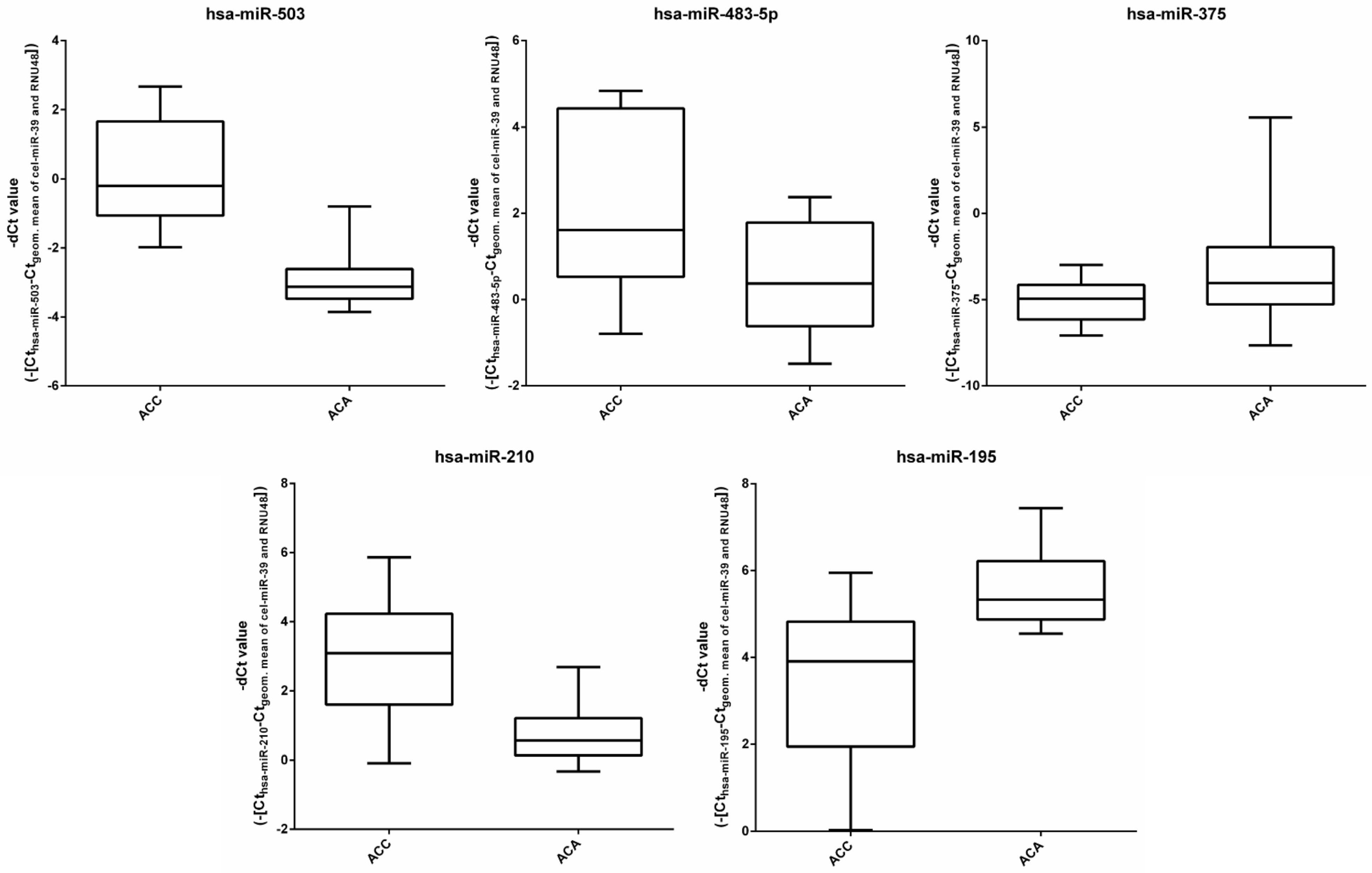
3.1. miRNA Expression in the Discovery Cohort by RT-qPCR
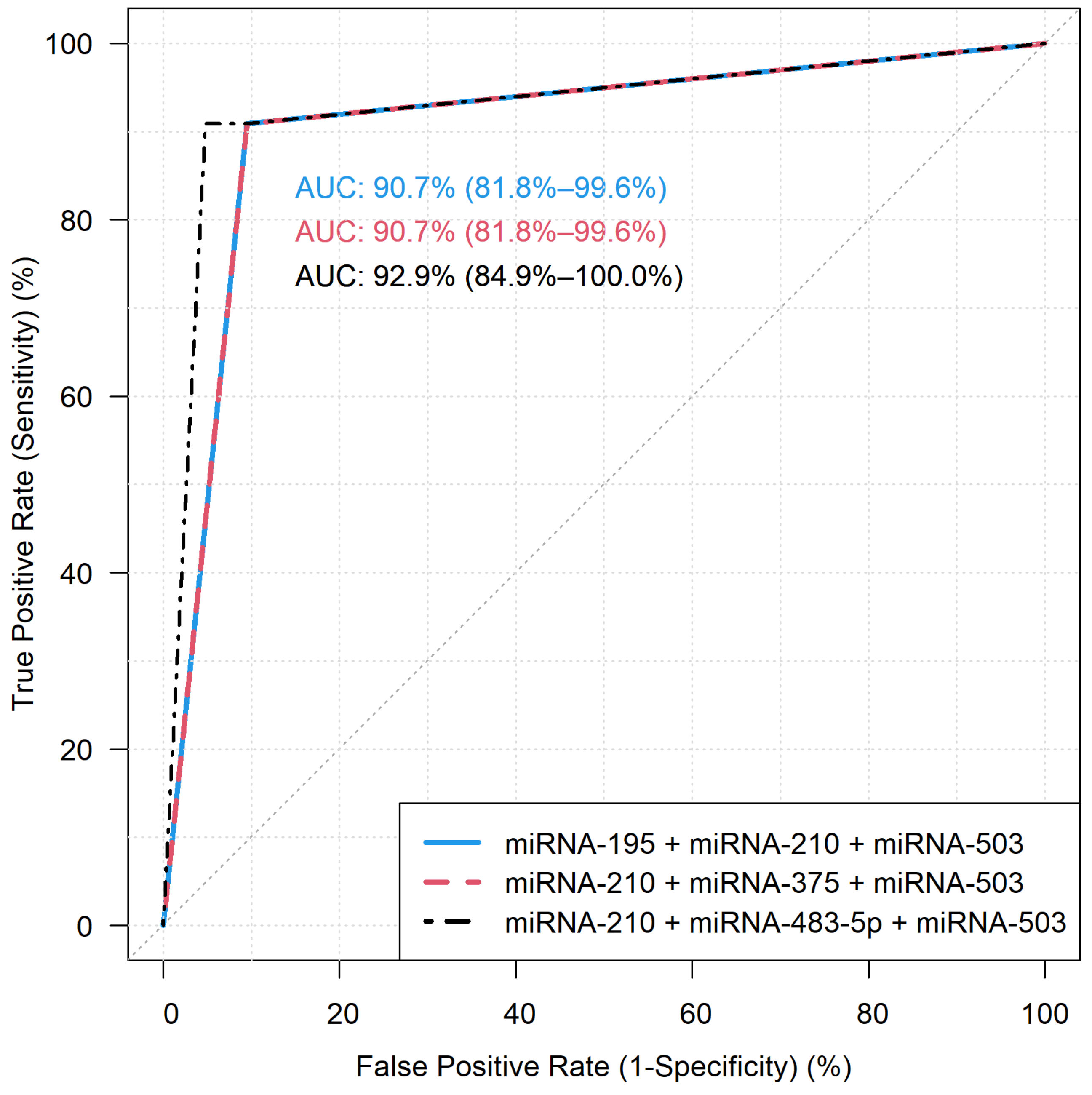
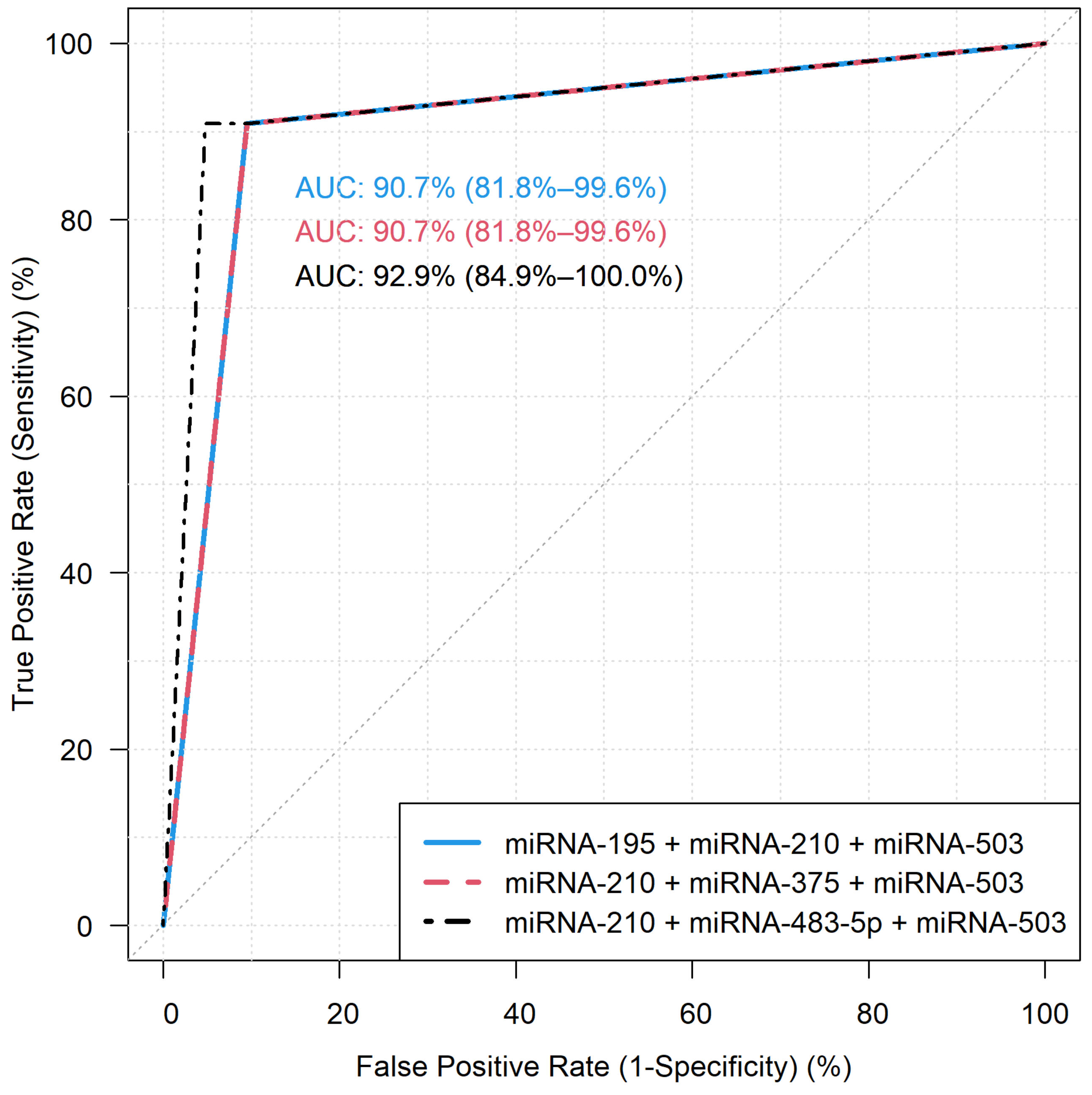
3.2. Diagnostic Performance of the miRNA Models by RT-qPCR
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bovio, S.; Cataldi, A.; Reimondo, G.; Sperone, P.; Novello, S.; Berruti, A.; Borasio, P.; Fava, C.; Dogliotti, L.; Scagliotti, G.V.; et al. Prevalence of adrenal incidentaloma in a contemporary computerized tomography series. J. Endocrinol. Investig. 2006, 29, 298–302. [Google Scholar] [CrossRef]
- Libé, R.; Borget, I.; Ronchi, C.L.; Zaggia, B.; Kroiss, M.; Kerkhofs, T.; Bertherat, J.; Volante, M.; Quinkler, M.; Chabre, O.; et al. Prognostic factors in stage III–IV adrenocortical carcinomas (ACC): An European Network for the Study of Adrenal Tumor (ENSAT) study. Ann. Oncol. 2015, 26, 2119–2125. [Google Scholar] [CrossRef] [PubMed]
- Fassnacht, M.; Dekkers, O.M.; Else, T.; Baudin, E.; Berruti, A.; De Krijger, R.R.; Haak, H.R.; Mihai, R.; Assie, G.; Terzolo, M. European Society of Endocrinology Clinical Practice Guidelines on the management of adrenocortical carcinoma in adults, in collaboration with the European Network for the Study of Adrenal Tumors. Eur. J. Endocrinol. 2018, 179, G1–G46. [Google Scholar] [CrossRef] [PubMed]
- Terzolo, M.; Daffara, F.; Ardito, A.; Zaggia, B.; Basile, V.; Ferrari, L.; Berruti, A. Management of adrenal cancer: A 2013 update. J. Endocrinol. Investig. 2014, 37, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Fassnacht, M.; Arlt, W.; Bancos, I.; Dralle, H.; Newell-Price, J.; Sahdev, A.; Tabarin, A.; Terzolo, M.; Tsagarakis, S.; Dekkers, O.M. Management of adrenal incidentalomas: European Society of Endocrinology Clinical Practice Guideline in collaboration with the European Network for the Study of Adrenal Tumors. Eur. J. Endocrinol. 2016, 175, G1–G34. [Google Scholar] [CrossRef] [Green Version]
- Lattin, G.E.; Sturgill, E.D.; Tujo, C.A.; Marko, J.; Sanchez-Maldonado, K.W.; Craig, W.D.; Lack, E.E. From the radiologic pathology archives: Adrenal tumors and tumor-like conditions in the adult: Radiologic-pathologic correlation. Radiographics 2014, 34, 805–829. [Google Scholar] [CrossRef] [Green Version]
- Bancos, I.; Arlt, W. Diagnosis of a malignant adrenal mass: The role of urinary steroid metabolite profiling. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 200–207. [Google Scholar] [CrossRef]
- Viëtor, C.L.; Creemers, S.G.; van Kemenade, F.J.; van Ginhoven, T.M.; Hofland, L.J.; Feelders, R.A. How to Differentiate Benign from Malignant Adrenocortical Tumors? Cancers 2021, 13, 4383. [Google Scholar] [CrossRef]
- Mazzaglia, P.J.; Monchik, J.M. Limited Value of Adrenal Biopsy in the Evaluation of Adrenal Neoplasm: A Decade of Experience. Arch. Surg. 2009, 144, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.R.; Hammer, G.D.; Else, T. Transcutaneous Biopsy of Adrenocortical Carcinoma is rarely helpful in diagnosis, potentially harmful, but does not affect patient outcome. Eur. J. Endocrinol. 2014, 170, 829. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.D.; Delivanis, D.A.; Eiken, P.W.; Atwell, T.D.; Bancos, I. Adrenal biopsy: Performance and use. Minerva Endocrinol. 2019, 44, 288–300. [Google Scholar] [CrossRef]
- Bancos, I.; Tamhane, S.; Shah, M.; Delivanis, D.A.; Alahdab, F.; Arlt, W.; Fassnacht, M.; Murad, M.H. DIAGNOSIS OF ENDOCRINE DISEASE: The diagnostic performance of adrenal biopsy: A systematic review and meta-analysis. Eur. J. Endocrinol. 2016, 175, R65–R80. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [Green Version]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2018, 20, 21–37. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Ritland Politz, J.C.; Hogan, E.M.; Pederson, T. MicroRNAs with a nucleolar location. RNA 2009, 15, 1705. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef]
- Igaz, I.; Igaz, P. Tumor surveillance by circulating microRNAs: A hypothesis. Cell. Mol. Life Sci. 2014, 71, 4081–4087. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; She, X.; Ming, Y.Z.; Wan, Q. quan miR-24 promotes the proliferation and invasion of HCC cells by targeting SOX7. Tumor Biol. 2014, 35, 10731–10736. [Google Scholar] [CrossRef]
- Yin, Y.; Zhong, J.; Li, S.W.; Li, J.Z.; Zhou, M.; Chen, Y.; Sang, Y.; Liu, L. TRIM11, a direct target of miR-24-3p, promotes cell proliferation and inhibits apoptosis in colon cancer. Oncotarget 2016, 7, 86755. [Google Scholar] [CrossRef] [Green Version]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Hall, J.S.; Taylor, J.; Valentine, H.R.; Irlam, J.J.; Eustace, A.; Hoskin, P.J.; Miller, C.J.; West, C.M.L. Enhanced stability of microRNA expression facilitates classification of FFPE tumour samples exhibiting near total mRNA degradation. Br. J. Cancer 2012, 107, 684–694. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155. [Google Scholar] [CrossRef]
- Igaz, P.; Igaz, I.; Nagy, Z.; Nyíro, G.; Szabó, P.M.; Falus, A.; Patócs, A.; Rácz, K. MicroRNAs in adrenal tumors: Relevance for pathogenesis, diagnosis, and therapy. Cell. Mol. Life Sci. 2014, 72, 417–428. [Google Scholar] [CrossRef]
- Decmann, A.; Perge, P.; Turai, P.I.; Patócs, A.; Igaz, P. Non-Coding RNAs in Adrenocortical Cancer: From Pathogenesis to Diagnosis. Cancers 2020, 12, 461. [Google Scholar] [CrossRef] [Green Version]
- Chehade, M.; Bullock, M.; Glover, A.; Hutvagner, G.; Sidhu, S. Key MicroRNA’s and Their Targetome in Adrenocortical Cancer. Cancers 2020, 12, 2198. [Google Scholar] [CrossRef]
- Singh, P.; Soon, P.S.H.; Feige, J.J.; Chabre, O.; Zhao, J.T.; Cherradi, N.; Lalli, E.; Sidhu, S.B. Dysregulation of microRNAs in adrenocortical tumors. Mol. Cell. Endocrinol. 2012, 351, 118–128. [Google Scholar] [CrossRef]
- Soon, P.S.H.; Tacon, L.J.; Gill, A.J.; Bambach, C.P.; Sywak, M.S.; Campbell, P.R.; Yeh, M.W.; Wong, S.G.; Clifton-Bligh, R.J.; Robinson, B.G.; et al. miR-195 and miR-483-5p Identified as Predictors of Poor Prognosis in Adrenocortical Cancer. Clin. Cancer Res. 2009, 15, 7684–7692. [Google Scholar] [CrossRef] [Green Version]
- Glover, A.R.; Zhao, J.T.; Gill, A.J.; Weiss, J.; Mugridge, N.; Kim, E.; Feeney, A.L.; Ip, J.C.; Reid, G.; Clarke, S.; et al. microRNA-7 as a tumor suppressor and novel therapeutic for adrenocortical carcinoma. Oncotarget 2015, 6, 36675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, A.M.; Sbiera, S.; Ribeiro, T.C.; Soares, I.C.; Mariani, B.M.P.; Freire, D.S.; De Sousa, G.R.V.; Lerario, A.M.; Ronchi, C.L.; Deutschbein, T.; et al. Expression of LIN28 and its regulatory microRNAs in adult adrenocortical cancer. Clin. Endocrinol. 2015, 82, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Khafaei, M.; Rezaie, E.; Mohammadi, A.; Shahnazi Gerdehsang, P.; Ghavidel, S.; Kadkhoda, S.; Zorrieh Zahra, A.; Forouzanfar, N.; Arabameri, H.; Tavallaie, M. miR-9: From function to therapeutic potential in cancer. J. Cell. Physiol. 2019, 234, 14651–14665. [Google Scholar] [CrossRef] [PubMed]
- Özata, D.M.; Caramuta, S.; Velázquez-Fernández, D.; Akçakaya, P.; Xie, H.; Höög, A.; Zedenius, J.; Bäckdahl, M.; Larsson, C.; Lui, W.O. The role of microRNA deregulation in the pathogenesis of adrenocortical carcinoma. Endocr. Relat. Cancer 2011, 18, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Romero, D.G.; Plonczynski, M.W.; Carvajal, C.A.; Gomez-Sanchez, E.P.; Gomez-Sanchez, C.E. Microribonucleic Acid-21 Increases Aldosterone Secretion and Proliferation in H295R Human Adrenocortical Cells. Endocrinology 2008, 149, 2477. [Google Scholar] [CrossRef]
- Chabre, O.; Libé, R.; Assie, G.; Barreau, O.; Bertherat, J.; Bertagna, X.; Feige, J.J.; Cherradi, N. Serum miR-483-5p and miR-195 are predictive of recurrence risk in adrenocortical cancer patients. Endocr. Relat. Cancer 2013, 20, 579–594. [Google Scholar] [CrossRef]
- Patterson, E.E.; Holloway, A.K.; Weng, J.; Fojo, T.; Kebebew, E. MicroRNA profiling of adrenocortical tumors reveals miR-483 as a marker of malignancy. Cancer 2011, 117, 1630. [Google Scholar] [CrossRef] [Green Version]
- Feinmesser, M.; Benbassat, C.; Meiri, E.; Benjamin, H.; Lebanony, D.; Lebenthal, Y.; De Vries, L.; Drozd, T.; Spector, Y. Specific microRNAs differentiate adrenocortical adenomas from carcinomas and correlate with weiss histopathologic system. Appl. Immunohistochem. Mol. Morphol. 2015, 23, 522–531. [Google Scholar] [CrossRef]
- Szabó, D.R.; Luconi, M.; Szabó, P.M.; Tóth, M.; Szücs, N.; Horányi, J.; Nagy, Z.; Mannelli, M.; Patócs, A.; Rácz, K.; et al. Analysis of circulating microRNAs in adrenocortical tumors. Lab. Investig. 2013, 94, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wang, W.; Hu, W.; Xu, W.; Xiao, G.; Nie, Q.; Ouyang, K.; Chen, S. MicroRNA-205 suppresses the growth of adrenocortical carcinoma SW-13 cells via targeting Bcl-2. Oncol. Rep. 2015, 34, 3104–3110. [Google Scholar] [CrossRef]
- Pereira, S.S.; Monteiro, M.P.; Antonini, S.R.; Pignatelli, D. Apoptosis regulation in adrenocortical carcinoma. Endocr. Connect. 2019, 8, R91. [Google Scholar] [CrossRef] [Green Version]
- Tömböl, Z.; Szabó, P.M.; Molnár, V.; Wiener, Z.; Tölgyesi, G.; Horányi, J.; Riesz, P.; Reismann, P.; Patócs, A.; Likó, I.; et al. Integrative molecular bioinformatics study of human adrenocortical tumors: MicroRNA, tissue-specific target prediction, and pathway analysis. Endocr. Relat. Cancer 2009, 16, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Koperski, L.; Kotlarek, M.; Swierniak, M.; Kolanowska, M.; Kubiak, A.; Górnicka, B.; Jazdzewski, K.; Wójcicka, A. Next-generation sequencing reveals microRNA markers of adrenocortical tumors malignancy. Oncotarget 2017, 8, 49191. [Google Scholar] [CrossRef]
- Assié, G.; Letouzé, E.; Fassnacht, M.; Jouinot, A.; Luscap, W.; Barreau, O.; Omeiri, H.; Rodriguez, S.; Perlemoine, K.; René-Corail, F.; et al. Integrated genomic characterization of adrenocortical carcinoma. Nat. Genet. 2014, 46, 607–612. [Google Scholar] [CrossRef]
- Schmitz, K.J.; Helwig, J.; Bertram, S.; Sheu, S.Y.; Suttorp, A.C.; Seggewiß, J.; Willscher, E.; Walz, M.K.; Worm, K.; Schmid, K.W. Differential expression of microRNA-675, microRNA-139-3p and microRNA-335 in benign and malignant adrenocortical tumours. J. Clin. Pathol. 2011, 64, 529–535. [Google Scholar] [CrossRef]
- Kwok, G.T.Y.; Zhao, J.T.; Glover, A.R.; Gill, A.J.; Clifton-Bligh, R.; Robinson, B.G.; Ip, J.C.Y.; Sidhu, S.B. microRNA-431 as a Chemosensitizer and Potentiator of Drug Activity in Adrenocortical Carcinoma. Oncologist 2019, 24, e241. [Google Scholar] [CrossRef] [Green Version]
- Duregon, E.; Rapa, I.; Votta, A.; Giorcelli, J.; Daffara, F.; Terzolo, M.; Scagliotti, G.V.; Volante, M.; Papotti, M. MicroRNA expression patterns in adrenocortical carcinoma variants and clinical pathologic correlations. Hum. Pathol. 2014, 45, 1555–1562. [Google Scholar] [CrossRef] [Green Version]
- Veronese, A.; Lupini, L.; Consiglio, J.; Visone, R.; Ferracin, M.; Fornari, F.; Zanesi, N.; Alder, H.; D’Elia, G.; Gramantieri, L.; et al. Oncogenic Role of miR-483-3p at the IGF2/483 Locus. Cancer Res. 2010, 70, 3140. [Google Scholar] [CrossRef] [Green Version]
- Perge, P.; Butz, H.; Pezzani, R.; Bancos, I.; Nagy, Z.; Pálóczi, K.; Nyíro, G.; Decmann, Á.; Pap, E.; Luconi, M.; et al. Evaluation and diagnostic potential of circulating extracellular vesicle-associated microRNAs in adrenocortical tumors. Sci. Rep. 2017, 7, 5474. [Google Scholar] [CrossRef]
- Salvianti, F.; Canu, L.; Poli, G.; Armignacco, R.; Scatena, C.; Cantini, G.; Di Franco, A.; Gelmini, S.; Ercolino, T.; Pazzagli, M.; et al. New insights in the clinical and translational relevance of miR483-5p in adrenocortical cancer. Oncotarget 2017, 8, 65525. [Google Scholar] [CrossRef] [Green Version]
- Decmann, A.; Bancos, I.; Khanna, A.; Thomas, M.A.; Turai, P.; Perge, P.; Pintér, J.Z.; Tóth, M.; Patócs, A.; Igaz, P. Comparison of plasma and urinary microRNA-483-5p for the diagnosis of adrenocortical malignancy. J. Biotechnol. 2019, 297, 49–53. [Google Scholar] [CrossRef]
- Zheng, S.; Cherniack, A.D.; Dewal, N.; Moffitt, R.A.; Danilova, L.; Murray, B.A.; Lerario, A.M.; Else, T.; Knijnenburg, T.A.; Ciriello, G.; et al. Comprehensive Pan-Genomic Characterization of Adrenocortical Carcinoma. Cancer Cell 2016, 29, 723. [Google Scholar] [CrossRef] [Green Version]
- Goh, G.; Scholl, U.I.; Healy, J.M.; Choi, M.; Prasad, M.L.; Nelson-Williams, C.; Kuntsman, J.W.; Korah, R.; Suttorp, A.C.; Dietrich, D.; et al. Recurrent activating mutation in PRKACA in cortisol-producing adrenal tumors. Nat. Genet. 2014, 46, 613. [Google Scholar] [CrossRef] [Green Version]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Modern Applied Statistics with S. Available online: https://link.springer.com/book/10.1007/978-0-387-21706-2 (accessed on 7 December 2021).
- Caret: Classification and Regression Training R Package Version 6.0-90. Available online: https://CRAN.R-project.org/package=caret (accessed on 7 December 2021).
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef]
- Soon, P.S.H.; Libe, R.; Benn, D.E.; Gill, A.; Shaw, J.; Sywak, M.S.; Groussin, L.; Bertagna, X.; Gicquel, C.; Bertherat, J.; et al. Loss of heterozygosity of 17p13, with possible involvement of ACADVL and ALOX15B, in the pathogenesis of adrenocortical tumors. Ann. Surg. 2008, 247, 157–164. [Google Scholar] [CrossRef]
- Bavelloni, A.; Ramazzotti, G.; Poli, A.; Piazzi, M.; Focaccia, E.; Blalock, W.; Faenza, I. MiRNA-210: A Current Overview. Anticancer Res. 2017, 37, 6511–6521. [Google Scholar]
- Pouysségur, J.; Dayan, F.; Mazure, N.M. Hypoxia signalling in cancer and approaches to enforce tumour regression. Nature 2006, 441, 437–443. [Google Scholar] [CrossRef]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar] [CrossRef]
- Grosso, S.; Doyen, J.; Parks, S.K.; Bertero, T.; Paye, A.; Cardinaud, B.; Gounon, P.; Lacas-Gervais, S.; Noël, A.; Pouysségur, J.; et al. MiR-210 promotes a hypoxic phenotype and increases radioresistance in human lung cancer cell lines. Cell Death Dis. 2013, 4, e544. [Google Scholar] [CrossRef]
- Nohata, N.; Hanazawa, T.; Kikkawa, N.; Mutallip, M.; Sakurai, D.; Fujimura, L.; Kawakami, K.; Chiyomaru, T.; Yoshino, H.; Enokida, H.; et al. Tumor suppressive microRNA-375 regulates oncogene AEG-1/MTDH in head and neck squamous cell carcinoma (HNSCC). J. Hum. Genet. 2011, 56, 595–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukamoto, Y.; Nakada, C.; Noguchi, T.; Tanigawa, M.; Nguyen, L.T.; Uchida, T.; Hijiya, N.; Matsuura, K.; Fujioka, T.; Seto, M.; et al. MicroRNA-375 Is Downregulated in Gastric Carcinomas and Regulates Cell Survival by Targeting PDK1 and 14-3-3ζ. Cancer Res. 2010, 70, 2339–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.M.; Poon, R.T.P.; Luk, J.M. MicroRNA-375 targets Hippo-signaling effector YAP in liver cancer and inhibits tumor properties. Biochem. Biophys. Res. Commun. 2010, 394, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Xu, Y.; Zhang, W.; Deng, Y.; Si, M.; Du, Y.; Yao, H.; Liu, X.; Ke, Y.; Si, J.; et al. MiR-375 frequently downregulated in gastric cancer inhibits cell proliferation by targeting JAK2. Cell Res. 2010, 20, 784–793. [Google Scholar] [CrossRef]
- Li, X.; Lin, R.; Li, J. Epigenetic silencing of microRNA-375 regulates PDK1 expression in esophageal cancer. Dig. Dis. Sci. 2011, 56, 2849–2856. [Google Scholar] [CrossRef]
- Ladeiro, Y.; Couchy, G.; Balabaud, C.; Bioulac-Sage, P.; Pelletier, L.; Rebouissou, S.; Zucman-Rossi, J. MicroRNA profiling in hepatocellular tumors is associated with clinical features and oncogene/tumor suppressor gene mutations. Hepatology 2008, 47, 1955–1963. [Google Scholar] [CrossRef]
- Tissier, F.; Cavard, C.; Groussin, L.; Perlemoine, K.; Fumey, G.; Hagneré, A.M.; René-Corail, F.; Jullian, E.; Gicquel, C.; Bertagna, X.; et al. Mutations of β-Catenin in Adrenocortical Tumors: Activation of the Wnt Signaling Pathway Is a Frequent Event in both Benign and Malignant Adrenocortical Tumors. Cancer Res. 2005, 65, 7622–7627. [Google Scholar] [CrossRef] [Green Version]
- Berthon, A.; Martinez, A.; Bertherat, J.; Val, P. Wnt/β-catenin signalling in adrenal physiology and tumour development. Mol. Cell. Endocrinol. 2012, 351, 87–95. [Google Scholar] [CrossRef]
- Decmann, A.; Perge, P.; Nyíro, G.; Darvasi, O.; Likó, I.; Borka, K.; Micsik, T.; Tóth, Z.; Bancos, I.; Pezzani, R.; et al. MicroRNA Expression Profiling in Adrenal Myelolipoma. J. Clin. Endocrinol. Metab. 2018, 103, 3522–3530. [Google Scholar] [CrossRef] [Green Version]
- Soon, P.S.H.; Gill, A.J.; Benn, D.E.; Clarkson, A.; Robinson, B.G.; McDonald, K.L.; Sidhu, S.B. Microarray gene expression and immunohistochemistry analyses of adrenocortical tumors identify IGF2 and Ki-67 as useful in differentiating carcinomas from adenomas. Endocr. Relat. Cancer 2009, 16, 573–583. [Google Scholar] [CrossRef] [Green Version]
- De Fraipont, F.; El Atifi, M.; Cherradi, N.; Le Moigne, G.; Defaye, G.; Houlgatte, R.; Bertherat, J.; Bertagna, X.; Plouin, P.F.; Baudin, E.; et al. Gene Expression Profiling of Human Adrenocortical Tumors Using Complementary Deoxyribonucleic Acid Microarrays Identifies Several Candidate Genes as Markers of Malignancy. J. Clin. Endocrinol. Metab. 2005, 90, 1819–1829. [Google Scholar] [CrossRef] [Green Version]
- Agosta, C.; Laugier, J.; Guyon, L.; Denis, J.; Bertherat, J.; Libé, R.; Boisson, B.; Sturm, N.; Feige, J.J.; Chabre, O.; et al. MiR-483-5p and miR-139-5p promote aggressiveness by targeting N-myc downstream-regulated gene family members in adrenocortical cancer. Int. J. Cancer 2018, 143, 944–957. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shi, M.; Hou, S.; Ding, B.; Liu, L.; Ji, X.; Zhang, J.; Deng, Y. MiR-483–5p suppresses the proliferation of glioma cells via directly targeting ERK1. FEBS Lett. 2012, 586, 1312–1317. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; He, B.; Xu, J.; Li, Y.; Guo, C.; Cai, Q.; Wang, S. miR-483-5p Targets MKNK1 to Suppress Wilms’ Tumor Cell Proliferation and Apoptosis In Vitro and In Vivo. Med. Sci. Monit. 2019, 25, 1459. [Google Scholar] [CrossRef]
- Zhao, J.J.; Yang, J.; Lin, J.; Yao, N.; Zhu, Y.; Zheng, J.; Xu, J.; Cheng, J.Q.; Lin, J.Y.; Ma, X. Identification of miRNAs associated with tumorigenesis of retinoblastoma by miRNA microarray analysis. Child’s Nerv. Syst. 2009, 25, 13–20. [Google Scholar] [CrossRef]
- Corbetta, S.; Vaira, V.; Guarnieri, V.; Scillitani, A.; Eller-Vainicher, C.; Ferrero, S.; Vicentini, L.; Chiodini, I.; Bisceglia, M.; Beck-Peccoz, P.; et al. Differential expression of microRNAs in human parathyroid carcinomas compared with normal parathyroid tissue. Endocr. Relat. Cancer 2010, 17, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Dey, B.K.; Dutta, A. MiR-322/424 and -503 Are Induced during Muscle Differentiation and Promote Cell Cycle Quiescence and Differentiation by Down-Regulation of Cdc25A. Mol. Biol. Cell 2010, 21, 2138. [Google Scholar] [CrossRef]
- Forrest, A.R.R.; Kanamori-Katayama, M.; Tomaru, Y.; Lassmann, T.; Ninomiya, N.; Takahashi, Y.; De Hoon, M.J.L.; Kubosaki, A.; Kaiho, A.; Suzuki, M.; et al. Induction of microRNAs, mir-155, mir-222, mir-424 and mir-503, promotes monocytic differentiation through combinatorial regulation. Leukemia 2009, 24, 460–466. [Google Scholar] [CrossRef] [Green Version]
- Vabalas, A.; Gowen, E.; Poliakoff, E.; Casson, A.J. Machine learning algorithm validation with a limited sample size. PLoS ONE 2019, 14, e0224365. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Cohort/Samples | Sex | Mean Age at Sample Taking (Years) | Mean Tumor Size (mm) | Ki-67 (%) | ENSAT Stage | Hormonal Activity |
|---|---|---|---|---|---|---|
| Discovery ACA | 10 F | 47.5 | 33.9 | - | - | 7 cortisol 3 NF |
| Discovery ACC | 6 F 4 M | 45.2 | 96.2 | 10–15 (1–40) | 5 II 5 III | 3 cortisol 5 NF 1 DOC 1 DOC + cortisol + estradiol |
| Validation ACA | 17 F 5 M | 53.9 | 35 | - | - | 11 cortisol 10 NF 1 DHEAS |
| Validation ACC | 14 F 7 M | 55.4 | 102 | 25–30 (8–50) | 1 I 4 II 5 III 11 IV | 7 cortisol 11 NF 2 cortisol + DHEAS 1 cortisol + androgen |
| miRNAs | Expression in ACC | References |
|---|---|---|
| hsa-miR-7 | Down-regulated | [30,31] |
| hsa-miR-9 | Up-regulated | [32,33] |
| hsa-miR-21 | Up-regulated | [34,35] |
| hsa-miR-195 | Down-regulated | [30,34,36,37,38,39] |
| hsa-miR-205 | Down-regulated | [40,41] |
| hsa-miR-210 | Up-regulated | [34,39,42,43] |
| hsa-miR-214 | Down-regulated | [38,42,44] |
| hsa-miR-335 | Down-regulated | [36,38,45] |
| hsa-miR-375 | Down-regulated | [42] |
| hsa-miR-431 | Down-regulated | [44,46] |
| hsa-miR-483-3p | Up-regulated | [34,38,47,48] |
| hsa-miR-483-5p | Up-regulated | [30,34,36,38,39,49,50,51] |
| hsa-miR-497 | Down-regulated | [34,38] |
| hsa-miR-503 | Up-regulated | [38,42] |
| hsa-miR-508 | Up-regulated | [36,44,52] |
| hsa-miR-511 | Down-regulated | [42,44,53] |
| Model Number | miRNA Combination |
|---|---|
| 1 | hsa-miR-9 + hsa-miR-375 |
| 2 | hsa-miR-9 + hsa-miR-503 |
| 3 | hsa-miR-375 + hsa-miR-503 |
| 4 | hsa-miR-210 + hsa-miR-503 |
| 5 | hsa-miR-375 + hsa-miR-497 |
| 6 | hsa-miR-483-3p + hsa-miR-503 |
| 7 | hsa-miR-503 + hsa-miR-508 |
| 8 | hsa-miR-195 + hsa-miR-503 + hsa-miR-508 |
| 9 | hsa-miR-195 + hsa-miR-210 + hsa-miR-503 |
| 10 | hsa-miR-9 + hsa-miR-195 + hsa-miR-503 |
| 11 | hsa-miR-9 + hsa-miR-210 + hsa-miR-503 |
| 12 | hsa-miR-9 + hsa-miR-375 + hsa-miR-503 |
| 13 | hsa-miR-9 + hsa-miR-483-3p + hsa-miR-503 |
| 14 | hsa-miR-9 + hsa-miR-497 + hsa-miR-503 |
| 15 | hsa-miR-195 + hsa-miR-375 + hsa-miR-497 |
| 16 | hsa-miR-210 + hsa-miR-375 + hsa-miR-503 |
| 17 | hsa-miR-210 + hsa-miR-483-5p + hsa-miR-503 |
| 18 | hsa-miR-375 + hsa-miR-503 + hsa-miR-508 |
| 19 | hsa-miR-375 + hsa-miR-483-3p + hsa-miR-503 |
| 20 | hsa-miR-9 + hsa-miR-195 + hsa-miR-375 + hsa-miR-503 |
| 21 | hsa-miR-9 + hsa-miR-210 + hsa-miR-483-5p + hsa-miR-503 |
| 22 | hsa-miR-210 + hsa-miR-375 + hsa-miR-503 + hsa-miR-508 |
| 23 | hsa-miR-375 + hsa-miR-483-5p + hsa-miR-503 + hsa-miR-508 |
| 24 | hsa-miR-375 + hsa-miR-497 + hsa-miR-503 + hsa-miR-508 |
| Model Number | Sensitivity | Specificity | Area under Curve (AUC) | Negative Predictive Value | Positive Predictive Value |
|---|---|---|---|---|---|
| 1 | 72.73% | 42.86% | 56.49% | 57.14% | 60.00% |
| 2 | 72.73% | 85.71% | 81.17% | 84.21% | 75.00% |
| 3 | 90.91% | 85.71% | 90.04% | 86.96% | 90.00% |
| 4 | 86.36% | 90.48% | 88.42% | 90.48% | 86.36% |
| 5 | 86.36% | 66.67% | 76.52% | 73.08% | 82.35% |
| 6 | 72.73% | 95.24% | 86.15% | 94.12% | 76.92% |
| 7 | 81.82% | 90.48% | 85.93% | 90.00% | 82.61% |
| 8 | 86.36% | 85.71% | 87.34% | 86.36% | 85.71% |
| 9 | 90.91% | 90.48% | 90.69% | 90.91% | 90.48% |
| 10 | 68.18% | 85.71% | 78.90% | 83.33% | 72.00% |
| 11 | 86.36% | 85.71% | 88.10% | 86.36% | 85.71% |
| 12 | 86.36% | 80.95% | 83.66% | 82.61% | 85.00% |
| 13 | 68.18% | 90.48% | 82.47% | 88.24% | 73.08% |
| 14 | 77.27% | 85.71% | 80.84% | 85.00% | 78.26% |
| 15 | 86.36% | 66.67% | 76.52% | 73.08% | 82.35% |
| 16 | 90.91% | 90.48% | 90.69% | 90.91% | 90.48% |
| 17 | 90.91% | 95.24% | 92.86% | 95.24% | 90.91% |
| 18 | 90.91% | 85.71% | 90.04% | 86.96% | 90.00% |
| 19 | 77.27% | 90.48% | 85.61% | 89.47% | 79.17% |
| 20 | 86.36% | 80.95% | 85.50% | 82.61% | 85.00% |
| 21 | 86.36% | 80.95% | 85.71% | 82.61% | 85.00% |
| 22 | 90.91% | 85.71% | 90.04% | 86.96% | 90.00% |
| 23 | 90.91% | 85.71% | 88.31% | 86.96% | 90.00% |
| 24 | 90.91% | 85.71% | 89.39% | 86.96% | 90.00% |
| miRNA | Sensitivity | Specificity | Area under Curve (AUC) | Negative Predictive Value | Positive Predictive Value |
|---|---|---|---|---|---|
| hsa-miR-9 | 54.55% | 61.90% | 59.52% | 60.00% | 56.52% |
| hsa-miR-195 | 86.36% | 71.43% | 78.90% | 76.00% | 83.33% |
| hsa-miR-210 | 68.18% | 80.95% | 76.41% | 78.95% | 70.83% |
| hsa-miR-375 | 81.82% | 23.81% | 53.68% | 52.94% | 55.56% |
| hsa-miR-483-3p | 54.55% | 90.48% | 74.57% | 85.71% | 65.52% |
| hsa-miR-483-5p | 81.82% | 90.48% | 86.15% | 90.00% | 82.61% |
| hsa-miR-497 | 86.36% | 80.95% | 83.66% | 82.61% | 85.00% |
| hsa-miR-503 | 81.82% | 90.48% | 86.15% | 90.00% | 82.61% |
| hsa-miR-508 | 59.09% | 52.38% | 58.33% | 56.52% | 55.00% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turai, P.I.; Herold, Z.; Nyirő, G.; Borka, K.; Micsik, T.; Tőke, J.; Szücs, N.; Tóth, M.; Patócs, A.; Igaz, P. Tissue miRNA Combinations for the Differential Diagnosis of Adrenocortical Carcinoma and Adenoma Established by Artificial Intelligence. Cancers 2022, 14, 895. https://doi.org/10.3390/cancers14040895
Turai PI, Herold Z, Nyirő G, Borka K, Micsik T, Tőke J, Szücs N, Tóth M, Patócs A, Igaz P. Tissue miRNA Combinations for the Differential Diagnosis of Adrenocortical Carcinoma and Adenoma Established by Artificial Intelligence. Cancers. 2022; 14(4):895. https://doi.org/10.3390/cancers14040895
Chicago/Turabian StyleTurai, Péter István, Zoltán Herold, Gábor Nyirő, Katalin Borka, Tamás Micsik, Judit Tőke, Nikolette Szücs, Miklós Tóth, Attila Patócs, and Peter Igaz. 2022. "Tissue miRNA Combinations for the Differential Diagnosis of Adrenocortical Carcinoma and Adenoma Established by Artificial Intelligence" Cancers 14, no. 4: 895. https://doi.org/10.3390/cancers14040895
APA StyleTurai, P. I., Herold, Z., Nyirő, G., Borka, K., Micsik, T., Tőke, J., Szücs, N., Tóth, M., Patócs, A., & Igaz, P. (2022). Tissue miRNA Combinations for the Differential Diagnosis of Adrenocortical Carcinoma and Adenoma Established by Artificial Intelligence. Cancers, 14(4), 895. https://doi.org/10.3390/cancers14040895