
A Novel High-Throughput Screening Method for a Human Multicentric Osteosarcoma-Specific Antibody and Biomarker Using a Phage Display-Derived Monoclonal Antibody


 , , , ,
, , , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Antibody Library and Screening
2.3. Analysis of Fluorescence Probe-Based ELISA
2.4. Flow Cytometry Analysis
2.5. Immunohistochemistry
2.6. Immunoprecipitation with Magnetic Beads and Electrophoresis
2.7. Protein Digestion and LC/MS Analysis
2.8. Confirmation with Purified Protein and Commercial Antibody
2.9. Statistical Analysis
3. Results
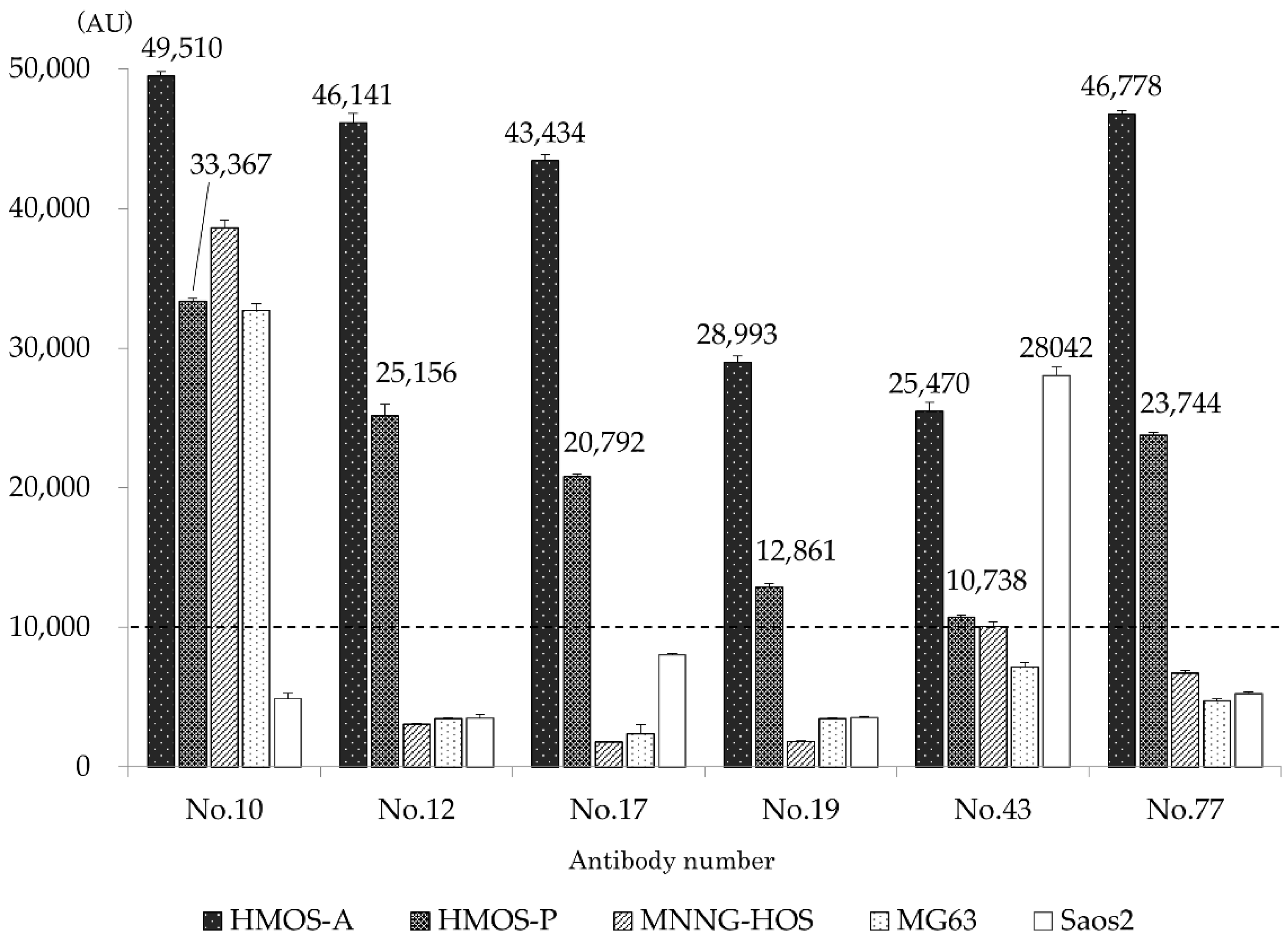
3.1. Antibody Selection at Fluorescence Probe-Based ELISA
3.2. Flow Cytometry
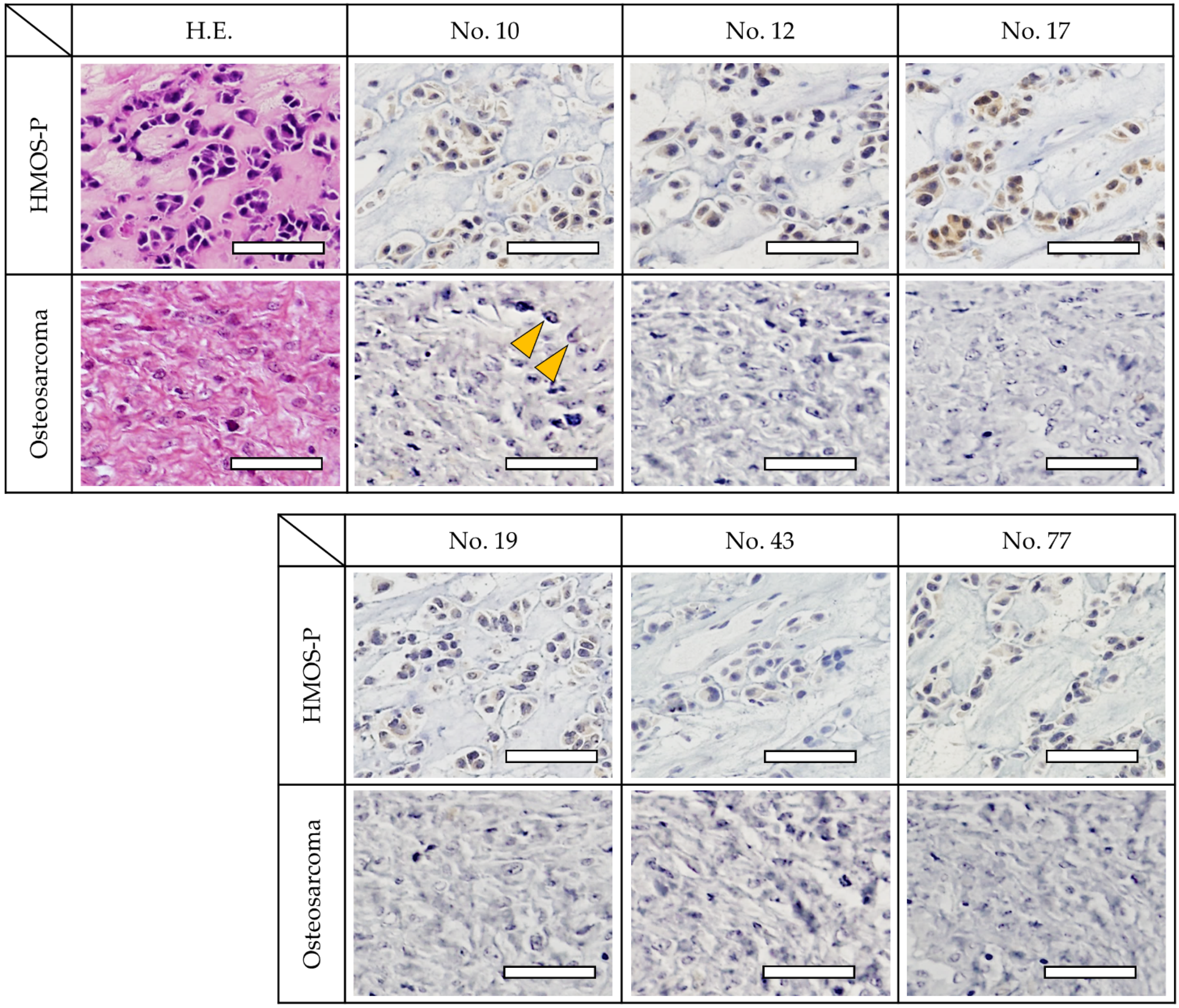
3.3. Immunohistochemistry
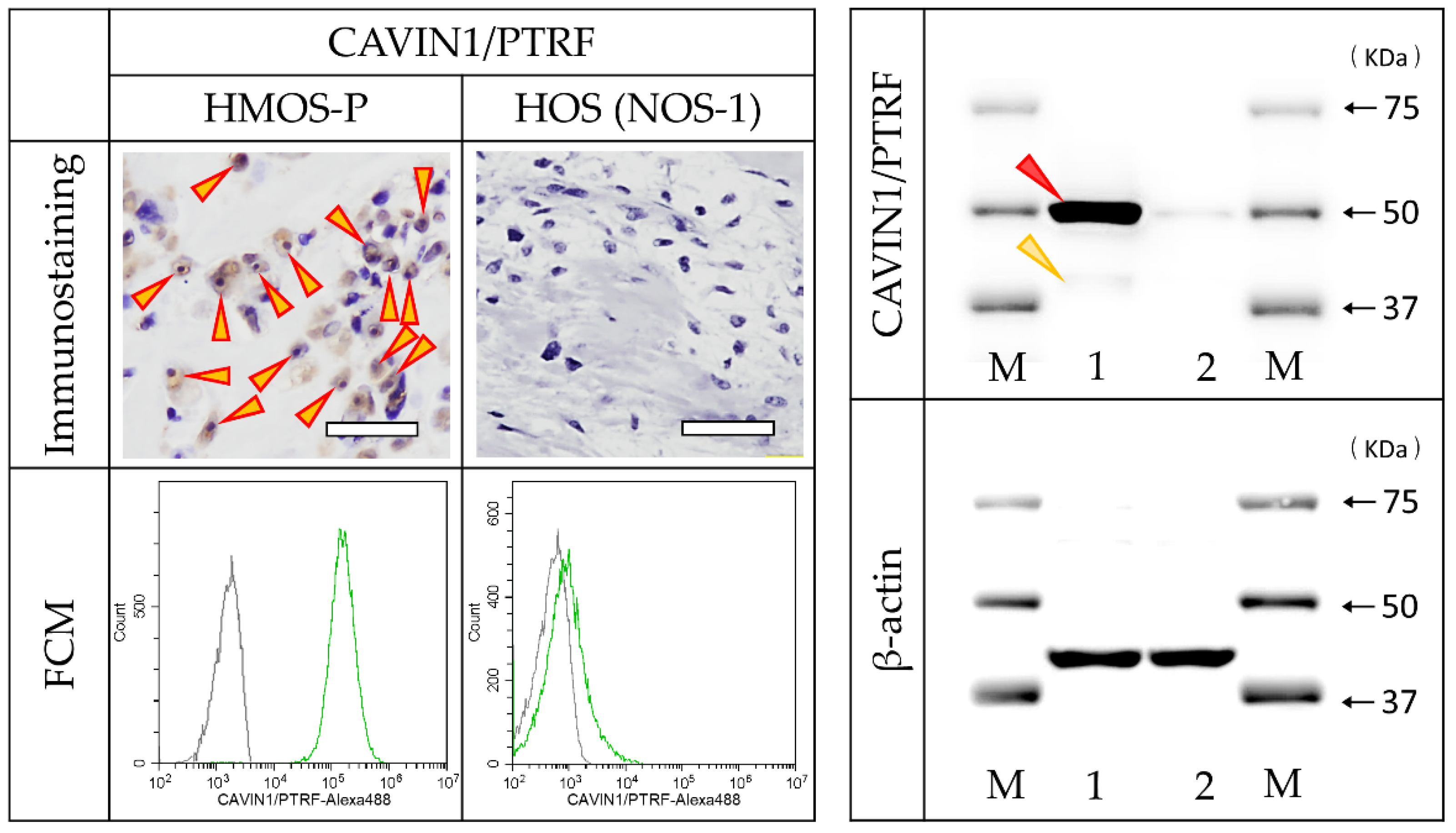
3.4. Identification of Antigen Reacting with Antibody
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, X.; Niu, G.; Yan, Y.; Yang, M.; Chen, K.; Ma, Y.; Chan, N.; Shen, B.; Chen, X. Phage display-derived peptides for osteosarcoma imaging. Clin. Cancer Res. 2010, 16, 4268–4277. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, L.; Roato, I. The Impact of Immune System in Regulating Bone Metastasis Formation by Osteotropic Tumors. J. Immunol. Res. 2015, 2015, 143526. [Google Scholar] [CrossRef] [PubMed]
- Senerchia, A.A.; Macedo, C.R.; Ferman, S.; Scopinaro, M.; Cacciavillano, W.; Boldrini, E.; Lins de Moraes, V.L.; Rey, G.; de Oliveira, C.T.; Castillo, L.; et al. Results of a randomized, prospective clinical trial evaluating metronomic chemotherapy in nonmetastatic patients with high-grade, operable osteosarcomas of the extremities: A report from the Latin American Group of Osteosarcoma Treatment. Cancer 2017, 123, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Wang, J.; Yang, L.; Zhao, Z.; Lu, K.; Zhao, L.; Xiao, J.; Li, Z.; Shi, Z. Targeting osteosarcoma vasculature with peptide obtained by phage display. Contemp. Oncol. 2014, 18, 165–170. [Google Scholar] [CrossRef]
- Fidler, I.J.; Yano, S.; Zhang, R.D.; Fujimaki, T.; Bucana, C.D. The seed and soil hypothesis: Vascularisation and brain metastases. Lancet Oncol. 2002, 3, 53–57. [Google Scholar] [CrossRef]
- Ramaswamy, S.; Ross, K.N.; Lander, E.S.; Golub, T.R. A molecular signature of metastasis in primary solid tumors. Nat. Genet. 2003, 33, 49–54. [Google Scholar] [CrossRef]
- Breuksch, I.; Weinert, M.; Brenner, W. The role of extracellular calcium in bone metastasis. J. Bone Oncol. 2016, 5, 143–145. [Google Scholar] [CrossRef]
- Satcher, R.L.; Pan, T.; Cheng, C.J.; Lee, Y.C.; Lin, S.C.; Yu, G.; Li, X.; Hoang, A.G.; Tamboli, P.; Jonasch, E.; et al. Cadherin-11 in renal cell carcinoma bone metastasis. PLoS ONE 2014, 9, e89880. [Google Scholar] [CrossRef]
- Pulido, C.; Vendrell, I.; Ferreira, A.R.; Casimiro, S.; Mansinho, A.; Alho, I.; Costa, L. Bone metastasis risk factors in breast cancer. Ecancermedicalscience 2017, 11, 715. [Google Scholar] [CrossRef]
- Holen, I.; Lefley, D.V.; Francis, S.E.; Rennicks, S.; Bradbury, S.; Coleman, R.E.; Ottewell, P. IL-1 drives breast cancer growth and bone metastasis in vivo. Oncotarget 2016, 7, 75571–75584. [Google Scholar] [CrossRef]
- Mi, F.; Gong, L. Secretion of interleukin-6 by bone marrow mesenchymal stem cells promotes metastasis in hepatocellular carcinoma. Biosci. Rep. 2017, 37, BSR20170181. [Google Scholar] [CrossRef] [PubMed]
- Arihiro, K.; Inai, K. Expression of CD31, Met/hepatocyte growth factor receptor and bone morphogenetic protein in bone metastasis of osteosarcoma. Pathol. Int. 2001, 51, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.D.; Doyonnas, R.; Newton, J.P.; Blystone, S.D.; Brown, E.J.; Watt, S.M.; Simmons, D.L. Identification of alpha v beta 3 as a heterotypic ligand for CD31/PECAM-1. J. Cell Sci. 1996, 109 Pt 2, 437–445. [Google Scholar] [CrossRef]
- Stuiver, I.; Ruggeri, Z.; Smith, J.W. Divalent cations regulate the organization of integrins alpha v beta 3 and alpha v beta 5 on the cell surface. J. Cell Physiol. 1996, 168, 521–531. [Google Scholar] [CrossRef]
- Bami, M.; Mavrogenis, A.F.; Angelini, A.; Milonaki, M.; Mitsiokapa, E.; Stamoulis, D.; Soucacos, P.N. Bone morphogenetic protein signaling in musculoskeletal cancer. J. Cancer Res. Clin. Oncol. 2016, 142, 2061–2072. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.; Scott, M.A.; Dry, S.M.; James, A.W. Roles of bone morphogenetic protein signaling in osteosarcoma. Int. Orthop. 2014, 38, 2313–2322. [Google Scholar] [CrossRef] [PubMed]
- Lammli, J.; Fan, M.; Rosenthal, H.G.; Patni, M.; Rinehart, E.; Vergara, G.; Ablah, E.; Wooley, P.H.; Lucas, G.; Yang, S.Y. Expression of Vascular Endothelial Growth Factor correlates with the advance of clinical osteosarcoma. Int. Orthop. 2012, 36, 2307–2313. [Google Scholar] [CrossRef]
- Parham, D.M.; Pratt, C.B.; Parvey, L.S.; Webber, B.L.; Champion, J. Childhood multifocal osteosarcoma. Clinicopathologic and radiologic correlates. Cancer 1985, 55, 2653–2658. [Google Scholar] [CrossRef]
- Corradi, D.; Wenger, D.E.; Bertoni, F.; Bacchini, P.; Bosio, S.; Goldoni, M.; Unni, K.K.; Sim, F.H.; Inwards, C.Y. Multicentric osteosarcoma: Clinicopathologic and radiographic study of 56 cases. Am. J. Clin. Pathol. 2011, 136, 799–807. [Google Scholar] [CrossRef]
- Bacci, G.; Fabbri, N.; Balladelli, A.; Forni, C.; Palmerini, E.; Picci, P. Treatment and prognosis for synchronous multifocal osteosarcoma in 42 patients. J. Bone Jt. Surg. Br. 2006, 88, 1071–1075. [Google Scholar] [CrossRef]
- Hameed, S.; Vijayan, S.; Naik, M.; Rao, S. Multicentric osteosarcoma. Singap. Med. J. 2012, 53, e214–e217. [Google Scholar]
- Yamamoto, Y.; Yamamoto, N.; Tajima, K.; Ohno, A.; Washimi, Y.; Ishimura, D.; Washimi, O.; Yamada, H. Characterization of human multicentric osteosarcoma using newly established cells derived from multicentric osteosarcoma. J. Cancer Res. Clin. Oncol. 2011, 137, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Akahori, Y.; Kurosawa, G.; Sumitomo, M.; Morita, M.; Muramatsu, C.; Eguchi, K.; Tanaka, M.; Suzuki, K.; Sugiura, M.; Iba, Y.; et al. Isolation of antigen/antibody complexes through organic solvent (ICOS) method. Biochem. Biophys. Res. Commun. 2009, 378, 832–835. [Google Scholar] [CrossRef]
- Kurosawa, G.; Sumitomo, M.; Ukai, Y.; Subere, J.; Muramatsu, C.; Eguchi, K.; Tanaka-Hashiba, M.; Sugiura, M.; Ando, M.; Sato, N.; et al. Selection and analysis of anti-cancer antibodies for cancer therapy obtained from antibody phage library. Cancer Sci. 2011, 102, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, G.; Sugiura, M.; Hattori, Y.; Tsuda, H.; Kurosawa, Y. Classification of 27 Tumor-Associated Antigens by Histochemical Analysis of 36 Freshly Resected Lung Cancer Tissues. Int. J. Mol. Sci. 2016, 17, 1862. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, G.; Kondo, M.; Kurosawa, Y. A variety of human monoclonal antibodies against epidermal growth factor receptor isolated from a phage antibody library. Biochem. Biophys. Res. Commun. 2016, 480, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, G.; Akahori, Y.; Morita, M.; Sumitomo, M.; Sato, N.; Muramatsu, C.; Eguchi, K.; Matsuda, K.; Takasaki, A.; Tanaka, M.; et al. Comprehensive screening for antigens overexpressed on carcinomas via isolation of human mAbs that may be therapeutic. Proc. Natl. Acad. Sci. USA 2008, 105, 7287–7292. [Google Scholar] [CrossRef]
- Ye, J.; Ma, N.; Madden, T.L.; Ostell, J.M. IgBLAST: An immunoglobulin variable domain sequence analysis tool. Nucleic Acids Res. 2013, 41, W34–W40. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Govender, P.; Romero, F.; Shah, D.; Paez, J.; Ding, S.Y.; Liu, L.; Gower, A.; Baez, E.; Aly, S.S.; Pilch, P.; et al. Cavin1; a regulator of lung function and macrophage phenotype. PLoS ONE 2013, 8, e62045. [Google Scholar] [CrossRef]
- Tillu, V.A.; Lim, Y.W.; Kovtun, O.; Mureev, S.; Ferguson, C.; Bastiani, M.; McMahon, K.A.; Lo, H.P.; Hall, T.E.; Alexandrov, K.; et al. A variable undecad repeat domain in cavin1 regulates caveola formation and stability. EMBO Rep. 2018, 19, e45775. [Google Scholar] [CrossRef]
- Lajoie, P.; Nabi, I.R. Lipid rafts, caveolae, and their endocytosis. Int. Rev. Cell Mol. Biol. 2010, 282, 135–163. [Google Scholar] [CrossRef]
- Aboulaich, N.; Vainonen, J.P.; Stralfors, P.; Vener, A.V. Vectorial proteomics reveal targeting, phosphorylation and specific fragmentation of polymerase I and transcript release factor (PTRF) at the surface of caveolae in human adipocytes. Biochem. J. 2004, 383, 237–248. [Google Scholar] [CrossRef]
- Kovtun, O.; Tillu, V.A.; Ariotti, N.; Parton, R.G.; Collins, B.M. Cavin family proteins and the assembly of caveolae. J. Cell Sci. 2015, 128, 1269–1278. [Google Scholar] [CrossRef]
- Hasegawa, T.; Takeuchi, A.; Miyaishi, O.; Xiao, H.; Mao, J.; Isobe, K. PTRF (polymerase I and transcript-release factor) is tissue-specific and interacts with the BFCOL1 (binding factor of a type-I collagen promoter) zinc-finger transcription factor which binds to the two mouse type-I collagen gene promoters. Biochem. J. 2000, 347 Pt 1, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Merchant, J.L. ZBP-89 promotes growth arrest through stabilization of p53. Mol. Cell. Biol. 2001, 21, 4670–4683. [Google Scholar] [CrossRef]
- Bai, L.; Deng, X.; Li, J.; Wang, M.; Li, Q.; An, W.; Deli, A.; Cong, Y.S. Regulation of cellular senescence by the essential caveolar component PTRF/Cavin-1. Cell Res. 2011, 21, 1088–1101. [Google Scholar] [CrossRef] [PubMed]
- Wiechen, K.; Diatchenko, L.; Agoulnik, A.; Scharff, K.M.; Schober, H.; Arlt, K.; Zhumabayeva, B.; Siebert, P.D.; Dietel, M.; Schäfer, R.; et al. Caveolin-1 is down-regulated in human ovarian carcinoma and acts as a candidate tumor suppressor gene. Am. J. Pathol. 2001, 159, 1635–1643. [Google Scholar] [CrossRef] [PubMed]
- Faggi, F.; Chiarelli, N.; Colombi, M.; Mitola, S.; Ronca, R.; Madaro, L.; Bouche, M.; Poliani, P.L.; Vezzoli, M.; Longhena, F.; et al. Cavin-1 and Caveolin-1 are both required to support cell proliferation, migration and anchorage-independent cell growth in rhabdomyosarcoma. Lab. Investig. 2015, 95, 585–602. [Google Scholar] [CrossRef]
- Huertas-Martinez, J.; Court, F.; Rello-Varona, S.; Herrero-Martin, D.; Almacellas-Rabaiget, O.; Sainz-Jaspeado, M.; Garcia-Monclus, S.; Lagares-Tena, L.; Buj, R.; Hontecillas-Prieto, L.; et al. DNA methylation profiling identifies PTRF/Cavin-1 as a novel tumor suppressor in Ewing sarcoma when co-expressed with caveolin-1. Cancer Lett. 2017, 386, 196–207. [Google Scholar] [CrossRef]
- Hill, M.M.; Bastiani, M.; Luetterforst, R.; Kirkham, M.; Kirkham, A.; Nixon, S.J.; Walser, P.; Abankwa, D.; Oorschot, V.M.; Martin, S.; et al. PTRF-Cavin, a conserved cytoplasmic protein required for caveola formation and function. Cell 2008, 132, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Nohe, A.; Keating, E.; Underhill, T.M.; Knaus, P.; Petersen, N.O. Dynamics and interaction of caveolin-1 isoforms with BMP-receptors. J. Cell Sci. 2005, 118, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Hartung, A.; Bitton-Worms, K.; Rechtman, M.M.; Wenzel, V.; Boergermann, J.H.; Hassel, S.; Henis, Y.I.; Knaus, P. Different routes of bone morphogenic protein (BMP) receptor endocytosis influence BMP signaling. Mol. Cell. Biol. 2006, 26, 7791–7805. [Google Scholar] [CrossRef]
- Takagi, S.; Sasaki, Y.; Koike, S.; Takemoto, A.; Seto, Y.; Haraguchi, M.; Ukaji, T.; Kawaguchi, T.; Sugawara, M.; Saito, M.; et al. Platelet-derived lysophosphatidic acid mediated LPAR1 activation as a therapeutic target for osteosarcoma metastasis. Oncogene 2021, 40, 5548–5558. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | No. 10 | No. 12 | No. 17 | No. 19 | No. 43 | No. 77 |
|---|---|---|---|---|---|---|
| HMOS-A | 90.3 ± 1.1 | 88.9 ± 1.5 | 88.8 ± 1.5 | 76.1 ± 1.4 | 92.1 ± 1.0 | 92.9 ± 1.1 |
| HMOS-P | 77.5 ± 2.1 | 78.8 ± 2.3 | 77.1 ± 1.9 | 63.2 ± 2.2 | 77.8 ± 2.0 | 82.9 ± 1.7 |
| MNNG-HOS | 96.8 ± 0.9 | 3.4 ± 0.2 | 0.3 ± 0.1 | 0.2 ± 0.1 | 91.1 ± 1.4 | 45.4 ± 5.1 |
| MG63 | 76.4 ± 1.5 | 1.0 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 | 41.5 ± 6.3 | 24.7 ± 2.1 |
| Saos2 | 22.3 ± 1.9 | 0.6 ± 0.2 | 68.2 ± 3.6 | 0.7 ± 0.2 | 96.1 ± 0.7 | 14.4 ± 1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayashi, T.; Yamamoto, N.; Kurosawa, G.; Tajima, K.; Kondo, M.; Hiramatsu, N.; Kato, Y.; Tanaka, M.; Yamaguchi, H.; Kurosawa, Y.; et al. A Novel High-Throughput Screening Method for a Human Multicentric Osteosarcoma-Specific Antibody and Biomarker Using a Phage Display-Derived Monoclonal Antibody. Cancers 2022, 14, 5829. https://doi.org/10.3390/cancers14235829
Hayashi T, Yamamoto N, Kurosawa G, Tajima K, Kondo M, Hiramatsu N, Kato Y, Tanaka M, Yamaguchi H, Kurosawa Y, et al. A Novel High-Throughput Screening Method for a Human Multicentric Osteosarcoma-Specific Antibody and Biomarker Using a Phage Display-Derived Monoclonal Antibody. Cancers. 2022; 14(23):5829. https://doi.org/10.3390/cancers14235829
Chicago/Turabian StyleHayashi, Takuma, Naoki Yamamoto, Gene Kurosawa, Kaori Tajima, Mariko Kondo, Noriko Hiramatsu, Yu Kato, Miho Tanaka, Hisateru Yamaguchi, Yoshikazu Kurosawa, and et al. 2022. "A Novel High-Throughput Screening Method for a Human Multicentric Osteosarcoma-Specific Antibody and Biomarker Using a Phage Display-Derived Monoclonal Antibody" Cancers 14, no. 23: 5829. https://doi.org/10.3390/cancers14235829
APA StyleHayashi, T., Yamamoto, N., Kurosawa, G., Tajima, K., Kondo, M., Hiramatsu, N., Kato, Y., Tanaka, M., Yamaguchi, H., Kurosawa, Y., Yamada, H., & Fujita, N. (2022). A Novel High-Throughput Screening Method for a Human Multicentric Osteosarcoma-Specific Antibody and Biomarker Using a Phage Display-Derived Monoclonal Antibody. Cancers, 14(23), 5829. https://doi.org/10.3390/cancers14235829