
Effects of Long Non-Coding RNAs Induced by the Gut Microbiome on Regulating the Development of Colorectal Cancer

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
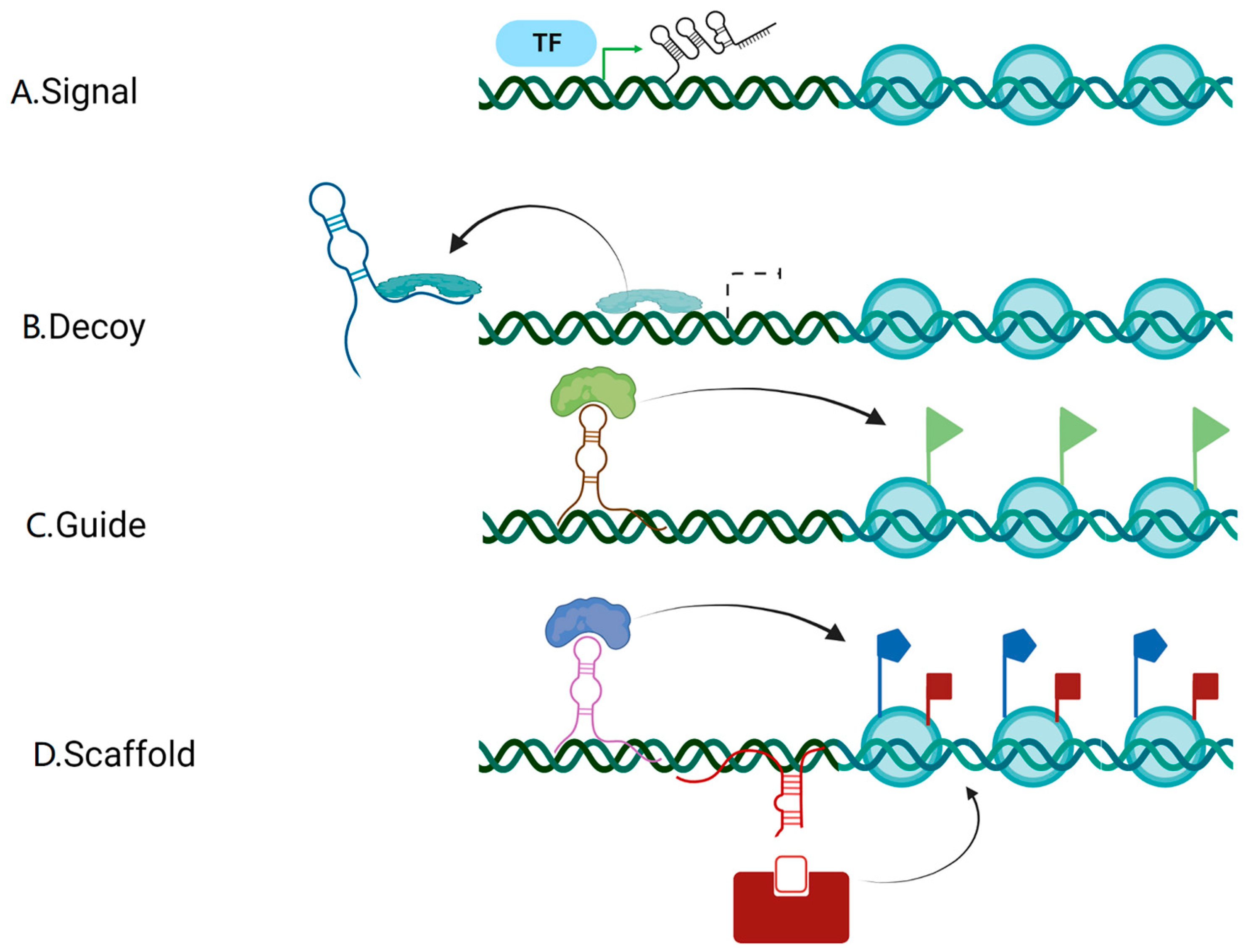
2. Biogenesis and the Mode of Action of lncRNAs
3. The Development of CRC Promoted by the Disturbance of the Gut Microbiome
4. Microbial Regulation of CRC Mediated by lncRNAs
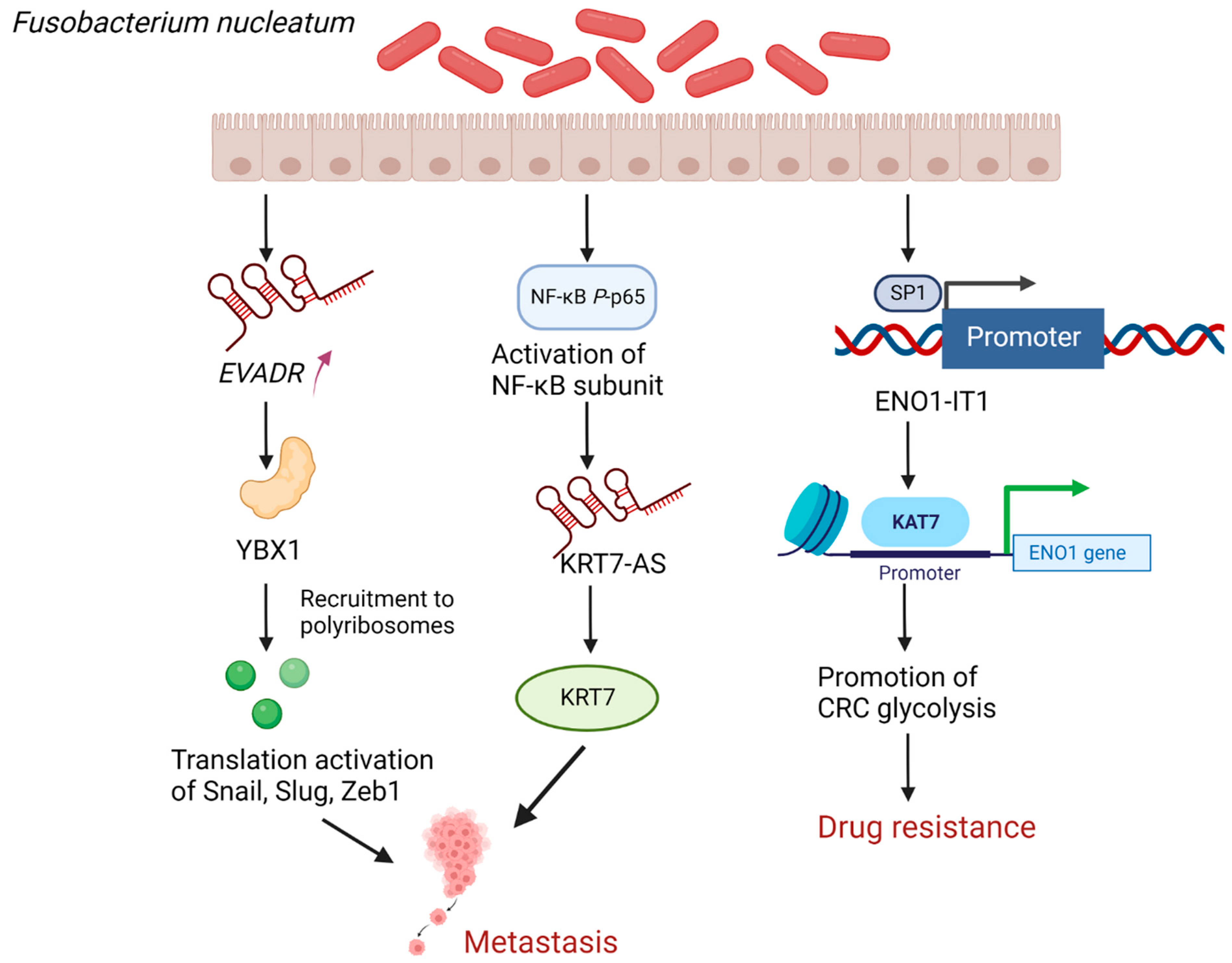
4.1. Fusobacterium Nucleatum (F. nucleatum) Promotes CRC Metastasis as Well as Enhances Drug Resistance by Regulating lncRNAs
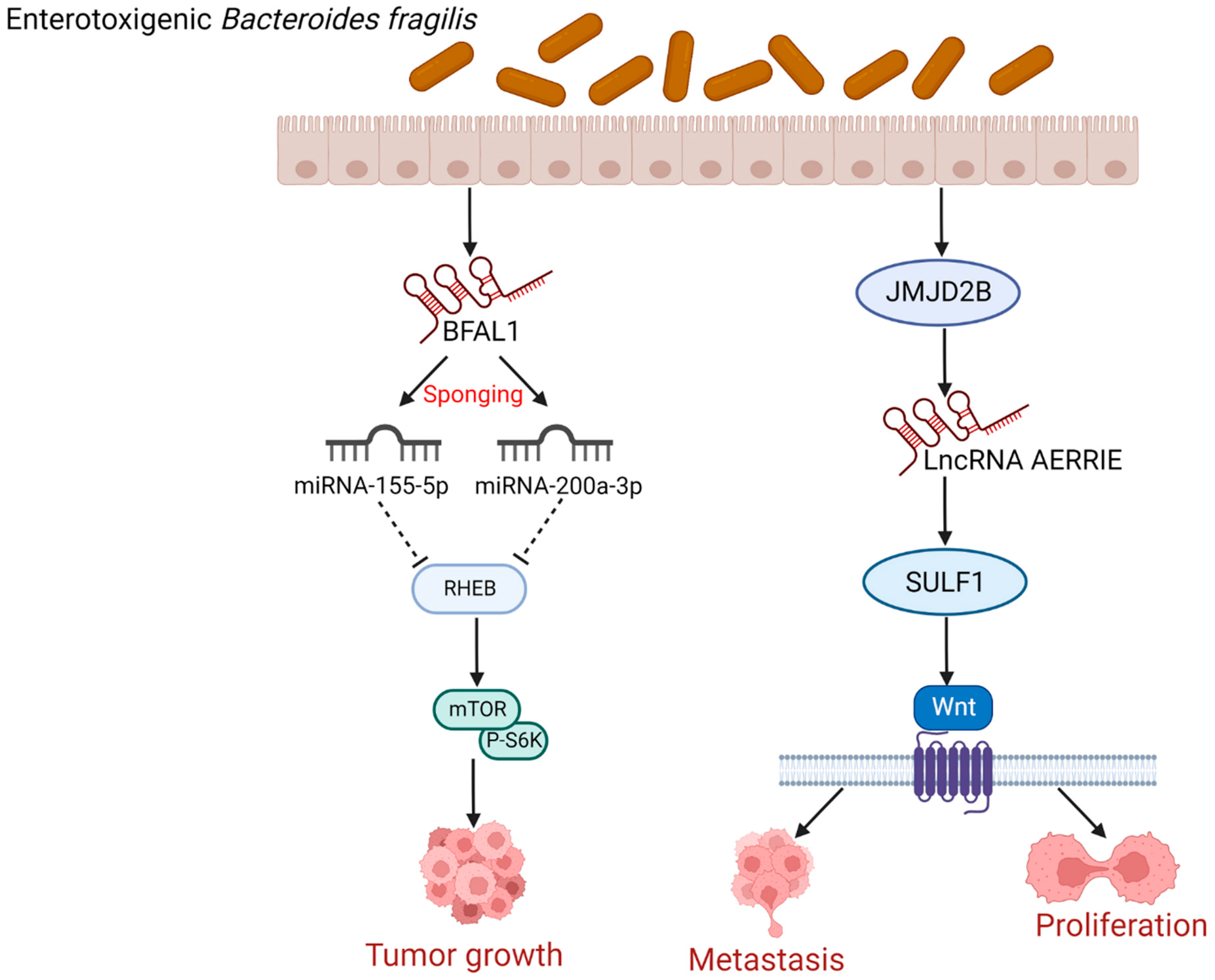
4.2. Enterotoxigenic Bacteroides Fragilis (ETBF) Induces CRC Cells Growth, Proliferation, and Metastasis by Regulating lncRNAs
5. The Gut Microbiome Metabolic Regulation of CRC Mediated by lncRNAs
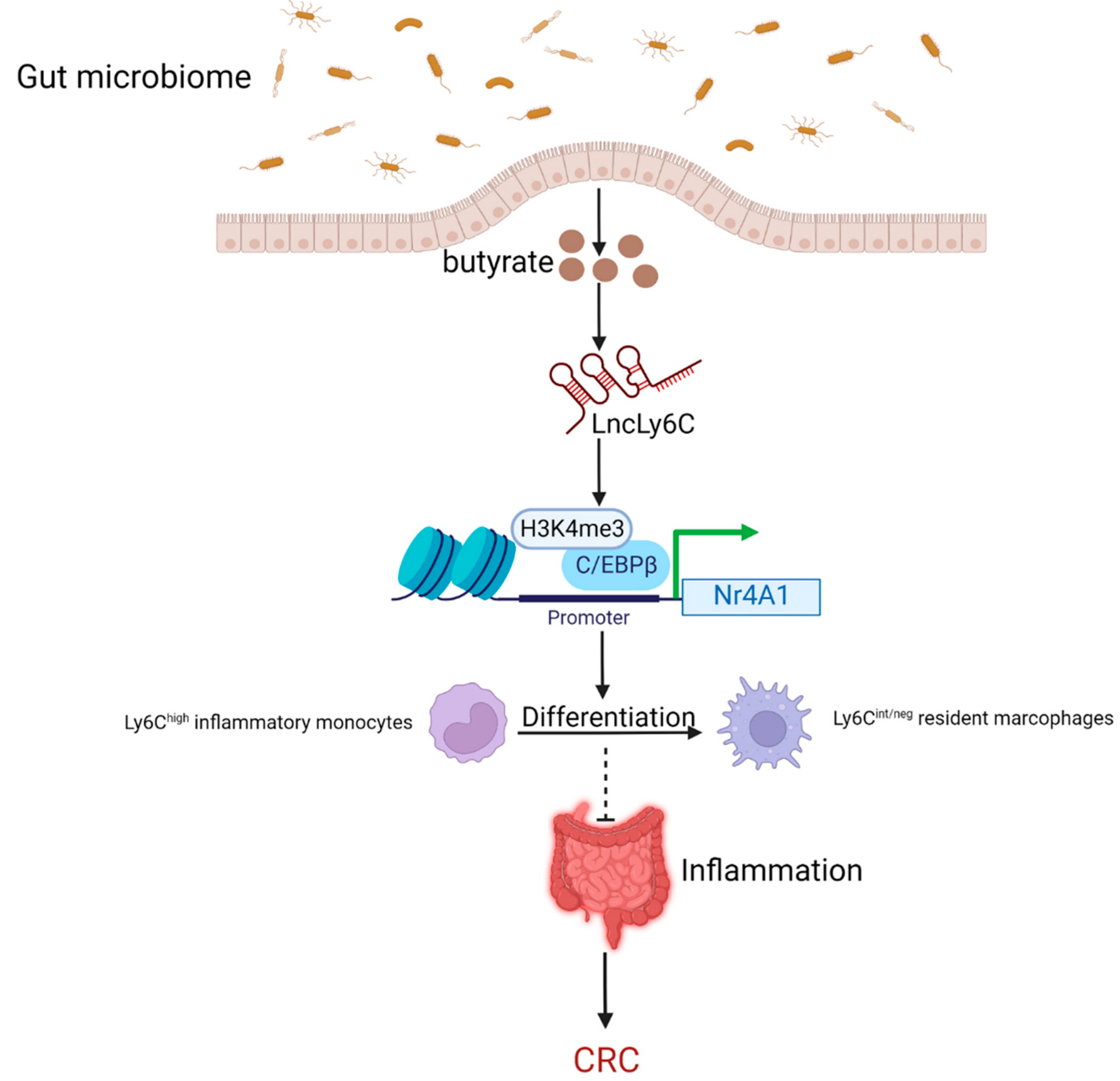
5.1. Butyrate as the Metabolite of the Gut Microbiome Inhibits the Intestinal Inflammation to Lower the Risk of CRC by Regulating lncRNAs
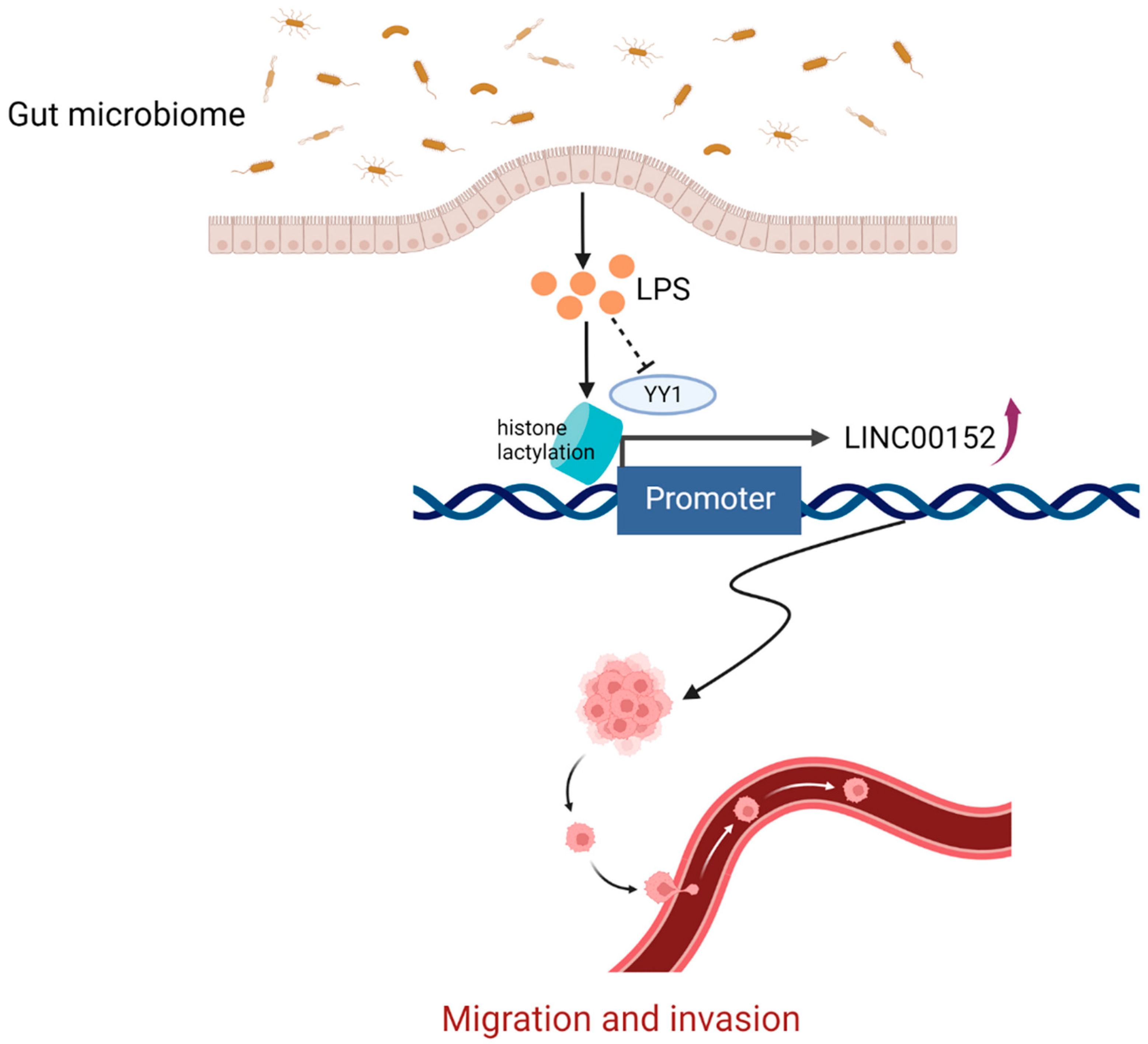
5.2. The Gut Microbiome-Derived Lipopolysaccharide (LPS) Promotes Cancer Cells Migration and Invasion by Regulating lncRNAs
6. Strategies of Preventing and Controlling CRC Based on the Regulatory Role of the Gut Microbiome on lncRNAs
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CRC | Colorectal cancer |
| lncRNAs | long non-coding RNAs |
| TME | tumor microenvironment |
| EMT | epithelial mesenchymal transition |
| NF-κB | nuclear factor kappa-B |
| Pol II | RNA polymerase II |
| NXF1 | nuclear RNA export factor 1 |
| TFs | transcription factors |
| KRT7-AS | Keratin7-antisense |
| KRT7 | Keratin7 |
| F. nucleatum | Fusobacterium nucleatum |
| EVADR | endogenous retroviral-associated adenocarcinoma lncRNA |
| EMT-TFs | EMT transcription factors |
| RBP | RNA-binding protein |
| ENO1-IT1 | enolase1-intronic transcript 1 |
| 5-FU | 5-fluorouracil |
| ETBF | Enterotoxigenic Bacteroides fragilis |
| BFT | Bacteroides fragilis toxin |
| BFAL1 | B. fragilis-associated lncRNA1 |
| RHEB | Ras homolog enriched in brain |
| CAFs | cancer-associated fibroblasts |
| HuR | human antigen R |
| FOXA1 | forkhead box protein A1 |
| JMJD2B | jumonji domain-containing protein 2B |
| SULF1 | sulfatase 1 |
| C/EBPβ | CCAAT/enhancer binding protein β |
| NR4A | nuclear subfamily 4 |
| IL | interleukin |
| IECs | intestinal epithelial cells |
| CAC | colitis-associated colorectal cancer |
| LPS | lipopolysaccharide |
| NGP | next-generation probiotics |
| SCFAs | short-chain fatty acids |
| FMT | fecal microbiota transplantation |
| CDI | Clostridium difficile infection |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Fang, J.-Y. Fusobacterium nucleatum, a key pathogenic factor and microbial biomarker for colorectal cancer. Trends Microbiol. 2022. [Google Scholar] [CrossRef]
- Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.; et al. Supporting data for the paper: “An integrated catalog of reference genes in the human gut microbiome”. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Nowak, J.A.; Milner, D.A., Jr.; Song, M.; Ogino, S. Integration of microbiology, molecular pathology, and epidemiology: A new paradigm to explore the pathogenesis of microbiome-driven neoplasms. J. Pathol. 2019, 247, 615–628. [Google Scholar] [CrossRef]
- Chen, Q.; Zhou, L.; De Ma, D.; Hou, J.; Lin, Y.; Wu, J.; Tao, M. LncRNA GAS6-AS1 facilitates tumorigenesis and metastasis of colorectal cancer by regulating TRIM14 through miR-370-3p/miR-1296-5p and FUS. J. Transl. Med. 2022, 20, 356. [Google Scholar] [CrossRef]
- Zhou, L.; Jiang, J.; Huang, Z.; Jin, P.; Peng, L.; Luo, M.; Zhang, Z.; Chen, Y.; Xie, N.; Gao, W.; et al. Hypoxia-induced lncRNA STEAP3-AS1 activates Wnt/beta-catenin signaling to promote colorectal cancer progression by preventing m(6)A-mediated degradation of STEAP3 mRNA. Mol. Cancer 2022, 21, 168. [Google Scholar] [CrossRef]
- Li, J.; Ji, Y.; Chen, N.; Wang, H.; Fang, C.; Yin, X.; Jiang, Z.; Dong, Z.; Zhu, D.; Fu, J.; et al. A specific upregulated long noncoding RNA in colorectal cancer promotes cancer progression. JCI Insight 2022, 7. [Google Scholar] [CrossRef]
- Chu, J.; Fang, X.; Sun, Z.; Gai, L.; Dai, W.; Li, H.; Yan, X.; Du, J.; Zhang, L.; Zhao, L.; et al. Non-Coding RNAs Regulate the Resistance to Anti-EGFR Therapy in Colorectal Cancer. Front. Oncol. 2022, 11, 801319. [Google Scholar] [CrossRef]
- Beermann, J.; Piccoli, M.-T.; Viereck, J.; Thum, T. Non-coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Yu, X.; Zheng, Q.; Zhang, S.; Guo, W. Oncogenic roles of LINC01234 in various forms of human cancer. Biomed. Pharmacother. 2022, 154, 113570. [Google Scholar] [CrossRef]
- da Silveira, W.A.; Renaud, L.; Hazard, E.S.; Hardiman, G. miRNA and lncRNA Expression Networks Modulate Cell Cycle and DNA Repair Inhibition in Senescent Prostate Cells. Genes 2022, 13, 208. [Google Scholar] [CrossRef] [PubMed]
- Ketley, R.F.; Gullerova, M. Jack of all trades? The versatility of RNA in DNA double-strand break repair. Essays Biochem. 2020, 64, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, M.; Zhang, M.; Sun, Q.; Zeng, S.; Zhao, H.; Yang, H.; Liu, M.; Ren, S.; Meng, X.; et al. The Role of non-coding RNAs in colorectal cancer, with a focus on its autophagy. Pharmacol. Ther. 2021, 226, 107868. [Google Scholar] [CrossRef]
- Liang, L.; Ai, L.; Qian, J.; Fang, J.-Y.; Xu, J. Long noncoding RNA expression profiles in gut tissues constitute molecular signatures that reflect the types of microbes. Sci. Rep. 2015, 5, 11763. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Su, T.; Zhang, Y.; Lee, A.; He, J.; Ge, Q.; Wang, L.; Si, J.; Zhuo, W.; Wang, L. Fusobacterium nucleatum promotes colorectal cancer metastasis by modulating KRT7-AS/KRT7. Gut Microbes 2020, 11, 511–525. [Google Scholar] [CrossRef]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 Catalog of Human Long Noncoding RNAs: Analysis of Their Gene Structure, Evolution, and Expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef]
- Nojima, T.; Proudfoot, N.J. Mechanisms of lncRNA biogenesis as revealed by nascent transcriptomics. Nat. Rev. Mol. Cell Biol. 2022, 23, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, S.; Ye, Z.; Wang, W.; Hu, X.; Hang, Q. Long Non-Coding RNAs in Pancreatic Cancer: Biologic Functions, Mechanisms, and Clinical Significance. Cancers 2022, 14, 2115. [Google Scholar] [CrossRef]
- Zuckerman, B.; Ron, M.; Mikl, M.; Segal, E.; Ulitsky, I. Gene Architecture and Sequence Composition Underpin Selective Dependency of Nuclear Export of Long RNAs on NXF1 and the TREX Complex. Mol. Cell 2020, 79, 251–267.e6. [Google Scholar] [CrossRef]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 206. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, L.; Liu, D. HOTAIR regulates colorectal cancer stem cell properties and promotes tumorigenicity by sponging miR-211-5p and modulating FLT-1. Cell Cycle 2021, 20, 1999–2009. [Google Scholar] [CrossRef]
- Saus, E.; Brunet-Vega, A.; Iraola-Guzmán, S.; Pegueroles, C.; Gabaldón, T.; Pericay, C. Long Non-Coding RNAs As Potential Novel Prognostic Biomarkers in Colorectal Cancer. Front. Genet. 2016, 7, 54. [Google Scholar] [CrossRef]
- Zhou, L.; Li, J.; Tang, Y.; Yang, M. Exosomal LncRNA LINC00659 transferred from cancer-associated fibroblasts promotes colorectal cancer cell progression via miR-342-3p/ANXA2 axis. J. Transl. Med. 2021, 19, 8. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zheng, L.; Ye, J.; Zhang, C.; Zhou, J.; Huang, Q.; Guo, Y.; Wang, L.; Yu, P.; Liu, S.; et al. CREB1 contributes colorectal cancer cell plasticity by regulating lncRNA CCAT1 and NF-kappaB pathways. Sci. China Life Sci. 2022, 65, 1481–1497. [Google Scholar] [CrossRef]
- Zhang, F.; Aschenbrenner, D.; Yoo, J.Y.; Zuo, T. The gut mycobiome in health, disease, and clinical applications in association with the gut bacterial microbiome assembly. Lancet Microbe 2022. [Google Scholar] [CrossRef] [PubMed]
- Kendong, S.M.A.; Ali, R.A.R.; Nawawi, K.N.M.; Ahmad, H.F.; Mokhtar, N.M. Gut Dysbiosis and Intestinal Barrier Dysfunction: Potential Explanation for Early-Onset Colorectal Cancer. Front. Cell. Infect. Microbiol. 2021, 11, 744606. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Ma, L.; Wei, W.; Li, X.; Chang, Y.; Pan, Z.; Gao, H.; Yang, R.; Bi, Y.; Ding, L. Metagenomics analysis of cultured mucosal bacteria from colorectal cancer and adjacent normal mucosal tissues. J. Med. Microbiol. 2022, 71, 001523. [Google Scholar] [CrossRef]
- Gao, R.; Xia, K.; Wu, M.; Zhong, H.; Sun, J.; Zhu, Y.; Huang, L.; Wu, X.; Yin, L.; Yang, R.; et al. Alterations of Gut Mycobiota Profiles in Adenoma and Colorectal Cancer. Front. Cell. Infect. Microbiol. 2022, 12, 839435. [Google Scholar] [CrossRef] [PubMed]
- Coker, O.O.; Liu, C.; Wu, W.K.K.; Wong, S.H.; Jia, W.; Sung, J.J.Y.; Yu, J. Altered gut metabolites and microbiota interactions are implicated in colorectal carcinogenesis and can be non-invasive diagnostic biomarkers. Microbiome 2022, 10, 35. [Google Scholar] [CrossRef]
- Sears, C.L.; Pardoll, D.M. Perspective: Alpha-Bugs, Their Microbial Partners, and the Link to Colon Cancer. J. Infect. Dis. 2011, 203, 306–311. [Google Scholar] [CrossRef]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A bacterial Driver–passenger model for colorectal cancer: Beyond the usual suspects. Nat. Rev. Microbiol. 2012, 10, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Fang, Y.; Zhang, W.; Zhang, H.; Tang, D.; Wang, D. Bacterial driver–passenger model in biofilms: A new mechanism in the development of colorectal cancer. Clin. Transl. Oncol. 2022, 24, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Z.; Xu, Y.; Wang, Y.; Wang, F.; Zhang, Q.; Ni, C.; Zhen, Y.; Xu, R.; Liu, Q.; et al. Enterobacterial LPS-inducible LINC00152 is regulated by histone lactylation and promotes cancer cells invasion and migration. Front. Cell. Infect. Microbiol. 2022, 12, 913815. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, J.; Qi, H.; Wei, J.; Yang, Y.; Yue, J.; Liu, X.; Zhang, Y.; Yang, R. LncRNA lncLy6C induced by microbiota metabolite butyrate promotes differentiation of Ly6C(high) to Ly6C(int/neg) macrophages through lncLy6C/C/EBPbeta/Nr4A1 axis. Cell Discov. 2020, 6, 87. [Google Scholar] [CrossRef]
- Lu, X.; Xu, Q.; Tong, Y.; Zhang, Z.; Dun, G.; Feng, Y.; Tang, J.; Han, D.; Mao, Y.; Deng, L.; et al. Long non-coding RNA EVADR induced by Fusobacterium nucleatum infection promotes colorectal cancer metastasis. Cell Rep. 2022, 40, 111127. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Lv, C.; Yu, H.; Wang, K.; Chen, C.; Tang, J.; Han, F.; Mai, M.; Ye, K.; Lai, M.; Zhang, H. ENO2 Promotes Colorectal Cancer Metastasis by Interacting with the LncRNA CYTOR and Activating YAP1-Induced EMT. Cells 2022, 11, 2363. [Google Scholar] [CrossRef]
- Puisieux, A.; Brabletz, T.; Caramel, J. Oncogenic roles of EMT-inducing transcription factors. Nat. Cell Biol. 2014, 16, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Pavlič, A.; Hauptman, N.; Boštjančič, E.; Zidar, N. Long Non-Coding RNAs as Potential Regulators of EMT-Related Transcription Factors in Colorectal Cancer—A Systematic Review and Bioinformatics Analysis. Cancers 2022, 14, 2280. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.; Todorova, K.; Wang, J.; Horos, R.; Lee, K.K.; Neel, V.A.; Negri, G.L.; Sorensen, P.H.; Lee, S.W.; Hentze, M.W.; et al. The RNA-binding protein YBX1 regulates epidermal progenitors at a posttranscriptional level. Nat. Commun. 2018, 9, 1734. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, A.M.; Veinotte, C.J.; Cheng, H.; Grunewald, T.G.; Negri, G.L.; Somasekharan, S.P.; Corkery, D.P.; Tirode, F.; Mathers, J.; Khan, D.; et al. Translational Activation of HIF1α by YB-1 Promotes Sarcoma Metastasis. Cancer Cell 2015, 27, 682–697. [Google Scholar] [CrossRef]
- Huang, B.; Song, J.H.; Cheng, Y.; Abraham, J.M.; Ibrahim, S.; Sun, Z.; Ke, X.; Meltzer, S.J. Long non-coding antisense RNA KRT7-AS is activated in gastric cancers and supports cancer cell progression by increasing KRT7 expression. Oncogene 2016, 35, 4927–4936. [Google Scholar] [CrossRef] [PubMed]
- Communal, L.; Roy, N.; Cahuzac, M.; Rahimi, K.; Köbel, M.; Provencher, D.; Mes-Masson, A.-M. A Keratin 7 and E-Cadherin Signature Is Highly Predictive of Tubo-Ovarian High-Grade Serous Carcinoma Prognosis. Int. J. Mol. Sci. 2021, 22, 5325. [Google Scholar] [CrossRef]
- Helfand, B.T.; Chang, L.; Goldman, R.D. Intermediate filaments are dynamic and motile elements of cellular architecture. J. Cell Sci. 2004, 117, 133–141. [Google Scholar] [CrossRef]
- Capece, D.; Verzella, D.; Flati, I.; Arboretto, P.; Cornice, J.; Franzoso, G. NF-kappaB: Blending metabolism, immunity, and inflammation. Trends Immunol. 2022, 43, 757–775. [Google Scholar] [CrossRef]
- Hong, J.; Guo, F.; Lu, S.-Y.; Shen, C.; Ma, D.; Zhang, X.; Xie, Y.; Yan, T.; Yu, T.; Sun, T.; et al. F. nucleatum targets lncRNA ENO1-IT1 to promote glycolysis and oncogenesis in colorectal cancer. Gut 2020, 70, 2123–2137. [Google Scholar] [CrossRef]
- Wang, G.; Wang, J.; Yin, P.; Xu, K.; Wang, Y.; Shi, F.; Gao, J.; Fu, X. New strategies for targeting glucose metabolism–mediated acidosis for colorectal cancer therapy. J. Cell. Physiol. 2018, 234, 348–368. [Google Scholar] [CrossRef]
- Ganapathy-Kanniappan, S.; Geschwind, J.-F.H. Tumor glycolysis as a target for cancer therapy: Progress and prospects. Mol. Cancer 2013, 12, 152. [Google Scholar] [CrossRef] [PubMed]
- Lan, R.; Wang, Q. Deciphering structure, function and mechanism of lysine acetyltransferase HBO1 in protein acetylation, transcription regulation, DNA replication and its oncogenic properties in cancer. Cell. Mol. Life Sci. 2020, 77, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Sapountzi, V.; Côté, J. MYST-family histone acetyltransferases: Beyond chromatin. Cell. Mol. Life Sci. 2010, 68, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Cho, H.I.; Park, S.H.; Kim, J.H.; Chai, Y.G.; Jang, Y.K. The histone acetyltransferase Myst2 regulates Nanog expression, and is involved in maintaining pluripotency and self-renewal of embryonic stem cells. FEBS Lett. 2015, 589, 941–950. [Google Scholar] [CrossRef]
- Huppertz, I.; Perez-Perri, J.I.; Mantas, P.; Sekaran, T.; Schwarzl, T.; Russo, F.; Ferring-Appel, D.; Koskova, Z.; Dimitrova-Paternoga, L.; Kafkia, E.; et al. Riboregulation of Enolase 1 activity controls glycolysis and embryonic stem cell differentiation. Mol. Cell 2022, 82, 2666–2680.e11. [Google Scholar] [CrossRef] [PubMed]
- Fakhr, E.; Zare, F.; Azadmanesh, K.; Teimoori-Toolabi, L. LEF1 silencing sensitizes colorectal cancer cells to oxaliplatin, 5-FU, and irinotecan. Biomed. Pharmacother. 2021, 143, 112091. [Google Scholar] [CrossRef]
- Kim, S.T.; Kim, S.Y.; Lee, J.; Yun, S.H.; Kim, H.C.; Lee, W.Y.; Kim, T.W.; Hong, Y.S.; Lim, S.; Baek, J.Y.; et al. Oxaliplatin (3 months v 6 months) With 6 Months of Fluoropyrimidine as Adjuvant Therapy in Patients With Stage II/III Colon Cancer: KCSG CO09-07. J. Clin. Oncol. 2022, 40, 3868–3877. [Google Scholar] [CrossRef]
- Zamani, S.; Taslimi, R.; Sarabi, A.; Jasemi, S.; Sechi, L.A.; Feizabadi, M.M. Enterotoxigenic Bacteroides fragilis: A Possible Etiological Candidate for Bacterially-Induced Colorectal Precancerous and Cancerous Lesions. Front. Cell. Infect. Microbiol. 2020, 9, 449. [Google Scholar] [CrossRef]
- Toprak, N.U.; Yagci, A.; Gulluoglu, B.M.; Akin, M.L.; Demirkalem, P.; Celenk, T.; Soyletir, G. A possible role of Bacteroides fragilis enterotoxin in the aetiology of colorectal cancer. Clin. Microbiol. Infect. 2006, 12, 782–786. [Google Scholar] [CrossRef]
- Boleij, A.; Hechenbleikner, E.M.; Goodwin, A.C.; Badani, R.; Stein, E.M.; Lazarev, M.G.; Ellis, B.; Carroll, K.C.; Albesiano, E.; Wick, E.C.; et al. The Bacteroides fragilis Toxin Gene Is Prevalent in the Colon Mucosa of Colorectal Cancer Patients. Clin. Infect. Dis. 2014, 60, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Haghi, F.; Goli, E.; Mirzaei, B.; Zeighami, H. The association between fecal enterotoxigenic B. fragilis with colorectal cancer. BMC Cancer 2019, 19, 879. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Tang, J.; Qian, Y.; Sun, T.; Chen, H.; Chen, Z.; Sun, D.; Zhong, M.; Chen, H.; Hong, J.; et al. Long noncoding RNA BFAL1 mediates enterotoxigenic Bacteroides fragilis-related carcinogenesis in colorectal cancer via the RHEB/mTOR pathway. Cell Death Dis. 2019, 10, 675. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, X.; Song, Y.; Si, M.; Sun, Y.; Liu, X.; Cui, S.; Qu, X.; Yu, X. Exosomal miR-146a-5p and miR-155-5p promote CXCL12/CXCR7-induced metastasis of colorectal cancer by crosstalk with cancer-associated fibroblasts. Cell Death Dis. 2022, 13, 380. [Google Scholar] [CrossRef] [PubMed]
- Al-Haidari, A.; Algaber, A.; Madhi, R.; Syk, I.; Thorlacius, H. MiR-155-5p controls colon cancer cell migration via post-transcriptional regulation of Human Antigen R (HuR). Cancer Lett. 2018, 421, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Di, Z.; Di, M.; Fu, W.; Tang, Q.; Liu, Y.; Lei, P.; Gu, X.; Liu, T.; Sun, M. Integrated Analysis Identifies a Nine-microRNA Signature Biomarker for Diagnosis and Prognosis in Colorectal Cancer. Front. Genet. 2020, 11, 192. [Google Scholar] [CrossRef]
- Liu, Q.-Q.; Li, C.-M.; Fu, L.-N.; Wang, H.-L.; Tan, J.; Wang, Y.-Q.; Sun, D.-F.; Gao, Q.-Y.; Chen, Y.-X.; Fang, J.-Y. Enterotoxigenic Bacteroides fragilis induces the stemness in colorectal cancer via upregulating histone demethylase JMJD2B. Gut Microbes 2020, 12, 1788900. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, P.; Liu, Y.; Ji, T.; Liu, X.; Yao, S.; Cheng, X.; Li, Y.; Chen, L.; Xiao, Z.; et al. An epigenetic role for PRL-3 as a regulator of H3K9 methylation in colorectal cancer. Gut 2013, 62, 571–581. [Google Scholar] [CrossRef]
- Pham, T.P.; van Bergen, A.S.; Kremer, V.; Glaser, S.F.; Dimmeler, S.; Boon, R.A. LncRNA AERRIE Is Required for Sulfatase 1 Expression, but Not for Endothelial-to-Mesenchymal Transition. Int. J. Mol. Sci. 2021, 22, 8088. [Google Scholar] [CrossRef]
- Morimoto-Tomita, M.; Uchimura, K.; Werb, Z.; Hemmerich, S.; Rosen, S.D. Cloning and Characterization of Two Extracellular Heparin-degrading Endosulfatases in Mice and Humans. J. Biol. Chem. 2002, 277, 49175–49185. [Google Scholar] [CrossRef]
- Esko, J.D.; Lindahl, U. Molecular diversity of heparan sulfate. J. Clin. Investig. 2001, 108, 169–173. [Google Scholar] [CrossRef]
- Ai, X.; Do, A.-T.; Lozynska, O.; Kusche-Gullberg, M.; Lindahl, U.; Emerson, C.P., Jr. QSulf1 remodels the 6-O sulfation states of cell surface heparan sulfate proteoglycans to promote Wnt signaling. J. Cell Biol. 2003, 162, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.L.; Wolin, M.J. Pathways of acetate, propionate, and butyrate formation by the human fecal microbial flora. Appl. Environ. Microbiol. 1996, 62, 1589–1592. [Google Scholar] [CrossRef] [PubMed]
- Artis, D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat. Rev. Immunol. 2008, 8, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Chen, B.; Huo, J.; Liu, X. Therapeutic potential of NR4A1 in cancer: Focus on metabolism. Front. Oncol. 2022, 12, 972984. [Google Scholar] [CrossRef] [PubMed]
- Bain, C.C.; Mowat, A.M. Macrophages in intestinal homeostasis and inflammation. Immunol. Rev. 2014, 260, 102–117. [Google Scholar] [CrossRef]
- Li, Y.-H.; Zhang, Y.; Pan, G.; Xiang, L.-X.; Luo, D.-C.; Shao, J.-Z. Occurrences and Functions of Ly6C(hi) and Ly6C(lo) Macrophages in Health and Disease. Front. Immunol. 2022, 13, 901672. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, E.; Varol, C.; Farache, J.; Elmaliah, E.; Satpathy, A.T.; Friedlander, G.; Mack, M.; Shpigel, N.; Boneca, I.G.; Murphy, K.M.; et al. Ly6C hi monocytes in the inflamed colon give rise to proinflammatory effector cells and migratory antigen-presenting cells. Immunity 2012, 37, 1076–1090. [Google Scholar] [CrossRef]
- Wu, S.; Yuan, W.; Luo, W.; Nie, K.; Wu, X.; Meng, X.; Shen, Z.; Wang, X. MiR-126 downregulates CXCL12 expression in intestinal epithelial cells to suppress the recruitment and function of macrophages and tumorigenesis in a murine model of colitis-associated colorectal cancer. Mol. Oncol. 2022, 16, 3465–3489. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Qian, S.; Zhang, J.; Feng, J.; Luo, K.; Sun, L.; Zhao, L.; Ran, Y.; Sun, L.; Wang, J.; et al. Lipopolysaccharide promotes metastasis via acceleration of glycolysis by the nuclear factor-kappaB/snail/hexokinase3 signaling axis in colorectal cancer. Cancer Metab. 2021, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Palsson-McDermott, E.M.; O’Neill, L.A.J. The Warburg effect then and now: From cancer to inflammatory diseases. BioEssays 2013, 35, 965–973. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic regulation of gene expression by histone lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Izzo, L.T.; Wellen, K.E. Histone lactylation links metabolism and gene regulation. Nature 2019, 574, 492–493. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Akopyan, G.; Garban, H.; Bonavida, B. Transcription factor YY1: Structure, function, and therapeutic implications in cancer biology. Oncogene 2006, 25, 1125–1142. [Google Scholar] [CrossRef]
- Zhang, N.; Li, X.; Wu, C.W.; Dong, Y.; Cai, M.; Mok, M.T.S.; Wang, H.; Chen, J.; Ng, S.S.M.; Chen, M.; et al. microRNA-7 is a novel inhibitor of YY1 contributing to colorectal tumorigenesis. Oncogene 2013, 32, 5078–5088. [Google Scholar] [CrossRef]
- Shen, X.; Zhong, J.; Yu, P.; Zhao, Q.; Huang, T. YY1-regulated LINC00152 promotes triple negative breast cancer progression by affecting on stability of PTEN protein. Biochem. Biophys. Res. Commun. 2019, 509, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Yin, Y.; Zhang, J.; Zhang, B.; Bian, Z.; Quan, C.; Zhou, L.; Hu, Y.; Wang, Q.; Ni, S.; et al. MiR-139-5p inhibits migration and invasion of colorectal cancer by downregulating AMFR and NOTCH1. Protein Cell 2014, 5, 851–861. [Google Scholar] [CrossRef]
- Liu, H.; Yin, Y.; Hu, Y.; Feng, Y.; Bian, Z.; Yao, S.; Li, M.; You, Q.; Huang, Z. miR-139-5p sensitizes colorectal cancer cells to 5-fluorouracil by targeting NOTCH-1. Pathol.-Res. Pract. 2016, 212, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Bian, Z.; Zhang, J.; Li, M.; Feng, Y.; Yao, S.; Song, M.; Qi, X.; Fei, B.; Yin, Y.; Hua, D.; et al. Long non-coding RNA LINC00152 promotes cell proliferation, metastasis, and confers 5-FU resistance in colorectal cancer by inhibiting miR-139-5p. Oncogenesis 2017, 6, 395. [Google Scholar] [CrossRef]
- Kvakova, M.; Kamlarova, A.; Stofilova, J.; Benetinova, V.; Bertkova, I. Probiotics and postbiotics in colorectal cancer: Prevention and complementary therapy. World J. Gastroenterol. 2022, 28, 3370–3382. [Google Scholar] [CrossRef]
- Tripathy, A.; Dash, J.; Kancharla, S.; Kolli, P.; Mahajan, D.; Senapati, S.; Jena, M. Probiotics: A Promising Candidate for Management of Colorectal Cancer. Cancers 2021, 13, 3178. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, D.Y.; Kang, J.H.; Kim, J.H.; Jeong, J.W.; Kim, H.W.; Oh, D.H.; Yoon, S.H.; Hur, S.J. Relationship between gut microbiota and colorectal cancer: Probiotics as a potential strategy for prevention. Food Res. Int. 2022, 156, 111327. [Google Scholar] [CrossRef]
- Rad, A.H.; Aghebati-Maleki, L.; Kafil, H.S.; Abbasi, A. Molecular mechanisms of postbiotics in colorectal cancer prevention and treatment. Crit. Rev. Food Sci. Nutr. 2020, 61, 1787–1803. [Google Scholar] [CrossRef] [PubMed]
- Cullin, N.; Cullin, N.; Antunes, C.; Straussman, R.; Stein-Thoeringer, C.; Elinav, E. Microbiome and cancer. Cancer Cell 2021, 39, 1317–1341. [Google Scholar] [CrossRef] [PubMed]
- Smits, L.P.; Bouter, K.E.; de Vos, W.M.; Borody, T.J.; Nieuwdorp, M. Therapeutic Potential of Fecal Microbiota Transplantation. Gastroenterology 2013, 145, 946–953. [Google Scholar] [CrossRef]
- Khoruts, A. Can FMT Cause or Prevent CRC? Maybe, But There Is More to Consider. Gastroenterology 2021, 161, 1103–1105. [Google Scholar] [CrossRef]
- Popov, J.; Caputi, V.; Nandeesha, N.; Rodriguez, D.A.; Pai, N. Microbiota-Immune Interactions in Ulcerative Colitis and Colitis Associated Cancer and Emerging Microbiota-Based Therapies. Int. J. Mol. Sci. 2021, 22, 11365. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Hu, W.; Feng, J.; Geng, Y. Promotion or remission: A role of noncoding RNAs in colorectal cancer resistance to anti-EGFR therapy. Cell Commun. Signal. 2022, 20, 150. [Google Scholar] [CrossRef]
- Wei, H.; Mao, T.; Zhang, Q.; Ren, K.; Qi, X.; Zhang, Y.; Cao, B.; Jin, Y.; Tian, Z.; Ren, L. The Novel LncRNA WASH5P Inhibits Colorectal Cancer Carcinogenesis via Targeting AKT Signaling Pathway. Front. Oncol. 2022, 12, 923425. [Google Scholar] [CrossRef]
- Zhuang, B.; Ni, X.; Min, Z.; Wu, D.; Wang, T.; Cui, P. Long Non-Coding RNA CKMT2-AS1 Reduces the Viability of Colo-rectal Cancer Cells by Targeting AKT/mTOR Signaling Pathway. Iran. J. Public Health 2022, 51, 327–335. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, S.; Xing, J.; Jiang, Z.; Zhang, Z.; Zhang, H.; Wang, D.; Tang, D. Effects of Long Non-Coding RNAs Induced by the Gut Microbiome on Regulating the Development of Colorectal Cancer. Cancers 2022, 14, 5813. https://doi.org/10.3390/cancers14235813
Fan S, Xing J, Jiang Z, Zhang Z, Zhang H, Wang D, Tang D. Effects of Long Non-Coding RNAs Induced by the Gut Microbiome on Regulating the Development of Colorectal Cancer. Cancers. 2022; 14(23):5813. https://doi.org/10.3390/cancers14235813
Chicago/Turabian StyleFan, Shiying, Juan Xing, Zhengting Jiang, Zhilin Zhang, Huan Zhang, Daorong Wang, and Dong Tang. 2022. "Effects of Long Non-Coding RNAs Induced by the Gut Microbiome on Regulating the Development of Colorectal Cancer" Cancers 14, no. 23: 5813. https://doi.org/10.3390/cancers14235813
APA StyleFan, S., Xing, J., Jiang, Z., Zhang, Z., Zhang, H., Wang, D., & Tang, D. (2022). Effects of Long Non-Coding RNAs Induced by the Gut Microbiome on Regulating the Development of Colorectal Cancer. Cancers, 14(23), 5813. https://doi.org/10.3390/cancers14235813