

Non-Invasive and Real-Time Monitoring of the Breast Cancer Metastasis Degree via Metabolomics


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
2.2. Mouse Model and Treatment
2.3. Ultrasound Imaging and Doppler Imaging
2.4. Logistic Curve Fitting and Leave-One-Out Cross Validation
2.5. Immunohistochemistry (IHC) and Immunofluorescence (IF)
2.6. Metabolomics
2.6.1. Sample Storage and Preparation
2.6.2. Instrumentation and Conditions
2.6.3. Data Processing and Analysis
2.7. Clinical Study Participants
2.8. Method of STRING and TCGA Database
3. Results
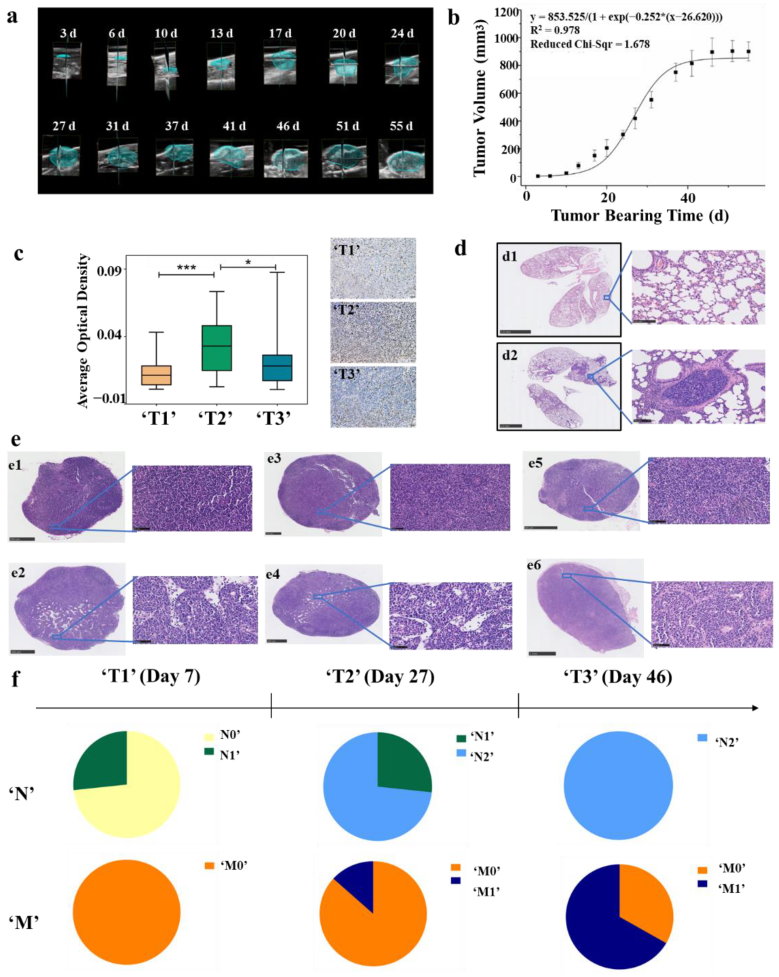
3.1. Measurement of Tumor Volume by Ultrasound Imaging and Fitting of Tumor Growth Curve
3.2. Division of ‘T’ Stage at the Animal Level According to Ki-67 Immunohistochemical and TCGA Database
3.3. Division of ‘N&M’ Stage at the Animal Level According to H&E Staining of Lymph Nodes and Lung Tissues
3.4. Metabolic Variations Associated with Tumor Metastasis
3.5. Correlations between Metabolomics Characteristics, Tumor Metastatic Status, and TME Evolution
3.6. Relationship among Intratumoral Hypoxia, TGF-β Contents in Tumor, and Metastasis-Related Metabolites Level
3.7. Level of Metastasis-Related Metabolites in Plasma of Clinical Patients with Different N Stages
3.8. Analysis of the Regulatory Relationship between Metastasis-Related Proteins and Metabolic Enzymes by Bioinformatics Analysis
4. Discussion
5. Conclusions and Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Dillekås, H.; Rogers, M.S.; Straume, O. Are 90% of deaths from cancer caused by metastases? Cancer Med. 2019, 8, 5574–5576. [Google Scholar] [CrossRef] [PubMed]
- Abu-Thuraia, A.; Goyette, M.A.; Boulais, J.; Delliaux, C.; Apcher, C.; Schott, C.; Chidiac, R.; Bagci, H.; Thibault, M.P.; Davidson, D.; et al. AXL confers cell migration and invasion by hijacking a PEAK1-regulated focal adhesion protein network. Nat. Commun. 2020, 11, 3586. [Google Scholar] [CrossRef] [PubMed]
- Prakash, V.; Carson, B.B.; Feenstra, J.M.; Dass, R.A.; Sekyrova, P.; Hoshino, A.; Petersen, J.; Guo, Y.; Parks, M.M.; Kurylo, C.M.; et al. Ribosome biogenesis during cell cycle arrest fuels EMT in development and disease. Nat. Commun. 2019, 10, 2110. [Google Scholar] [CrossRef]
- Goldhirsch, A.; Winer, E.P.; Coates, A.S.; Gelber, R.D.; Piccart-Gebhart, M.; Thürlimann, B.; Senn, H.J. Personalizing the treatment of women with early breast cancer: Highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2013. Ann. Oncol. 2013, 24, 2206–2223. [Google Scholar] [CrossRef]
- Coates, A.S.; Winer, E.P.; Goldhirsch, A.; Gelber, R.D.; Gnant, M.; Piccart-Gebhart, M.; Thürlimann, B.; Senn, H.J. Panel Members Tailoring therapies—Improving the management of early breast cancer: St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2015. Ann. Oncol. 2015, 26, 1533–1546. [Google Scholar] [CrossRef]
- Zheng, Q.H.; Yang, L.; Zeng, B.; Li, J.H.; Guo, K.X.; Liang, Y.J.; Liao, G.Q. Artificial intelligence performance in detecting tumor metastasis from medical radiology imaging: A systematic review and meta-analysis. EClinicalMedicine 2021, 31, 100669. [Google Scholar] [CrossRef]
- Takahiro, H.; Tomoko, H.; Yasuhiro, S.; Hiroki, M.; Yuichi, O.; Takashi, T.; Takumi, N.; Shingo, A.; Yuichi, M.; Tomoaki, T.; et al. Endogenous Angiogenesis Inhibitor Vasohibin1 Exhibits Broad-Spectrum Antilymphangiogenic Activity and Suppresses Lymph Node Metastasis. Am. J. Pathol. 2010, 176, 1950–1958. [Google Scholar]
- Sand, J.M.; Hafeez, B.B.; Jamal, M.S.; Witkowsky, O.; Siebers, E.M.; Fischer, J.; Verma, A.K. Plumbagin (5-hydroxy-2-methyl-1,4-naphthoquinone), isolated from Plumbago zeylanica, inhibits ultraviolet radiation-induced development of squamous cell carcinomas. Carcinogenesis 2012, 33, 184–190. [Google Scholar] [CrossRef]
- Byrd, D.R.; Brierley, J.D.; Baker, T.P.; Sullivan, D.C.; Gress, D.M. Current and future cancer staging after neoadjuvant treatment for solid tumors. CA Cancer J. Clin. 2021, 71, 140–148. [Google Scholar] [CrossRef]
- Van Roessel, S.; Kasumova, G.G.; Verheij, J.; Najarian, R.M.; Maggino, L.; de Pastena, M.; Malleo, G.; Marchegiani, G.; Salvia, R.; Ng, S.C.; et al. International Validation of the Eighth Edition of the American Joint Committee on Cancer (AJCC) TNM Staging System in Patients with Resected Pancreatic Cancer. JAMA Surg. 2019, 153, e183617. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Chavez-MacGregor, M.; Lichtensztajn, D.Y.; Yi, M.; Tadros, A.; Hortobagyi, G.N.; Giordano, S.H.; Hunt, K.K.; Mittendrof, E.A. Validation Study of the American Joint Committee on Cancer Eighth Edition Prognostic Stage Compared with the Anatomic Stage in Breast Cancer. JAMA Oncol. 2017, 4, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, B.I.; Santos, B.; Link, W.; De Sousa-Coelho, A.L. Tribbles Pseudokinases in Colorectal Cancer. Cancers 2021, 13, 2825. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Kadota, K.; Sima, C.S.; Nitadori, J.I.; Rusch, V.W.; Travis, W.D.; Sadelain, M.; Adusumilli, P.S. Clinical Impact of Immune Microenvironment in Stage I Lung Adenocarcinoma: Tumor Interleukin-12 Receptor β2 (IL-12Rβ2), IL-7R, and Stromal FoxP3/CD3 Ratio Are Independent Predictors of Recurrence. J. Clin. Oncol. 2012, 45, 2052. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.Y.; Li, W.H.; Cai, S.X.; Yang, C.S.; Liu, Y.; Lin, Z.Z. Large tumor size is a poor prognostic factor of gastric cancer with signet ring cell. Medicine 2019, 98, 40. [Google Scholar] [CrossRef]
- Boughey, J.C.; Ballman, K.V.; Hunt, K.K.; McCall, L.M.; Mittendorf, E.A.; Ahrendt, G.M.; Wilke, L.G.; Le-Petross, H.T. Axillary Ultrasound After Neoadjuvant Chemotherapy and Its Impact on Sentinel Lymph Node Surgery: Results from the American College of Surgeons Oncology Group Z1071 Trial (Alliance). J. Clin. Oncol. 2014, 57, 8401. [Google Scholar] [CrossRef]
- Hiroyuki, T.; Kimito, S.; Masafumi, K.; Yoshio, H.; Takashi, Y.; Jun, N.; Miho, Y.; Yasutaka, H.; Mari, K.; Yuji, H.; et al. Recurrence after sentinel lymph node biopsy with or without axillary lymph node dissection in patients with breast cancer. Breast Cancer 2007, 14, 16–24. [Google Scholar]
- Stokes, J.B.; Graw, K.S.; Dengel, L.T. Patients with Merkel Cell Carcinoma Tumors ≤1.0 cm in Diameter Are Unlikely to Harbor Regional Lymph Node Metastasis. J. Clin. Oncol. 2009, 27, 3772–3777. [Google Scholar] [CrossRef]
- Nikshoar, M.S.; Khayamian, M.A.; Ansaryan, S.; Sanati, H.; Gharooni, M.; Farahmand, L.; Rezakhanloo, F.; Majidzadeh-A, K.; Hoseinpour, P.; Dadgari, S.; et al. Metas-Chip precisely identifies presence of micrometastasis in live biopsy samples by label free approach. Nat. Commun. 2017, 8, 2175. [Google Scholar] [CrossRef]
- Peiris, P.M.; Deb, P.; Doolittle, E.; Doron, G.; Goldberg, A.; Govender, P.; Shah, S.; Rao, S.; Carbone, S.; Cotey, T.; et al. Vascular targeting of a gold nanoparticle to breast cancer metastasis. J. Pharm. Sci. 2015, 104, 2600–2610. [Google Scholar] [CrossRef] [PubMed]
- Hockel, A.; Dornhofer, N. The hydra phenomenon of cancer: Why tumors recur locally after microscopically complete resection. Cancer Res. 2005, 65, 2997–3002. [Google Scholar] [CrossRef] [PubMed]
- Wall, M.A.; Shaffer, T.M.; Harmsen, S.; Tschaharganeh, D.F.; Huang, C.H.; Lowe, S.W.; Drain, C.M.; Kircher, M.F. Chelator-Free Radiolabeling of SERRS Nanoparticles for Whole-Body PET and Intraoperative Raman Imaging. Theranostics 2017, 7, 3068–3077. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.H.; Zhao, Y.; Huang, W.Z.; Gao, Y.B.; Xu, W.H.; Tao, J.S.; Yang, M.; Li, L.Q.; Ping, W.; Shen, H.; et al. Non-invasive diagnosis of early-stage lung cancer using high-throughput targeted DNA methylation sequencing of circulating tumor DNA (ctDNA). Theranostics 2019, 9, 2056–2070. [Google Scholar] [CrossRef]
- Passero, V.A.; Branstetter, B.F.; Shuai, Y.; Heron, D.E.; Gibson, M.K.; Lai, S.Y.; Kim, S.W.; Grandis, J.R.; Ferris, R.L.; Johnson, J.T.; et al. Response assessment by combined PET–CT scan versus CT scan alone using RECIST in patients with locally advanced head and neck cancer treated with chemoradiotherapy. Ann. Oncol. 2010, 21, 2278–2283. [Google Scholar] [CrossRef]
- Apolo, A.B.; Riches, J.; Schoder, H.; Akin, O.; Trout, A.; Milowsky, M.I.; Bajorin, D.F. Clinical Value of Fluorine-18 2-Fluoro-2-Deoxy-D-Glucose Positron Emission Tomography/Computed Tomography in Bladder Cancer. J. Clin. Oncol. 2010, 28, 3973–3978. [Google Scholar] [CrossRef]
- Zhou, X.; Dierks, A.; Kertels, O.; Kircher, M.; Schirbel, A.; Samnick, S.; Buck, A.K.; Knorz, S.; Bockle, D.; Scheller, L.; et al. 18F-FDG, 11C-Methionine, and 68Ga-Pentixafor PET/CT in Patients with Smoldering Multiple Myeloma: Imaging Pattern and Clinical Features. Cancers 2020, 12, 2333. [Google Scholar] [CrossRef]
- Krall, J.A.; Reinhardt, F.; Mercury, O.A.; Pattabiraman, D.R.; Brooks, M.W.; Dougan, M.; Lambert, A.W.; Bierie, B.; Ploegh, H.L.; Dougan, S.K.; et al. The systemic response to surgery triggers the outgrowth of distant immune-controlled tumors in mouse models of dormancy. Sci. Transl. Med. 2018, 10, eaan3464. [Google Scholar] [CrossRef]
- Chang, K.C.; Diermeier, S.D.; Yu, A.T.; Brine, L.D.; Russo, S.; Bhatia, S.; Alsudani, H.; Kostroff, K.; Bhuiya, T.; Brogi, E.; et al. MaTAR25 lncRNA regulates the Tensin1 gene to impact breast cancer progression. Nat. Commun. 2020, 11, 6438. [Google Scholar] [CrossRef]
- Smith, S.D.; Redman, M.; Dunleavy, K. FDG PET-CT in follicular lymphoma: A case-based evidence review. Blood 2015, 125, 1078–1082. [Google Scholar] [CrossRef]
- Echeverria, G.V.; Powell, E.; Seth, S.; Ge, Z.; Carugo, A.; Bristow, C.; Peoples, M.; Robinson, F.; Qiu, H.; Shao, J.S.; et al. More High-resolution clonal mapping of multi-organ metastasis in triple negative breast cancer. Nat. Commun. 2018, 9, 5079. [Google Scholar] [CrossRef] [PubMed]
- Gertler, F.; Condeelis, J. Metastasis: Tumor cells becoming Menacing. Trends Cell Biol. 2011, 21, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, G.S.; Pastoriza, J.M.; Wang, Y.R.; Harney, A.S.; Entenberg, D.; Pignatelli, J.; Sharma, V.P.; Xue, E.A.; Cheng, E.; D’Alfonso, T.M.; et al. Neoadjuvant chemotherapy induces breast cancer metastasis through a TMEM-mediated mechanism. Sci. Transl. Med. 2017, 9, eaan0026. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.L.; Escara-Wilke, J.; Keller, J.M.; Jung, Y.; Taichman, R.; Pienta, K.J.; Keller, E.T. Primary prostate cancer educates bone stroma through exosomal pyruvate kinase M2 to promote bone metastasis. J. Exp. Med. 2020, 216, 2883–2899. [Google Scholar] [CrossRef] [PubMed]
- Elia, I.; Haigis, M.C. Metabolites and the tumour microenvironment: From cellular mechanisms to systemic metabolism. Nat. Metabol. 2021, 3, 21–32. [Google Scholar] [CrossRef]
- Ahmed, N.; Kidane, B.; Wang, L.; Nugent, Z.; Moldovan, N.; McElrea, A.; Shariati-Ievari, S.; Qing, G.F.; Tan, L.; Buduhan, G.; et al. Metabolic Changes in Early-Stage Non–Small Cell Lung Cancer Patients after Surgical Resection. Cancers 2021, 13, 3012. [Google Scholar] [CrossRef]
- Serpa, J. Metabolic remodeling as a way of adapting to tumor microenvironment (TME), a job of several holders. In Tumor Microenvironment: The Main Driver of Metabolic Adaptation; Springer: Cham, Switzerland, 2020; Volume 1219, pp. 1–34. [Google Scholar]
- Cumsille, P.; Coronel, A.; Conca, C.; Quininao, C.; Escudero, C. Proposal of a hybrid approach for tumor progression and tumor-induced angiogenesis. Theor. Biol. Med. Model 2015, 12, 13. [Google Scholar] [CrossRef][Green Version]
- Giuliano, A.E.; Connolly, J.L.; Edge, S.B.; Mittendorf, E.A.; Rugo, H.S.; Solin, L.J.; Weaver, D.L.; Winchester, D.J.; Hortobagyi, G.N. Breast Cancer-Major changes in the American Joint Committee on Cancer eighth edition cancer staging manual. CA Cancer J. Clin. 2017, 67, 291–303. [Google Scholar] [CrossRef]
- Hao, Y.; Baker, D.; ten Dijke, P. TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. Int. J. Mol. Sci. 2019, 20, 2767. [Google Scholar] [CrossRef]
- Nobre, A.R.; Entenberg, D.; Wang, Y.R.; Condeelis, J.; Aguirre-Ghiso, J.A. The Different Routes to Metastasis via Hypoxia-Regulated Programs. Trends Cell Biol. 2018, 28, 941–956. [Google Scholar] [CrossRef]
- Han, L.F.; Wang, Y.M.; Huang, X.X.; Liu, F.L.; Ma, C.Y.; Feng, F.; Zhang, J.G.; Liu, W.Y.; Qu, W.; Pang, H.; et al. Specific-oxygen-supply functionalized core-shell nanoparticles for smart mutual-promotion between photodynamic therapy and gambogic acid-induced chemotherapy. Biomaterials 2020, 257, 120228. [Google Scholar] [CrossRef] [PubMed]
- Raes, F.; Sobilo, J.; LeMee, M.; Retif, S.; Natkunarajah, S.; Lerondel, S.; Le, P.A. High Resolution Ultrasound and Photoacoustic Imaging of Orthotopic Lung Cancer in Mice: New Perspectives for Onco-Pharmacology. PLoS ONE 2016, 11, e0153532. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Sultan, L.R.; Tian, J.W.; Cary, T.W.; Sehgal, C.M. Machine learning for diagnostic ultrasound of triple-negative breast cancer. Breast Cancer Res. Treat. 2019, 173, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Zvrko, E.; Mikic, A.; Vuckovic, L. CD105 expression as a measure of microvessel density in supraglottic laryngeal squamous cell carcinoma. Eur. Arch. Oto-Rhino-Laryngol. 2009, 266, 1971–1976. [Google Scholar] [CrossRef]
- Karwicka, M.; Pucelik, B.; Gonet, M.; Elas, M.; Dąbrowski, J.M. Effects of Photodynamic Therapy with Redaporfin on Tumor Oxygenation and Blood Flow in a Lung Cancer Mouse Model. Sci. Rep. 2019, 9, 12655. [Google Scholar] [CrossRef]
- Tan, G.G.; Wang, X.; Liu, K.; Dong, X.; Liao, W.T.; Wu, H. Correlation of drug-induced and drug-related ultra-high performance liquid chromatography-mass spectrometry serum metabolomic profiles yields discovery of effective constituents of Sini decoction against myocardial ischemia in rats. Food Funct. 2018, 9, 5528–5535. [Google Scholar] [CrossRef]
- Tan, G.G.; Liu, M.; Dong, X.; Wu, S.; Fan, L.; Qiao, Y.B.; Chai, Y.F.; Wu, H. A strategy for rapid analysis of xenobiotic metabolome of Sini decoction in vivo using ultra-performance liquid chromatography-electrospray ionization quadrupole-time-of-flight mass spectrom- etry combined with pattern recognition approach. J. Pharm. Biomed. Anal. 2014, 96, 187–196. [Google Scholar] [CrossRef]
- Zhang, X.H.; Shi, X.Y.; Lu, X.; Li, Y.Q.; Zhan, C.; Akhtar, M.L.; Yang, L.J.; Bai, Y.F.; Zhao, J.X.; Wang, Y.; et al. Novel Metabolomics Serum Biomarkers for Pancreatic Ductal Adenocarcinoma by the Comparison of Pre-.; Postoperative and Normal Samples. J. Cancer 2020, 11, 4641–4651. [Google Scholar] [CrossRef]
- Hsin, C.H.; Chen, M.K.; Tang, C.H.; Lin, H.P.; Chou, M.Y.; Lin, C.W.; Yang, S.F. High level of plasma matrix metalloproteinase-11 is associated with clinicopathological characteristics in patients with oral squamous cell carcinoma. PLoS ONE 2017, 9, e113129. [Google Scholar] [CrossRef]
- Ni, J.; Cozzi, P.; Hung, T.T.; Hao, J.L.; Graham, P.; Li, Y. Monitoring Prostate Tumor Growth in an Orthotopic Mouse Model Using Three-Dimensional Ultrasound Imaging Technique. Transl. Oncol 2016, 9, 41–45. [Google Scholar] [CrossRef]
- Sun, X.M.; Kaufman, P.D. Ki-67: More than a proliferation marker. Chromosoma 2018, 127, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, R.; Sukhbaatar, A.; Sakamoto, M.; Mori, S.; Kodama, T. A model system for studying superselective radiotherapy of lymph node metastasis in mice with swollen lymph nodes. Clin. Transl. Oncol. 2020, 20, 53–57. [Google Scholar] [CrossRef]
- Thiele, W.; Rothley, M.; Dimmler, A.; Bugert, P.; Coll, C.S.; Sleeman, J.P. Platelet deficiency in Tpo−/− mice can both promote and suppress the metastasis of experimental breast tumors in an organ-specific manner. Clin. Exp. Metastasis 2018, 35, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Jobard, E.; Pontoizeau, C.; Blaise, B.J.; Bachelot, T.; Elena-Herrmann, B.; Tredan, O. A serum nuclear magnetic resonance-based metabolomic signature of advanced metastatic human breast cancer. Cancer Lett. 2014, 343, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Imamichi, Y.; Menke, A. Signaling pathways involved in collagen-induced disruption of the E-cadherin complex during epithelial-mesenchymal transition. Cells Tissues Organs 2007, 185, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Van’t Veer, L.J.; Dai, H.Y.; van de Vijver, M.J.; He, Y.D.D.; Hart, A.A.M.; Mao, M.; Peterse, H.L.; van der Kooy, K.; Marton, M.J.; Witteveen, A.T.; et al. Gene expression profiling predicts clinical outcome of breast cancer. Nature 2002, 415, 530–536. [Google Scholar] [CrossRef]
- Anggorowati, N.; Kurniasari, C.R.; Damayanti, K.; Cahyanti, T.; Widodo, I.; Ghozali, A.; Romi, M.M.; Sari, D.C.R.; Arfian, N. Histochemical and Immunohistochemical Study of α-SMA; Collagen; and PCNA in Epithelial Ovarian Neoplasm. Asian Pac. J. Cancer Prev. 2017, 18, 667–671. [Google Scholar]
- Minhajat, R.; Mori, D.; Yamasaki, F.; Sugita, Y.; Satoh, T.; Tokunaga, O. Organ-specific endoglin (CD105) expression in the angiogenesis of human cancers. Pathol. Int. 2006, 56, 717–723. [Google Scholar] [CrossRef]
- Catteau, X.; Simon, P.; Noël, J.C. Myofibroblastic stromal reaction and lymph node status in invasive breast carcinoma: Possible role of the TGF-β1/TGF-βR1 pathway. BMC Cancer 2014, 14, 499. [Google Scholar] [CrossRef]
- Yu, J.X.; Zhang, X.T.; Liao, Y.Q.; Zhang, Q.Y.; Chen, H.; Lin, M.; Kumar, S. Relationship between expression of CD105 and growth factors in malignant tumors of gastrointestinal tract and its significance. World J. Gastroenterol. 2003, 9, 2866–2869. [Google Scholar] [CrossRef]
- Verrecchia, F.; Mauviel, A. Transforming Growth Factor-β Signaling Through the Smad Pathway: Role in Extracellular Matrix Gene Expression and Regulation. J. Investig. Dermatol. 2002, 118, 211–215. [Google Scholar] [CrossRef]
- Natarajan, S.; Foreman, K.M.; Soriano, M.I.; Rossen, N.S.; Shehade, H.; Fregoso, D.R.; Eggold, J.T.; Krishnan, V.; Dorigo, O.; Krieg, A.J.; et al. Collagen Remodeling in the Hypoxic Tumor-Mesothelial Niche Promotes Ovarian Cancer Metastasis. Cancer Res. 2019, 79, 2271–2284. [Google Scholar] [CrossRef]
- Deniz, H.; Karakök, M.; Yagcı, F.; Güldür, M.E. Evaluation of relationship between HIF-1α immunoreactivity and stage, grade, angiogenic profile and proliferative index in bladder urothelial carcinomas. Int. Urol. Nephrol. 2010, 42, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Niu, W.; Dong, H.Y.; Liu, M.L.; Luo, Y.; Li, Z.C. Hypoxia induces endothelial-mesenchymal transition in pulmonary vascular remodeling. Int. J. Mol. Med. 2018, 42, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, Y.; Li, Z.; Liu, X.H. Untargeted metabolomics profiles delineate metabolic alterations in mouse plasma during lung carcinoma development using UPLCQTOF/MS in MSE mode. R. Soc. Open Sci. 2018, 5, 181143. [Google Scholar] [CrossRef] [PubMed]
- Vaisman, B.L.; Klein, H.G.; Rouis, M.; Berard, A.M.; Kindt, M.R.; Talley, G.D.; Meyn, S.M.; Hoyt, R.F.; Marcovina, S.M.; Albers, J.J.; et al. Overexpression of Human Lecithin Cholesterol Acyltransferase Leads to Hyperalphalipoproteinemia in Transgenic Mice. J. Biol. Chem. 1995, 270, 12269–12275. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, M.; Korporaal, S.J.A.; van der Sluis, R.J.; Hirsch-Reinshagen, V.; Bochem, A.E.; Wellington, C.L.; Van Berkel, T.J.C.; Kuivenhoven, J.A.; Van Eck, M. LCAT deficiency in mice is associated with a diminished adrenal glucocorticoid function. J. Lipid Res. 2013, 54, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Pentikainen, M.O.; Oorni, K.; Lassila, R.; Kovanen, P.T. The Proteoglycan Decorin Links Low Density Lipoproteins with Collagen Type I. J. Biol. Chem. 1997, 272, 7633–7638. [Google Scholar] [CrossRef]
- Rochet, M.L.B.; Angevin, L.; Bazellières, E.; Ordas, L.; Comunale, F.; Denisov, E.V.; Tashireva, L.A.; Perelmuter, V.M.; Bieche, I.; Vacher, S.; et al. P-cadherin-induced decorin secretion is required for collagen fiber alignment and directional collective cell migration. J. Cell Sci. 2019, 132, jcs233189. [Google Scholar] [CrossRef]
- Xu, L.; Ping, F.; Yin, J.H.; Xiao, X.H.; Xiang, H.D.; Ballantyne, C.M. Elevated Plasma SPARC Levels Are Associated with Insulin Resistance, Dyslipidemia, and Inflammation in Gestational Diabetes Mellitus. PLoS ONE 2013, 8, e81615. [Google Scholar] [CrossRef]
- Bradshaw, A.D. The role of SPARC in extracellular matrix assembly. J. Cell Commun. Signal 2009, 3, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.G.; Tang, L.L. SPARC in Tumor Pathophysiology and as a Potential Therapeutic Target. Curr. Pharm. Des. 2014, 20, 6182–6190. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shoshan, J.; Afek, A.; Maysel-Auslender, S.; Barzelay, A.; Rubinstein, A.; Keren, G.; George, J. HIF-1α Overexpression and Experimental Murine Atherosclerosis. Arter. Thromb. Vasc. Biol. 2009, 29, 665–670. [Google Scholar] [CrossRef]
- Xu, F.; Liu, Y.; Shi, L.; Cai, H.J.; Liu, W.; Hu, Y.J.; Li, Y.L. RGS3 inhibits TGF-β1/Smad signalling in adventitial fibroblasts. Cell Biochem. Funct. 2017, 35, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Jackstadt, R.; van Hoof, S.R.; Leach, J.D.; Cortes-Lavaud, X.; Lohuis, J.O.; Ridgway, R.A.; Wouters, V.M.; Roper, J.; Kendall, T.J.; Roxburgh, C.S.; et al. Epithelial NOTCH Signaling Rewires the Tumor Microenvironment of Colorectal Cancer to Drive Poor-Prognosis Subtypes and Metastasis. Cancer Cell 2019, 36, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Costa, N.M.M.; Fialho, A.D.V.; Proietti, C.C.; Kataoka, M.S.S.; Jaeger, R.G.; Alves-Junior, S.M.J.V.J. Role of hypoxia-related proteins in invasion of ameloblastoma cells: Crosstalk between NOTCH1; hypoxiainducible factor 1α, a disintegrin and metalloproteinase 12; and heparin-binding epidermal growth factor. Histopathology 2016, 69, 99–106. [Google Scholar] [CrossRef]
- Yeh, C.M.; Lin, C.W.; Yang, J.S.; Yang, W.E.; Su, S.C.; Yang, S.F. Melatonin inhibits TPA-induced oral cancer cell migration by suppressing matrix metalloproteinase-9 activation through the histone acetylation. Oncotarget 2016, 7, 21952–21967. [Google Scholar] [CrossRef]
- Freund, J.; Brandmaier, A.M.; Lewejohann, L.; Kirste, I.; Kritzler, M.; Kruger, A.; Sachser, N.; Lindenberger, U.; Kempermann, G. Emergence of Individuality in Genetically Identical Mice. Science 2013, 340, 756–759. [Google Scholar] [CrossRef]
- Belmaker, R.H.; Bersudsky, Y.; Agam, G. Individual differences and evidence-based psychopharmacology. BMC Med. 2012, 10, 110. [Google Scholar] [CrossRef]
- Pascual, G.; Avgustinova, A.; Mejetta, S.; Martin, M.; Castellanos, A.; Attolini, C.S.O.; Berenguer, A.; Prats, N.; Toll, A.; Hueto, J.A.; et al. Targeting metastasis-initiating cells through the fatty acid receptor CD36. Nature 2017, 541, 41. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, W.; Qian, W.; Liao, W.; Huang, X.; Xu, J.; Qu, W.; Xue, J.; Feng, F.; Liu, W.; Liu, F.; et al. Non-Invasive and Real-Time Monitoring of the Breast Cancer Metastasis Degree via Metabolomics. Cancers 2022, 14, 5589. https://doi.org/10.3390/cancers14225589
Zhu W, Qian W, Liao W, Huang X, Xu J, Qu W, Xue J, Feng F, Liu W, Liu F, et al. Non-Invasive and Real-Time Monitoring of the Breast Cancer Metastasis Degree via Metabolomics. Cancers. 2022; 14(22):5589. https://doi.org/10.3390/cancers14225589
Chicago/Turabian StyleZhu, Wanfang, Wenxin Qian, Wenting Liao, Xiaoxian Huang, Jiawen Xu, Wei Qu, Jingwei Xue, Feng Feng, Wenyuan Liu, Fulei Liu, and et al. 2022. "Non-Invasive and Real-Time Monitoring of the Breast Cancer Metastasis Degree via Metabolomics" Cancers 14, no. 22: 5589. https://doi.org/10.3390/cancers14225589
APA StyleZhu, W., Qian, W., Liao, W., Huang, X., Xu, J., Qu, W., Xue, J., Feng, F., Liu, W., Liu, F., & Han, L. (2022). Non-Invasive and Real-Time Monitoring of the Breast Cancer Metastasis Degree via Metabolomics. Cancers, 14(22), 5589. https://doi.org/10.3390/cancers14225589