Simple Summary
Breast cancer is a heterogeneous disease and treatment needs to be adapted to individual tumors. Two thirds of breast tumors may benefit from treatment with drugs targeting a specific protein, the estrogen receptor alpha, a regulator of gene expression activated by female sex hormones. However, a significant percentage of tumors will recur and progress. To help detect more accurately the expression status and activity of this protein, a gene that it upregulates, the progesterone receptor (PR), is routinely used in the clinic as an additional marker. Here, we show that PR status is an imperfect reflection of the expression and/or overall activity of estrogen receptor alpha and identify another marker that can perform this task more consistently. Overall, use of CAXII as a marker of ER+ tumors should reinforce the diagnosis of ER status and prediction of its activity and enhance the accuracy of hormonal therapy delivery.
Abstract
Estrogen receptor alpha (ERα) expression in ~2/3 breast tumors selects patients for hormonal therapies. Tumors negative for ERα but positive for the progesterone receptor (PR, encoded by PGR) have also been candidates for ER-targeting therapies, as PR expression may reflect undetected ER activity. Conversely, PR− status in ER+ tumors predicts a worse therapeutic response. Our analysis of breast tumor transcriptome datasets, however, revealed that in tumors with lower PGR expression, the clinical PR status does not correlate accurately with the expression of ESR1 or of ER target genes, including PGR itself. We identified carbonic anhydrase 12 (CA12) as an estrogen target gene better correlated with ESR1 than PGR, reflecting CA12 regulation by both ERα and the luminal factor and upstream ESR1 regulator GATA3. Immunostaining supported strong positive correlations at the protein level with ERα and GATA3 in a cohort of 118 tumors. Most ER+PR− tumors expressed CAXII at levels similar to those of ER+PR+ tumors, consistent with observations in tumor transcriptome datasets and with active estrogenic signaling in some ER+PR− breast cancer cell lines. The few ER−PR+ tumors did not express CAXII or the other luminal markers FOXA1 and GATA3. Overall, CAXII is a luminal marker that can help interpret ER status in single ER/PR positive tumors.
Keywords:
breast cancer; luminal tumors; ERα; PR; CA12/CAXII; GATA3; estrogen signaling; ER status; immunostaining 1. Introduction
Breast cancer is a complex and heterogeneous disease, as is apparent from the differential expression of therapeutic targets ERα and HER2, and by the identification of intrinsic subtypes based on whole tumor transcriptomic profiles. Different gene signatures have been proposed to identify these subtypes, resulting in various classification schemes [1,2,3,4,5]. In spite of variations between classifiers, luminal subtypes essentially correspond to ERα-positive (ER+) tumors and display the highest prevalence, representing together more than 70% of all malignancies. ERα, a ligand-dependent transcription factor and member of the nuclear receptor family, is the main therapeutic target in these tumors as it drives cell proliferation and survival. Targeted treatments include anti-estrogens (AEs), which modulate ERα activity and/or accelerate its turnover, and aromatase inhibitors (AIs), which prevent the synthesis of estrogens [6]. Both ERα and PR, encoded by PGR, a target gene of ERα, are used as markers of tumors that may benefit from hormonal therapies [7,8,9,10], their expression at the protein level being routinely detected by immunohistochemistry (IHC). ERα and PR protein levels in ER+ tumors are often stronger and more homogeneous than in adjacent normal tissue, where they are heterogeneous within luminal cells of the mammary ducts and alveoli [11]. However, ER+ tumors can also display intra-tumor heterogeneity for ERα and PR expression levels. The threshold for positivity for both markers is nuclear expression in at least 1% of the cells according to the 2010 American Society of Clinical Oncology (ASCO) guidelines [12], but can be as high as 10% in clinical practice [13]. ER− tumors include both tumors over-expressing the ERBB2 gene, which codes for the ERBB2/HER2 membrane receptor, and those negative for HER2 as well as ERα and PR (triple negative, TN).
The causes of intra-tumoral heterogeneity of ER expression are currently still speculative and could correspond to the co-existence of different tumor cell states or of different biological subtypes, the latter potentially resulting in intrinsic resistance to hormonal treatment. Low ERα expression (1–10% cells) has indeed been associated with increased rates of relapse and progression on hormonal therapies [14,15,16,17], and may result in an unstable ER status during tumor progression [14]. The St Gallen international expert consensus on the primary therapy of early breast cancer 2015 describes ER values between 1 and 9% as equivocal [18]. A 2020 update from ASCO recommends use of a new reporting category for these tumors, ER Low Positive, with additional steps to confirm and interpret the results [19]. Although detection of PR as well as ER reinforces recommendation of hormonal therapies in double hormone receptor (HR) positive tumors (55–67% of all tumors), especially at the low end of the ER expression spectrum, it also yields single positive tumors, mainly ER+PR− tumors (7–16% of all tumors) and a small fraction of ER−PR+ tumors (1–4%) [9,20]. The ER−PR+ tumor category has been proposed to reflect staining artifacts, warranting reanalysis of the ER− diagnosis [21,22]. However, others have found that ER−PR+ tumors are a rare but distinct entity [23,24,25]. These tumors were reported to have survival rates and incidence of BRCA mutations similar to ER−PR− tumors, with which they share phenotypic traits [10,26,27,28]. Nevertheless, ER−PR+ tumors have been traditionally considered for hormonal therapies and even included in the Luminal A group in the absence of ERBB2 amplification in surrogate tumor subtype classifications [10]. On the other hand, ER+PR− tumors have 5-yr survival rates intermediate between those of ER+PR+ and ER−PR− tumors, conferring a prognostic value to PR expression [8,9,20]. The contribution of PR expression to prognosis has been attributed to its role as a surrogate marker of ERα expression/activity [13,29,30] and also to the direct contribution of PR isoforms to tumor cell biology and progression [31,32,33]. ER+PR− tumors, nevertheless, are still candidates for hormonal therapies, in association or not with chemotherapy, as response rates are lower but not null in this tumor group [34].
Apart from PGR, other estrogen target genes have been considered for their association with ER status and/or prediction of response to hormonal therapies, including pS2/TFF1 [35] and Growth Regulation by Estrogen in Breast cancer 1 (GREB1) [36]. Contrary to PGR, neither are included in the PAM50 classifier [3], although both are part of the 256 genes CIT breast cancer classifier [5]. They also play roles in ER+ tumorigenesis that may confound their significance as ER surrogate markers. The TFF1 protein was reported to stimulate breast cancer cell migration [37] and to exert apoptosis-protecting effects in doxorubicin-treated cells [38]. Gene deletion in mice reduced breast tumorigenesis [39]. Nevertheless, high intratumoral expression of TFF1 was found to have a low but significant predictive value for response to tamoxifen [40]. GREB1 has been reported to act as an ERα coactivator, but also to modify it by glycosylation, to increase its stability and decrease response to tamoxifen [41,42]. It may also have ER-independent roles as a proliferation regulator in breast cancer [43]. In ovarian cancer, it acts as a tumor promoter, promoting cell proliferation, migration and mesenchymal morphology [44]. It was also identified as a target of the Wnt/β-catenin pathway required for hepatoblastoma progression [45].
Another estrogen target gene known to be enriched in ER+ tumors is CA12, encoding the membrane protein carbonic anhydrase 12 (CAXII) [46,47], one of the 15 members of the carbonic anhydrase family catalyzing the hydration of carbon dioxide [48]. This family of metalloenzymes includes a total of four classes depending on their subcellular location [48]. CAXII belongs to the class of proteins found at the plasma membrane [49]. Regulation of CA12 by estrogens has been observed in MCF-7 and T-47D breast cancer cells, and ChIP experiments have indicated binding of ERα to an enhancer at −6 kb [46]. Although CA12 is regulated in breast cancer cells by other transcription factors (TFs) such as AP2γ and by hypoxia, albeit much less than its paralog CA9 [50,51], we show that its RNA expression levels are amongst the most highly correlated with those of the ESR1 gene in several transcriptome datasets. We confirmed that ERα is a major regulator of CA12 expression, acting via two ERE-containing enhancers, and that the luminal factor GATA3, itself an upstream regulator of the ESR1 gene [52], also induces CA12 expression in ER+ tumors. We further validated the strong association of CAXII and ERα positivity at the protein level in a 118-tumor cohort. Comparison with PR status indicated that CAXII was broadly expressed in ER+PR− tumors but was absent from ER−PR+ tumors. We propose that CAXII may usefully complement PR for the determination of ERα expression and activity.
2. Materials and Methods
2.1. Cell Culture Conditions
Breast cell lines were purchased from the American Type Culture Collection (ATCC) and maintained in a humidified 37 °C, 5% CO2 incubator. MCF-7, SKBR-3 and MDA-MB-453 breast cancer cells were cultured in Dulbecco’s Minimal Eagle’s Medium (DMEM; Wisent Inc., Saint-Jean Baptiste, QC, Canada) supplemented with 10% fetal bovine serum (FBS; Sigma, St. Louis, MO, USA) and 1% penicillin/streptomycin (P/S, Wisent Inc., Saint-Jean Baptiste, QC, Canada). MDA-MB-231 were maintained in DMEM with 5% FBS and 1% P/S. T-47D, ZR-75-1 and HCC-70 cells were maintained in RPMI 1640 (Wisent Inc., Saint-Jean Baptiste, QC, Canada) with 10% FBS, 1% P/S, 10 mM HEPES pH 7.0 (BioBasic, Markham, ON, Canada), and 1% sodium pyruvate (Wisent Inc., Saint-Jean Baptiste, QC, Canada). MCF-10A cells were maintained in Dulbecco’s Modified Eagle’s Medium F-12 (DMEM F-12; Wisent Inc., Saint-Jean Baptiste, QC, Canada) without calcium chloride with 10% FBS and 1% P/S. For estrogenic treatments, cells were cultured in hormone-depleted media (without phenol red and supplemented with charcoal-dextran treated FBS (FBST)).
2.2. RNA Extraction, Reverse Transcription and Real-Time Quantitative PCR
Total RNA was prepared using 1 mL of QIAzol Lysis Reagent (QIAGEN, Hilden, Germany), 0.2 mL of chloroform (Sigma-Aldrich, St. Louis, MO, USA) and 0.5 mL isopropanol 99.5% (Fisher, Hampton, NH, USA) per cell pellet. RNA aliquots (1 µg) were reverse-transcribed using the RevertAid H first minus strand cDNA synthesis kit (Thermo Fisher Scientific, Waltham, MA, USA) with oligo(dT)18 primers. Expression levels of target genes were assessed by real-time quantitative PCR (qPCR) using the Universal ProbeLibrary system (Roche, Basel, Switzerland) and the ViiA 7 Real-Time PCR System (ThermoFisher Scientific, Waltham, MA, USA) using YWHAZ and RPLP0 as housekeeping genes. RT-qPCR primer sequences and UPL probe numbers are listed in Supplementary Table S1.
2.3. Protein Extraction and Western Blotting
Whole cell lysates were prepared using RIPA lysis buffer (Tris-HCl pH 7.5 50 mM, NaCl 150 mM, Triton X-100 1%, SDS 0.1%, sodium deoxycholate 0.5% and freshly added protease inhibitors (PMSF 10 mM and leupeptin, pepstatin, and aprotinin, 1 µg/mL)). Extracts were homogenized for 10 min before sonication (4 °C, 10 min; Bioruptor, Diagenode, Denville, NJ, USA) and quantified using a Lowry assay (BioRad, Hercules, CA, USA). Equal amounts of proteins (20–40 µg) were lysed in Laemmli blue 1X and electrophoresed on an 8% SDS-polyacrylamide gel at 80V for 2 h. Proteins were transferred on a PVDF membrane using a Trans-Blot Turbo Transfer System (BioRad, Hercules, CA, USA). Membranes were blocked using PBS-T (PBS 1X, Tween 20 0.05%) with 5% milk for 1h and blotted with rabbit or mouse antibodies directed against CAXII, ERα, FOXA1, GATA3, β-actin or lamin B1 overnight (see Supplementary Table S2), and then incubated with horseradish peroxidase–conjugated secondary anti-mouse or anti-rabbit IgG (Cedarlane, Burlington, Canada) for 45 min. Immunodetection was performed using the Clarity Western ECL substrate (BioRad, Hercules, CA, USA) for enhanced chemiluminescence. Original blots see supplementary File S1.
2.4. siRNA Transfection
Cells were maintained in hormone-depleted media for three days before transfection. Cells were seeded and transfected when attached. A SMARTpool of four siRNAs was used against CA12 and TFAP2C and two different ON-TARGETplus siRNAs against ESR1, FOXA1 or GATA3 (Dharmacon, Lafayette, CO, USA). One ON-TARGETplus Non-Targeting siRNA (“si-Control”, Dharmacon, Lafayette, CO, USA) was used as a negative control. Transfection was performed with each siRNA (40 nM) using the SilentFect reagent (BioRad, Hercules, CA, USA) for a total duration of 72 h. Cells were then treated or not with E2 (25 nM) or with vehicle (EtOH 0.024%), 24 h before cell collection for subsequent RNA and protein extractions. Sequences of all siRNAs are provided in Supplementary Table S3.
2.5. Chromatin Immunoprecipitation (ChIP)
Cells maintained in a hormone-depleted medium for three days were either treated or not with E2 (25 nM) for 1 h before crosslinking at room temperature by addition of formaldehyde (1%) for 10 min. Crosslinking was stopped by adding glycine (0.125 M) for 5 min and washing cells twice with ice-cold PBS 1X. Collected cells were lysed on ice using lysis buffer (Tris-HCl pH 8.0 10 mM, EDTA 10 mM, EGTA 0.5 mM, Triton X-100 0.25%, and protease inhibitors) for 5 min. After centrifugation, cell pellets were washed with a second lysis buffer (Tris pH 8.0 10 mM, NaCl 200 mM, EDTA 1 mM, EGTA 0.5 mM and protease inhibitors), incubated for 30 min and centrifugated. Cells resuspended in sonication buffer (Tris pH 8.0 10 mM, NaCl 140 mM, EDTA 1 mM, EGTA 0.5 mM, SDS 0.5%, Triton X-100 0.5%, sodium deoxycholate 0.05%, and protease inhibitors) were sonicated using a Bioruptor (Diagenode, Denville, NJ, USA; maximum intensity, 30 s intervals between pulses). IP was performed on sonicated chromatin prepared from three million cells by addition of 3 µg of each antibody (see Supplementary Table S2) with a 1:1 mix of Dynabeads A and G (Invitrogen, Waltham, MA, USA) in ChIP dilution buffer (Tris pH 8.0 10 mM, NaCl 150 mM, EDTA 2 mM, Triton X-100 1%). Mixes were then incubated on a rotor O/N at 4 °C to capture antibody-protein-DNA complexes. Beads were washed with buffer (Tris pH 8.0 20 mM, EDTA 2 mM, Triton X-100 1% and SDS 0.1%) containing decreasing concentrations of NaCl (from 500 to 50 mM). Input DNA and IP samples were decrosslinked (NaHCO3 10 mM, SDS 1%) O/N at 65 °C. Eluates were subsequently incubated with RNase A (BioBasic, Markham, ON, Canada) for 30 min at 65 °C and Proteinase K (ThermoScientific, Waltham, MA, USA) for 1 h at 65 °C. Finally, DNA fragments were purified on EZ-10 columns (BioBasic, Markham, ON, Canada). ChIP results are shown as percentage of input relative to IgG (% input with antibody/% input with IgG). ChIP qPCR primer sequences and the respective TaqMan probe numbers are provided in Supplementary Table S4.
2.6. Tissue Microarrays and Immunohistochemitry
Tissue micro-arrays (TMAs) were prepared from cores (1 mm) extracted from each of 118 formalin-fixed and paraffin-embedded (FFPE) breast tumor tissues as previously described [53]. These samples were obtained from patients with primary breast tumors who underwent surgery at the Hôtel Dieu Hospital and at the Centre Hospitalier de l’Université de Montréal (CHUM) between 2001 and 2018. The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the CHUM ethics committee (SL 05-019). Tumor characteristics extracted from the pathological reports include ER, PR and HER2 status as well as histological classification and grades (Supplementary Table S5).
Sections (4 µm) were prepared from TMAs or FFPE normal mammary gland tissues. Deparaffinization and antigen retrieval were performed using a Discovery XT automatic stainer (Ventana Medical Systems, Oro Valley, AZ, USA) and incubated with primary antibodies directed against CAXII, ERα, FOXA1 or GATA3 (see Supplementary Table S6 for antibodies and dilutions). Single staining was performed using Bond polymer DAB refine kits (#DS9800, Leica Biosystems, Wetzlar, Germany) on a Bond RX stainer (Leica Biosystems, Buffalo Grove, IL, USA). Dual staining was performed by staining CAXII as above, and then using the Green Chromogen (DC9913, Leica Biosystems, Wetzlar, Germany) in conjunction with BOND Polymer Refine HRP PLEX Detection (#DS9914) for ERα, FOXA1 or GATA3. The sections were then stained with Gill hematoxylin to visualize nuclei. Stained tissue sections were scanned using the C9600 NanoZoomer System (Hamamatsu Corporation, Bridgewater, NJ, USA). The NDP Scan software (version 2.2.9; Hamamatsu Corporation, Bridgewater, NJ, USA) was used to extract all images. The stained tissue sections were scored using the QuPath software (version 0.3.0, https://github.com/qupath/qupath/releases, accessed on 26 September 2022). Training was performed on several cores for cell identification based on hematoxylin intensity staining and nuclei shapes, and for scoring of each marker based on a range of expression levels (from null to high expression) in tumor cells. QuPath provides the proportion of positive cells (0 to 5 scale) and staining intensities (0 to 3 scale), with a maximum total score of 8 [54]. HRP staining was analyzed in the nuclear or membrane compartment for ERα, FOXA1 and GATA3 or CAXII, respectively. Score cut-offs were set based on the trough in the bi-modal histogram representation for each protein (scores ≥ 4 for ERα, ≥ 3 for CAXII). IHC and co-IHC staining conditions for CAXII, ERα, GATA3 and FOXA1 and controls are listed in Supplementary Tables S6 and S7.
3. Results
3.1. The Clinical PR Status Is Not an Accurate Predictor of ESR1 Expression Levels or of ERα Activity
To examine whether the PR clinical status reflects ER expression and/or activity, we generated a pair-wise scatterplot representation of ESR1 and PGR expression levels in an RNA-seq breast tumor transcriptome dataset from the Cancer Genome Atlas (TCGA, 754 tumors) using MiSTIC, a visualization platform for gene–gene correlation studies and enrichment analysis [55]. This analysis revealed that, although PGR expression levels were well correlated with those of ESR1 (Pearson correlation 0.68), tumors with low PGR expression levels have a broad range of ESR1 expression levels. Indeed, selecting a PGR expression level cut-off leading to the exclusion of ~90% of ESR1low tumors (~22.5% of all tumors; Figure 1A, left box) excluded many tumors with high ESR1 expression levels (12.2% of all tumors, ~16.3% of ESR1high tumors; Figure 1A, right box). Note that the ESR1 expression levels correlated well with ER status, with only 5.4% of tumors having a discrepant status (Figure 1A, ER+ tumors in blue, ER− tumors in red). On the other hand, PGR expression levels displayed more discrepancies with the PR clinical status (13.4% overall; Figure 1B, PR+ tumors in orange, PR− tumors in cyan). This may suggest either post-transcriptional regulation of PGR expression, or variability in calling PR status, especially for tumors with low to intermediate RNA levels (Figure 1B, boxes).
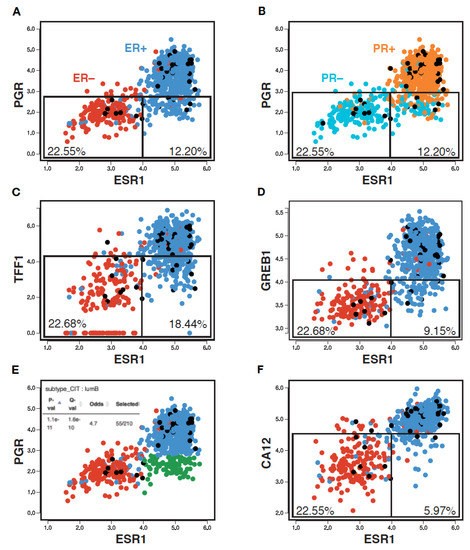
Figure 1.
PR status imperfectly reflects ESR1 expression, ER status and activity. Pair-wise scatterplots for the expression of ESR1 vs PGR (A,B,E), TFF1 (C), GREB1 (D) and CA12 (F) generated using MiSTIC [55]. Boxes identify 90% of ESR1low tumors, and the portion of ESR1high tumors identified using the same threshold of positivity for different ER target genes. Percentages of total tumors are shown. Enrichment of ESR1highPGRlow tumors (highlighted in green) in the CIT luminal B tumor subtype is shown in the inset of panel (E).
Other well-characterized ER target genes such as TFF1 or GREB1 (Figure 1C,D) displayed similar correlation coefficients with ESR1 at the RNA expression level (0.70 and 0.68, resp., Figure S1). The overlap in expression levels between ESR1high and ESR1low tumors was higher for TFF1 (exclusion of 18.4% of total tumors, i.e., 24.6% of ESR1high tumors for an expression cut-off excluding 90% of ESR1low tumors), but lower for GREB1 (exclusion of 9.2% total tumors, i.e., 12.3% of ESR1high tumors). Notably, a significant fraction of the ESR1highPGRlow tumors (green dots in Figure S2A) express high levels of TFF1 (~75.3%) and/or GREB1 (~77.4) (purple dots in Figure S2B,C). Conversely, most of the rare ESR1lowPGRhigh tumors (18 tumors, ~2.4%) express low levels of these other ER target genes (14/18 TFF1low, 15/18 GREB1low, not shown).
Together, these results indicate that PR scores do not appear to provide highly accurate information on either ESR1 expression levels or ERα activity. Rather, PGR expression may reflect the activity of other regulatory pathways with prognostic value, as the ESR1highPGRlow tumor group was enriched in LumB tumors (Figure 1E, Q-value 1.6 × 10−10; Figure 1).
3.2. CA12 mRNA Levels Correlate with Those of Luminal Transcription Factor Genes ESR1, GATA3 and FOXA1 in Breast Tumor Transcriptome Datasets
Gene correlation analysis in the same TCGA breast tumor dataset (754 tumors) using MiSTIC revealed a cluster of genes with highly correlated expression biased for luminal tumors (Figure 2A). The strong correlation of cluster genes is not the result of a copy number variation (CNV) event, as most genes are located on different chromosomes. This cluster includes six luminal transcription factor genes: ESR1, FOXA1, GATA3, AR, SPDEF and XBP1 (in orange, Figure 2A), suggesting that clustering reflects transcriptional regulation. Apart from genes located proximal to and likely co-regulated with ESR1 (CCDC170, C6orf211/ARMT1 and RMND1), known estrogen targets are also found in this cluster, including TFF1, AGR3 and CA12 (in red, Figure 2A). CA12 encodes a carbonic anhydrase whose expression is regulated by estrogens in MCF-7 and T-47D breast cancer cell lines [46]. Interestingly, ESR1 expression levels were more highly correlated with those of CA12 (Pearson correlation coefficient of 0.81) than of PGR, TFF1 and GREB1. CA12 expression was also correlated, albeit to a slightly lower level, with luminal transcription factors GATA3 and FOXA1 (correlation coefficients of 0.77 and 0.76, respectively) (Figure 2B).
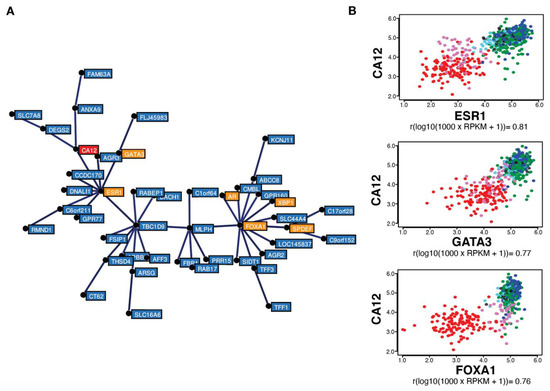
Figure 2.
CA12 mRNA levels are highly correlated with those of ESR1, FOXA1 and GATA3 in breast tumors. (A) Minimum spanning tree representing a cluster of genes expressed preferentially in luminal and molecular apocrine breast tumors, identified by gene correlation analysis from a TCGA breast cancer transcriptomic dataset comprising 754 tumors using MiSTIC software [55]. Genes encoding transcription factors (ESR1, FOXA1, GATA3, SPDEF, XBP1 and AR) are highlighted in orange, whereas CA12 is indicated in red. (B) Pair-wise correlation scatterplots for CA12 and ESR1 (top), GATA3 (middle) or FOXA1 (bottom) expression levels in the same cohort, with CIT tumor subtypes highlighted by different colors. Luminal A tumors appear in dark blue, luminal B in green, luminal C in light blue, molecular apocrine in magenta and basal-like in red. Normal-like or non-classified tumors are shown in black. Pearson correlation coefficients are indicated at the bottom of each pair-wise correlation scatterplot.
High correlation between CA12 and ESR1 expression is reproducible in the TCGA Firehose Legacy dataset comprising 960 breast tumor transcriptomes (Pearson correlation coefficient of 0.80, Figure S3A) and in the 1904 breast cancer samples from the METABRIC dataset (Pearson correlation coefficient of 0.79, Figure S3B). Outlier tumors in this correlation were those expressing low levels of CA12 mRNA in spite of high ESR1 levels, which were identified as a small fraction of luminal B tumors, and tumors expressing higher CA12 mRNA levels than predicted from ESR1 expression, representing a fraction of molecular apocrine (mApo) tumors in the CIT classification [5] (Figure 2B). mApo tumors are ER– and express high levels of the androgen receptor (AR) and FOXA1. They include most ER–HER2+ and some TN tumors [56,57,58,59,60]. Interestingly, mApo tumors with high CA12 levels also had high levels of GATA3, whereas other mApo tumors have GATA3 levels comparable to those of basal-like tumors (Figure 2B). Basal-like tumors expressed low levels, whereas luminal tumors expressed high levels of both ESR1 and CA12 (Figure 2A,B). Thus, setting a threshold of CA12 RNA positivity excluding 90% of ESR1low tumors led to exclusion of a much lower number of ESR1high tumors compared to PGR (about half, Figure 1F).
3.3. CAXII Is Detected Predominantly in ER+ Breast Cancer Cells
Using a transcriptome dataset for 51 breast cancer cell lines including luminal, ERBB2-amplified, basal and claudin-low lines [61], we determined whether expression of CA12 is also associated with that of ESR1 (Figure 3A) and how it compares with PGR expression (Figure 3B). CA12 expression at the RNA level was consistently high in all luminal cell lines. Basal tumors and claudin-low cell lines expressed lower levels of CA12. ERBB2-amp lines displayed a range of ESR1 expression and CA12 levels, with some overlap in CA12 expression between ESR1low and ESR1high lines. PGR RNA levels were much lower in comparison, being absent in most claudin-low, basal and ESR1low ERBB2-amp cell lines but also in some ESR1high lines. Notably, as observed in tumors, several ESR1highPGRnull lines expressed high CA12 levels. These lines, classified as luminal or ERBB2-amplified lines, include estrogen-responsive ZR75-30 and 600MPE lines [62], confirming that PR− status does not necessarily reflect lack of estrogen response. Levels of PGR were also relatively low in estrogen-responsive MCF7 and MDA-MB-134VI [63].
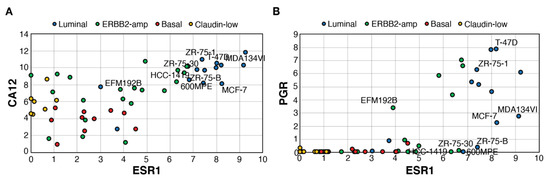
Figure 3.
ESR1high cell lines express high levels of CA12, but variable levels of PGR. Pair-wise scatterplots for the expression of ESR1 vs those of CA12 (A) or PGR (B). Values are Log2(Rpkm+1). RNA levels and cell line subtyping are from [61]. Selected cell lines are labeled.
Next, we compared levels of CA12 RNA (RT-qPCR, Figure S4A) and CAXII protein (western analysis, Figure S4B) in cell lines corresponding to different breast cancer subtypes using a polyclonal antibody previously validated for CAXII detection in western and immunohistochemistry analyses [51,64]. MCF-7, T-47D and ZR-75-1 are luminal breast cancer lines, SKBR-3 and MDA-MB-453 are ER− but express FOXA1 and AR and are representative of the mApo subtype, with ERBB2-amplification and high expression in SKBR-3 and MDA-MB-453, respectively. HCC70 and MDA-MB-231 belong respectively to the basal-like and claudin-low subtypes of triple-negative breast cancer cell lines [5,65]. In addition, the MCF-10A cell line is immortalized but non-tumorigenic. ESR1 RNA and ERα protein levels were, as expected, high in MCF-7, T-47D and ZR-75-1 cells and low in all other cell lines (Figure S4A,B). Strong RNA and protein expression of the CA12 gene was detected in T-47D and ZR-75-1 cells and in the mApo SKBR-3 cell line. Weaker expression was observed in MCF-7 and in immortalized MCF-10A. Expression was low to undetectable in mApo MDA-MB-453, basal HCC70 cells and claudin-low MDA-MB-231 (Figure S4A,B). Western analysis further revealed the existence of multiple bands detected by the CAXII antibody. Different CA12 transcripts are responsible for the translation of two main CAXII isoforms, including a longer form at 39 kDa (Gen-Bank accession # NM_001218.5) and a 37 kDa truncated isoform, resulting from translation of an RNA transcript lacking exon 9 (Gen-Bank accession # NM_206925.3). However, multiple glycosylation events result in different CAXII forms, including, for the longer isoform, a fully glycosylated form at 43 kDa [66]. Alternative splicing and variants presenting amino acid substitution at the level of glycosylation sites, such as p.His121Gln and p.Glu143Lys [66] could be responsible for the different ratios of CAXII isoforms. Migration patterns indicate that only ER+ breast cancer cells express the high-molecular weight CAXII form (upper band, Figure S4B).
Globally, the CAXII protein expression patterns mirrored those at the RNA level, confirming the specificity of the antibody and suggesting that post-transcriptional regulation does not play a major role in the control of CAXII protein expression in breast cancer cell lines.
3.4. CA12 Is Regulated by Luminal Transcription Factors ERα and GATA3 in ER+ Cell Lines
ENCODE ChIP-Seq datasets visualized on the UCSC Genome Browser identified CA12 regulatory regions bound by luminal transcription factors with associated binding motifs (Figure 4A). TFs ERα, FOXA1 and GATA3 bound three potential CA12 enhancers, with predicted estrogen response elements (ERE) motifs in enhancers 1 and 2 and predicted GATA3 binding motifs in enhancer 2. Regulation of CA12 by estrogens in MCF-7 and T-47D cells was previously described and binding to enhancer 1 was confirmed by ChIP-qPCR [46]. Accordingly, we observed time-dependent induction of CA12 upon treatment of MCF-7 and T-47D cells with 25 nM 17β-estradiol (E2), with maximal expression at 16 h, but did not observe induction of CA12 by E2 in ZR-75-1 cells within 24 h (Figure 4B). These results were reproduced at the protein level, CAXII expression in these cells being maximal at 16 h in MCF-7 and at 24 h in T-47D cells but remaining mostly stable in ZR-75-1 cells (Figure 4C). Further, ChIP-qPCR confirmed estradiol-induced recruitment of ERα on enhancers 1 and 2, compared to control IgG in MCF-7 cells (Figure 4D) and to gene desert regions (not shown).
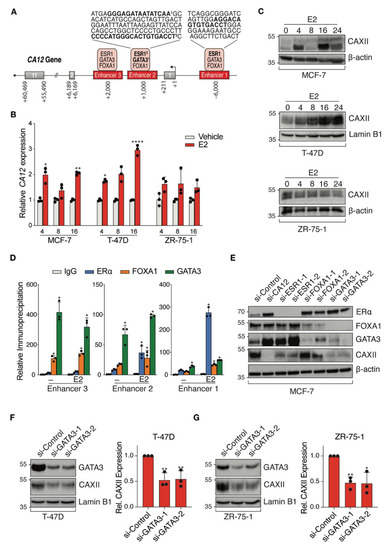
Figure 4.
ERα and GATA3 positively regulate CA12 expression in luminal breast cancer cell lines. (A) Schematic CA12 gene organization with exons (grey boxes) and putative enhancers (red boxes). Predicted binding motifs for ERα and GATA3 are highlighted in bold in sequences for enhancers 1 and 2. (B) MCF-7, T-47D and ZR-75-1 cell lines were cultured in hormone-depleted medium for five days and then treated with E2 (25 nM). CA12 mRNA levels were measured by RT-qPCR at 4, 8 and 16 h. RPLP0 and YWHAZ were used as housekeeping genes for normalization. Values are presented as ratios over the expression in vehicle-treated samples for each time point. Asterisks indicate statistically significant regulations (Average of n = 3; *, p ≤ 0.05; **, p ≤ 0.01; ****, p ≤ 0.0001; Student’s unpaired t-test). (C) CAXII protein levels from MCF-7, T-47D and ZR-75-1 cells were analyzed at 0, 4, 8, 16 and 24 h after treatment with E2 by Western blotting (n = 3, a representative blot is shown). β-actin or Lamin B1 housekeeping proteins were used as loading controls. (D) MCF-7 cells were cultured in hormone-depleted medium for three days and then treated with E2 (25 nM) for 1 h before fixation and collection. Binding of ERα, FOXA1 and GATA3 to CA12 enhancers was examined by ChIP-qPCR. Relative immunoprecipitation levels (ratios to IgG in vehicle-treated cells) are shown. The assay was performed twice with similar results. Asterisks indicate statistically significant binding compared to IgG in vehicle-treated cells for each enhancer from one experiment performed in triplicates (*, p ≤ 0.05; one-way ANOVA test, Dunnett’s multiple comparisons test). (E) MCF-7 cells were cultured in hormone-depleted medium for three days and transfected with a SMARTpool of siRNAs targeting CA12 and two different siRNAs targeting ESR1, FOXA1 or GATA3. Cells were then collected two days after transfection. Protein levels of CAXII, ERα, FOXA1 and GATA3 were analyzed by Western blotting (n = 2). β-actin was used as a loading control. (F,G) T-47D and ZR-75-1 cells were cultured in hormone-depleted medium for three days and transfected with two different siRNAs targeting GATA3. Cells were then collected two days after transfection. Protein levels of CAXII and GATA3 were analyzed by Western blotting in T-47D (F) and ZR-75-1 (G) cells; CAXII levels were quantified for both cell lines using ImageJ (average of n = 3 for quantification, one representative blot shown; *, p ≤ 0.05; **, p ≤ 0.01; Student’s unpaired t-test). Lamin B1 was used as a loading control.
We also confirmed GATA3 recruitment as well as that of FOXA1 on all three enhancers in MCF-7 cells (Figure 4D). To further explore regulation of CA12 by luminal transcription factors, we targeted ESR1, FOXA1 and GATA3 by siRNAs (2 siRNAs per gene) as well as CA12 (siRNA pool) in MCF-7 cells cultured in a hormone-depleted medium (Figure 4E). These experiments revealed that GATA3 in addition to ERα regulates CAXII protein expression. As GATA3 acts in a positive regulatory loop with ERα [52], GATA3 could be involved in the regulation of CA12 indirectly. However, ERα levels were not significantly depleted by siRNAs against GATA3 (or conversely) in MCF-7 cells under our experimental conditions. To support the evidence of regulation of CA12 by GATA3, we transfected siRNAs against GATA3 in two other cell lines, T-47D (Figure 4F) and ZR-75-1 (Figure 4G). The results showed a significant decreased expression of CAXII, supported by a quantification of the protein levels using Image J.
Taken together, these results indicate that CA12 is a luminal gene that is regulated by ERα and GATA3, although potential regulation by FOXA1 in some settings cannot be excluded.
3.5. CAXII Expression Is Increased with Lack of Polarity in Invasive ER+ Tumors
CA12 is expressed physiologically in epithelial cells in the pancreas, kidney and colon as well as in the mammary gland [48,67,68]. We probed CAXII levels in breast tumors as well as in normal tissue by immunohistochemistry (IHC). Different controls were used to validate and optimize CAXII staining of tissue sections, including the pancreas, kidney and colon as positive expression controls and lymph node, liver and spleen for negative controls (Supplementary Figure S5). CAXII levels are higher in breast ductal carcinoma in situ compared to normal lobules and ducts [51]. In pulmonary adenocarcinomas, increased CAXII levels correlate with tumor grade and aggressiveness [69]. Accordingly, in normal mammary gland and normal tumor-adjacent mammary tissues, CAXII levels, detected by IHC using the same antibody as above, were low and heterogeneous (Figure 5A), similar to those of ERα. Conversely, in breast tumors, CAXII levels were high and homogeneous. The osmotic exchange of bicarbonates by CAXII occurs at the basolateral level and not in the lumen in renal and pancreatic epithelial cells [67,68]. CAXII basolateral polarity was observable in columnar cell lesions, in which bilayer integrity is preserved (Figure 5B) but was lost in intra-ductal proliferating cells in ductal carcinoma in situ (DCIS) (Figure 5C) and in cells migrating into the stromal compartment in invasive ductal carcinoma (IDC) (Figure 5D).
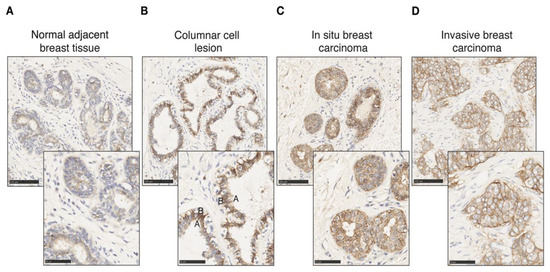
Figure 5.
CAXII expression is increased in tumoral epithelial cells with loss of polarity. CAXII staining (in brown) of normal adjacent mammary tissue (A), columnar cell lesion (B), in situ breast ductal carcinoma (C) and invasive breast ductal carcinoma (D) at 20× and 40× magnification. The apical (A) and basal (B) poles of a luminal cell are labeled.
3.6. Levels of CAXII Correlate with Those of ERα, GATA3 and FOXA1 in Breast Tumor Arrays
Immunohistochemistry analysis of ERα, GATA3, FOXA1 and CAXII was then carried out in consecutive slices of seven tissue micro-arrays (TMAs) displaying 118 tumors represented in triplicates. Scores for nuclear (for ERα, GATA3 and FOXA1) or membrane (for CAXII) staining in individual tumors were computed using QuPath [54] after training for identification of epithelial, stromal and immune compartments and validation by a pathologist (Figure S6A,B). Cut-offs for positivity were set at 4 for ERα and at 3 for CAXII, based on their bimodal distributions (Figure S7A,B). QuPath scores for ERα were largely concordant with the ER clinical status. Linear regression analysis of score distributions for CAXII and ERα, GATA3 or FOXA1 revealed a high determination coefficient (R2 ~0.77, ~0.61 and ~0.51, resp., Figure 6A–C), supporting at the protein level the correlations observed at the RNA level between CA12 and ESR1, GATA3 or FOXA1 expression. Accordingly, co-staining of CAXII (membrane localization, brown) and ERα, GATA3 or FOXA1 (nuclear localization, green) in tumors further supported their co-expression (Figure 6A–C).
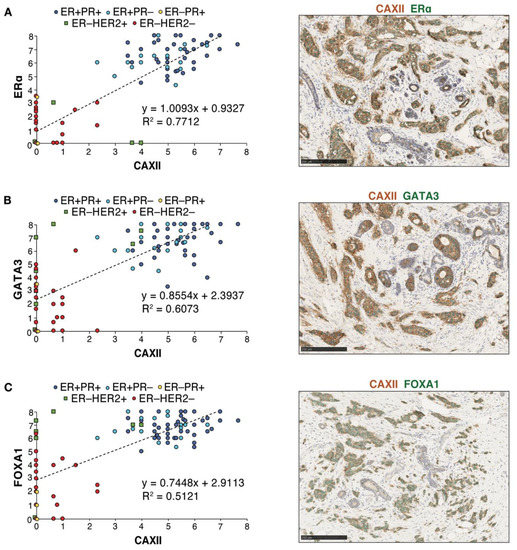
Figure 6.
Protein expression levels of CAXII and of the luminal transcription factors ERα, FOXA1 and GATA3 are correlated in breast tumors. Correlations between CAXII and ERα (A), GATA3 (B) or FOXA1 (C) expression scores are shown for all tumors in the TMAs. Scores were computed by QuPath using membrane (CAXII) and nuclear signal intensities (ERα, GATA3, FOXA1). ER+PR+ tumors are highlighted in dark blue circles, ER+PR− tumors in light blue circles, ER−PR+ tumors in yellow diamonds, ER−HER2+ tumors in green squares and ER−HER2− tumors in red circles. Representative co-staining for CAXII (brown) and each of these luminal transcription factors (green) is also shown.
3.7. CAXII Is Mainly Expressed in ER+ Breast Tumors
CAXII score distributions within each clinical tumor subtype indicate that in 72 ER+ tumors, 71 (98.6%) were positive for CAXII. In these tumors, the 55 ER+PR+ and the 17 ER+PR− have significantly higher scores for CAXII expression compared to ER−HER2+ and TN clinical tumors (Figure 7A). However, no significant difference in CAXII scores was observed between ER+PR+ and ER+PR− tumors. The only ER+ tumor negative for CAXII (score 2.3) is also negative for PR, suggesting low or absent ERα activity. Conversely, 7 out 9 ER−HER2+ tumors and all 34 TN tumors (ER−PR−HER2−) are negative for CAXII expression (Figure 7A). The 2 ER−HER2+ tumors positive for CAXII express intermediate CAXII levels (scores at 3.67 and 4.0). These tumors also express FOXA1 and GATA3 [70], but are negative for PR. The 3 tumors classified as ER−PR+ in the clinic (ER scores below 1%, PR scores between 1–10%) were negative for both ERα and CAXII in our analysis (yellow diamonds in Figure 6A, yellow dots in the ER−HER2− group in Figure 7A). These tumors were also all negative for expression of GATA3 and FOXA1 (Figure 6B,C). Two out of 3 tumors expressed high levels of the basal marker FOXC1 (not shown), suggesting a dominant basal-like phenotype. The third one was null for all subtype markers assessed, suggestive of a different TN subtype. Representative staining results for ER and CAXII expression are shown from co-staining corresponding to the two most frequently observed phenotypes, ER+CAXII+ (71 tumors) and ER−CAXII− (37 tumors), and the two rarer phenotypes ER−CAXII+ (2 tumors, both HER2+) and ER+CAXII− (1 tumor) (Figure 7B).
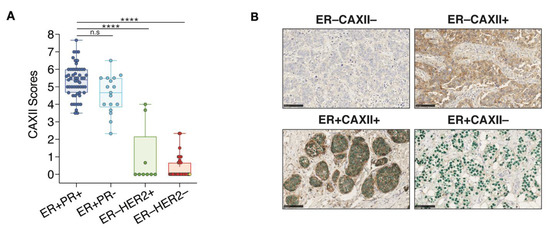
Figure 7.
CAXII expression discriminates ER+ from ER− breast tumors. (A) Boxplot representing CAXII score distribution according to ER, PR and HER2 status (7 arrays, n = 118; ns, p > 0.05; ****, p ≤ 0.0001; Dunn’s multiple comparisons post-hoc test performed after Kruskall-Wallis test (p < 0.0001)). ER−PR+ tumors are highlighted in yellow in the ER−HER2− group. (B) Representative cores for the four observed phenotypes of CAXII and ER status after co-staining with CAXII (brown) and ERα (green). Scores were computed by QuPath using membrane (CAXII) and nuclear (ERα) signal intensities.
4. Discussion
ER status remains the main diagnostic factor for selection of tumors for hormonal therapies. Lack of PR expression in ER+ tumors is a bad prognosis factor and may be used as an indication for additional chemotherapy, whereas its expression in the absence of ER may prompt re-examination of ER staining and inclusion of controls without clear guidelines as to whether or not hormonal therapies may be of benefit. Additional markers that could reinforce the diagnosis of ER expression and activity are thus desirable. In this study, we identified CA12 as one of the ERα target genes whose RNA expression is most correlated with that of ESR1, GATA3 and FOXA1 in breast cancer datasets. This high degree of correlation at the RNA level stems both from the very high correlation of ESR1 RNA and protein levels and the regulation of CA12 by both ERα and by GATA3, which may cooperate by synergizing for recruitment of coactivators or of the basal transcription machinery, and via cross-regulation [52,71,72,73,74]. Our TMA study further confirmed the value of the CAXII protein as a marker of luminal tumors and of ERα activity in IHC assays. In particular, CAXII IHC status correlated better with ER clinical status than PR clinical status.
Our dataset contained three ER− tumors with a positive but low PR score (1–10%), which would have led to the diagnosis of two tumors as PR− if a 10% threshold was applied. CAXII was absent in all three ER−PR+ tumors, compatible with lack of ER expression. Low FOXA1 levels in all three tumors and high FOXC1 levels in two of them further suggest TN phenotypes. Thus, staining with CAXII in addition to ER and PR, and possibly in combination with subtype markers such as FOXA1 and FOXC1, may spare some patients with ER−PR+ tumors futile long-term hormonal therapy treatments, and emphasize the need for TNBC-targeting therapies and genetic testing for BRCA mutations.
A large fraction of ER+ tumors (23.6%, 17 out of 72) in our dataset had a negative PR status, in spite of the low threshold for PR positivity, and most of these (15/17) expressed ER in more than 10% of the cells. Strikingly, 16/17 ER+PR− tumors were positive for CAXII, suggesting a luminal phenotype (supported by positive scores for FOXA1 and GATA3), and, potentially, active ER signaling. Observations at the RNA level in the TCGA dataset also support active ER signaling in an important fraction of ER+PR− tumors. Indeed, the majority (~75%) of ESR1highPGRlow tumors (12.2% of all tumors) express high levels of another ER target gene (GREB1, TFF1 or CA12), suggesting that few tumors express fully inactive ER proteins. Lack of PR positivity, but not of other ERα target genes, may result from differential activity of ERα on its different target genes, possibly due to variable levels of ER cofactors. Other explanations for the lack of PR expression in a subset of ER+ tumors include copy number loss or CpG island methylation, independent from ER regulation. Irrespective of the mechanism involved, loss of PR expression likely contributes to bad prognosis for reasons other than loss of ER signaling, including its own action as a transcriptional regulator and/or as a modulator of ER action [33]. In this respect, it would be important to determine whether the higher degree of discrepancy between PGR RNA expression and PR clinical status compared to ESR1/ERα results from scoring variability, affecting the accuracy of PR status as a prognostic biomarker, or from potential post-transcriptional regulation of PR levels, the significant entity for progesterone signaling.
In the TCGA dataset, RNA expression levels of CA12 comparable to those in some ER+ tumors were observed in a subset of molecular apocrine tumors, which are ER−, but positive for AR and FOXA1 expression, and often ERBB2-amplified [56,57,58,59,60]. Two ER−HER2+FOXA1+ tumors indeed had moderate expression levels of CAXII in our study cohort. Although CA12 expression is highly correlated with ER status in breast tumors at the RNA and protein levels, genes are rarely regulated by a single transcription factor. Other TFs such as GATA3 or AP2γ [70], or environmental factors such as hypoxia [49,50] may contribute to CAXII positivity in the absence of ER expression. This may result in one or several CA12 enhancers remaining active in some tumors. Indeed, DNA methylation patterns on CA12 enhancers 1 (at –6 kb) and 2 (+2 kb) were intermediate in two mApo cell lines (SKBR-3 and MDA-MB-453), with partial CpG methylation on one of the two regions, vs complete methylation in basal-like and claudin-low cell lines (HCC-1954, Hs578T and MDA-MB-231), and lack of methylation in luminal cell lines (MCF-7 and T-47D) [70]. Tumors expressing CA12 in spite of lack of ERα are, however, likely to be negative for other markers of ER signaling such as PR, as was observed here for our two ER−CAXII+ tumors.
An advantage of CAXII as a biomarker is its membrane localization, enabling combining it usefully with nuclear ER detection in co-staining. In addition, the intra-tumoral heterogeneity of the ERα protein observed in several mammary tumors suggests that its expression is differentially regulated from one cell to another. Whether intra-tumor heterogeneity of ERα protein levels reflects the co-existence of different subtypes within the same tumor remains unclear. A surrogate marker of ESR1 expression expressed on cell plasma membranes such as CAXII may enable the isolation of ER+ cells in heterogeneous tissues by fluorescence-activated cell sorting (FACS), and ultimately characterization of sorted tumor subpopulations via functional genomics approaches to determine genetic/epigenetic differences between these populations. CAXII may also be used in combination with other markers such as EpCAM and/or CD49f [75] to enhance purification of normal luminal mammary epithelial cells.
Limitations of this study include the small number of lobular carcinoma (nine samples) and absence of less frequent luminal subtypes such as tubular or mucinous tumors. It will be interesting in the future to explore whether the high correlation between CA12 and ESR1 remains valid in these subtypes. In addition, only three ER−PR+ cases were present in our cohort. This specific subgroup should be further studied in the future in larger cohorts, especially in retrospective studies in order to assess the usefulness of CAXII in distinguishing tumors that may benefit from hormonal therapies from those that do not. Similarly, further studies are needed to determine the frequency of ER+ tumors that are negative for both PR and CAXII (one tumor in our cohort, positive for FOXA1 and GATA3), and whether these tumors are also negative for other ER target genes and are unresponsive to hormonal therapies.
5. Conclusions
Altogether, CAXII is a luminal marker that would likely prove useful in conjunction with ERα and PR to identify tumors that may benefit from hormonal therapies, with or without adjuvant chemotherapy. CAXII may in addition prove useful to identify and sort heterogeneous cell populations in tumors and normal tissues.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cancers14215453/s1, Figure S1: Correlations between mRNA levels of ESR1 and of several estrogen target genes in breast cancer. Figure S2: ESR1highPGRlow status does not imply lack of ER signaling. Figure S3: CA12 is mainly expressed in luminal tumors. Figure S4. CA12 is highly expressed in luminal and mApo breast cancer cell lines. Figure S5. CAXII staining optimization. Figure S6. Tumor cell detection and scoring by QuPath in breast tumor sections. Figure S7. Bimodal distribution of CAXII and ER QuPath scores. Table S1. RT-qPCR primer sequences. Table S2. Immunoblotting and immunoprecipitation conditions. Table S3. SiRNA oligonucleotide sequences. Table S4. ChIP-qPCR primer sequences. Table S5. Clinicopathological characteristics of the 118 breast tumors. Table S6. Antibodies and IHC staining conditions. Table S7. Co-IHC staining conditions. File S1: Original blots.
Author Contributions
Conceptualization, L.P. and S.M.; methodology, L.P., F.G., N.K.T. and S.M.; validation, L.P., F.G., N.K.T., L.G. and S.M.; formal analysis, L.P., F.G. and S.M.; writing—original draft preparation, L.P.; writing—review and editing, S.M.; visualization, L.P.; and F.G.; supervision, S.M.; project administration, S.M.; funding acquisition, S.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Canadian Institutes for Health Research, grants # PJT153178 and PJT156199 to S.M.; by studentships from the Molecular Biology program and Institute for Research in Immunology and Cancer to L.P.; and by a studentship from the Faculté des Etudes Supérieures et Post-doctorales of Université de Montréal to F.G.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) Centre Hospitalier de l’Université de Montréal (protocol SL 05-019).
Informed Consent Statement
All patients were informed before surgery that their surgical specimens might be used for research purposes.
Data Availability Statement
Public datasets analyzed for figure generation are referenced in the figure legends (cBioportal: https://www.cbioportal.org; MiSTIC: http://mistic.iric.ca. Data generated during this study is available upon request.
Acknowledgments
The authors are grateful to William Shuford Sly and Abdul Waheed for generously sharing the CAXII antibody. We thank Layane Duarte, Fatima Zohra Khadri, Sarah Pasquin and Hend Al-Bizri for TMA generation and tumor characterization, and the IRIC Histology platform for slide preparation and IHC staining of FFPE blocks.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Fan, C.; Oh, D.S.; Marron, J.S.; He, X.; Qaqish, B.F.; Livasy, C.; Carey, L.A.; Reynolds, E.; Dressler, L.; et al. The molecular portraits of breast tumors are conserved across microarray platforms. BMC Genom. 2006, 7, 96. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.S.; Mullins, M.; Cheang, M.C.; Leung, S.; Voduc, D.; Vickery, T.; Davies, S.; Fauron, C.; He, X.; Hu, Z.; et al. Supervised risk predictor of breast cancer based on intrinsic subtypes. J. Clin. Oncol. 2009, 27, 1160–1167. [Google Scholar] [CrossRef]
- Jonsson, G.; Staaf, J.; Vallon-Christersson, J.; Ringner, M.; Holm, K.; Hegardt, C.; Gunnarsson, H.; Fagerholm, R.; Strand, C.; Agnarsson, B.A.; et al. Genomic subtypes of breast cancer identified by array-comparative genomic hybridization display distinct molecular and clinical characteristics. Breast Cancer Res. 2010, 12, R42. [Google Scholar] [CrossRef] [PubMed]
- Guedj, M.; Marisa, L.; de Reynies, A.; Orsetti, B.; Schiappa, R.; Bibeau, F.; MacGrogan, G.; Lerebours, F.; Finetti, P.; Longy, M.; et al. A refined molecular taxonomy of breast cancer. Oncogene 2012, 31, 1196–1206. [Google Scholar] [CrossRef]
- Harbeck, N.; Penault-Llorca, F.; Cortes, J.; Gnant, M.; Houssami, N.; Poortmans, P.; Ruddy, K.; Tsang, J.; Cardoso, F. Breast cancer. Nat. Rev. Dis. Prim. 2019, 5, 66. [Google Scholar] [CrossRef]
- Schultz, J.R.; Petz, L.N.; Nardulli, A.M. Estrogen receptor alpha and Sp1 regulate progesterone receptor gene expression. Mol. Cell. Endocrinol. 2003, 201, 165–175. [Google Scholar] [CrossRef]
- Bardou, V.J.; Arpino, G.; Elledge, R.M.; Osborne, C.K.; Clark, G.M. Progesterone receptor status significantly improves outcome prediction over estrogen receptor status alone for adjuvant endocrine therapy in two large breast cancer databases. J. Clin. Oncol. 2003, 21, 1973–1979. [Google Scholar] [CrossRef]
- Rakha, E.A.; El-Sayed, M.E.; Green, A.R.; Paish, E.C.; Powe, D.G.; Gee, J.; Nicholson, R.I.; Lee, A.H.; Robertson, J.F.; Ellis, I.O. Biologic and clinical characteristics of breast cancer with single hormone receptor positive phenotype. J. Clin. Oncol. 2007, 25, 4772–4778. [Google Scholar] [CrossRef]
- Onitilo, A.A.; Engel, J.M.; Greenlee, R.T.; Mukesh, B.N. Breast cancer subtypes based on ER/PR and Her2 expression: Comparison of clinicopathologic features and survival. Clin. Med. Res. 2009, 7, 4–13. [Google Scholar] [CrossRef]
- Petersen, O.W.; Hoyer, P.E.; van Deurs, B. Frequency and distribution of estrogen receptor-positive cells in normal, nonlactating human breast tissue. Cancer Res. 1987, 47, 5748–5751. [Google Scholar] [PubMed]
- Hammond, M.E.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer (unabridged version). Arch. Pathol. Lab. Med. 2010, 134, e48–e72. [Google Scholar] [CrossRef] [PubMed]
- Goldhirsch, A.; Glick, J.H.; Gelber, R.D.; Coates, A.S.; Thurlimann, B.; Senn, H.J.; Panel, M. Meeting highlights: International expert consensus on the primary therapy of early breast cancer 2005. Ann. Oncol. 2005, 16, 1569–1583. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, L.S.; Karlsson, E.; Wilking, U.M.; Johansson, U.; Hartman, J.; Lidbrink, E.K.; Hatschek, T.; Skoog, L.; Bergh, J. Clinically used breast cancer markers such as estrogen receptor, progesterone receptor, and human epidermal growth factor receptor 2 are unstable throughout tumor progression. J. Clin. Oncol. 2012, 30, 2601–2608. [Google Scholar] [CrossRef]
- Lindstrom, L.S.; Yau, C.; Czene, K.; Thompson, C.K.; Hoadley, K.A.; Van’t Veer, L.J.; Balassanian, R.; Bishop, J.W.; Carpenter, P.M.; Chen, Y.Y.; et al. Intratumor Heterogeneity of the Estrogen Receptor and the Long-term Risk of Fatal Breast Cancer. J. Natl. Cancer Inst. 2018, 110, 726–733. [Google Scholar] [CrossRef]
- Yi, M.; Huo, L.; Koenig, K.B.; Mittendorf, E.A.; Meric-Bernstam, F.; Kuerer, H.M.; Bedrosian, I.; Buzdar, A.U.; Symmans, W.F.; Crow, J.R.; et al. Which threshold for ER positivity? A retrospective study based on 9639 patients. Ann. Oncol. 2014, 25, 1004–1011. [Google Scholar] [CrossRef]
- Fujii, T.; Kogawa, T.; Dong, W.; Sahin, A.A.; Moulder, S.; Litton, J.K.; Tripathy, D.; Iwamoto, T.; Hunt, K.K.; Pusztai, L.; et al. Revisiting the definition of estrogen receptor positivity in HER2-negative primary breast cancer. Ann. Oncol. 2017, 28, 2420–2428. [Google Scholar] [CrossRef]
- Coates, A.S.; Winer, E.P.; Goldhirsch, A.; Gelber, R.D.; Gnant, M.; Piccart-Gebhart, M.; Thurlimann, B.; Senn, H.J.; Panel, M. Tailoring therapies--improving the management of early breast cancer: St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2015. Ann. Oncol. 2015, 26, 1533–1546. [Google Scholar] [CrossRef]
- Allison, K.H.; Hammond, M.E.H.; Dowsett, M.; McKernin, S.E.; Carey, L.A.; Fitzgibbons, P.L.; Hayes, D.F.; Lakhani, S.R.; Chavez-MacGregor, M.; Perlmutter, J.; et al. Estrogen and Progesterone Receptor Testing in Breast Cancer: ASCO/CAP Guideline Update. J. Clin. Oncol. 2020, 38, 1346–1366. [Google Scholar] [CrossRef]
- Bae, S.Y.; Kim, S.; Lee, J.H.; Lee, H.C.; Lee, S.K.; Kil, W.H.; Kim, S.W.; Lee, J.E.; Nam, S.J. Poor prognosis of single hormone receptor- positive breast cancer: Similar outcome as triple-negative breast cancer. BMC Cancer 2015, 15, 138. [Google Scholar] [CrossRef]
- Hefti, M.M.; Hu, R.; Knoblauch, N.W.; Collins, L.C.; Haibe-Kains, B.; Tamimi, R.M.; Beck, A.H. Estrogen receptor negative/progesterone receptor positive breast cancer is not a reproducible subtype. Breast Cancer Res. 2013, 15, R68. [Google Scholar] [CrossRef]
- Foley, N.M.; Coll, J.M.; Lowery, A.J.; Hynes, S.O.; Kerin, M.J.; Sheehan, M.; Brodie, C.; Sweeney, K.J. Re-Appraisal of Estrogen Receptor Negative/Progesterone Receptor Positive (ER−/PR+) Breast Cancer Phenotype: True Subtype or Technical Artefact? Pathol. Oncol. Res. 2018, 24, 881–884. [Google Scholar] [CrossRef]
- Ng, C.H.; Pathy, N.B.; Taib, N.A.; Mun, K.S.; Rhodes, A.; Yip, C.H. The estrogen receptor negative-progesterone receptor positive breast carcinoma is a biological entity and not a technical artifact. Asian Pac. J. Cancer Prev. 2012, 13, 1111–1113. [Google Scholar] [CrossRef] [PubMed]
- De Maeyer, L.; Van Limbergen, E.; De Nys, K.; Moerman, P.; Pochet, N.; Hendrickx, W.; Wildiers, H.; Paridaens, R.; Smeets, A.; Christiaens, M.R.; et al. Does estrogen receptor negative/progesterone receptor positive breast carcinoma exist? J. Clin. Oncol. 2008, 26, 335–336. [Google Scholar] [CrossRef]
- Beltjens, F.; Molly, D.; Bertaut, A.; Richard, C.; Desmoulins, I.; Loustalot, C.; Charon-Barra, C.; Courcet, E.; Bergeron, A.; Ladoire, S.; et al. ER−/PR+ breast cancer: A distinct entity, which is morphologically and molecularly close to triple-negative breast cancer. Int. J. Cancer 2021, 149, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Sanford, R.A.; Song, J.; Gutierrez-Barrera, A.M.; Profato, J.; Woodson, A.; Litton, J.K.; Bedrosian, I.; Albarracin, C.T.; Valero, V.; Arun, B. High incidence of germline BRCA mutation in patients with ER low-positive/PR low-positive/HER-2 neu negative tumors. Cancer 2015, 121, 3422–3427. [Google Scholar] [CrossRef] [PubMed]
- Keshgegian, A.A.; Cnaan, A. Estrogen receptor-negative, progesterone receptor-positive breast carcinoma: Poor clinical outcome. Arch. Pathol. Lab. Med. 1996, 120, 970–973. [Google Scholar]
- Dowsett, M.; Houghton, J.; Iden, C.; Salter, J.; Farndon, J.; A’Hern, R.; Sainsbury, R.; Baum, M. Benefit from adjuvant tamoxifen therapy in primary breast cancer patients according oestrogen receptor, progesterone receptor, EGF receptor and HER2 status. Ann. Oncol. 2006, 17, 818–826. [Google Scholar] [CrossRef]
- Horwitz, K.B.; McGuire, W.L. Predicting response to endocrine therapy in human breast cancer: A hypothesis. Science 1975, 189, 726–727. [Google Scholar] [CrossRef]
- McGuire, W.L.; Horwitz, K.B.; Pearson, O.H.; Segaloff, A. Current status of estrogen and progesterone receptors in breast cancer. Cancer 1977, 39, 2934–2947. [Google Scholar] [CrossRef]
- Creighton, C.J.; Kent Osborne, C.; van de Vijver, M.J.; Foekens, J.A.; Klijn, J.G.; Horlings, H.M.; Nuyten, D.; Wang, Y.; Zhang, Y.; Chamness, G.C.; et al. Molecular profiles of progesterone receptor loss in human breast tumors. Breast Cancer Res. Treat. 2009, 114, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Daniel, A.R.; Hagan, C.R.; Lange, C.A. Progesterone receptor action: Defining a role in breast cancer. Expert Rev. Endocrinol. Metab. 2011, 6, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, H.; Russell, I.A.; Stark, R.; Rueda, O.M.; Hickey, T.E.; Tarulli, G.A.; Serandour, A.A.; Birrell, S.N.; Bruna, A.; Saadi, A.; et al. Progesterone receptor modulates ERalpha action in breast cancer. Nature 2015, 523, 313–317. [Google Scholar] [CrossRef]
- Ravdin, P.M.; Green, S.; Dorr, T.M.; McGuire, W.L.; Fabian, C.; Pugh, R.P.; Carter, R.D.; Rivkin, S.E.; Borst, J.R.; Belt, R.J.; et al. Prognostic significance of progesterone receptor levels in estrogen receptor-positive patients with metastatic breast cancer treated with tamoxifen: Results of a prospective Southwest Oncology Group study. J. Clin. Oncol. 1992, 10, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Henry, J.A.; Piggott, N.H.; Mallick, U.K.; Nicholson, S.; Farndon, J.R.; Westley, B.R.; May, F.E. pNR-2/pS2 immunohistochemical staining in breast cancer: Correlation with prognostic factors and endocrine response. Br. J. Cancer 1991, 63, 615–622. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hnatyszyn, H.J.; Liu, M.; Hilger, A.; Herbert, L.; Gomez-Fernandez, C.R.; Jorda, M.; Thomas, D.; Rae, J.M.; El-Ashry, D.; Lippman, M.E. Correlation of GREB1 mRNA with protein expression in breast cancer: Validation of a novel GREB1 monoclonal antibody. Breast Cancer Res. Treat. 2010, 122, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Prest, S.J.; May, F.E.; Westley, B.R. The estrogen-regulated protein, TFF1, stimulates migration of human breast cancer cells. FASEB J. 2002, 16, 592–594. [Google Scholar] [CrossRef]
- Pelden, S.; Insawang, T.; Thuwajit, C.; Thuwajit, P. The trefoil factor 1 (TFF1) protein involved in doxorubicininduced apoptosis resistance is upregulated by estrogen in breast cancer cells. Oncol. Rep. 2013, 30, 1518–1526. [Google Scholar] [CrossRef]
- Buache, E.; Etique, N.; Alpy, F.; Stoll, I.; Muckensturm, M.; Reina-San-Martin, B.; Chenard, M.P.; Tomasetto, C.; Rio, M.C. Deficiency in trefoil factor 1 (TFF1) increases tumorigenicity of human breast cancer cells and mammary tumor development in TFF1-knockout mice. Oncogene 2011, 30, 3261–3273. [Google Scholar] [CrossRef]
- Corte, M.D.; Tamargo, F.; Alvarez, A.; Rodriguez, J.C.; Vazquez, J.; Sanchez, R.; Lamelas, M.L.; Gonzalez, L.O.; Allende, M.T.; Garcia-Muniz, J.L.; et al. Cytosolic levels of TFF1/pS2 in breast cancer: Their relationship with clinical-pathological parameters and their prognostic significance. Breast Cancer Res. Treat. 2006, 96, 63–72. [Google Scholar] [CrossRef]
- Mohammed, H.; D’Santos, C.; Serandour, A.A.; Ali, H.R.; Brown, G.D.; Atkins, A.; Rueda, O.M.; Holmes, K.A.; Theodorou, V.; Robinson, J.L.; et al. Endogenous purification reveals GREB1 as a key estrogen receptor regulatory factor. Cell Rep. 2013, 3, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.M.; Huynh, V.T.; Neja, S.A.; Liu, C.Y.; Raju, A.; Tan, K.; Tan, N.S.; Gunaratne, J.; Bi, X.; Iyer, L.M.; et al. GREB1: An evolutionarily conserved protein with a glycosyltransferase domain links ERalpha glycosylation and stability to cancer. Sci. Adv. 2021, 7, eabe2470. [Google Scholar] [CrossRef]
- Haines, C.N.; Braunreiter, K.M.; Mo, X.M.; Burd, C.J. GREB1 isoforms regulate proliferation independent of ERalpha co-regulator activities in breast cancer. Endocr. Relat. Cancers 2018, 25, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Hodgkinson, K.; Forrest, L.A.; Vuong, N.; Garson, K.; Djordjevic, B.; Vanderhyden, B.C. GREB1 is an estrogen receptor-regulated tumour promoter that is frequently expressed in ovarian cancer. Oncogene 2018, 37, 5873–5886. [Google Scholar] [CrossRef]
- Matsumoto, S.; Yamamichi, T.; Shinzawa, K.; Kasahara, Y.; Nojima, S.; Kodama, T.; Obika, S.; Takehara, T.; Morii, E.; Okuyama, H.; et al. GREB1 induced by Wnt signaling promotes development of hepatoblastoma by suppressing TGFbeta signaling. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Barnett, D.H.; Sheng, S.; Charn, T.H.; Waheed, A.; Sly, W.S.; Lin, C.Y.; Liu, E.T.; Katzenellenbogen, B.S. Estrogen receptor regulation of carbonic anhydrase XII through a distal enhancer in breast cancer. Cancer Res. 2008, 68, 3505–3515. [Google Scholar] [CrossRef]
- Li, Y.; Lei, B.; Zou, J.; Wang, W.; Chen, A.; Zhang, J.; Fu, Y.; Li, Z. High expression of carbonic anhydrase 12 (CA12) is associated with good prognosis in breast cancer. Neoplasma 2019, 66, 420–426. [Google Scholar] [CrossRef]
- Waheed, A.; Sly, W.S. Carbonic anhydrase XII functions in health and disease. Gene 2017, 623, 33–40. [Google Scholar] [CrossRef]
- Chiche, J.; Ilc, K.; Laferriere, J.; Trottier, E.; Dayan, F.; Mazure, N.M.; Brahimi-Horn, M.C.; Pouyssegur, J. Hypoxia-inducible carbonic anhydrase IX and XII promote tumor cell growth by counteracting acidosis through the regulation of the intracellular pH. Cancer Res. 2009, 69, 358–368. [Google Scholar] [CrossRef]
- Wykoff, C.C.; Beasley, N.J.; Watson, P.H.; Turner, K.J.; Pastorek, J.; Sibtain, A.; Wilson, G.D.; Turley, H.; Talks, K.L.; Maxwell, P.H.; et al. Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res. 2000, 60, 7075–7083. [Google Scholar]
- Wykoff, C.C.; Beasley, N.; Watson, P.H.; Campo, L.; Chia, S.K.; English, R.; Pastorek, J.; Sly, W.S.; Ratcliffe, P.; Harris, A.L. Expression of the hypoxia-inducible and tumor-associated carbonic anhydrases in ductal carcinoma in situ of the breast. Am. J. Pathol. 2001, 158, 1011–1019. [Google Scholar] [CrossRef]
- Eeckhoute, J.; Keeton, E.K.; Lupien, M.; Krum, S.A.; Carroll, J.S.; Brown, M. Positive cross-regulatory loop ties GATA-3 to estrogen receptor alpha expression in breast cancer. Cancer Res. 2007, 67, 6477–6483. [Google Scholar] [CrossRef] [PubMed]
- Khadri, F.Z.; Issac, M.S.M.; Gaboury, L.A. Impact of Epithelial-Mesenchymal Transition on the Immune Landscape in Breast Cancer. Cancers 2021, 13, 5099. [Google Scholar] [CrossRef] [PubMed]
- Bankhead, P.; Loughrey, M.B.; Fernandez, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef]
- Lemieux, S.; Sargeant, T.; Laperriere, D.; Ismail, H.; Boucher, G.; Rozendaal, M.; Lavallee, V.P.; Ashton-Beaucage, D.; Wilhelm, B.; Hebert, J.; et al. MiSTIC, an integrated platform for the analysis of heterogeneity in large tumour transcriptome datasets. Nucleic Acids Res. 2017, 45, e122. [Google Scholar] [CrossRef]
- Farmer, P.; Bonnefoi, H.; Becette, V.; Tubiana-Hulin, M.; Fumoleau, P.; Larsimont, D.; Macgrogan, G.; Bergh, J.; Cameron, D.; Goldstein, D.; et al. Identification of molecular apocrine breast tumours by microarray analysis. Oncogene 2005, 24, 4660–4671. [Google Scholar] [CrossRef]
- Robinson, J.L.; Macarthur, S.; Ross-Innes, C.S.; Tilley, W.D.; Neal, D.E.; Mills, I.G.; Carroll, J.S. Androgen receptor driven transcription in molecular apocrine breast cancer is mediated by FoxA1. EMBO J. 2011, 30, 3019–3027. [Google Scholar] [CrossRef]
- Lehmann-Che, J.; Hamy, A.S.; Porcher, R.; Barritault, M.; Bouhidel, F.; Habuellelah, H.; Leman-Detours, S.; de Roquancourt, A.; Cahen-Doidy, L.; Bourstyn, E.; et al. Molecular apocrine breast cancers are aggressive estrogen receptor negative tumors overexpressing either HER2 or GCDFP15. Breast Cancer Res. 2013, 15, R37. [Google Scholar] [CrossRef]
- Turner, N.C.; Reis-Filho, J.S. Tackling the diversity of triple-negative breast cancer. Clin. Cancer Res. 2013, 19, 6380–6388. [Google Scholar] [CrossRef]
- Lakis, S.; Kotoula, V.; Eleftheraki, A.G.; Batistatou, A.; Bobos, M.; Koletsa, T.; Timotheadou, E.; Chrisafi, S.; Pentheroudakis, G.; Koutras, A.; et al. The androgen receptor as a surrogate marker for molecular apocrine breast cancer subtyping. Breast 2014, 23, 234–243. [Google Scholar] [CrossRef]
- Daemen, A.; Griffith, O.L.; Heiser, L.M.; Wang, N.J.; Enache, O.M.; Sanborn, Z.; Pepin, F.; Durinck, S.; Korkola, J.E.; Griffith, M.; et al. Modeling precision treatment of breast cancer. Genome Biol. 2013, 14, R110. [Google Scholar] [CrossRef]
- Ruffalo, M.; Thomas, R.; Chen, J.; Lee, A.V.; Oesterreich, S.; Bar-Joseph, Z. Network-guided prediction of aromatase inhibitor response in breast cancer. PLoS Comput. Biol. 2019, 15, e1006730. [Google Scholar] [CrossRef] [PubMed]
- Reiner, G.C.; Katzenellenbogen, B.S. Characterization of estrogen and progesterone receptors and the dissociated regulation of growth and progesterone receptor stimulation by estrogen in MDA-MB-134 human breast cancer cells. Cancer Res. 1986, 46, 1124–1131. [Google Scholar]
- Tureci, O.; Sahin, U.; Vollmar, E.; Siemer, S.; Gottert, E.; Seitz, G.; Parkkila, A.K.; Shah, G.N.; Grubb, J.H.; Pfreundschuh, M.; et al. Human carbonic anhydrase XII: cDNA cloning, expression, and chromosomal localization of a carbonic anhydrase gene that is overexpressed in some renal cell cancers. Proc. Natl. Acad. Sci. USA 1998, 95, 7608–7613. [Google Scholar] [CrossRef] [PubMed]
- Prat, A.; Parker, J.S.; Karginova, O.; Fan, C.; Livasy, C.; Herschkowitz, J.I.; He, X.; Perou, C.M. Phenotypic and molecular characterization of the claudin-low intrinsic subtype of breast cancer. Breast Cancer Res. 2010, 12, R68. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Vecchio-Pagan, B.; Sharma, N.; Waheed, A.; Li, X.; Raraigh, K.S.; Robbins, S.; Han, S.T.; Franca, A.L.; Pellicore, M.J.; et al. Loss of carbonic anhydrase XII function in individuals with elevated sweat chloride concentration and pulmonary airway disease. Hum. Mol. Genet. 2016, 25, 1923–1933. [Google Scholar] [CrossRef]
- Kivela, A.J.; Parkkila, S.; Saarnio, J.; Karttunen, T.J.; Kivela, J.; Parkkila, A.K.; Pastorekova, S.; Pastorek, J.; Waheed, A.; Sly, W.S.; et al. Expression of transmembrane carbonic anhydrase isoenzymes IX and XII in normal human pancreas and pancreatic tumours. Histochem. Cell Biol. 2000, 114, 197–204. [Google Scholar] [CrossRef]
- Purkerson, J.M.; Schwartz, G.J. The role of carbonic anhydrases in renal physiology. Kidney Int. 2007, 71, 103–115. [Google Scholar] [CrossRef]
- Ilie, M.I.; Hofman, V.; Ortholan, C.; Ammadi, R.E.; Bonnetaud, C.; Havet, K.; Venissac, N.; Mouroux, J.; Mazure, N.M.; Pouyssegur, J.; et al. Overexpression of carbonic anhydrase XII in tissues from resectable non-small cell lung cancers is a biomarker of good prognosis. Int. J. Cancer 2011, 128, 1614–1623. [Google Scholar] [CrossRef]
- Franke, C.M.; Gu, V.W.; Grimm, B.G.; Cassady, V.C.; White, J.R.; Weigel, R.J.; Kulak, M.V. TFAP2C regulates carbonic anhydrase XII in human breast cancer. Oncogene 2020, 39, 1290–1301. [Google Scholar] [CrossRef]
- Carroll, J.S.; Meyer, C.A.; Song, J.; Li, W.; Geistlinger, T.R.; Eeckhoute, J.; Brodsky, A.S.; Keeton, E.K.; Fertuck, K.C.; Hall, G.F.; et al. Genome-wide analysis of estrogen receptor binding sites. Nat. Genet. 2006, 38, 1289–1297. [Google Scholar] [CrossRef]
- Kong, S.L.; Li, G.; Loh, S.L.; Sung, W.K.; Liu, E.T. Cellular reprogramming by the conjoint action of ERalpha, FOXA1, and GATA3 to a ligand-inducible growth state. Mol. Syst. Biol. 2011, 7, 526. [Google Scholar] [CrossRef] [PubMed]
- Theodorou, V.; Stark, R.; Menon, S.; Carroll, J.S. GATA3 acts upstream of FOXA1 in mediating ESR1 binding by shaping enhancer accessibility. Genome Res. 2013, 23, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Eeckhoute, J.; Carroll, J.S.; Geistlinger, T.R.; Torres-Arzayus, M.I.; Brown, M. A cell-type-specific transcriptional network required for estrogen regulation of cyclin D1 and cell cycle progression in breast cancer. Genes Dev. 2006, 20, 2513–2526. [Google Scholar] [CrossRef] [PubMed]
- Stingl, J.; Raouf, A.; Emerman, J.T.; Eaves, C.J. Epithelial progenitors in the normal human mammary gland. J. Mammary Gland Biol. Neoplasia 2005, 10, 49–59. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).