
MicroRNAs and the Diagnosis of Childhood Acute Lymphoblastic Leukemia: Systematic Review, Meta-Analysis and Re-Analysis with Novel Small RNA-Seq Tools




Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
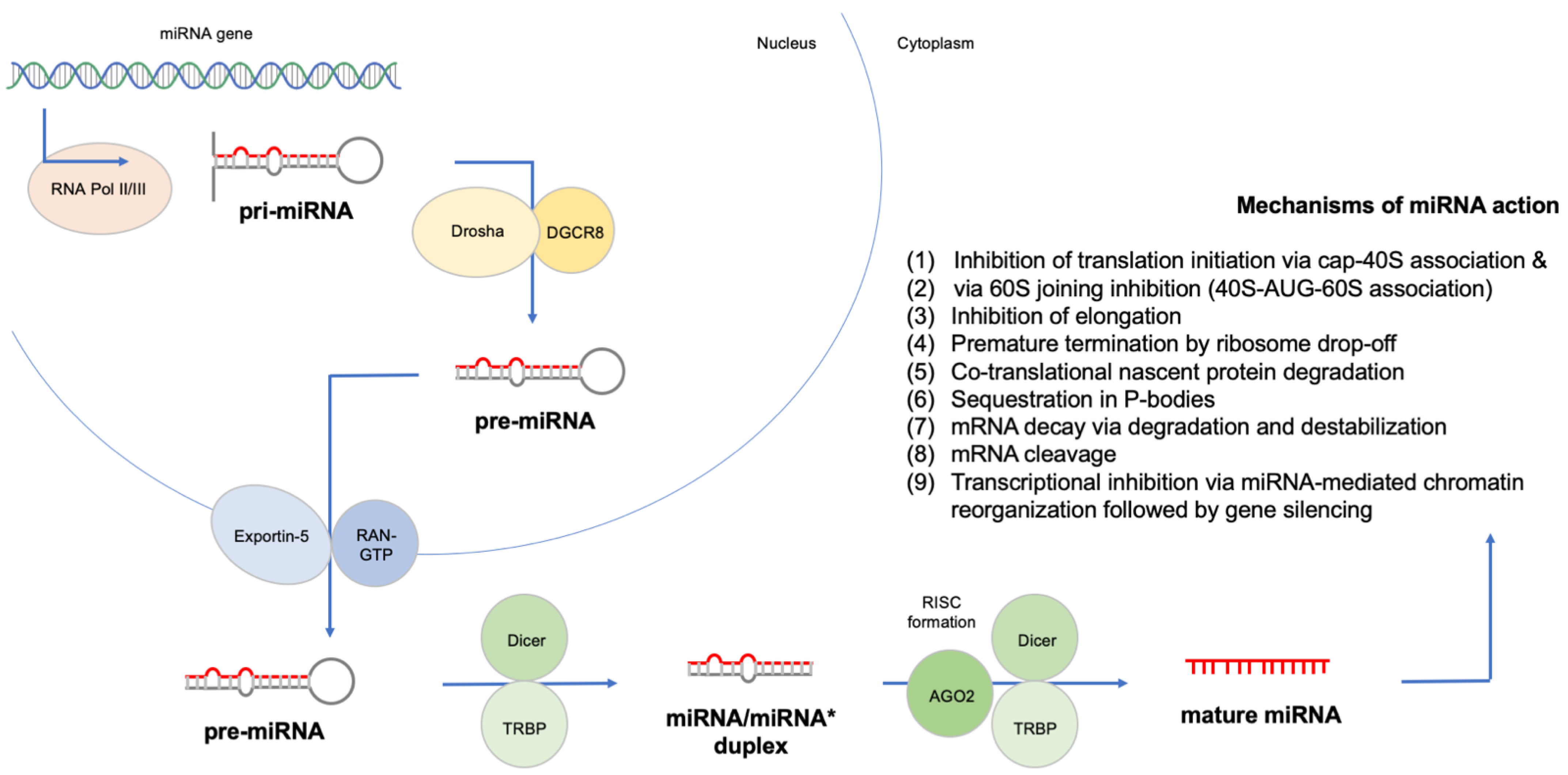
3. Results
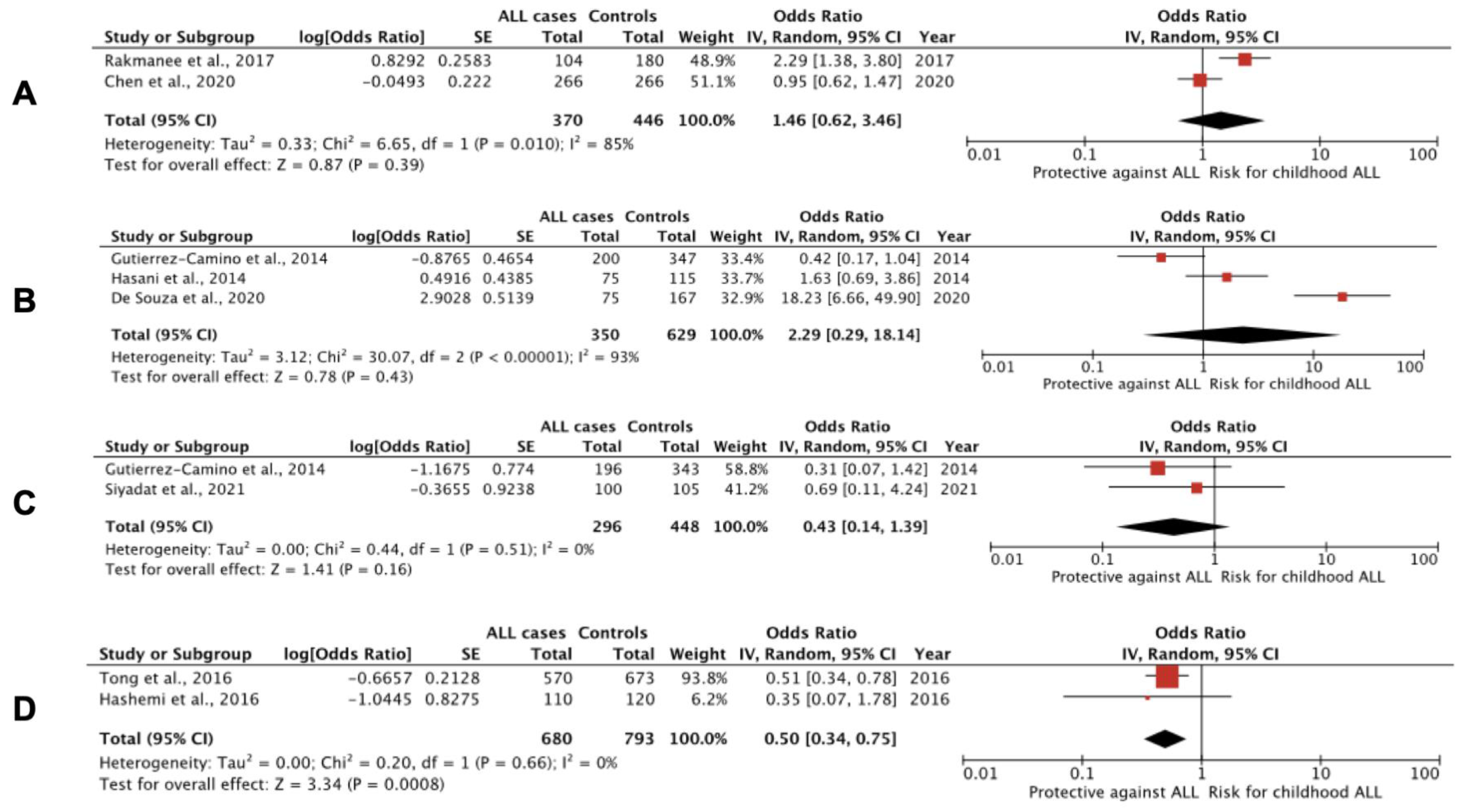
3.1. miRNA Polymorphisms and Childhood ALL


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Cohort Characteristics | Methods | Outcomes |
|---|---|---|---|
| Siyadat et al., 2021 [92] | 100 childhood B-ALL patients and 105 controls; Iran | RT-PCR, HRM and Sanger sequencing; BM or whole blood | No significant relationship was confirmed between rs12803915 variants of miR-612 and susceptibility to B-ALL. |
| Pei et al., 2020 [90] | 266 childhood ALL patients and 266 controls; Taiwan | PCR-RFLP; whole blood | The G allele of miR-146a polymorphism rs2910164 seems to be a protective biomarker for childhood ALL (OR, 0.66; 95% CI: 0.52 to 0.85; p = 0.0011), whereas patients with the GG genotype were associated with decreased susceptibility to ALL (OR, 0.4; 95% CI: 0.23 to 0.67; p = 0.0004) compared with healthy controls. Conspicuously, children bearing CG and GG genotypes were found to have earlier onset compared to those with CC genotype (p = 0.025 and p = 0.0001, respectively). The G>C polymorphism leads to reduced amounts of mature miR-146a, whereas the C allele has reduced capacity to inhibit target genes. |
| C.C. Chen et al., 2020 [93] | 266 childhood ALL patients and 266 controls; Taiwan | PCR-RFLP; whole blood | The T>C polymorphism of rs11614913 in miR-196a-2 does not seem to be associated with susceptibility to childhood ALL. |
| De Souza et al., 2020 [88] | 100 childhood B-ALL patients and 180 adult controls; Brazil | qRT-PCR; whole blood | The mutant homozygote (AA) genotype of DROSHA rs3805500 was associated with a threefold increase in the risk of developing ALL (recessive model: GG + GA vs. AA; OR, 2.913; 95% CI: 1.415 to 5.998; p = 0.004). The mutant homozygote (GG) genotype of the miR-499 gene rs3746444 was associated with a 17.8-fold increase in the risk of developing ALL (recessive model: AA + AG vs. GG; 95% CI: 5.55 to 57.016; p < 0.001). In contrast, the wild homozygous (CC) genotype of the miR-938 gene rs2505901 seems to confer a protective effect against developing ALL (dominant model: CC vs. CT + TT; OR, 0.359; 95% CI: 0.160 to 0.805; p = 0.013). No significant associations for AGO1 rs636832, MIR219A1 rs213210 and rs107822, MIR146A rs2910164, MIR330 rs12894467 and MIR608 rs4919510. |
| Jemimah Devanandan et al., 2019 [47] | 71 childhood ALL patients and 74 controls; India | qRT-PCR; whole blood | No statistically significant association was found between miR-146a gene SNPs rs2910164 G>C and rs57095329 A>G and ALL risk. |
| Xue et al., 2019 [42] | 831 childhood ALL patients and 1079 controls; validation: 88 childhood ALL cases and 99 controls; China | qRT-PCR; whole blood; plasma from validation cohort | Subjects carrying the mutant homozygous TT genotype of miR-100 rs543412 had a statistically significantly decreased risk of childhood ALL (OR, 0.73; 95% CI: 0.55 to 0.97; p = 0.029). No association was been found for miR-146a rs2910164 and miR-210 rs7395206 polymorphisms and ALL. |
| Gutierrez-Camino et al., 2018 [94] | 217 children with B-ALL and 330 controls from Spain and 75 children with B-ALL and 96 controls from Slovenia | GoldenGate genotyping assay and OpenArray genotyping; whole blood or BM | The AA genotype of rs12402181 in miR-3117-3p was associated with B-ALL risk (OR, 1.44; 95% CI: 1.01 to 2.08; p = 0.047; GG vs. AG vs. AA) in the Spanish cohort. The same effect was observed in the Slovenian cohort (OR, 2.01; 95% CI: 1.02 to 3.95; p = 0.041). When both populations were analyzed together, they displayed a more significant trend (OR, 1.53; 95% CI: 1.12 to 2.09; p = 0.006). With respect to allele frequency analysis, minor allele A showed a 1.51-fold increased risk of B-ALL in total (95% CI: 1.11 to 2.05; p = 0.007). The CT/CC genotype in rs62571442 of miR-3689d-2 was associated with a significantly increased risk of developing B-ALL (OR, 1.48; 95% CI: 1.02 to 2.15; p = 0.039) in the Spanish cohort, whereas the risk was even greater in the Slovenian cohort (OR, 3.57; 95% CI: 1.57 to 8.12; p = 0.001). Analyzed together, both cohorts confirmed the latter association (OR, 1.31; 95% CI: 1.06 to 1.60; p = 0.011). Allele frequency analysis in both populations showed that minor allele C is associated with a 1.31-fold increased risk of B-ALL (95% CI: 1.06 to 1.6; p = 0.012). |
| Liu et al., 2018 [95] | 200 childhood ALL patients and 100 controls; China | RFLP; whole blood | The miR-146a rs2910164 CC or CG genotype significantly increased the risk of ALL (frequency of GG, GC and CC genotypes in the patient group and in the control group was 16%, 44.5%, 39.5%, and 29%, 41%, 30%, respectively). The expression of GC/CC genotypes were significantly higher in patients than in controls (GG genotype as reference; for the GC genotype: OR, 1.967; 95% CI: 1.054 to 3.672; p = 0.037; and for the CC genotype: OR, 2.386; 95%CI: 1.239 to 4.595; p = 0.012). |
| Rakmanee et al., 2017 [96] | 104 childhood ALL patients and 180 controls; Thailand | PCR-RFLP; whole blood | Variant CC of rs11614913 in miR-196a-2 (OR, 4.321; 95% CI: 2.091 to 8.930; p < 0.001), TC heterozygote (OR, 2.248; 95% CI: 1.103 to 4.579; p = 0.024) and CC+TC genotypes (OR, 2.921; 95% CI: 1.504 to 5.673; p = 0.001) were associated with childhood ALL susceptibility compared with the TT wild type. CC homozygotes were associated with significantly increased miR-196a-2 expression. |
| Chansing et al., 2016 [97] | 100 childhood ALL patients and 200 controls; Thailand | PCR-RFLP; whole blood | There was no association between miR-146a rs2910164 G>C polymorphism and susceptibility to childhood ALL. |
| Hashemi et al., 2016 [98] | 110 childhood ALL patients and 120 controls; Iran | PCR-RFLP; whole blood | A study of polymorphism rs4938723 in pri-miR-34b/c in ALL and healthy children revealed decreased risk of ALL in heterozygous (OR, 0.48; 95% CI: 0.28 to 0.84; p = 0.012; TC vs. TT), and overdominant (OR, 0.51; 95% CI: 0.3 to 0.89; p = 0.02; TC vs. TT+CC) inheritance models. The C allele significantly decreased the risk of childhood ALL compared to the T allele (OR, 0.52; 95% CI: 0.33 to 0.83; p = 0.006). |
| Tong et al., 2016 [99] | 570 childhood ALL patients at diagnosis and 673 controls; China | RT-PCR; whole blood | The CC genotype in rs4938723 of pri-miR-34b/c was associated with significantly reduced ALL risk (CC vs. TT: OR, 0.51; 95% CI: 0.33 to 0.8; p = 0.003, and CC vs. TT+TC: OR, 0.49; 95% CI: 0.32 to 0.75; p = 0.002). |
| Gutierrez-Camino et al., 2014 [91] | 213 childhood precursor B-ALL patients and 387 adult controls; Spain | OpenArray; remission BM or whole blood | The A allele of rs12803915 in premature mir-612 was found to be protective (GG vs. GA vs. AA; OR, 0.61; 95% CI: 0.42 to 0.88; p = 0.007) against ALL risk. The G allele of rs3746444 in the seed region of mature miR-499-3p was also found to be protective (AA vs. AG vs. GG; OR, 0.67; 95% CI: 0.49 to 0.91; p = 0.009). An association with rs10061133 in mir-449b was also identified (AA vs. GA+GG; OR, 0.52; 95% CI: 0.31 to 0.89; p = 0.012). Eight SNPs present in the six miRNA biogenesis pathway genes (TNRC6B, DROSHA, DGCR8, EIF2C1 or AGO1, CNOT1 and CNOT6) were also identified. |
| Hasani et al., 2014 [89] | 75 childhood ALL patients and 115 controls; Iran | T-ARMS-PCR; whole blood | The G>C variant of rs2910164 in hsa-miR-146a was found to significantly increase risk of ALL (CC vs. GG; OR, 4.24; 95% CI: 1.52 to 11.87; p = 0.006, and GC vs. GG; OR, 3.55; 95% CI: 1.41 to 8.93; p = 0.007; C vs. G; OR 1.73; 95% CI: 1.13 to 2.67; p = 0.012). No association was found between rs3746444 of miR-499 and ALL risk. |
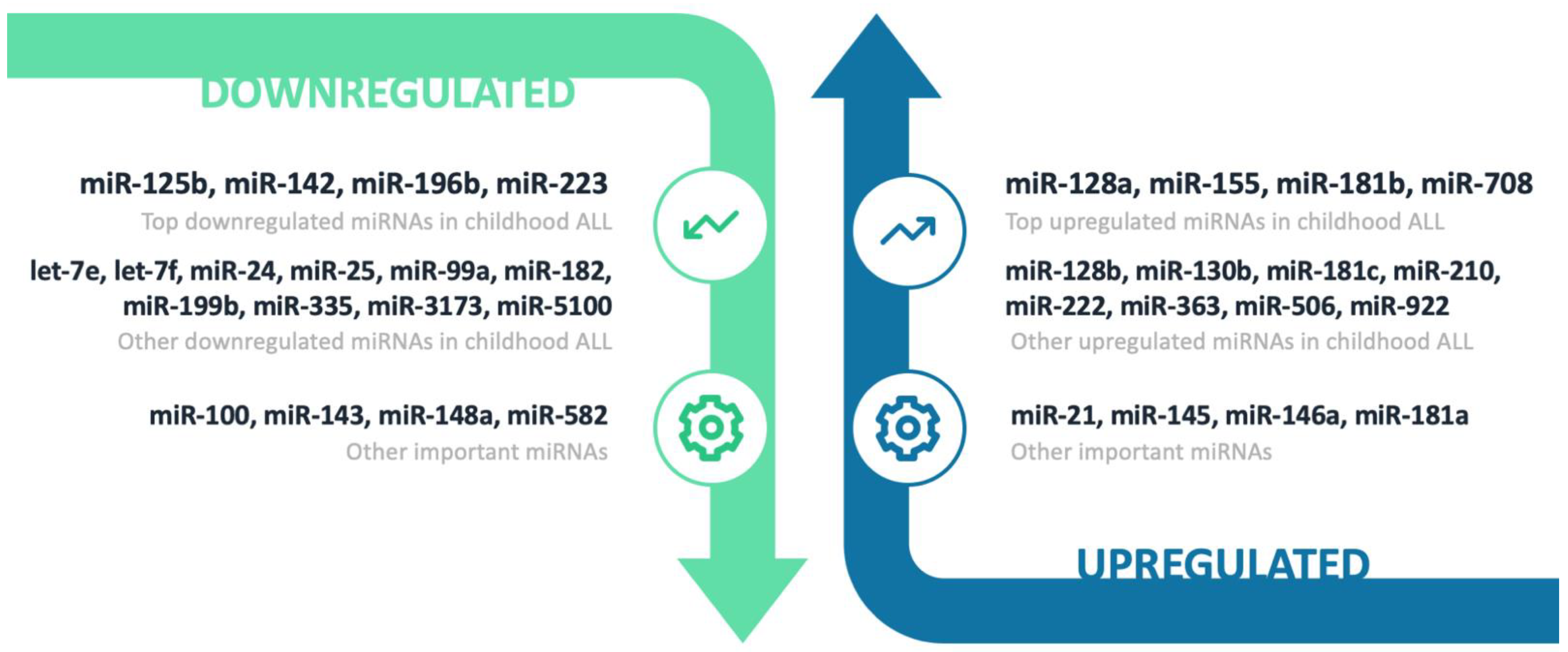
3.2. Downregulated miRNAs
3.2.1. miR-125b
3.2.2. miR-142
3.2.3. miR-196b
3.2.4. miR-223
3.2.5. Other Downregulated miRNAs
3.3. Upregulated miRNAs
3.3.1. miR-128 Family
3.3.2. miR-155
3.3.3. miR-181 Family
3.3.4. miR-708
3.3.5. Other Upregulated miRNAs
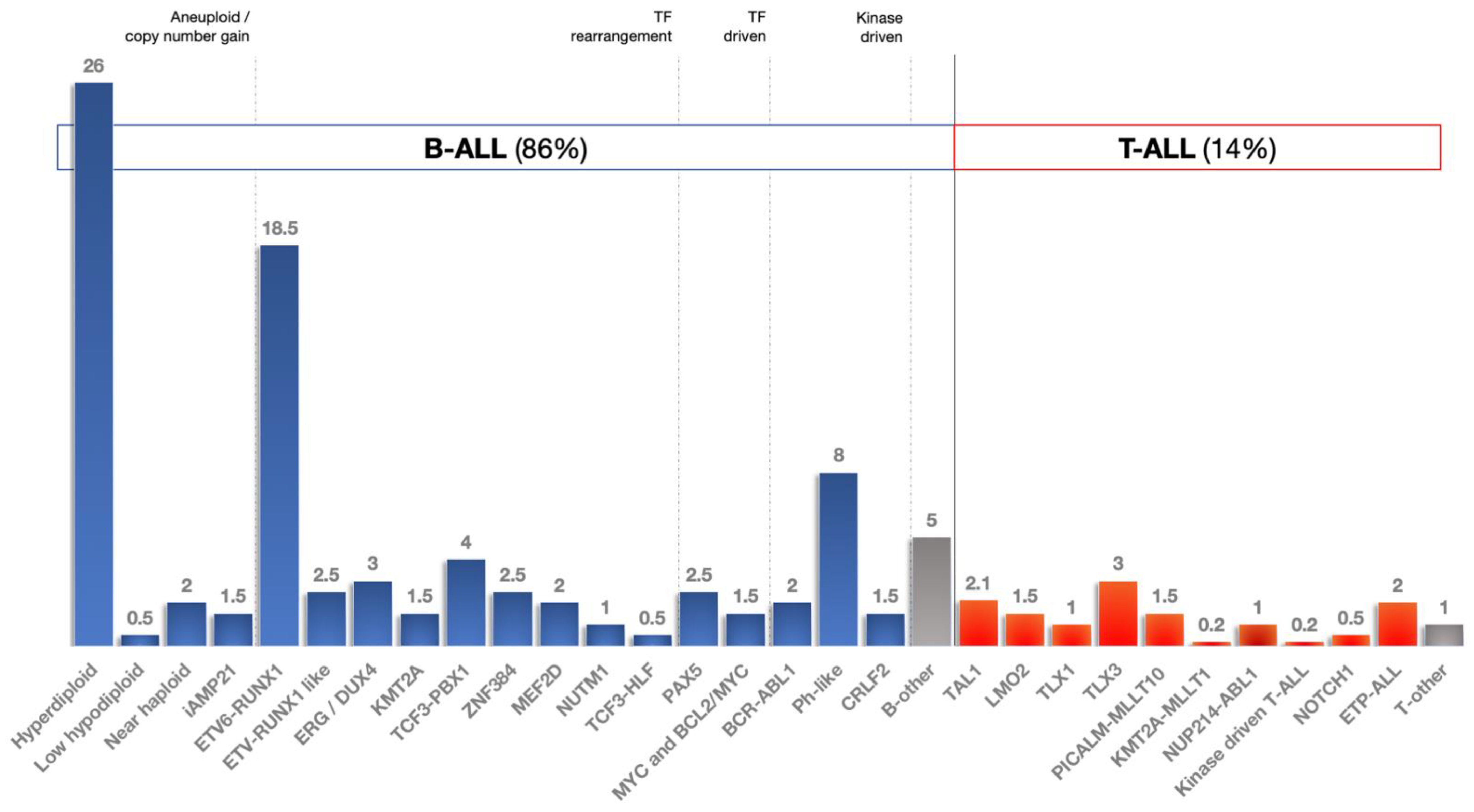
3.4. miRNAs and ALL Subtypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hunger, S.P.; Mullighan, C.G. Redefining ALL classification: Toward detecting high-risk ALL and implementing precision medicine. Blood 2015, 125, 3977–3987. [Google Scholar] [CrossRef] [PubMed]
- Qian, M.; Zhang, H.; Kham, S.K.Y.; Liu, S.; Jiang, C.; Zhao, X.; Lu, Y.; Goodings, C.; Lin, T.N.; Zhang, R.; et al. Whole-transcriptome sequencing identifies a distinct subtype of acute lymphoblastic leukemia with predominant genomic abnormalities of EP300 and CREBBP. Genome Res. 2017, 27, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Dai, Y.T.; Lilljebjörn, H.; Shen, S.H.; Cui, B.W.; Bai, L.; Liu, Y.F.; Qian, M.X.; Kubota, Y.; Kiyoi, H.; et al. Transcriptional landscape of B cell precursor acute lymphoblastic leukemia based on an international study of 1223 cases. Proc. Natl. Acad. Sci. USA 2018, 115, E11711–E11720. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.G. Genetics and prognosis of ALL in children vs. adults. Hematology 2018, 2018, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Malard, F.; Mohty, M. Acute lymphoblastic leukaemia. Lancet 2020, 395, 1146–1162. [Google Scholar] [CrossRef]
- Kyriakidis, I.; Vasileiou, E.; Rossig, C.; Roilides, E.; Groll, A.H.; Tragiannidis, A. Invasive Fungal Diseases in Children with Hematological Malignancies Treated with Therapies That Target Cell Surface Antigens: Monoclonal Antibodies, Immune Checkpoint Inhibitors and CAR T-Cell Therapies. J. Fungi 2021, 7, 186. [Google Scholar] [CrossRef]
- Morozova, N.; Zinovyev, A.; Nonne, N.; Pritchard, L.L.; Gorban, A.N.; Harel-Bellan, A. Kinetic signatures of microRNA modes of action. RNA 2012, 18, 1635–1655. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Rodriguez, P.D.; Paculova, H.; Kogut, S.; Heath, J.; Schjerven, H.; Frietze, S. Non-coding rna signatures of b-cell acute lymphoblastic leukemia. Int. J. Mol. Sci. 2021, 22, 2683. [Google Scholar] [CrossRef]
- Dawidowska, M.; Jaksik, R.; Drobna, M.; Szarzyńska-Zawadzka, B.; Kosmalska, M.; Sędek, Ł.; Machowska, L.; Lalik, A.; Lejman, M.; Ussowicz, M.; et al. Comprehensive Investigation of miRNome Identifies Novel Candidate miRNA-mRNA Interactions Implicated in T-Cell Acute Lymphoblastic Leukemia. Neoplasia 2019, 21, 294–310. [Google Scholar] [CrossRef]
- Olson, N.D.; Wagner, J.; McDaniel, J.; Stephens, S.H.; Westreich, S.T.; Prasanna, A.G.; Johanson, E.; Boja, E.; Maier, E.J.; Serang, O.; et al. PrecisionFDA Truth Challenge V2: Calling variants from short and long reads in difficult-to-map regions. Cell Genom. 2022, 2, 100129. [Google Scholar] [CrossRef]
- Chapman, B.; Kirchner, R.; Pantano, L.; Naumenko, S.; De Smet, M.; Beltrame, L.; Khotiainsteva, T.; Sytchev, I.; Guimera, R.V.; Kern, J.; et al. Bcbio/Bcbio-Nextgen. Available online: https://doi.org/10.5281/ZENODO.5781867 (accessed on 16 August 2022).
- Tsuji, J.; Weng, Z. DNApi: A De Novo Adapter Prediction Algorithm for Small RNA Sequencing Data. PLoS ONE 2016, 11, e0164228. [Google Scholar] [CrossRef] [PubMed]
- Didion, J.P.; Martin, M.; Collins, F.S. Atropos: Specific, sensitive, and speedy trimming of sequencing reads. PeerJ 2017, 2017, e3720. [Google Scholar] [CrossRef]
- Pantano, L.; Estivill, X.; Martí, E. SeqBuster, a bioinformatic tool for the processing and analysis of small RNAs datasets, reveals ubiquitous miRNA modifications in human embryonic cells. Nucleic Acids Res. 2010, 38, e34. [Google Scholar] [CrossRef] [PubMed]
- Pantano, L.; Estivill, X.; Martí, E. A non-biased framework for the annotation and classification of the non-miRNA small RNA transcriptome. Bioinformatics 2011, 27, 3202–3203. [Google Scholar] [CrossRef]
- Mangan, M.E.; Williams, J.M.; Kuhn, R.M.; Lathe, W.C. The UCSC Genome Browser: What Every Molecular Biologist Should Know. Curr. Protoc. Mol. Biol. 2014, 107, 19.9.1–19.9.28. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wallaert, A.; Van Loocke, W.; Hernandez, L.; Taghon, T.; Speleman, F.; Van Vlierberghe, P. Comprehensive miRNA expression profiling in human T-cell acute lymphoblastic leukemia by small RNA-sequencing. Sci. Rep. 2017, 7, 7901. [Google Scholar] [CrossRef]
- Tavakoli, F.; Jaseb, K.; Far, M.A.J.; Soleimani, M.; Khodadi, E.; Saki, N. Evaluation of microRNA-146a expression in acute lymphoblastic leukemia. Front. Biol. 2016, 11, 53–58. [Google Scholar] [CrossRef]
- Miao, M.H.; Ji, X.Q.; Zhang, H.; Xu, J.; Zhu, H.; Shao, X.J. miR-590 promotes cell proliferation and invasion in T-cell acute lymphoblastic leukaemia by inhibiting RB1. Oncotarget 2016, 7, 39527–39534. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Lu, J.; Sun, M.; Li, Z.; Zhang, H.; Neilly, M.B.; Wang, Y.; Qian, Z.; Jin, J.; Zhang, Y.; et al. MicroRNA expression signatures accurately discriminate acute lymphoblastic leukemia from acute myeloid leukemia. Proc. Natl. Acad. Sci. USA 2007, 104, 19971–19976. [Google Scholar] [CrossRef] [PubMed]
- Rashed, W.M.; Hammad, A.M.; Saad, A.M.; Shohdy, K.S. MicroRNA as a diagnostic biomarker in childhood acute lymphoblastic leukemia; systematic review, meta-analysis and recommendations. Crit. Rev. Oncol. Hematol. 2019, 136, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Guan, G.; Dai, Y.; Zhao, P.; Liu, L.; Wang, Q.; Li, X. microRNA-155-5p initiates childhood acute lymphoblastic leukemia by regulating the IRF4/CDK6/CBL axis. Lab. Investig. 2021, 102, 411–421. [Google Scholar] [CrossRef]
- Zamani, A.; Fattahi Dolatabadi, N.; Houshmand, M.; Nabavizadeh, N. miR-324-3p and miR-508-5p expression levels could serve as potential diagnostic and multidrug-resistant biomarkers in childhood acute lymphoblastic leukemia. Leuk. Res. 2021, 109, 106643. [Google Scholar] [CrossRef]
- Liang, C.; Li, Y.; Wang, L.N.; Zhang, X.L.; Luo, J.S.; Peng, C.J.; Tang, W.Y.; Huang, L.B.; Tang, Y.L.; Luo, X.Q. Up-regulated miR-155 is associated with poor prognosis in childhood acute lymphoblastic leukemia and promotes cell proliferation targeting ZNF238. Hematology 2021, 26, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Duyu, M.; Durmaz, B.; Gunduz, C.; Vergin, C.; Yilmaz Karapinar, D.; Aksoylar, S.; Kavakli, K.; Cetingul, N.; Irken, G.; Yaman, Y.; et al. Prospective evaluation of whole genome microrna expression profiling in childhood acute lymphoblastic leukemia. BioMed Res. Int. 2014, 2014, 967585. [Google Scholar] [CrossRef] [PubMed]
- Schotte, D.; de Menezes, R.X.; Moqadam, F.A.; Khankahdani, L.M.; Lange-Turenhout, E.; Chen, C.; Pieters, R.; den Boer, M.L. MicroRNA characterize genetic diversity and drug resistance in pediatric acute lymphoblastic leukemia. Haematologica 2011, 96, 703–711. [Google Scholar] [CrossRef]
- El-maadawy, E.A.; Bakry, R.M.; Moussa, M.M.; El-Naby, S.H.; Talaat, R.M. Alteration in miRNAs expression in paediatric acute lymphocyticleukaemia: Insight into patients’ therapeutic response. Clin. Exp. Pharmacol. Physiol. 2021, 48, 35–43. [Google Scholar] [CrossRef]
- Zhu, Y.-Y.; Wu, R.-N.; Li, X.; Chen, X.-B. Value of serum miR-922 and miR-506 expression levels in the diagnosis and prognostic assessment of childhood acute lymphoblastic leukemia. Chin. J. Contemp. Pediatr. 2021, 23, 1021–1026. [Google Scholar] [CrossRef]
- Fayed, D.; Donia, T.; El-Shanshory, M.; Ali, E.M.M.; Mohamed, T.M. Evaluation of MicroRNA92, MicroRNA638 in Acute Lymphoblastic Leukemia of Egyptian Children. Asian Pac. J. Cancer Prev. 2021, 22, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Li, S.W.; Li, H.; Zhang, Z.P.; Zhuo, F.; Li, Z.X. Expression and Clinical Significance of MiR-146a and MiR-221 in Childhood Acute T Lymphoblastic Leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2020, 28, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, T.; Nie, L.; Zou, Y.; Zhang, Q. MicroRNA-223 decreases cell proliferation, migration, invasion, and enhances cell apoptosis in childhood acute lymphoblastic leukemia via targeting forkhead box O 1. Biosci. Rep. 2020, 40, BSR20200485. [Google Scholar] [CrossRef] [PubMed]
- Shafik, R.E.; Wahab, N.A.; El Mokhtar, M.M.; Taweel, M.A.E.; Ebeid, E. Expression of microRNA-181a and microRNA-196b in Egyptian Pediatric acute Lymphoblastic Leukemia. Asian Pac. J. Cancer Prev. 2020, 21, 3429–3434. [Google Scholar] [CrossRef]
- Akpinar, F.; Polat, A.; Balci, Y.I.; Akca, H.; Senol, H.; Tokgun, O. Microrna expression profiles and changes with treatment on childhood leukemias. UHOD—Uluslar. Hematol. Derg. 2020, 30, 72–80. [Google Scholar] [CrossRef]
- Chen, L.; Shi, Y.; Li, J.; Yang, X.; Li, R.; Zhou, X.; Zhu, L. LncRNA CDKN2B-AS1 contributes to tumorigenesis and chemoresistance in pediatric T-cell acute lymphoblastic leukemia through miR-335-3p/TRAF5 axis. Anticancer Drugs 2020. [Google Scholar] [CrossRef]
- Hassan, N.M.; Refaat, L.A.; Ismail, G.N.; Abdellateif, M.; Fadel, S.A.; AbdelAziz, R.S. Diagnostic, prognostic and predictive values of miR-100 and miR-210 in pediatric acute lymphoblastic Leukemia. Hematology 2020, 25, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.S.; El-Khazragy, N.; Elshimy, A.A.; Aboelhussein, M.M.; Saleh, S.A.; Fadel, S.; Atia, H.A.; Matbouly, S.; Tamer, N. In vitro knock-out of miR-155 suppresses leukemic and HCV virus loads in pediatric HCV-4–associated acute lymphoid leukemia: A promising target therapy. J. Cell. Biochem. 2020, 121, 2811–2817. [Google Scholar] [CrossRef]
- Al Nakeeb, R.H.; Al- Rubaye, D. The expression of different micrornas in iraqi patients with childhood acute leukemia and their association to C/EBP-Β serum level. Iraqi J. Sci. 2020, 61, 2879–2887. [Google Scholar] [CrossRef]
- Sheybani, Z.; Rahgozar, S.; Ghodousi, E.S. The hedgehog signal transducer smoothened and microRNA-326: Pathogenesis and regulation of drug resistance in pediatric B-cell acute lymphoblastic leukemia. Cancer Manag. Res. 2019, 11, 7621–7630. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Yang, X.; Hu, S.; Kang, M.; Chen, J.; Fang, Y. A genetic variant in miR-100 is a protective factor of childhood acute lymphoblastic leukemia. Cancer Med. 2019, 8, 2553–2560. [Google Scholar] [CrossRef] [PubMed]
- Pouyanrad, S.; Rahgozar, S.; Ghodousi, E.S. Dysregulation of miR-335-3p, targeted by NEAT1 and MALAT1 long non-coding RNAs, is associated with poor prognosis in childhood acute lymphoblastic leukemia. Gene 2019, 692, 35–43. [Google Scholar] [CrossRef] [PubMed]
- El-Khazragy, N.; Noshi, M.A.; Abdel-Malak, C.; Zahran, R.F.; Swellam, M. miRNA-155 and miRNA-181a as prognostic biomarkers for pediatric acute lymphoblastic leukemia. J. Cell. Biochem. 2019, 120, 6315–6321. [Google Scholar] [CrossRef] [PubMed]
- El-Khazragy, N.; Elshimy, A.A.; Hassan, S.S.; Matbouly, S.; Safwat, G.; Zannoun, M.; Riad, R.A. Dysregulation of miR-125b predicts poor response to therapy in pediatric acute lymphoblastic leukemia. J. Cell. Biochem. 2019, 120, 7428–7438. [Google Scholar] [CrossRef] [PubMed]
- Rzepiel, A.; Kutszegi, N.; Gézsi, A.; Sági, J.C.; Egyed, B.; Péter, G.; Butz, H.; Nyíro, G.; Müller, J.; Kovács, G.T.; et al. Circulating microRNAs as minimal residual disease biomarkers in childhood acute lymphoblastic leukemia. J. Transl. Med. 2019, 17, 372. [Google Scholar] [CrossRef] [PubMed]
- Jemimah Devanandan, H.; Venkatesan, V.; Scott, J.X.; Magatha, L.S.; Durairaj Paul, S.F.; Koshy, T. MicroRNA 146a Polymorphisms and Expression in Indian Children with Acute Lymphoblastic Leukemia. Lab Med. 2019, 50, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, H.; Lei, D. Microrna-146a promotes growth of acute leukemia cells by downregulating ciliary neurotrophic factor receptor and activating jak2/stat3 signaling. Yonsei Med. J. 2019, 60, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, H.; Zhang, B.; Zhang, X. Expression and role of microRNA-663b in childhood acute lymphocytic leukemia and its mechanism. Open Med. 2019, 14, 863–871. [Google Scholar] [CrossRef]
- Piatopoulou, D.; Avgeris, M.; Drakaki, I.; Marmarinos, A.; Xagorari, M.; Baka, M.; Pourtsidis, A.; Kossiva, L.; Gourgiotis, D.; Scorilas, A. Clinical utility of miR-143/miR-182 levels in prognosis and risk stratification specificity of BFM-treated childhood acute lymphoblastic leukemia. Ann. Hematol. 2018, 97, 1169–1182. [Google Scholar] [CrossRef] [PubMed]
- Swellam, M.; Hashim, M.; Mahmoud, M.S.; Ramadan, A.; Hassan, N.M. Aberrant Expression of Some Circulating miRNAs in Childhood Acute Lymphoblastic Leukemia. Biochem. Genet. 2018, 56, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Ghodousi, E.S.; Rahgozar, S. MicroRNA-326 and microRNA-200c: Two novel biomarkers for diagnosis and prognosis of pediatric acute lymphoblastic leukemia. J. Cell. Biochem. 2018, 119, 6024–6032. [Google Scholar] [CrossRef] [PubMed]
- Shafik, R.E.; Abd El Wahab, N.; Senoun, S.A.; Ebeid, E.; El Taweel, M.A. Expression of micro-RNA 128 and let-7b in pediatric acute lymphoblastic leukemia cases. Asian Pac. J. Cancer Prev. 2018, 19, 2263–2267. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Lu, X.; Huang, P.; Gao, C.; Zhao, X.; Xing, T.; Li, G.; Bao, S.; Zheng, H. Expression of miR-652-3p and Effect on Apoptosis and Drug Sensitivity in Pediatric Acute Lymphoblastic Leukemia. BioMed Res. Int. 2018, 2018, 5724686. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Yu, R.; Bai, Y.; Chen, X. MicroRNA-9 suppresses cancer proliferation and cell cycle progression in acute lymphoblastic leukemia with inverse association of neuropilin-1. J. Cell. Biochem. 2018, 119, 6604–6613. [Google Scholar] [CrossRef] [PubMed]
- Asnafi, A.A.; Khodadi, E.; Golchin, N.; Alghasi, A.; Tavakolifar, Y.; Saki, N. Association between microRNA-21, microRNA-150, and micro-RNA-451 expression and clinical outcome of patients with acute lymphoblastic leukemia. Front. Biol. 2017, 12, 63–70. [Google Scholar] [CrossRef]
- Piatopoulou, D.; Avgeris, M.; Marmarinos, A.; Xagorari, M.; Baka, M.; Doganis, D.; Kossiva, L.; Scorilas, A.; Gourgiotis, D. MiR-125b predicts childhood acute lymphoblastic leukaemia poor response to BFM chemotherapy treatment. Br. J. Cancer 2017, 117, 801–812. [Google Scholar] [CrossRef]
- Nabhan, M.; Louka, M.L.; Khairy, E.; Tash, F.; Ali-Labib, R.; El-Habashy, S. MicroRNA-181a and its target Smad 7 as potential biomarkers for tracking child acute lymphoblastic leukemia. Gene 2017, 628, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Wu, S.; Huang, S.; Jiang, W.; Ke, Y. Association between expression of plasma miRNA and the risk of childhood acute lymphocytic leukemia. Chin. J. Epidemiol. 2017, 38, 1252–1258. [Google Scholar] [CrossRef]
- Labib, H.A.; Elantouny, N.G.; Ibrahim, N.F.; Alnagar, A.A. Upregulation of microRNA-21 is a poor prognostic marker in patients with childhood B cell acute lymphoblastic leukemia. Hematology 2017, 22, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Cao, J.; Ji, Q.; Zhang, C.; Qian, T.; Song, X.; Huang, B.; Tian, X. The downregulation of miR-3173 in B-cell acute lymphoblastic leukaemia promotes cell invasion via PTK2. Biochem. Biophys. Res. Commun. 2017, 494, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Ramani, R.; Megason, G.; Schallheim, J.; Karlson, C.; Vijayakumar, V.; Vijayakumar, S.; Hicks, C. Integrative analysis of microRNA-mediated gene signatures and pathways modulating white blood cell count in childhood acute lymphoblastic leukemia. Biomark. Insights 2017, 12, 1177271917702895. [Google Scholar] [CrossRef]
- Cao, L.; Wang, N.; Pan, J.; Hu, S.; Zhao, W.; He, H.; Wang, Y.; Gu, G.; Chai, Y. Clinical significance of microRNA-34b expression in pediatric acute leukemia. Mol. Med. Rep. 2016, 13, 2777–2784. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Liu, L.; Zhan, L.; Wang, X.; Fan, H. MIR-187-5p regulates cell growth and apoptosis in acute lymphoblastic Leukemia via DKK2. Oncol. Res. 2016, 24, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Swellam, M.; El-Khazragy, N. Clinical impact of circulating microRNAs as blood-based marker in childhood acute lymphoblastic leukemia. Tumor Biol. 2016, 37, 10571–10576. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, J.C.; Scrideli, C.A.; Brassesco, M.S.; Yunes, J.A.; Brandalise, S.R.; Tone, L.G. MiR-708-5p is differentially expressed in childhood acute lymphoblastic leukemia but not strongly associated to clinical features. Pediatr. Blood Cancer 2015, 62, 177–178. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Eguchi-Ishimae, M.; Yagi, C.; Iwabuki, H.; Gao, W.; Tauchi, H.; Inukai, T.; Sugita, K.; Ishii, E.; Eguchi, M. HMGA2 as a potential molecular target in KMT2A-AFF1-positive infant acute lymphoblastic leukaemia. Br. J. Haematol. 2015, 171, 818–829. [Google Scholar] [CrossRef] [PubMed]
- Nemes, K.; Csóka, M.; Nagy, N.; Márk, Á.; Váradi, Z.; Dankó, T.; Kovács, G.; Kopper, L.; Sebestyén, A. Expression of Certain Leukemia/Lymphoma Related microRNAs and its Correlation with Prognosis in Childhood Acute Lymphoblastic Leukemia. Pathol. Oncol. Res. 2015, 21, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Luna-Aguirre, C.M.; Martinez-Fierro, M.D.L.L.; Mar-Aguilar, F.; Garza-Veloz, I.; Treviño-Alvaradod, V.; Rojas-Martinez, A.; Jaime-Perez, J.C.; Malagon-Santiago, G.I.; Gutierrez-Aguirre, C.H.; Gonzalez-Llano, O.; et al. Circulating microRNA expression profile in B-cell acute lymphoblastic leukemia. Cancer Biomark. 2015, 15, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Organista-Nava, J.; Gómez-Gómez, Y.; Illades-Aguiar, B.; Del Carmen Alarcón-Romero, L.; Saavedra-Herrera, M.V.; Rivera-Ramírez, A.B.; Garzón-Barrientos, V.H.; Leyva-Vázquez, M.A. High miR-24 expression is associated with risk of relapse and poor survival in acute leukemia. Oncol. Rep. 2015, 33, 1639–1649. [Google Scholar] [CrossRef]
- Oliveira, L.H.; Schiavinato, J.L.; Fráguas, M.S.; Lucena-Araujo, A.R.; Haddad, R.; Araújo, A.G.; Dalmazzo, L.F.; Rego, E.M.; Covas, D.T.; Zago, M.A.; et al. Potential roles of microRNA-29a in the molecular pathophysiology of T-cell acute lymphoblastic leukemia. Cancer Sci. 2015, 106, 1264–1277. [Google Scholar] [CrossRef]
- Malik, D.; Kaul, D.; Chauhan, N.; Marwaha, R.K. MiR-2909-mediated regulation of KLF4: A novel molecular mechanism for differentiating between B-cell and T-cell pediatric acute lymphoblastic leukemias. Mol. Cancer 2014, 13, 175. [Google Scholar] [CrossRef] [PubMed]
- Gimenes-Teixeira, H.L.; Lucena-Araujo, A.R.; dos Santos, G.A.; Zanette, D.L.; Scheucher, P.S.; Oliveira, L.C.; Dalmazzo, L.F.; Silva-Júnior, W.A.; Falcão, R.P.; Rego, E.M. Increased expression of miR-221 is associated with shorter overall survival in T-cell acute lymphoid leukemia. Exp. Hematol. Oncol. 2013, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, D.; Zhuang, Y.; Shi, Q.; Wei, W.; Zhang, H.; Ju, X. li [The expression and regulatory mechanism of microRNA-708 in pediatric common B-cell acute lymphoblastic leukemia]. Zhonghua Xue Ye Xue Za Zhi 2013, 34, 138–143. [Google Scholar] [PubMed]
- Li, X.J.; Luo, X.Q.; Han, B.W.; Duan, F.T.; Wei, P.P.; Chen, Y.Q. MicroRNA-100/99a, deregulated in acute lymphoblastic leukaemia, suppress proliferation and promote apoptosis by regulating the FKBP51 and IGF1R/mTOR signalling pathways. Br. J. Cancer 2013, 109, 2189–2198. [Google Scholar] [CrossRef]
- De Oliveira, J.C.; Scrideli, C.A.; Brassesco, M.S.; Morales, A.G.; Pezuk, J.A.; de Paula Queiroz, R.; Yunes, J.A.; Brandalise, S.R.; Tone, L.G. Differential MiRNA expression in childhood acute lymphoblastic leukemia and association with clinical and biological features. Leuk. Res. 2012, 36, 293–298. [Google Scholar] [CrossRef]
- Borze, I.; Guled, M.; Musse, S.; Raunio, A.; Elonen, E.; Saarinen-Pihkala, U.; Karjalainen-Lindsberg, M.L.; Lahti, L.; Knuutila, S. MicroRNA microarrays on archive bone marrow core biopsies of leukemias-Method validation. Leuk. Res. 2011, 35, 188–195. [Google Scholar] [CrossRef]
- Bhatia, S.; Kaul, D.; Varma, N. Functional genomics of tumor suppressor miR-196b in T-cell acute lymphoblastic leukemia. Mol. Cell. Biochem. 2011, 346, 103–116. [Google Scholar] [CrossRef]
- Mavrakis, K.J.; Van Der Meulen, J.; Wolfe, A.L.; Liu, X.; Mets, E.; Taghon, T.; Khan, A.A.; Setti, M.; Rondou, P.; Vandenberghe, P.; et al. A cooperative microRNA-tumor suppressor gene network in acute T-cell lymphoblastic leukemia (T-ALL). Nat. Genet. 2011, 43, 673–678. [Google Scholar] [CrossRef]
- Stumpel, D.J.P.M.; Schotte, D.; Lange-Turenhout, E.A.M.; Schneider, P.; Seslija, L.; De Menezes, R.X.; Marquez, V.E.; Pieters, R.; Den Boer, M.L.; Stam, R.W. Hypermethylation of specific microRNA genes in MLL-rearranged infant acute lymphoblastic leukemia: Major matters at a micro scale. Leukemia 2011, 25, 429–439. [Google Scholar] [CrossRef]
- Schotte, D.; Moqadam, F.A.; Lange-Turenhout, E.A.M.; Chen, C.; Van Ijcken, W.F.J.; Pieters, R.; Den Boer, M.L. Discovery of new microRNAs by small RNAome deep sequencing in childhood acute lymphoblastic leukemia. Leukemia 2011, 25, 1389–1399. [Google Scholar] [CrossRef]
- Bhatia, S.; Kaul, D.; Varma, N. Potential tumor suppressive function of miR-196b in B-cell lineage acute lymphoblastic leukemia. Mol. Cell. Biochem. 2010, 340, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Schotte, D.; Chau, J.C.K.; Sylvester, G.; Liu, G.; Chen, C.; van der Velden, V.H.J.; Broekhuis, M.J.C.; Peters, T.C.J.M.; Pieters, R.; den Boer, M.L. Identification of new microRNA genes and aberrant microRNA profiles in childhood acute lymphoblastic leukemia. Leukemia 2009, 23, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, J.H.; Zheng, Y.S.; Zhang, P.; Chen, X.; Wu, J.; Xu, L.; Luo, X.Q.; Ke, Z.Y.; Zhou, H.; et al. Genome-wide analysis of small RNA and novel microRNA discovery in human acute lymphoblastic leukemia based on extensive sequencing approach. PLoS ONE 2009, 4, e6849. [Google Scholar] [CrossRef]
- Ju, X.; Li, D.; Shi, Q.; Hou, H.; Sun, N.; Shen, B. Differential microRNA expression in childhood B-cell precursor acute lymphoblastic leukemia. Pediatr. Hematol. Oncol. 2009, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Luo, X.Q.; Zhang, P.; Huang, L.B.; Zheng, Y.S.; Wu, J.; Zhou, H.; Qu, L.H.; Xu, L.; Chen, Y.Q. MicroRNA patterns associated with clinical prognostic parameters and CNS relapse prediction in pediatric acute leukemia. PLoS ONE 2009, 4, e7826. [Google Scholar] [CrossRef]
- Zou, D.; Yin, J.; Ye, Z.; Zeng, Q.; Tian, C.; Wang, Y.; Chen, Q.; Chen, R. Association Between the miR-146a Rs2910164 Polymorphism and Childhood Acute Lymphoblastic Leukemia Susceptibility in an Asian Population. Front. Genet. 2020, 11, 886. [Google Scholar] [CrossRef]
- De Souza, T.P.; de Carvalho, D.C.; Wanderley, A.V.; Fernandes, S.M.; Gomes Rodrigues, J.C.; Cohen-Paes, A.; Fernandes, M.R.; Mello, F.A.R.; Pastana, L.F.; Vinagre, L.W.M.S.; et al. Influence of variants of the drosha, mir499a, and mir938 genes on susceptibility to acute lymphoblastic leukemia in an admixed population from the brazilian amazon. Am. J. Transl. Res. 2021, 12, 8216–8224. [Google Scholar]
- Hasani, S.S.; Hashemi, M.; Eskandari-Nasab, E.; Naderi, M.; Omrani, M.; Sheybani-Nasab, M. A functional polymorphism in the miR-146a gene is associated with the risk of childhood acute lymphoblastic leukemia: A preliminary report. Tumor Biol. 2014, 35, 219–225. [Google Scholar] [CrossRef]
- Pei, J.S.; Chang, W.S.; Hsu, P.C.; Chen, C.C.; Chin, Y.T.; Huang, T.L.; Hsu, Y.N.; Kuo, C.C.; Wang, Y.C.; Tsai, C.W.; et al. Significant association βetween the mIR146a genotypes and susceptibility to childhood acute lymphoblastic leukemia in Taiwan. Cancer Genom. Proteom. 2020, 17, 175–180. [Google Scholar] [CrossRef]
- Gutierrez-Camino, A.; Lopez-Lopez, E.; Martin-Guerrero, I.; Piñan, M.A.; Garcia-Miguel, P.; Sanchez-Toledo, J.; Carbone Bañeres, A.; Uriz, J.; Navajas, A.; Garcia-Orad, A. Noncoding RNA-related polymorphisms in pediatric acute lymphoblastic leukemia susceptibility. Pediatr. Res. 2014, 75, 767–773. [Google Scholar] [CrossRef]
- Siyadat, P.; Ayatollahi, H.; Barati, M.; Sheikhi, M.; Shahidi, M. High Resolution Melting Analysis for Evaluation of mir-612 (Rs12803915) Genetic Variant with Susceptibility to Pediatric Acute Lymphoblastic Leukemia. Rep. Biochem. Mol. Biol. 2021, 9, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Hsu, P.C.; Shih, L.C.; Hsu, Y.N.; Kuo, C.C.; Chao, C.Y.; Chang, W.S.; Tsai, C.W.; Bau, D.T.; Pei, J.S. MiR-196a-2 Genotypes determine the susceptibility and early onset of childhood acute lymphoblastic leukemia. Anticancer Res. 2020, 40, 4465–4469. [Google Scholar] [CrossRef]
- Gutierrez-Camino, A.; Martin-Guerrero, I.; Dolzan, V.; Jazbec, J.; Carbone-Bañeres, A.; de Andoin, N.G.; Sastre, A.; Astigarraga, I.; Navajas, A.; Garcia-Orad, A. Involvement of SNPs in miR-3117 and miR-3689d2 in childhood acute lymphoblastic leukemia risk. Oncotarget 2018, 9, 22907–22914. [Google Scholar] [CrossRef] [PubMed]
- LIU, X.; LIU, L.; CAO, Z.; GUO, B.; LI, M. Association between miR146a(rs2910164)G>C polymorphism and susceptibility to acute lymphoblastic leuke-mia in children. Chin. J. Appl. Clin. Pediatr. 2018, 24, 200–202. [Google Scholar] [CrossRef]
- Rakmanee, S.; Pakakasama, S.; Hongeng, S.; Sanguansin, S.; Thongmee, A.; Pongstaporn, W. Increased risk of Thai childhood acute lymphoblastic leukemia with the MiR196a2 T > C polymorphism. Asian Pac. J. Cancer Prev. 2017, 18, 1117–1120. [Google Scholar] [CrossRef]
- Chansing, K.; Pakakasama, S.; Hongeng, S.; Thongmee, A.; Pongstaporn, W. Lack of association between the MiR146a polymorphism and susceptibility to Thai childhood acute lymphoblastic leukemia. Asian Pac. J. Cancer Prev. 2016, 17, 2435–2438. [Google Scholar] [CrossRef]
- Hashemi, M.; Bahari, G.; Naderi, M.; Sadeghi-Bojd, S.; Taheri, M. Pri-miR-34b/c rs4938723 polymorphism is associated with the risk of childhood acute lymphoblastic leukemia. Cancer Genet. 2016, 209, 493–496. [Google Scholar] [CrossRef]
- Tong, N.; Chu, H.; Wang, M.; Xue, Y.; Du, M.; Lu, L.; Zhang, H.; Wang, F.; Fang, Y.; Li, J.; et al. Pri-miR-34b/c rs4938723 polymorphism contributes to acute lymphoblastic leukemia susceptibility in Chinese children. Leuk. Lymphoma 2016, 57, 1436–1441. [Google Scholar] [CrossRef]
- Akbari Moqadam, F.; Lange-Turenhout, E.A.M.; Van Der Veer, A.; Marchante, J.R.M.; Boer, J.M.; Pieters, R.; Den Boer, M. MicroRNA signature in BCR-ABL1-like and BCR-ABL1-positive childhood acute lymphoblastic leukemia: Similarities and dissimilarities. Leuk. Lymphoma 2014, 55, 1942–1945. [Google Scholar] [CrossRef]
- Gefen, N.; Binder, V.; Zaliova, M.; Linka, Y.; Morrow, M.; Novosel, A.; Edry, L.; Hertzberg, L.; Shomron, N.; Williams, O.; et al. Hsa-mir-125b-2 is highly expressed in childhood ETV6/RUNX1 (TEL/AML1) leukemias and confers survival advantage to growth inhibitory signals independent of p53. Leukemia 2010, 24, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Vendramini, E.; Giordan, M.; Giarin, E.; Michielotto, B.; Fazio, G.; Cazzaniga, G.; Biondi, A.; Silvestri, D.; Valsecchi, M.G.; Muckenthaler, M.U.; et al. High expression of miR-125b-2 and SNORD116 noncoding RNA clusters characterize ERG-related B cell precursor acute lymphoblastic leukemia. Oncotarget 2017, 8, 42398–42413. [Google Scholar] [CrossRef]
- Shaham, L.; Binder, V.; Gefen, N.; Borkhardt, A.; Izraeli, S. MiR-125 in normal and malignant hematopoiesis. Leukemia 2012, 26, 2011–2018. [Google Scholar] [CrossRef] [PubMed]
- Tassano, E.; Acquila, M.; Tavella, E.; Micalizzi, C.; Panarello, C.; Morerio, C. MicroRNA-125b-1 and BLID upregulation resulting from a novel IGH translocation in childhood B-cell precursor acute lymphoblastic leukemia. Genes Chromosom. Cancer 2010, 49, 682–687. [Google Scholar] [CrossRef]
- Doerrenberg, M.; Kloetgen, A.; Hezaveh, K.; Wössmann, W.; Bleckmann, K.; Stanulla, M.; Schrappe, M.; McHardy, A.C.; Borkhardt, A.; Hoell, J.I. T-cell acute lymphoblastic leukemia in infants has distinct genetic and epigenetic features compared to childhood cases. Genes Chromosom. Cancer 2017, 56, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Renou, L.; Boelle, P.Y.; Deswarte, C.; Spicuglia, S.; Benyoucef, A.; Calvo, J.; Uzan, B.; Belhocine, M.; Cieslak, A.; Landman-Parker, J.; et al. Homeobox protein TLX3 activates miR-125b expression to promote T-cell acute lymphoblastic leukemia. Blood Adv. 2017, 1, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Hu, H.; Gu, W. Regulation Effects of Circular RNA CircPVT1 and miR-125b on NF-κB Signal Pathway in Childhood ALL. Clin. Lab. 2021, 67, 2491–2500. [Google Scholar] [CrossRef] [PubMed]
- Vafadar, A.; Mokaram, P.; Erfani, M.; Yousefi, Z.; Farhadi, A.; Elham Shirazi, T.; Tamaddon, G. The effect of decitabine on the expression and methylation of the PPP1CA, BTG2, and PTEN in association with changes in miR-125b, miR-17, and miR-181b in NALM6 cell line. J. Cell. Biochem. 2019, 120, 13156–13167. [Google Scholar] [CrossRef]
- Puissegur, M.P.; Eichner, R.; Quelen, C.; Coyaud, E.; Mari, B.; Lebrigand, K.; Broccardo, C.; Nguyen-Khac, F.; Bousquet, M.; Brousset, P. B-cell regulator of immunoglobulin heavy-chain transcription (Bright)/ARID3a is a direct target of the oncomir microRNA-125b in progenitor B-cells. Leukemia 2012, 26, 2224–2232. [Google Scholar] [CrossRef] [PubMed]
- Willimott, S.; Wagner, S.D. miR-125b and miR-155 contribute to BCL2 repression and proliferation in response to CD40 ligand (CD154) in human leukemic B-cells. J. Biol. Chem. 2012, 287, 2608–2617. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; Li, J.; Zheng, D.; Li, Y.; Gao, X.; Xu, C.; Gao, L.; Wang, L.; Yu, L. MicroRNA-142-3p inhibits cell proliferation in human acute lymphoblastic leukemia by targeting the MLL-AF4 oncogene. Mol. Biol. Rep. 2013, 40, 6811–6819. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, D.; Zhang, G.; Li, X.; Liang, Y.; Kasukurthi, M.V.; Li, S.; Borchert, G.M.; Huang, J. A semantics-oriented computational approach to investigate microRNA regulation on glucocorticoid resistance in pediatric acute lymphoblastic leukemia. BMC Med. Inform. Decis. Mak. 2018, 18, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Zhang, X.; Jia, H.; Li, D.; Zhang, B.; Zhang, H.; Hong, M.; Jiang, T.; Jiang, Q.; Lu, J.; et al. An oncogenic role of miR-142-3p in human T-cell acute lymphoblastic leukemia (T-ALL) by targeting glucocorticoid receptor-α and cAMP/PKA pathways. Leukemia 2012, 26, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Croce, C.M. MicroRNA-cancer connection: The beginning of a new tale. Cancer Res. 2006, 66, 7390–7394. [Google Scholar] [CrossRef] [PubMed]
- Kramer, N.J.; Wang, W.L.; Reyes, E.Y.; Kumar, B.; Chen, C.C.; Ramakrishna, C.; Cantin, E.M.; Vonderfecht, S.L.; Taganov, K.D.; Chau, N.; et al. Altered lymphopoiesis and immunodeficiency in miR-142 null mice. Blood 2015, 125, 3720–3730. [Google Scholar] [CrossRef] [PubMed]
- Schotte, D.; Lange-Turenhout, E.A.M.; Stumpel, D.J.P.M.; Stam, R.W.; Buijs-Gladdines, J.G.C.A.M.; Meijerink, J.P.P.; Pieters, R.; den Boer, M.L. Expression of miR-196b is not exclusively MLL-driven but is especially linked to activation of HOXA genes in pediatric acute lymphoblastic leukemia. Haematologica 2010, 95, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, H.; Chen, P.; He, M.; Li, Y.; Arnovitz, S.; Jiang, X.; He, C.; Hyjek, E.; Zhang, J.; et al. miR-196b directly targets both HOXA9/MEIS1 oncogenes and FAS tumour suppressor in MLL-rearranged leukaemia. Nat. Commun. 2012, 3, 688. [Google Scholar] [CrossRef] [PubMed]
- Avigad, S.; Verly, I.R.; Lebel, A.; Kordi, O.; Shichrur, K.; Ohali, A.; Hameiri-Grossman, M.; Kaspers, G.J.; Cloos, J.; Fronkova, E.; et al. miR expression profiling at diagnosis predicts relapse in pediatric precursor B-cell acute lymphoblastic leukemia. Genes Chromosom. Cancer 2016, 55, 328–339. [Google Scholar] [CrossRef]
- Zhou, W.; Shunqing, W.; Yi, Y.; Zhou, R.; Mao, P. MiR-196b/miR-1290 participate in the antitumor effect of resveratrol via regulation of IGFBP3 expression in acute lymphoblastic leukemia. Oncol. Rep. 2017, 37, 1075–1083. [Google Scholar] [CrossRef]
- Roman-Gomez, J.; Agirre, X.; Jiménez-Velasco, A.; Arqueros, V.; Vilas-Zornoza, A.; Rodriguez-Otero, P.; Martin-Subero, I.; Garate, L.; Corden, L.; José-Eneriz, E.S.; et al. Epigenetic regulation of MicroRNAs in acute lymphoblastic leukemia. J. Clin. Oncol. 2009, 27, 1316–1322. [Google Scholar] [CrossRef]
- Hu, X.-Y.; Yin, J.-J.; Chao, R.; Zhu, S.-D.; Deng, W.; Pan, M. [Correlation of Plasma MiR-146a, MiR-223 Levels with the Prognosis of Childhood with Acute Lymphoblastic Leukemia]. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2021, 29, 1727–1732. [Google Scholar] [CrossRef]
- Zhou, K.; Yi, S.; Yu, Z.; Li, Z.; Wang, Y.; Zou, D.; Qi, J.; Zhao, Y.; Qiu, L. MicroRNA-223 expression is uniformly down-regulated in B cell lymphoproliferative disorders and is associated with poor survival in patients with chronic lymphocytic leukemia. Leuk. Lymphoma 2012, 53, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.Y.; Wang, F.; Yu, J.; Yang, G.H.; Liu, X.L.; Zhang, J.W. MicroRNA-223 reversibly regulates erythroid and megakaryocytic differentiation of K562 cells. J. Cell. Mol. Med. 2009, 13, 4551–4559. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.R.; Sanda, T.; Lawton, L.N.; Li, X.; Kreslavsky, T.; Novina, C.D.; Brand, M.; Gutierrez, A.; Kelliher, M.A.; Jamieson, C.H.M.; et al. The TAL1 complex targets the FBXW7 tumor suppressor by activating miR-223 in human T cell acute lymphoblastic leukemia. J. Exp. Med. 2013, 210, 1545–1557. [Google Scholar] [CrossRef] [PubMed]
- Mets, E.; Van Der Meulen, J.; Van Peer, G.; Boice, M.; Mestdagh, P.; Van De Walle, I.; Lammens, T.; Goossens, S.; De Moerloose, B.; Benoit, Y.; et al. MicroRNA-193b-3p acts as a tumor suppressor by targeting the MYB oncogene in T-cell acute lymphoblastic leukemia. Leukemia 2015, 29, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.D.; Wang, L.; Sun, C.; Fan, L.; Zhu, D.X.; Fang, C.; Wang, Y.H.; Zou, Z.J.; Zhang, S.J.; Li, J.Y.; et al. Distinctive microRNA signature is associated with the diagnosis and prognosis of acute leukemia. Med. Oncol. 2012, 29, 2323–2331. [Google Scholar] [CrossRef] [PubMed]
- Rainer, J.; Ploner, C.; Jesacher, S.; Ploner, A.; Eduardoff, M.; Mansha, M.; Wasim, M.; Panzer-Grümayer, R.; Trajanoski, Z.; Niederegger, H.; et al. Glucocorticoid-regulated microRNAs and mirtrons in acute lymphoblastic leukemia. Leukemia 2009, 23, 746–752. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Metformin may antagonize Lin28 and/or Lin28B activity, thereby boosting let-7 levels and antagonizing cancer progression. Med. Hypotheses 2012, 78, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Silver, S.J.; Hagen, J.W.; Okamura, K.; Perrimon, N.; Lai, E.C. Functional screening identifies miR-315 as a potent activator of Wingless signaling. Proc. Natl. Acad. Sci. USA 2007, 104, 18151–18156. [Google Scholar] [CrossRef] [PubMed]
- Nishi, M.; Eguchi-Ishimae, M.; Wu, Z.; Gao, W.; Iwabuki, H.; Kawakami, S.; Tauchi, H.; Inukai, T.; Sugita, K.; Hamasaki, Y.; et al. Suppression of the let-7b microRNA pathway by DNA hypermethylation in infant acute lymphoblastic leukemia with MLL gene rearrangements. Leukemia 2013, 27, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Krzanowski, J.; Madzio, J.; Pastorczak, A.; Tracz, A.; Braun, M.; Tabarkiewicz, J.; Pluta, A.; Młynarski, W.; Zawlik, I. Selected miRNA levels are associated with IKZF1 microdeletions in pediatric acute lymphoblastic leukemia. Oncol. Lett. 2017, 14, 3853–3861. [Google Scholar] [CrossRef] [PubMed]
- Manvati, S.; Mangalhara, K.C.; Kalaiarasan, P.; Srivastava, N.; Bamezai, R.N.K. miR-24-2 regulates genes in survival pathway and demonstrates potential in reducing cellular viability in combination with docetaxel. Gene 2015, 567, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Scheibner, K.A.; Teaboldt, B.; Hauer, M.C.; Chen, X.; Cherukuri, S.; Guo, Y.; Kelley, S.M.; Liu, Z.; Baer, M.R.; Heimfeld, S.; et al. MiR-27a Functions as a Tumor Suppressor in Acute Leukemia by Regulating 14-3-3θ. PLoS ONE 2012, 7, e50895. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Dong, L.; Zou, D.; Zhao, H.; Ren, Y.; Li, F.; Yi, P.; Li, L.; Zhu, Y.; Ma, Y.; et al. microRNA-23a, -27a and -24 synergistically regulate JAK1/Stat3 cascade and serve as novel therapeutic targets in human acute erythroid leukemia. Oncogene 2016, 35, 6001–6014. [Google Scholar] [CrossRef]
- Drobna, M.; Szarzyńska-Zawadzka, B.; Daca-Roszak, P.; Kosmalska, M.; Jaksik, R.; Witt, M.; Dawidowska, M. Identification of Endogenous Control miRNAs for RT-qPCR in T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2018, 19, 2858. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Ma, J.; Wu, M.; Qin, W.; Zhao, B.; Shi, Y.; Jin, Y.; Xie, Y. Aberrant microRNA-182 expression is associated with glucocorticoid resistance in lymphoblastic malignancies. Leuk. Lymphoma 2012, 53, 2465–2473. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Shen, T.; Wang, H.; Ke, Z.; Liang, Y.; Ouyang, J.; Jiang, T. MicroRNA-185-5p restores glucocorticoid sensitivity by suppressing the mammalian target of rapamycin complex (mTORC) signaling pathway to enhance glucocorticoid receptor autoregulation. Leuk. Lymphoma 2017, 58, 2657–2667. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Jiang, N.; Huang, G.; Tay, J.L.S.; Lin, B.; Bi, C.; Koh, G.S.; Li, Z.; Tan, J.; Chung, T.H.; et al. Deregulated MIR335 that targets MAPK1 is implicated in poor outcome of paediatric acute lymphoblastic leukaemia. Br. J. Haematol. 2013, 163, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Scarola, M.; Schoeftner, S.; Schneider, C.; Benetti, R. miR-335 directly targets Rb1 (pRb/p105) in a proximal connection to p53-dependent stress response. Cancer Res. 2010, 70, 6925–6933. [Google Scholar] [CrossRef]
- Akbari Moqadam, F.; Lange-Turenhout, E.A.M.; Ariës, I.M.; Pieters, R.; den Boer, M.L. MiR-125b, miR-100 and miR-99a co-regulate vincristine resistance in childhood acute lymphoblastic leukemia. Leuk. Res. 2013, 37, 1315–1321. [Google Scholar] [CrossRef]
- Dou, L.; Zheng, D.; Li, J.; Li, Y.; Gao, L.; Wang, L.; Yu, L. Methylation-mediated repression of microRNA-143 enhances MLL-AF4 oncogene expression. Oncogene 2012, 31, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Song, Y.; Zheng, W.; Ma, W. miRNA143 Induces K562 Cell Apoptosis Through Downregulating BCR-ABL. Med. Sci. Monit. 2016, 22, 2761–2767. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.Z.; Zhang, Y.Y.; Fu, H.Y.; Wu, D.S.; Zhou, H.R. Overexpression of microRNA-143 inhibits growth and induces apoptosis in human leukemia cells. Oncol. Rep. 2014, 31, 2035–2042. [Google Scholar] [CrossRef] [PubMed]
- Volinia, S.; Galasso, M.; Costinean, S.; Tagliavini, L.; Gamberoni, G.; Drusco, A.; Marchesini, J.; Mascellani, N.; Sana, M.E.; Jarour, R.A.; et al. Reprogramming of miRNA networks in cancer and leukemia. Genome Res. 2010, 20, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ma, T.; Huang, C.; Zhang, L.; Xu, T.; Hu, T.; Li, J. MicroRNA-148a: A potential therapeutic target for cancer. Gene 2014, 533, 456–457. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, S.; Tyybäkinoja, A.; Borze, I.; Räty, R.; Saarinen-Pihkala, U.M.; Usvasalo, A.; Elonen, E.; Knuutila, S. Integrated analysis of gene copy number, copy neutral LOH, and microRNA profiles in adult acute lymphoblastic leukemia. Cytogenet. Genome Res. 2012, 136, 246–255. [Google Scholar] [CrossRef]
- Zanette, D.L.; Rivadavia, F.; Molfetta, G.A.; Barbuzano, F.G.; Proto-Siqueira, R.; Falcão, R.P.; Zago, M.A.; Silva, W.A. miRNA expression profiles in chronic lymphocytic and acute lymphocytic leukemia. Braz. J. Med. Biol. Res. 2007, 40, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Kotani, A.; Ha, D.; Hsieh, J.; Rao, P.K.; Schotte, D.; Den Boer, M.L.; Armstrong, S.A.; Lodish, H.F. miR-128b is a potent glucocorticoid sensitizer in MLL-AF4 acute lymphocytic leukemia cells and exerts cooperative effects with miR-221. Blood 2009, 114, 4169–4178. [Google Scholar] [CrossRef] [PubMed]
- Fulci, V.; Colombo, T.; Chiaretti, S.; Messina, M.; Citarella, F.; Tavolaro, S.; Guarini, A.; Foà, R.; Macino, G. Characterization of B- and T-lineage acute lymphoblastic leukemia by integrated analysis of microRNA and mRNA expression profiles. Genes Chromosom. Cancer 2009, 48, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Kotani, A.; Ha, D.; Schotte, D.; Den Boer, M.L.; Armstrong, S.A.; Lodish, H.F. A novel mutation in the miR-128b gene reduces miRNA processing and leads to glucocorticoid resistance of MLL-AF4 acute lymphocytic leukemia cells. Cell Cycle 2010, 9, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Mets, E.; Van Peer, G.; Van der Meulen, J.; Boice, M.; Taghon, T.; Goossens, S.; Mestdagh, P.; Benoit, Y.; De Moerloose, B.; Van Roy, N.; et al. MicroRNA-128-3p is a novel oncomiR targeting PHF6 in T-cell acute lymphoblastic leukemia. Haematologica 2014, 99, 1326–1333. [Google Scholar] [CrossRef] [PubMed]
- Guzman, H.; Sanders, K.; Idica, A.; Bochnakian, A.; Jury, D.; Daugaard, I.; Zisoulis, D.G.; Pedersen, I.M. miR-128 inhibits telomerase activity by targeting TERT mRNA. Oncotarget 2018, 9, 13244–13253. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Yen, C.T.; Pai, C.H.; Chen, H.Y.; Yu, S.L.; Lin, C.Y.; Hu, C.Y.; Jou, S.T.; Lin, D.T.; Lin, S.R.; et al. A double negative loop comprising ETV6/RUNX1 and MIR181A1 contributes to differentiation block in t(12;21)-positive acute lymphoblastic leukemia. PLoS ONE 2015, 10, e0142863. [Google Scholar] [CrossRef] [PubMed]
- Durmaz, B.; Bagca, B.G.; Cogulu, O.; Susluer, S.Y.; Alpay, A.; Aksoylar, S.; Gunduz, C. Antileukemic Effects of Anti-miR-146a, Anti-miR-155, Anti-miR-181a, and Prednisolone on Childhood Acute Lymphoblastic Leukemia. BioMed Res. Int. 2021, 2021, 3207328. [Google Scholar] [CrossRef]
- Schneider, E.; Staffas, A.; Röhner, L.; Krowiorz, K.; Heuser, M.; Döhner, K.; Bullinger, L.; Döhner, H.; Fogelstrand, L.; Rouhi, A.; et al. MicroRNA-155 is upregulated in MLL-rearranged AML but its absence does not affect leukemia development. Exp. Hematol. 2016, 44, 1166–1171. [Google Scholar] [CrossRef]
- Hickey, C.; Schwind, S.; Becker, H.; Alachkar, H.; Garzon, R.; Wu, Y.-Z.; Liu, S.; Perrotti, D.; Marcucci, G. MicroRNA-181a Targets TEL/AML1 Expression and Impairs Cell Proliferation in t(12;21) Acute Lymphocytic Leukemia (ALL) Cells. Blood 2009, 114, 766. [Google Scholar] [CrossRef]
- Dahlhaus, M.; Schult, C.; Lange, S.; Freund, M.; Junghanss, C. MicroRNA 181a influences the expression of HMGB1 and CD4 in acute Leukemias. Anticancer Res. 2013, 33, 445–452. [Google Scholar] [PubMed]
- Havelange, V.; Stauffer, N.; Heaphy, C.C.E.; Volinia, S.; Andreeff, M.; Marcucci, G.; Croce, C.M.; Garzon, R. Functional implications of microRNAs in acute myeloid leukemia by integrating microRNA and messenger RNA expression profiling. Cancer 2011, 117, 4696–4706. [Google Scholar] [CrossRef]
- Markopoulos, G.S.; Roupakia, E.; Tokamani, M.; Alabasi, G.; Sandaltzopoulos, R.; Marcu, K.B.; Kolettas, E. Roles of NF-κB Signaling in the Regulation of miRNAs Impacting on Inflammation in Cancer. Biomedicines 2018, 6, 40. [Google Scholar] [CrossRef]
- Iliopoulos, D.; Jaeger, S.A.; Hirsch, H.A.; Bulyk, M.L.; Struhl, K. STAT3 activation of miR-21 and miR-181b-1 via PTEN and CYLD are part of the epigenetic switch linking inflammation to cancer. Mol. Cell 2010, 39, 493–506. [Google Scholar] [CrossRef]
- Verduci, L.; Azzalin, G.; Gioiosa, S.; Carissimi, C.; Laudadio, I.; Fulci, V.; Macino, G. MicroRNA-181a enhances cell proliferation in acute lymphoblastic leukemia by targeting EGR1. Leuk. Res. 2015, 39, 479–485. [Google Scholar] [CrossRef]
- Zeng, M.N.; Ma, W.L.; Zheng, W.L. [Bioinformatics analysis of microRNA comprehensive regulatory network in B- cell acute lymphoblastic leukemia]. Zhonghua Xue Ye Xue Za Zhi 2016, 37, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Z.; He, C.; Wang, D.; Yuan, X.; Chen, J.; Jin, J. MicroRNAs expression signatures are associated with lineage and survival in acute leukemias. Blood Cells Mol. Dis. 2010, 44, 191–197. [Google Scholar] [CrossRef]
- Han, B.W.; Feng, D.D.; Li, Z.G.; Luo, X.Q.; Zhang, H.; Li, X.J.; Zhang, X.J.; Zheng, L.L.; Zeng, C.W.; Lin, K.Y.; et al. A set of miRNAs that involve in the pathways of drug resistance and leukemic stem-cell differentiation is associated with the risk of relapse and glucocorticoid response in childhood ALL. Hum. Mol. Genet. 2011, 20, 4903–4915. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.S.; Costa e Silva, M.; Coutinho, L.L.; Garcia Gomes, R.; Pedrosa, F.; Massaro, J.D.; Donadi, E.A.; Lucena-Silva, N. MicroRNA expression profiles discriminate childhood T- from B-acute lymphoblastic leukemia. Hematol. Oncol. 2019, 37, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Suresh, S.; McCallum, L.; Lu, W.; Lazar, N.; Perbal, B.; Irvine, A.E. MicroRNAs 130a/b are regulated by BCR-ABL and downregulate expression of CCN3 in CML. J. Cell Commun. Signal. 2011, 5, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Medina, R.; Zaidi, S.K.; Liu, C.G.; Stein, J.L.; VanWijnen, A.J.; Croce, C.M.; Stein, G.S. MicroRNAs 221 and 222 bypass quiescence and compromise cell survival. Cancer Res. 2008, 68, 2773–2780. [Google Scholar] [CrossRef]
- Zhang, C.; Kang, C.; Wang, P.; Cao, Y.; Lv, Z.; Yu, S.; Wang, G.; Zhang, A.; Jia, Z.; Han, L.; et al. MicroRNA-221 and -222 regulate radiation sensitivity by targeting the PTEN pathway. Int. J. Radiat. Oncol. Biol. Phys. 2011, 80, 240–248. [Google Scholar] [CrossRef]
- Wang, M.; Ge, X.; Zheng, J.; Li, D.; Liu, X.; Wang, L.; Jiang, C.; Shi, Z.; Qin, L.; Liu, J.; et al. Role and mechanism of miR-222 in arsenic-transformed cells for inducing tumor growth. Oncotarget 2016, 7, 17805–17814. [Google Scholar] [CrossRef] [PubMed]
- Coskun, E.; Neumann, M.; Schlee, C.; Liebertz, F.; Heesch, S.; Goekbuget, N.; Hoelzer, D.; Baldus, C.D. MicroRNA profiling reveals aberrant microRNA expression in adult ETP-ALL and functional studies implicate a role for miR-222 in acute leukemia. Leuk. Res. 2013, 37, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Gao, C.; Wang, K.; Cui, L.; Li, W.; Zhao, X.; Liu, F.; Wu, M.; Deng, G.; Ding, W.; et al. Effect of microRNA-210 on prognosis and response to chemotherapeutic drugs in pediatric acute lymphoblastic leukemia. Cancer Sci. 2014, 105, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.J.; Croce, C.M. MicroRNAs play a central role in molecular dysfunctions linking inflammation with cancer. Immunol. Rev. 2013, 253, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Yan, J.; Liu, C.; Zhou, H.; Zheng, Y. Tumor Suppressor Role of miR-363-3p in Gastric Cancer. Med. Sci. Monit. 2015, 21, 4074–4080. [Google Scholar] [CrossRef] [PubMed]
- Busche, S.; Ge, B.; Vidal, R.; Spinella, J.F.; Saillour, V.; Richer, C.; Healy, J.; Chen, S.H.; Droit, A.; Sinnett, D.; et al. Integration of high-resolution methylome and transcriptome analyses to dissect epigenomic changes in childhood acute lymphoblastic leukemia. Cancer Res. 2013, 73, 4323–4336. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Li, L.; Li, S. Circulating microRNA-21 as a biomarker for the detection of various carcinomas: An updated meta-analysis based on 36 studies. Tumour Biol. 2015, 36, 1973–1981. [Google Scholar] [CrossRef] [PubMed]
- Seca, H.; Lima, R.; Lopes-Rodrigues, V.; Guimaraes, J.; Gabriela, G.; Vasconcelos, M. Targeting miR-21 induces autophagy and chemosensitivity of leukemia cells. Curr. Drug Targets 2013, 14, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.H.; Yang, L.; Liu, X.J.; Nie, Z.Y.; Luo, J.M. [Targeted suppression of miRNA-21 inhibit K562 cells growth through PTEN-PI3K/AKT signaling pathway]. Zhonghua Xue Ye Xue Za Zhi 2016, 37, 982–986. [Google Scholar] [CrossRef]
- Bai, H.; Xu, R.; Cao, Z.; Wei, D.; Wang, C. Involvement of miR-21 in resistance to daunorubicin by regulating PTEN expression in the leukaemia K562 cell line. FEBS Lett. 2011, 585, 402–408. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, X.; Li, X.; Zhang, L.; Li, L.; Sun, Z.; Fu, X.; Wu, J.; Chang, Y.; Li, W.; et al. Effects of microRNA-21 on the biological functions of T-cell acute lymphoblastic lymphoma/leukemia. Oncol. Lett. 2016, 12, 4173–4180. [Google Scholar] [CrossRef]
- Aster, J.C. Dicing up T-ALL. Blood 2015, 126, 929–930. [Google Scholar] [CrossRef][Green Version]
- Starczynowski, D.T.; Morin, R.; McPherson, A.; Lam, J.; Chari, R.; Wegrzyn, J.; Kuchenbauer, F.; Hirst, M.; Tohyama, K.; Humphries, R.K.; et al. Genome-wide identification of human microRNAs located in leukemia-associated genomic alterations. Blood 2011, 117, 595–607. [Google Scholar] [CrossRef]
- Cui, S.Y.; Wang, R.; Chen, L.B. MicroRNA-145: A potent tumour suppressor that regulates multiple cellular pathways. J. Cell. Mol. Med. 2014, 18, 1913–1926. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, M.; Mo, Y.Y. miR-145-mediated suppression of cell growth, invasion and metastasis. Am. J. Transl. Res. 2010, 2, 170. [Google Scholar] [PubMed]
- Almeida, M.I.; Calin, G.A. The miR-143/miR-145 cluster and the tumor microenvironment: Unexpected roles. Genome Med. 2016, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, Q.; Zhang, H.M.; Liu, W.; Liu, C.J.; Li, Q.; Guo, A.Y. Transcription factor and miRNA co-regulatory network reveals shared and specific regulators in the development of B cell and T cell. Sci. Rep. 2015, 5, 15215. [Google Scholar] [CrossRef] [PubMed]
- Labbaye, C.; Spinello, I.; Quaranta, M.T.; Pelosi, E.; Pasquini, L.; Petrucci, E.; Biffoni, M.; Nuzzolo, E.R.; Billi, M.; Foà, R.; et al. A three-step pathway comprising PLZF/miR-146a/CXCR4 controls megakaryopoiesis. Nat. Cell Biol. 2008, 10, 788–801. [Google Scholar] [CrossRef]
- Tong, L.G.; Wu, W.Z.; Zhang, Y.P.; Zhou, Z.G.; Chen, Y.F.; Huang, W.J.; Xu, H.; Su, Q.Q. [Expression of miR-16 in patients with T lymphoblastic lymphoma/acute lymphoblastic leukemia]. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2014, 22, 99–103. [Google Scholar] [CrossRef]
- Azimi, A.; Hagh, M.F.; Yousefi, B.; Rahnama, M.A.; Khorrami, A.; Heydarabad, M.Z.; Najafpour, M.; Hallajzadeh, J.; Ghahremani, A. The Effect of Prednisolone on miR 15a and miR16-1 Expression Levels and Apoptosis in Acute Lymphoblastic Leukemia Cell Line: CCRF-CEM. Drug Res. 2016, 66, 432–435. [Google Scholar] [CrossRef]
- Kaddar, T.; Chien, W.W.; Bertrand, Y.; Pages, M.P.; Rouault, J.P.; Salles, G.; Ffrench, M.; Magaud, J.P. Prognostic value of miR-16 expression in childhood acute lymphoblastic leukemia relationships to normal and malignant lymphocyte proliferation. Leuk. Res. 2009, 33, 1217–1223. [Google Scholar] [CrossRef]
- Li, J.; Li, P.; Wang, J.; Xi, Y.X. [Significance of microRNA-16 and bcl-2 expression in T lymphoblastic lymphoma/leukemia and its relation with prognosis]. Zhonghua Bing Li Xue Za Zhi 2013, 42, 748–752. [Google Scholar] [CrossRef]
- McBride, A.; Houtmann, S.; Wilde, L.; Vigil, C.; Eischen, C.M.; Kasner, M.; Palmisiano, N. The role of inhibition of apoptosis in acute leukemias and myelodysplastic syndrome. Front. Oncol. 2019, 9, 192. [Google Scholar] [CrossRef]
- Xi, Y.; Li, J.; Zan, L.; Wang, J.; Wang, G.; Ning, Y. Micro-RNA-16 expression in paraffin-embedded specimen correlates with overall survival of T-lymphoblastic lymphoma/leukemia. Hum. Pathol. 2013, 44, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Mavrakis, K.J.; Wolfe, A.L.; Oricchio, E.; Palomero, T.; De Keersmaecker, K.; McJunkin, K.; Zuber, J.; James, T.; Chang, K.; Khan, A.A.; et al. Genome-wide RNA-mediated interference screen identifies miR-19 targets in Notch-induced T-cell acute lymphoblastic leukaemia. Nat. Cell Biol. 2010, 12, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Slovak, M.L.; Mannoor, K.; Chen, C.; Hunger, S.P.; Carroll, A.J.; Schultz, R.A.; Shaffer, L.G.; Ballif, B.C.; Ning, Y. Microarray detection of multiple recurring submicroscopic chromosomal aberrations in pediatric T-cell acute lymphoblastic leukemia. Leukemia 2011, 25, 1042–1046. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mesrian Tanha, H.; Mojtabavi Naeini, M.; Rahgozar, S.; Moafi, A.; Honardoost, M.A. Integrative computational in-depth analysis of dysregulated miRNA-mRNA interactions in drug-resistant pediatric acute lymphoblastic leukemia cells: An attempt to obtain new potential gene-miRNA pathways involved in response to treatment. Tumor Biol. 2016, 37, 7861–7872. [Google Scholar] [CrossRef]
- Malouf, C.; Antunes, E.T.B.; O’Dwyer, M.; Jakobczyk, H.; Sahm, F.; Landua, S.L.; Anderson, R.A.; Soufi, A.; Halsey, C.; Ottersbach, K. miR-130b and miR-128a are essential lineage-specific codrivers of t(4;11) MLL-AF4 acute leukemia. Blood 2021, 138, 2066–2092. [Google Scholar] [CrossRef]
- Haque, S.; Vaiselbuh, S.R. Silencing of exosomal miR-181a reverses pediatric acute lymphocytic leukemia cell proliferation. Pharmaceuticals 2020, 13, 241. [Google Scholar] [CrossRef]
- Longjohn, M.N.; Squires, W.R.B.; Christian, S.L. Meta-analysis of microRNA profiling data does not reveal a consensus signature for B cell acute lymphoblastic leukemia. Gene 2022, 821, 146211. [Google Scholar] [CrossRef]
- Shi, L.H.; Tian, L.; Wang, Y.F.; Liu, J.S.; Guo, M.F.; Liu, W. Effects of MiR-221-Mediated Wnt/β- Catenin Signaling Pathway on Biological Activity of Childhood Acute Lymphoblastic Leukemia Cells. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2019, 27, 1367–1373. [Google Scholar] [CrossRef]
- De Leeuw, D.C.; Van Den Ancker, W.; Denkers, F.; De Menezes, R.X.; Westers, T.M.; Ossenkoppele, G.J.; Van De Loosdrecht, A.A.; Smit, L. MicroRNA profiling can classify acute leukemias of ambiguous lineage as either acute myeloid leukemia or acute lymphoid leukemia. Clin. Cancer Res. 2013, 19, 2187–2196. [Google Scholar] [CrossRef]
- Mraz, M.; Pospisilova, S.; Malinova, K.; Slapak, I.; Mayer, J. MicroRNAs in chronic lymphocytic leukemia pathogenesis and disease subtypes. Leuk. Lymphoma 2009, 50, 506–509. [Google Scholar] [CrossRef]
- Alsuwaidi, L.; Hachim, M.; Senok, A. Novel Markers in Pediatric Acute Lymphoid Leukemia: The Role of ADAM6 in B Cell Leukemia. Front. Cell Dev. Biol. 2021, 9, 1680. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Zhou, Y.; Zhou, D.; Zhang, Y.; Shang, D.; Qi, J. The latest progress on miR-374 and its functional implications in physiological and pathological processes. J. Cell. Mol. Med. 2019, 23, 3063–3076. [Google Scholar] [CrossRef] [PubMed]
- Ohyashiki, J.H.; Umezu, T.; Kobayashi, C.; Hamamura, R.S.; Tanaka, M.; Kuroda, M.; Ohyashiki, K. Impact on cell to plasma ratio of miR-92a in patients with acute leukemia: In vivo assessment of cell to plasma ratio of miR-92a. BMC Res. Notes 2010, 3, 347. [Google Scholar] [CrossRef] [PubMed]
- Git, A.; Dvinge, H.; Salmon-Divon, M.; Osborne, M.; Kutter, C.; Hadfield, J.; Bertone, P.; Caldas, C. Systematic comparison of microarray profiling, real-time PCR, and next-generation sequencing technologies for measuring differential microRNA expression. RNA 2010, 16, 991. [Google Scholar] [CrossRef] [PubMed]
- Dzikiewicz-Krawczyk, A.; Macieja, A.; Mały, E.; Januszkiewicz-Lewandowska, D.; Mosor, M.; Fichna, M.; Strauss, E.; Nowak, J. Polymorphisms in microRNA target sites modulate risk of lymphoblastic and myeloid leukemias and affect microRNA binding. J. Hematol. Oncol. 2014, 7, 43. [Google Scholar] [CrossRef]
- GeneCards—Human Genes | Gene Database | Gene Search. Available online: https://www.genecards.org/ (accessed on 23 February 2021).
- Kovaleva, V.; Mora, R.; Park, Y.J.; Plass, C.; Chiramel, A.I.; Bartenschlager, R.; Doḧner, H.; Stilgenbauer, S.; Pscherer, A.; Lichter, P.; et al. miRNA-130a targets ATG2B and DICER1 to inhibit autophagy and trigger killing of chronic lymphocytic leukemia cells. Cancer Res. 2012, 72, 1763–1772. [Google Scholar] [CrossRef]
- Jiang, L.H.; Zhang, H.D.; Tang, J.H. MiR-30a: A Novel Biomarker and Potential Therapeutic Target for Cancer. J. Oncol. 2018, 2018, 5167829. [Google Scholar] [CrossRef]
- Ortega, M.; Bhatnagar, H.; Lin, A.P.; Wang, L.; Aster, J.C.; Sill, H.; Aguiar, R.C.T. A microRNA-mediated regulatory loop modulates NOTCH and MYC oncogenic signals in B- and T-cell malignancies. Leukemia 2015, 29, 968–976. [Google Scholar] [CrossRef]
- Mukherjee, S.; Shelar, B.; Krishna, S. Versatile role of miR-24/24-1*/24-2* expression in cancer and other human diseases. Am. J. Transl. Res. 2022, 14, 20. [Google Scholar]
- Yang, H.; Zhang, J.; Li, J.; Zhao, F.; Shen, Y.; Xing, X. Overexpression of miR-574-3p suppresses proliferation and induces apoptosis of chronic myeloid leukemia cells via targeting IL6/JAK/STAT3 pathway. Exp. Ther. Med. 2018, 16, 4296–4302. [Google Scholar] [CrossRef]
- Fu, A.; Hoffman, A.E.; Liu, R.; Jacobs, D.I.; Zheng, T.; Zhu, Y. Targetome profiling and functional genetics implicate miR-618 in lymphomagenesis. Epigenetics 2014, 9, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.M.; Brown, L.M.; Ryland, G.; Lonsdale, A.; Kosasih, H.J.; Ludlow, L.E.; Majewski, I.J.; Blombery, P.; Ekert, P.G.; Davidson, N.; et al. ALLSorts: A RNA-Seq subtype classifier for B-Cell Acute Lymphoblastic Leukemia. Blood Adv. 2022, 6, 4093–4097. [Google Scholar] [CrossRef] [PubMed]
- Bařinka, J.; Hu, Z.; Wang, L.; Wheeler, D.A.; Rahbarinia, D.; McLeod, C.; Gu, Z.; Mullighan, C.G. RNAseqCNV: Analysis of large-scale copy number variations from RNA-seq data. Leukemia 2022, 36, 1492–1498. [Google Scholar] [CrossRef]
- Li, X.; Sanda, T.; Thomas Look, A.; Novina, C.D.; von Boehmer, H. Repression of tumor suppressor miR-451 is essential for NOTCH1-induced oncogenesis in T-ALL. J. Exp. Med. 2011, 208, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Akbari Moqadam, F.; Boer, J.M.; Lange-Turenhout, E.A.M.; Pieters, R.; Den Boer, M.L. Altered expression of miR-24, miR-126 and miR-365 does not affect viability of childhood TCF3-rearranged leukemia cells. Leukemia 2014, 28, 1008–1014. [Google Scholar] [CrossRef]
- Diakos, C.; Zhong, S.; Xiao, Y.; Zhou, M.; Vasconcelos, G.M.; Krapf, G.; Yeh, R.F.; Zheng, S.; Kang, M.; Wiencke, J.K.; et al. TEL-AML1 regulation of survivin and apoptosis via miRNA-494 and miRNA-320a. Blood 2010, 116, 4885–4893. [Google Scholar] [CrossRef]
- Scherr, M.; Elder, A.; Battmer, K.; Barzan, D.; Bomken, S.; Ricke-Hoch, M.; Schröder, A.; Venturini, L.; Blair, H.J.; Vormoor, J.; et al. Differential expression of miR-17∼92 identifies BCL2 as a therapeutic target in BCR-ABL-positive B-lineage acute lymphoblastic leukemia. Leukemia 2014, 28, 554–565. [Google Scholar] [CrossRef]
- Agueli, C.; Cammarata, G.; Salemi, D.; Dagnino, L.; Nicoletti, R.; La Rosa, M.; Messana, F.; Marfia, A.; Bica, M.G.; Coniglio, M.L.; et al. 14q32/miRNA clusters loss of heterozygosity in acute lymphoblastic leukemia is associated with up-regulation of BCL11a. Am. J. Hematol. 2010, 85, 575–578. [Google Scholar] [CrossRef]
- Amankwah, E.K.; Devidas, M.; Teachey, D.T.; Rabin, K.R.; Brown, P.A. Six Candidate miRNAs Associated With Early Relapse in Pediatric B-Cell Acute Lymphoblastic Leukemia. Anticancer Res. 2020, 40, 3147–3153. [Google Scholar] [CrossRef]
- Gutierrez-Camino, A.; Richer, C.; St-Onge, P.; Lopez-Lopez, E.; Bañeres, A.C.; De Andoin, N.G.; Sastre, A.; Astigarraga, I.; Martin-Guerrero, I.; Sinnett, D.; et al. Role of rs10406069 in miR-5196 in hyperdiploid childhood acute lymphoblastic leukemia. Epigenomics 2020, 12, 1949–1955. [Google Scholar] [CrossRef]
- Piroozian, F.; Bagheri Varkiyani, H.; Koolivand, M.; Ansari, M.; Afsa, M.; AtashAbParvar, A.; MalekZadeh, K. The impact of variations in transcription of DICER and AGO2 on exacerbation of childhood B-cell lineage acute lymphoblastic leukaemia. Int. J. Exp. Pathol. 2019, 100, 184–191. [Google Scholar] [CrossRef] [PubMed]



| Study | Cohort Characteristics | Methods | Outcomes |
|---|---|---|---|
| Sun et al., 2021 [25] | 43 childhood ALL patients and 14 controls; validation: 28 childhood ALL patients and 28 controls; China | qRT-PCR and microarray; BM | Significant upregulation of miR-155-5p in ALL samples compared with controls (p < 0.05). |
| Zamani et al., 2021 [26] | 59 children with ALL at diagnosis and 50 non-cancer controls; Iran | qRT-PCR; BM | Significantly lower miR-324-3p and miR-508-5p expression in children with ALL (p < 0.0001 and p < 0.005, respectively). ROC analysis evaluated the utilization of miR-324-3p (AUC = 0.73; sensitivity, 44% (95% CI: 29.99 to 58.75); specificity, 100% (95% CI: 92.75 to 100); cutoff value, 0.9506; p < 0.0001) and miR-508-5p (AUC = 0.664; sensitivity, 40%; 95% CI: 26.41 to 54.82; specificity, 95.92%; 95% CI: 86.02 to 99.5; cutoff value, 0.1812; p = 0.005) as potential diagnostic biomarkers in pediatric ALL. |
| Liang et al., 2021 [27] | 42 childhood ALL patients at diagnosis and controls from GEO databases GSE56489 (43 children with ALL and 14 controls) [28] and GSE23024 (81 children with ALL and 7 controls) [29]; China | qRT-PCR and microarray; BM | Differentially expressed miR-155 and miR-199b were documented in all three datasets. Analysis of datasets showed several other differentially expressed miRNAs. With respect to the GSE56489 dataset, significant differential expression of miR-606, miR-640, miR-199b-3p, miR-145, miR-297, miR-181a*, miR-1322, miR-155, miR-146a, miR-587, miR-1323, let-7b*, miR-548i, miR-3121, miR-449a, miR-369-3p, miR-708 and miR-181b was observed in ALL patients compared to controls (p < 0.05). With respect to the GSE23024 dataset, significant differential expression of miR-199b, miR-432, miR-224, miR-376a, miR-148a, miR-485-3p, miR-411, miR-382, miR-503, miR-223, miR-199a, miR-450, miR-338, miR-424, miR-145, miR-143, miR-618, miR-128b, miR-363, miR-497, miR-153, miR-659, miR-34a, miR-181b, miR-181d, miR-181c, miR-130a, miR-210, miR-130b, miR-579 and miR-155 was observed in children with ALL compared with healthy children (p < 0.05). |
| El-maadawy et al., 2021 [30] | 43 childhood ALL patients and 42 controls; Egypt | qRT-PCR; PBMCs | Significant elevation in miR-21 (p < 0.05), miR-148a (p < 0.01) and miR-24 (p < 0.05) expression levels in ALL patients compared to controls. No significant change in miR-26a and miR-133b levels. ROC curve analysis demonstrated the highest AUC for miR-24, followed by miR148a and miR-133b (0.785, 0.719 and 0.669, respectively). No significant results with respect to miR-21 and miR-26 ROC curves. At a cutoff value of 2.928, miR-24 showed 72% sensitivity and 81% specificity (p < 0.001) in detecting ALL cases. Compared to healthy subjects, ALL patients showed a positive correlation between miR-148a and miR-24 (r = 0.347; p < 0.05). Another positive correlation was found between miR-26a and miR-24 (r = 0.353; p < 0.05) in the ALL group compared to the control group. |
| Zhu et al., 2021 [31] | 132 childhood ALL patients and 80 controls; China | qRT-PCR; serum | Significantly higher expression levels of serum miR-922 and miR-506 in children with ALL than controls (p < 0.001). ROC curve analysis reported optimal cutoff values of miR-922 and miR-506 for the diagnosis of childhood ALL of 1.46 and 2.17, respectively. |
| Fayed et al., 2021 [32] | 71 childhood ALL patients (32 newly diagnosed, 21 relapsed and 18 remitted) and 30 controls; Egypt | qRT-PCR; plasma | AUC of miR-92a (0.755; cutoff 8.77) with 41.5% sensitivity and 100% specificity, whereas miR-638 showed an AUC of 0.862 and sensitivity and specificity of 54.7% and 100%, respectively (cutoff, 6.79). Levels of miR-92a and miR-638, as well as the miR-92a/miR-638 ratio, were significantly higher (17.89, 10.19 and 1.75-fold, respectively) in children newly diagnosed with ALL compared to controls. There was a significant positive correlation of miR-92a and miR-638 levels in children with ALL (r = 0.955; p ˂ 0.0001). |
| Li Shao-Wu et al., 2020 [33] | 130 newly diagnosed children with T-ALL and 50 controls; China | qRT-PCR; PBMCs | Expression of miR-146a and miR-221 in T-ALL subjects was significantly higher than in controls (5.83 ± 1.54 vs. 0.96 ± 0.17 and 7.13 ± 2.6 vs. 1.64 ± 0.51, respectively; p < 0.01). Diagnostic cutoff values of miR-146a and miR-221 in childhood T-ALL were determined to be 3.9 and 5.28, respectively, by means of ROC curve analysis. The AUC of T-ALL jointly diagnosed by miR-146a and miR-221 was 0.835 (95% CI: 0.764 to 0.892) with high sensitivity (85%) and specificity (77.2%). In addition, levels of miR-146a were positively correlated with miR-221 levels at diagnosis (r = 0.784, p < 0.01). |
| Li Chunyu et al., 2020 [34] | 59 childhood ALL patients at diagnosis and 30 controls; China | qRT-PCR; BM | Expression of miR-223 was markedly reduced in patients with ALL compared with controls (p < 0.001). The ROC curve confirmed the diagnostic value of miR-223 in ALL: AUC, 0.978 ± 0.013; sensitivity, 93.22%; and specificity, 93.33% (cutoff, 0.705; p < 0.0001). miR-223 seems to inhibit cell proliferation, migration and invasion and to promote apoptosis by targeting FOXO1 (which was upregulated in ALL patients). |
| Shafik et al., 2020 [35] | 70 childhood ALL patients at diagnosis and 7 controls; Egypt | qRT-PCR; BM | The expression of miR-181a was statistically significantly elevated in ALL patients compared to controls (p < 0.001). However, miR-196b expression was significantly downregulated in ALL patients compared to controls (p = 0.038). High expression of miR-181a was reported in 68 out of 70 ALL patients (97.1%; cutoff, 0.015), whereas 49 of 56 children with ALL (87.5%; cutoff, 0.001) were found to exhibit low miR-196b expression. A significant positive correlation was observed between miR-181a and miR-196b expression levels (r = 0.344; p = 0.009). |
| Akpinar et al., 2020 [36] | 13 childhood ALL patients at diagnosis and 5 controls; Turkey | qRT-PCR; whole blood | Significant downregulation of miR-375 and upregulation of miR-21, miR-222, miR-30, miR-145, miR-146a and miR-155 levels in ALL patients compared with controls. |
| Chen et al., 2020 [37] | 42 childhood T-ALL patients (21 primary and 21 recurrent) and 20 controls; China | qRT-PCR; PBMCs | Significant downregulation of miR-335-3p in T-ALL patients compared with controls (p < 0.05). |
| N. Hassan et al., 2020 [38] | 85 childhood ALL patients at diagnosis and 12 controls; Egypt | qRT-PCR; BM | Expression of miR-100 was significantly downregulated in ALL patients compared to controls (p = 0.035). Expression of miR-210 was significantly upregulated in ALL patients compared to controls (p = 0.025). ROC curve analysis revealed an AUC 0.642 for miR-100 (95% CI: 0.519 to 0.764; cutoff, 2.6; 64.7% sensitivity and 62.5% specificity), whereas the AUC for miR-210 was 0.65 (95% CI: 0.511 to 0.79; cutoff, 3.5; 60% sensitivity and 58.3% specificity). |
| S.S. Hassan et al., 2020 [39] | 60 childhood ALL patients (30 HCV and 30 non-HCV) at diagnosis, 10 controls with HCV and 2 healthy controls; Egypt | qRT-PCR; PBMCs | Hepatitis C virus genotype 4 (HCV-4)-associated ALL cases displayed a 3-fold increase in the expression of miR-155 compared to children with chronic HCV infection (47.8 vs. 16.2), suggesting it as a therapeutic target in the respective cases. In addition, ALL patients seemed to express higher levels of miR-155, regardless of whether they were HCV-infected or not. |
| Al Nakeeb et al., 2020 [40] | 20 childhood ALL patients and 30 controls; Iraq | qRT-PCR; serum | Expression levels of miR-142-3p and miR-146a-3p were significantly downregulated, whereas miR-223-3p expression was significantly upregulated in ALL patients compared with healthy children. |
| Dawidowska et al., 2019 [10] | 34 childhood T-ALL patients and 5 controls; Poland | RNA-seq and qRT-PCR; BM | Significant overexpression of miR-548a-3p, miR-128-3p, miR-181b-5p, miR-20b-5p, miR-6500-3p, miR-331-5p, miR-363-3p, miR-153-3p, miR-466 and miR-130a-3p (p < 0.01; also miR-20b-3p, miR-210-3p, miR-181a-3p, miR-4421, miR-18b-5p, miR-181a-2-3p, miR-181a-5p, miR-625-3p, miR-130b-3p, miR-4687-5p, miR-4437, miR-625-5p and miR-3609 with 0.01 ≤ p < 0.05) in T-ALL samples compared with controls. Significantly lower levels of miR-574-5p, miR-10a-5p, miR-582-3p, miR-143-3p, miR-941, miR-145-5p, miR-27a-5p, miR-618, miR-24-2-3p, miR-145-3p, miR-504-5p, miR-3690, miR-223-5p, miR-199b-5p, miR-550a-5p, miR-4695-3p, miR-30a-5p, miR-3909, miR-2115-3p, miR-582-5p, miR-504-3p, miR-23a-5p, miR-10b-5p, miR-4494 and miR-151a-3p (p < 0.01; also miR-223-3p, miR-6865-3p, miR-7849-3p, miR-1275, miR-338-5p, miR-3150b-5p, miR-3154, miR-6823-5p, miR-10a-3p, miR-143-5p and miR-4745-3p with 0.01 < p < 0.05) in T-ALL samples compared with controls. |
| Sheybani et al., 2019 [41] | 27 childhood B-ALL patients (non-Ph-positive; 19 at diagnosis and 8 at relapse) and 16 controls; Iran | qRT-PCR; BM | Significant downregulation of miR-326 was noted in B-ALL patients compared with controls. |
| Xue et al., 2019 [42] | 831 childhood ALL patients and 1079 controls; validation: 88 childhood ALL cases and 99 controls; China | qRT-PCR; whole blood; plasma from validation cohort | Expression levels of miR-100 in plasma of childhood ALL cases were 3.25-fold higher than in controls (p < 0.001). |
| Pouyanrad et al., 2019 [43] | 64 childhood ALL patients (46 at diagnosis and 18 at relapse) and 30 controls; Iran | qRT-PCR; BM | Expression of miR-335-3p was found to be significantly downregulated in children with ALL compared with controls (0.33 ± 0.04 vs. 0.7325 ± 0.13; p = 0.005). Moreover, miR-335-3p was noticeably downregulated in relapsed patients (0.08 ± 0.02) compared with controls (0.73 ± 0.13; p = 0.0002), and miR-335-3p levels were also lower in newly diagnosed ALL patients compared with controls (p = 0.018). |
| El-Khazragy et al., 2019a [44] | 45 childhood ALL patients and 10 controls; Egypt | qRT-PCR; BM | Expression levels of miR-155 and miR-181a were significantly higher in ALL patients compared to controls (p < 0.01). |
| El-Khazragy et al., 2019b [45] | 120 childhood ALL patients at diagnosis and 30 controls; Egypt | qRT-PCR; PBMCs and BM | Pediatric ALL samples exhibited significantly lower expression of miR-125b (p < 0.001) compared with controls. In contrast, Bcl-2 expression levels were significantly higher in ALL patients compared with controls (p < 0.001). ROC curve analysis illustrated that miR-125b and Bcl-2 expression could be potential biomarkers for discrimination children with ALL from healthy control, with an AUC 0.99, 0.9 reciprocally. However, miR-125b was superior to Bcl-2: miR-125b sensitivity 98.3% and specificity 96.7% (cutoff ≤ 1.5); Bcl-2 sensitivity 96.6% and specificity 70% (cutoff > 2.14). |
| Rzepiel et al., 2019 [46] | 20 childhood BCP-ALL patients (15 at diagnosis and 5 at relapse) and 10 controls; Hungary | qRT-PCR and TLDA; plasma and BM | Various circulating miRNAs in plasma displayed significantly different expressions between BCP-ALL patients and controls: upregulation of miR-34a-5p, miR-128-3p, miR-146a-5p, miR-155-5p, miR-181a-5p, miR-181b-5p, miR-181c-5p, miR-222-3p, miR-532-5p (p < 0.001), miR-21-5p, miR-92a-3p, miR-125b-5p, miR-320a, miR-361-3p, miR-660-5p (p < 0.01), miR-16, miR-30d-5p and miR-93-5p (p < 0.05); downregulation of miR-223-3p, miR-494-3p (p < 0.001) and miR-374a-5p (p < 0.05). There was no significant correlation between miR expression levels in bone marrow and peripheral blood. |
| Jemimah Devanandan et al., 2019 [47] | 71 childhood ALL patients and 74 controls; India | qRT-PCR; whole blood | Downregulation of miR-146a in ALL patients compared to controls, although not significant. |
| Wang et al., 2019 [48] | 28 childhood ALL patients and 10 controls; China | qRT-PCR; BM | The mRNA expression of miR-146a was significantly increased in children with ALL compared with controls (p < 0.05). |
| Liu et al., 2019 [49] | 30 childhood T-ALL patients and 30 controls; China | qRT-PCR; whole blood | Significantly higher expression of miR-663b in children with T-ALL compared with controls (p < 0.01). |
| Piatopoulou et al., 2018 [50] | 125 childhood ALL cases at diagnosis and 64 controls; Greece | qRT-PCR; BM | Levels of miR-143 and miR-182 were significantly lower in ALL patients compared to controls (p < 0.001). Univariate logistic regression for miR-143 (OR, 0.108; 95% CI: 0.056 to 0.207; p < 0.001) and miR-182 (OR, 0.221; 95% CI: 0.126 to 0.386; p < 0.001) and ROC curve analysis revealed a strong clinical significance of both miR-143 and miR-182 in differential diagnosis of childhood ALL (AUC, 0.88; 95% CI: 0.831 to 0.928; p < 0.001 and AUC, 0.762; 95% CI: 0.694 to 0.829; p < 0.001; respectively). Multivariate logistic regression analysis highlighted that lower levels of miR-143 (OR, 0.115; 95% CI: 0.054 to 0.245; p < 0.001) can discriminate leukemic from normal BM specimens, independently of patient age and gender. |
| Swellam et al., 2018 [51] | 43 childhood ALL patients at diagnosis and 23 controls; Egypt | qRT-PCR; whole blood | Expression levels of miR-125b-1 were 32.6-fold higher in ALL cases as compared to controls (median: 66.67 vs. 2.04; p < 0.0001), whereas expression of miR-203 was 30.76-fold higher in controls than in ALL patients (median: 20.34 vs. 0.66; p < 0.0001). ROC analysis for miR-203 displayed better sensitivity scores than miR-125b-1: (a) miR-203: AUC, 0.874; 95% CI: 0.769 to 0.942; p < 0.0001; cutoff 0.973; 97.7% sensitivity; 87% specificity, (b) miR-125b-1: AUC, 0.858; 95% CI: = 0.715 to 0.847; p < 0.0001; cutoff 3.209; 83.7% sensitivity; 100% specificity; (c) miR-125b-1 combined with miR-203: 100% sensitivity; 87% specificity; 93.5% positive predictive value; 100% negative predictive value; 95.5% accuracy. These two miRNA expression levels were negatively correlated (r = −0.302; p = 0.014). |
| Ghodousi et al., 2018 [52] | 46 childhood ALL patients at diagnosis and 16 controls; Iran | qRT-PCR; BM | Both miR-326 and miR-200c expression levels were significantly lower in ALL patients than controls (p = 0.0002 and 0.035, respectively). ROC curve analysis revealed a high AUC for miR-326 (0.813; 95% CI: 0.671 to 0.954; cutoff, 0.29; sensitivity, 83.3%; and specificity, 70.8%; p < 0.001), whereas the AUC for miR-200c was 0.79 (95% CI: 0.649 to 0.932; cutoff, 0.42; sensitivity, 66.7%; and specificity, 71.4%; p = 0.004). |
| Shafik et al., 2018 [53] | 70 childhood ALL patients at diagnosis and 7 controls; Egypt | qRT-PCR; BM | Expression of miR-128 was significantly elevated in ALL patients compared with controls (p < 0.001). |
| Jiang et al., 2018 [54] | 92 children with ALL (both at diagnosis and relapsed) and 3 controls; China | qRT-PCR; BM and plasma | Expression of miR-652-3p was significantly lower in ALL patients at diagnosis compared with healthy controls (p < 0.05). In the same study, a total of 45 miRNAs were documented as significantly downregulated in newly diagnosed patients compared with controls (i.e., 3661.7-fold downregulation of miR-22 in ALL patients compared to controls; p = 00002): miR-22, miR-624, miR-29a*, miR-183, miR-190b, miR-339-5p, miR-501-3p, miR-200b, miR-195, mmu-miR-140, miR-487b, miR-301, miR-200c, miR-106b, miR-502-3p, miR-18a, miR-27a, SNORD48 X2, miR-142-5p, miR-142-3p, miR-15a*, miR-196b, miR-340*, miR-183*, miR-191*, miR-205, miR-370, miR-29a, miR-532-3p, miR-532, miR-324-3p, miR-20b, miR-138, miR-26b, miR-433, miR-550, miR-19b, miR-136*, miR-1290, miR-374, miR-1180, miR-642, miR-10b, mmu-miR-93, miR-15b, miR-20a*, miR-500, miR-223, miR-652, miR-20a, miR-27b, miR-19a, dme-miR-7, miR-22*, miR-30c, miR-30b and miR-769-5p. |
| Zang et al., 2018 [55] | 81 childhood ALL patients and 83 controls; China | qRT-PCR; whole blood | Significant downregulation of miR-9 in T-cells obtained from ALL patients relative to controls (p < 0.05). Induced overexpression of miR-9 inhibited ALL development in vitro via its downstream target, neuropilin-1 (NRP1). |
| Asnafi et al., 2017 [56] | 41 children with ALL and 41 controls; Iran | qRT-PCR; BM and whole blood | Expression of miR-21 and miR-150 was downregulated in ALL patients compared with controls (unknown significance), whereas miR-451 expression displayed no difference. |
| Piatopoulou et al., 2017 [57] | 125 childhood ALL patients at diagnosis and 64 controls; Greece | qRT-PCR; BM | Expression analysis revealed the significant downregulation of miR-125b levels in childhood ALL patients compared to controls (p < 0.004). The discriminatory significance of miR-125b for childhood ALL from healthy children was confirmed by univariate logistic regression (OR, 0.477; 95% CI: 0.288 to 0.790; p = 0.004) and ROC analysis (AUC, 0.628; 95% CI: 0.548 to 0.707; p = 0.004). The discriminatory value of miR-125b seems to be independent of patient age and gender according to the adjusted multivariate logistic regression model (OR, 0.507; 95% CI: 0.305 to 0.842; p = 0.009). |
| Nabhan et al., 2017 [58] | 30 childhood ALL patients at diagnosis and 30 controls; Egypt | qRT-PCR; serum | Significant downregulation of miR-181a in children with ALL compared with controls (p < 0.01). The previous finding were associated with SMAD7 overexpression and TGF-β1 downregulation (increasing proliferation and decreasing apoptosis). ROC curve analysis revealed an AUC of 0.93 (cutoff, 0.97; 86.5% sensitivity; 93.3% specificity; miR-181a positivity rate, 86.7%; p < 0.01). Serum levels of SMAD7, TGF-β1 (cutoff values ≥400.1 pg/mL and ≤0.25 ng/mL, respectively) and miR-181a seem to discriminate ALL cases from controls with 100% sensitivity, 93.3% specificity, 96.7% accuracy, 100% negative predictive value and 93.7% positive predictive value. |
| Yuan et al., 2017 [59] | 111 childhood ALL patients and 111 controls with fractures; China | qRT-PCR; plasma | The expression levels of let-7f-5p, miR-5100 and miR-25-3p in ALL patients were significantly lower than in controls (p < 0.01). After adjusting for confounders, risk for ALL (OR and 95% CI) was calculated to be 0.84 (0.76 to 0.92), 0.81 (0.73 to 0.9) and 0.81 (0.74 to 0.89), respectively (p < 0.01 in all instances). ROC analysis and reclassification analysis showed that utilization of all three miRNAs, compared to established risk factors, can improve the AUC and improve diagnosis (0.78; 95% CI: 0.72 to 0.84; p = 0.012). |
| Labib et al., 2017 [60] | 75 childhood B-ALL patients at diagnosis and 50 controls; Egypt | qRT-PCR; BM and serum | Significant upregulation of miR-21 in children with B-ALL compared to controls (9.62 ± 3.23 vs. 2.56 ± 0.83; p < 0.001; cutoff, 9.83 for high and low expression). ROC curve analysis revealed an AUC of 0.879 (88.7% sensitivity; 71.8% specificity; cutoff, 3.23) in distinguishing ALL patients from controls. |
| Tian et al., 2017 [61] | 189 childhood B-ALL cases and 189 controls; China | qRT-PCR; N/A | Significant downregulation of miR-3173 in children with B-ALL compared to controls (p < 0.001). |
| Ramani et al., 2017 [62] | 60 childhood B-ALL patients and 17 controls; the Netherlands | qRT-PCR; BM or PBMCs | The analysis revealed a signature of 136 significantly (p < 0.05; 45 of them with p < 10−5) differentially expressed miRNAs distinguishing pediatric patients with ALL from controls. Significant upregulation in ALL: miR-133b, miR-302, miR-190, miR-520, miR-10b, miR-515-5p, miR-517b, miR-501, miR-129, miR-155, miR-217, miR-330, miR-513, miR-585, miR-645, miR-617, miR-15b, miR-23a, miR-362, miR-368, miR-425-5p, miR-576 and miR-369-5p. Significant downregulation in ALL: miR-193b, miR-325, miR-514, miR-22, miR-7g, miR-7d, miR-302d, miR-206, miR-494, miR-101, miR-126, miR-100, miR-29a, miR-299-3p, miR-146, miR-20a, miR-374, miR-216, miR-532, miR-25, miR-30c and miR-30e-3p. |
| Cao et al., 2016 [63] | 42 childhood ALL patients and 20 controls; China | qRT-PCR; BM | Significant downregulation of miR-34b in ALL patients compared with controls (1.65 ± 0.69 vs. 5.22 ± 1.15; p = 0.012). |
| Lou et al., 2016 [64] | 20 childhood B-ALL patients and 20 controls; China | qRT-PCR; BM | Expression of miR-187-5p was significantly upregulated in B-ALL samples compared with controls (p < 0.001). This miRNA seems to modulate the Wnt/b-catenin signaling pathway via direct targeting of DDK2, thus promoting ALL cell proliferation and inhibiting apoptosis. |
| Swellam et al., 2016 [65] | 85 childhood ALL patients at diagnosis and 25 controls; Egypt | qRT-PCR; PBMCs | Significant upregulation of miR-100 (median: 61.777 vs. 2.04; range: 0.14–259.39 vs. 0.173–3.029; p = 0.007) and miR-146a (121.3 vs. 2.05; 14.3–222.55 vs. 0–144; p < 0.0001) in ALL patients compared with controls. Significant downregulation of miR-196a (0.369 vs. 0.511; 0.001–144 vs. 0.08–0.973; p = 0.028) in ALL samples compared with controls. ROC analysis revealed a significant AUC for miRNA-146a (1; 95% CI: 0.956 to 1; SE 0; 100% sensitivity; 100% specificity; cutoff, 3.727; p < 0.0001) and miR-100 (0.87; 95% CI: 0.779 to 0.934; SE 0.038; 82.76% sensitivity; 100% specificity; cutoff, 3.029; p = 0.0001) in discriminating ALL patients from controls. |
| de Oliveira et al., 2015 [66] | 128 childhood ALL patients at diagnosis and 11 controls; Brazil | qRT-PCR; BM | Significantly higher miR-708-5p expression in ALL samples compared with controls (p < 0.05). High expression was documented specifically for pre-B ALL (p < 0.01). |
| Wu et al., 2015 [67] | 40 infants with KMT2A-rearranged ALL (26 KMT2A-AFF1, 4 KMT2A-MLLT3, 5 KMT2A-MLLT1 and 5 KMT2A-germline) at diagnosis and 8 controls; Japan | qRT-PCR; BM or whole blood | In infant ALL cells with KMT2A fusion, let-7b was significantly downregulated compared with controls according to DNA hypermethylation of its gene-promoter region. |
| Nemes et al., 2015 [68] | 24 childhood ALL patients and no controls; Hungary | qRT-PCR; bone marrow and whole blood | Significant downregulation of miR-21, miR-24 and miR-29b in T-ALL patients compared to controls (p < 0.05). Significant upregulation of miR-155 in B-ALL patients compared to controls (p < 0.05). Significant upregulation of miR-128b in both B- and T-ALL children compared with controls (p < 0.05). Expression profiles of miRNAs were not significantly different between peripheral blood and bone marrow samples derived from the same patient. |
| Luna-Aguirre et al., 2015 [69] | 19 childhood B-ALL patients at diagnosis and 7 adult controls; Mexico | qRT-PCR; plasma | A total of 40 significantly overexpressed miRNAs in B-ALL patients compared to controls: miR-511, miR-34a*, miR-565, miR-34a, miR-10b*, miR-630, miR-610, miR-181a, miR-181c, miR-222, miR-138-1*, miR-363, miR-144*, miR-451, miR-99a, miR-155, miR-886-3p, miR-223*, miR-422a, miR-146a, miR-192, miR-190b, miR-95, miR-140-3p, miR-660, miR-886-5p, miR-25, miR-320, miR-30e, miR-16, miR-19b, miR-500, miR-29a, miR-502-3p, miR-195, miR-20b, miR-579, miR-7, miR-19a and miR-768-3p. A total of 37 miRNAs significantly downregulated in B-ALL patients compared with controls: miR-199a-3p, miR-340*, miR-151-3p, miR-335, miR-99b, miR-425*, miR-224, miR-221, miR-744, miR-15b, miR-223, miR-26a, miR-454*, miR-452, miR-491-5p, miR-340, miR-196b, miR-301a, miR-324-5p, miR-126*, miR-152, miR-330-3p, miR-652, miR-374b, miR-148b, miR-671-3p, miR-18a, let-7d, miR-339-3p, miR-126, miR-30b, miR-148b*, miR-27a, miR-30c, miR-374a, miR-331-3p and miR-28-5p. Diagnostic capacity for miR-511 (overexpressed in B-ALL; range, 3.32 to 12.98; cutoff, 9.458; AUC, 1; 100% sensitivity, specificity, PPV and NPV), miR-34a (overexpressed; range, 0.73 to 7.55; cutoff, 7.179; AUC, 0.98; 92% sensitivity; 100% specificity and PPV; 70% NPV), miR-222 (overexpressed; range, −1.5 to 3.12; cutoff, −0.1325; AUC, 0.91; 79% sensitivity; 100% specificity and PPV; 54% NPV), miR-26a (underexpressed; range, −1.63 to −9.17; cutoff, 2.073; AUC, 0.91; 79% sensitivity; 100% specificity and PPV; 47% NPV), miR-221 (underexpressed; range, 1.1 to −8.21; cutoff, −0.1861; AUC, 0.92; 83% sensitivity; 100% specificity and PPV; 54% NPV) and miR-223 (underexpressed; range, 0.31 to −8.52; cutoff, −4.309; AUC, 0.93; 89% sensitivity; 100% specificity and PPV; 64% NPV) showed that miR-511 could be used as a circulating biomarker for B-ALL detection. Moreover, miR-199a-3p (most underexpressed compared to controls; RQ, -13.48; p < 0.001), along with miR-511 (most overexpressed; RQ, 159.55; p = 0.002), might be associated with the pathogenesis of B-ALL. |
| Organista-Nava et al., 2015 [70] | 111 childhood ALL patients and 100 controls; Mexico | qRT-PCR; BM and/or whole blood | Significantly lower miR-24 expression in ALL patients compared with controls (median, 0.84; p = 0.002). |
| Oliveira et al., 2015 [71] | 37 childhood T-ALL patients and normal T-cells; Brazil | qRT-PCR; BM or whole blood | Significantly lower miR-29a expression in T-ALL patients compared with control T-cells. |
| Duyu et al., 2014 [28] | 43 childhood ALL patients at diagnosis and 14 controls; Turkey | qRT-PCR and microarray; BM and whole blood | A total of 13 miRNAs (miR-548i, miR-708, miR-181b, miR-449a, miR-146a, miR-155, miR-181a*, miR-3121, miR-181a, miR-128, miR-1323, miR-195 and miR-587) showed upregulation, and 2 miRNAs (miR-640 and miR-145) showed downregulation in the microarray study. Confirmation analysis by qRT-PCR demonstrated only five upregulated miRNAs: miR-128, miR-146a, miR-155, miR-181a and miR-195 (p < 0.05). Microarrays revealed significant upregulation of miR-548i, miR-3140, miR-181b, miR-3115, miR-548d-5p, miR-449a, miR-181c, miR-181a, miR-181a*, let-7b*, miR-1827, miR-92a-1*, miR-299-3p, miR-155, miR-181a-2*, miR-1323, miR-587, miR-7-1*, miR-28-3p, miR-130b, miR-27b and miR-361-5p and significant downregulation of miR-633, miR-326, miR-501-3p, miR-802, miR-4260, miR-3130-5p, miR-145, miR-186, miR-593*, miR-574-3p, miR-4262, miR-640 and miR-606 in T-ALL patients compared to controls. Significant upregulation of miR-708, miR-181b, miR-369-3p, miR-146a, miR-155, miR-195 and miR-128 and significant downregulation of miR-143 and miR-145 in children with B-ALL compared with healthy controls. Intriguingly, miRNAs from peripheral blood samples were not correlated with BM samples. |
| Malik et al., 2014 [72] | 30 childhood B-ALL and 20 T-ALL patients and 50 controls; India | qRT-PCR; PBMCs | Significantly higher expression of miR-2909 in both B- and T-ALL samples compared with controls (p < 0.01). |
| Gimenes-Teixeira et al., 2013 [73] | 48 childhood T-ALL patients and 10 controls; Brazil | qRT-PCR; BM, PBMCs and thymic samples | Significantly higher miR-221 and miR-374 expression in T-ALL samples compared with controls (p < 0.05). |
| Xue Li et al., 2013 [74] | 34 childhood common-ALL patients and 5 controls; China | Microarray; BM | Significantly upregulated expression of miR-708 (16.886 ± 16.854 vs. 1.872 ± 0.339; p < 0.01), miR-181b (5.710 ± 4.652 vs. 1.276 ± 0.531; p = 0.006) and miR-210 (9.789 ± 1.178 vs. 1.005 ± 0.08; p < 0.01) in children with common ALL compared with controls. Significant downregulation of miR-345 (0.675 ± 0.086 vs. 1.204 ± 0.143; p = 0.007) and miR-27a (0.523 ± 0.085 vs. 1.123 ± 0.066; p = 0.004) in common ALL compared with controls. |
| Li et al., 2013 [75] | 111 childhood ALL patients and 10 controls; China | qRT-PCR; BM | The expression levels of miR-100 and miR-99a were significantly downregulated in ALL patients compared with controls (p = 0.001 and p = 0.0086, respectively). |
| de Oliveira et al., 2012 [76] | 128 childhood ALL patients at diagnosis and 11 controls; Brazil | qRT-PCR; BM | Significantly lower expression of miR-100 (median, 0.21; range, 0.002 to 10.76; vs. 1.04 (0.34 to 3.9); p < 0.01)), miR-196b (0.03 (0.001 to 65.68) vs. 1.07 (0.29 to 3.7); p < 0.01) and let-7e (0.59 (0.04 to 5.15) vs. 1.16 (0.54 to 3.25); p < 0.01) and significantly higher expression of miR-128a (4.32 (0.16 to 167.91) vs. 1.02 (0.57 to 2.23); p < 0.01) and miR-181b (5.06 (0.07 to 45.09) vs. 1.41 (0.33 to 3.4); p < 0.01) in ALL samples compared to controls. The expression of miR-92a and miR-125a-5p was similar for both groups. |
| Borze et al., 2011 [77] | 9 childhood ALL patients and 8 controls of N/A age; Finland | Microarray; BM | Significant upregulation of miR-142-3p (p = 0.001), miR-146a (p = 0.004), miR-222 (p = 0.012), miR-142-5p (p = 0.022), miR-150* (p = 0.032), miR-144 (p = 0.041) and miR-155 (p = 0.043) in ALL samples compared with controls. Significant downregulation of miR-768-5p (p < 0.001), miR-125b (p = 0.006), miR-223 (p = 0.007), miR-22 (p = 0.012), miR-27a (p = 0.027) and miR-15b (p = 0.032) in ALL patients compared with healthy donors. Similar expression levels of miRNAs between BM core biopsies (under decalcification and fixation with formalin) and BM aspirates. |
| Bhatia et al., 2011 [78] | 10 childhood T-ALL patients at diagnosis and control T-cells; India | Semi-qRT-PCR; PBMCs | Significantly lower expression levels of miR-196b and miR-148a and significantly higher expression levels of miR-30b and miR-151 in children with T-ALL compared with normal T-cells (p < 0.05). Consequently, miR-196b was unable to downregulate MYC protooncogene expression in T-ALL patients as a result of mutations in the 3′-untranslated region (3′-UTR) of the target. |
| Mavrakis et al., 2011 [79] | 50 childhood T-ALL patients at diagnosis and normal T-cell populations from pediatric thymuses; Belgium and France | qRT-PCR; BM and thymic T-cells | Significantly higher miR-223, miR-19b, miR-20a, miR-92, miR-142-3p, miR-150, miR-93, miR-26a, miR-16 and miR-342 expression in T-ALL samples compared with controls (p < 0.05). |
| Stumpel et al., 2011 [80] | 5 t(4;11)-positive ALL infants at diagnosis and 7 controls; the Netherlands | qRT-PCR; BM | Significant downregulation of miR-10a, miR-101, miR-148a, miR-152, miR-200a, miR-200b, miR-424, miR-429, miR-432, miR-486 and miR-503 in KMT2A-rearranged ALL samples compared with controls (via hypermethylation). |
| Schotte et al., 2011 [81] | 70 childhood ALL patients at diagnosis and 10 controls; the Netherlands | RNA-seq and qRT-PCR; BM and PBMCs | Significant downregulation of miR-3136 and miR-4474 in ALL patients compared to controls. |
| Bhatia et al., 2010 [82] | 10 ALL patients of N/A age and normal B-cells; India | RT-PCR; PBMCs | Significant downregulation of miR-151, miR-196b and miR-148a in patient samples compared with B-cells obtained from healthy volunteers (p < 0.05). Among them, only miR-196b was found to be downregulated in the EB-3 cell line (potential tumor-suppressive function), whereas it seemed to downregulate the highly expressed MYC and, in turn, to decrease the expression of MYC effector genes (TERT, BCL2 and AATF). |
| Schotte et al., 2009 [83] | 92 childhood ALL patients at diagnosis (20 MLL-rearranged, 57 B-ALL and 15 T-ALL) and normal CD34+ progenitor cells from 2 children with brain tumors; the Netherlands | qRT-PCR; BM or whole blood | Significant upregulation of miR-128a, miR-142-3p and -5p, as well as miR-150, miR-151-5p (not in KMT2A-rearranged cases), miR-181a, miR-181b, miR-181c, miR-193a, miR-30e-5p, miR-34b, miR-365, miR-582 and miR-708 (p ≤ 0.001) in ALL patients compared with controls. Significant downregulation of miR-100, miR-125b, miR-99a and let-7e in children with ALL compared with controls (p < 0.001). Conversely, miR-196b was found to be significantly downregulated in B-other cases (p < 0.001) and significantly upregulated in KMT2A-rearranged cases (p = 0.021) compared to controls. |
| Zhang, Yang, et al., 2009 [84] | 24 childhood ALL patients at diagnosis and 3 controls; China | qRT-PCR; BM | Significantly higher expression levels of miR-9, miR-9*, miR-181a, miR-128, miR-181b, miR-130b and miR-363 in children with ALL compared with controls. Significantly lower expression of miR-7e, miR-30a, miR-199b-3p, miR-126, miR-143, miR-223 and miR-582-5p in ALL samples compared to controls. With respect to novel miRNAs miR-1943, miR-1841, miR-1931, miR-1987, miR-1890 and miR-1902 were upregulated, and miR-1859, miR-1859*, miR-1947, miR-1971, miR-1866, miR-1986, miR-1843, miR-1852, miR-1852*, miR-1842, miR-1834, miR-1971* and miR-1893 were down regulated in ALL specimens compared with controls (p < 0.001). |
| Ju et al., 2009 [85] | 40 childhood pre-B-ALL patients at diagnosis and 6 controls; China | Microarray and qRT-PCR; BM | Significantly higher expression levels of miR-339 (p < 0.001), miR-222 (p = 0.0061) and miR-142-3p (p < 0.001) and significantly lower expression of miR-451 and miR-373* (p < 0.001) in ALL samples compared with controls. Upregulation of miR-361, miR-487b and miR-519e and downregulation of miR-296, miR-485-3p and miR-483 in ALL specimens compared with controls in microarrays, although not been confirmed by qRT-PCR. |
| Zhang, Luo, et al., 2009 [86] | 18 childhood ALL patients and 7 controls (validation cohort: 31 ALL cases and 5 controls); China | Microarray and qRT-PCR; BM | Significantly higher expression of miR-146a, miR-34a, miR-210, miR-213, miR-181c, miR-126, miR-181a, miR-181d, miR-130a, miR-195, miR-181b, miR-130b, miR-155, miR-17-3p, miR-128b, miR-128a, miR-18b, miR-28, miR-331, miR-505 and miR-363 and significantly lower expression of miR-582, miR-142-5p, miR-145, miR-143, miR-338, miR-148a, miR-424, miR-24, miR-29a, miR-199b and miR-199a in children with ALL compared with controls. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyriakidis, I.; Kyriakidis, K.; Tsezou, A. MicroRNAs and the Diagnosis of Childhood Acute Lymphoblastic Leukemia: Systematic Review, Meta-Analysis and Re-Analysis with Novel Small RNA-Seq Tools. Cancers 2022, 14, 3976. https://doi.org/10.3390/cancers14163976
Kyriakidis I, Kyriakidis K, Tsezou A. MicroRNAs and the Diagnosis of Childhood Acute Lymphoblastic Leukemia: Systematic Review, Meta-Analysis and Re-Analysis with Novel Small RNA-Seq Tools. Cancers. 2022; 14(16):3976. https://doi.org/10.3390/cancers14163976
Chicago/Turabian StyleKyriakidis, Ioannis, Konstantinos Kyriakidis, and Aspasia Tsezou. 2022. "MicroRNAs and the Diagnosis of Childhood Acute Lymphoblastic Leukemia: Systematic Review, Meta-Analysis and Re-Analysis with Novel Small RNA-Seq Tools" Cancers 14, no. 16: 3976. https://doi.org/10.3390/cancers14163976
APA StyleKyriakidis, I., Kyriakidis, K., & Tsezou, A. (2022). MicroRNAs and the Diagnosis of Childhood Acute Lymphoblastic Leukemia: Systematic Review, Meta-Analysis and Re-Analysis with Novel Small RNA-Seq Tools. Cancers, 14(16), 3976. https://doi.org/10.3390/cancers14163976