
RETRACTED: HOXA11-AS1 Promotes PD-L1-Mediated Immune Escape and Metastasis of Hypopharyngeal Carcinoma by Facilitating PTBP1 and FOSL1 Association
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Cell Culture and Transfection
2.3. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR) Assay
2.4. Western Blot
2.5. Immunohistochemistry (IHC)
2.6. CCK-8 Assay
2.7. Colony Formation Assay
2.8. Wound Healing Assay
2.9. Transwell Assay
2.10. Co-Culture of Human PBMCs and HSCC Cells
2.11. Flow Cytometry
2.12. Enzyme-Linked Immunosorbent Assay (ELISA)
2.13. Immunofluorescence Assay
2.14. Fractionation of Nuclear/Cytoplasmic RNA
2.15. Chromatin Immunoprecipitation (ChIP) Assay
2.16. Dual-Luciferase Reporter Assay
2.17. mRNA Stability Analysis
2.18. RNA Pulldown
2.19. RNA Immunoprecipitation (RIP) Assay
2.20. In Vivo Xenograft Experiments
2.21. Statistical Analysis
3. Results
3.1. HOXA11-AS1 and PD-L1 Were Highly Expressed in HSCC, and HOXA11-AS1 Positively Correlated with PD-L1
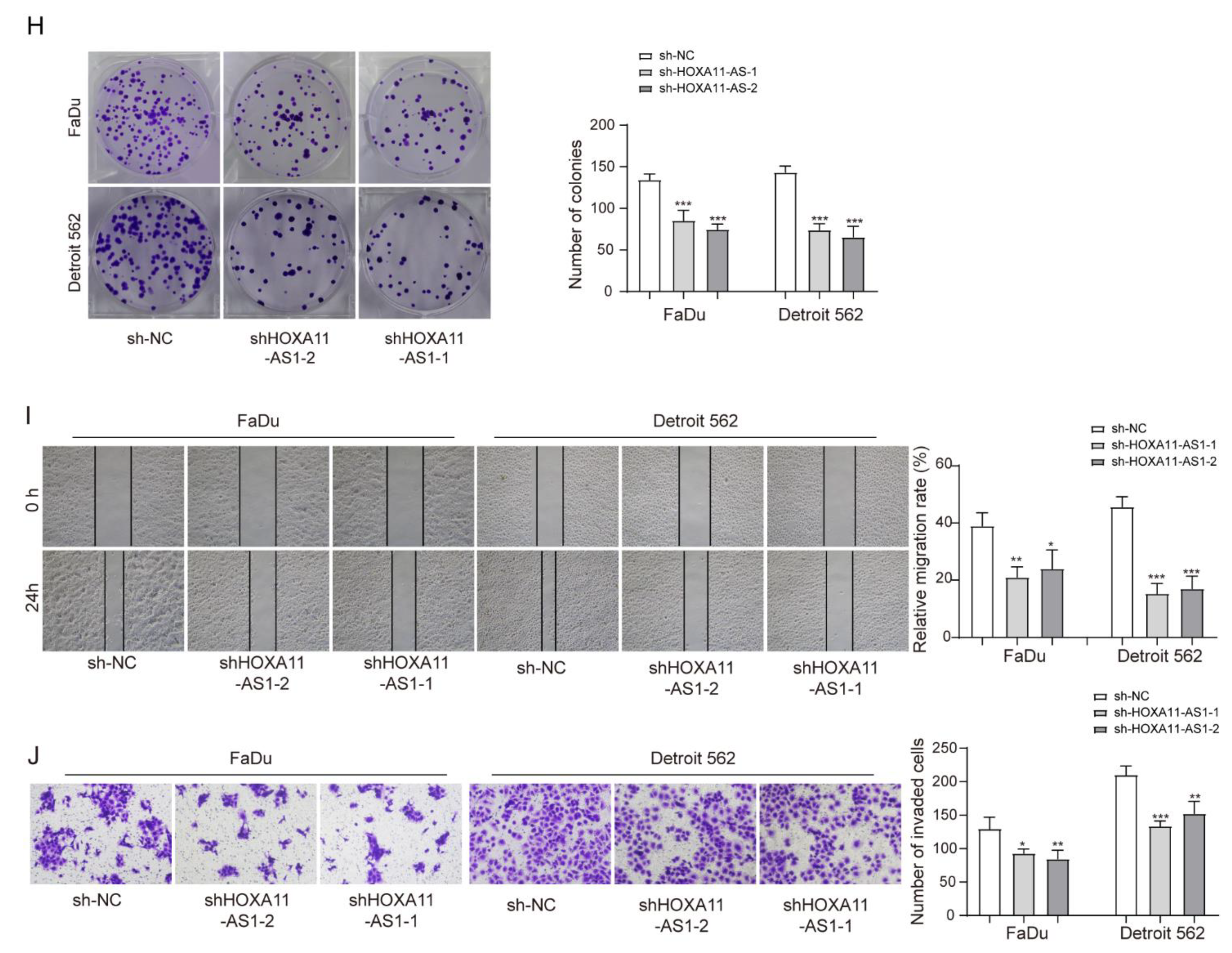
3.2. HOXA11-AS1 Knockdown Suppressed PD-L1 Expression and Immune Escape, Proliferation, and Metastasis of HSCC Cells
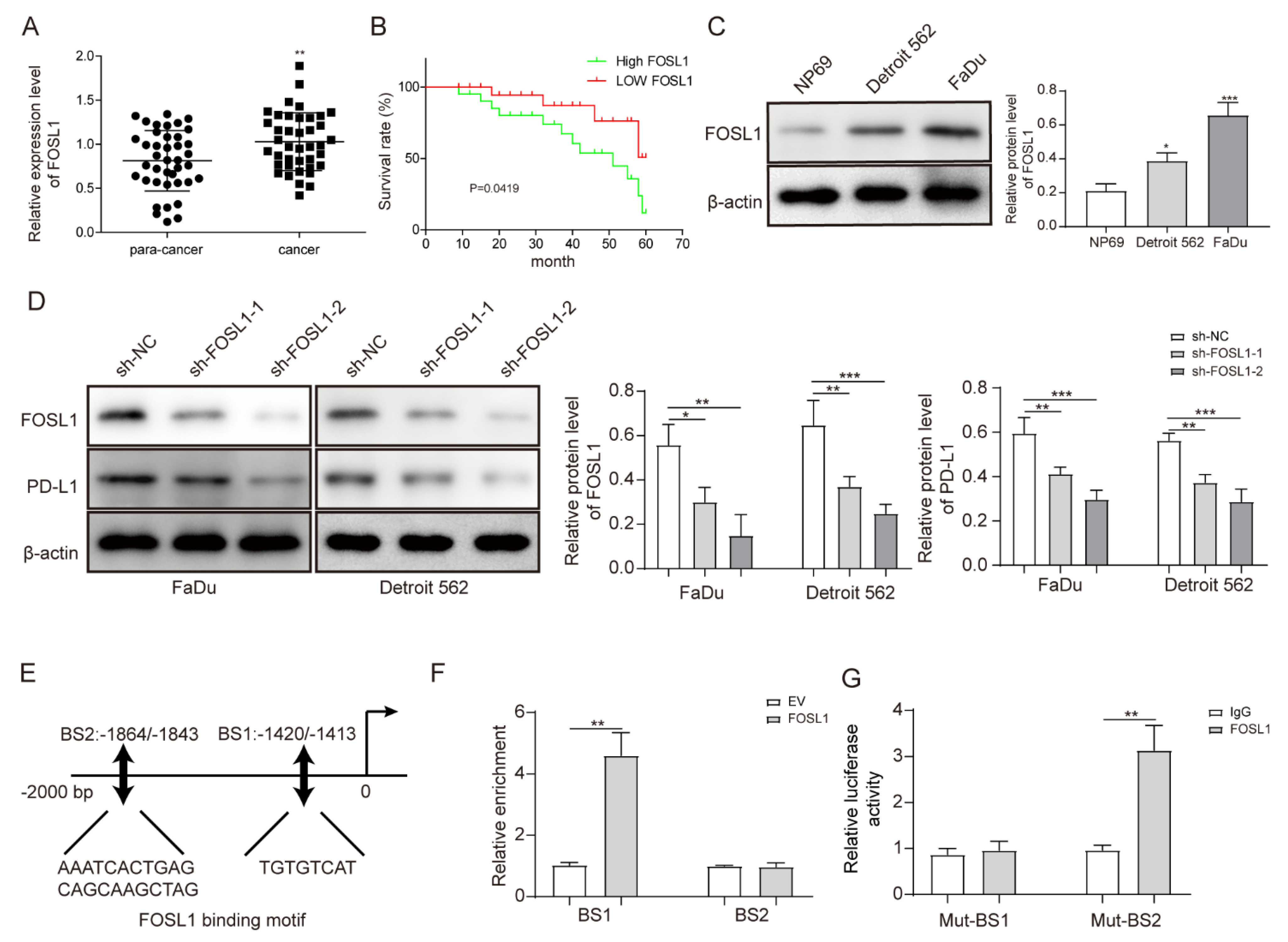
3.3. FOSL1 was Highly Expressed in HSCC Cells and Positively Regulated PD-L1 Expression by Binding the PD-L1 Promoter
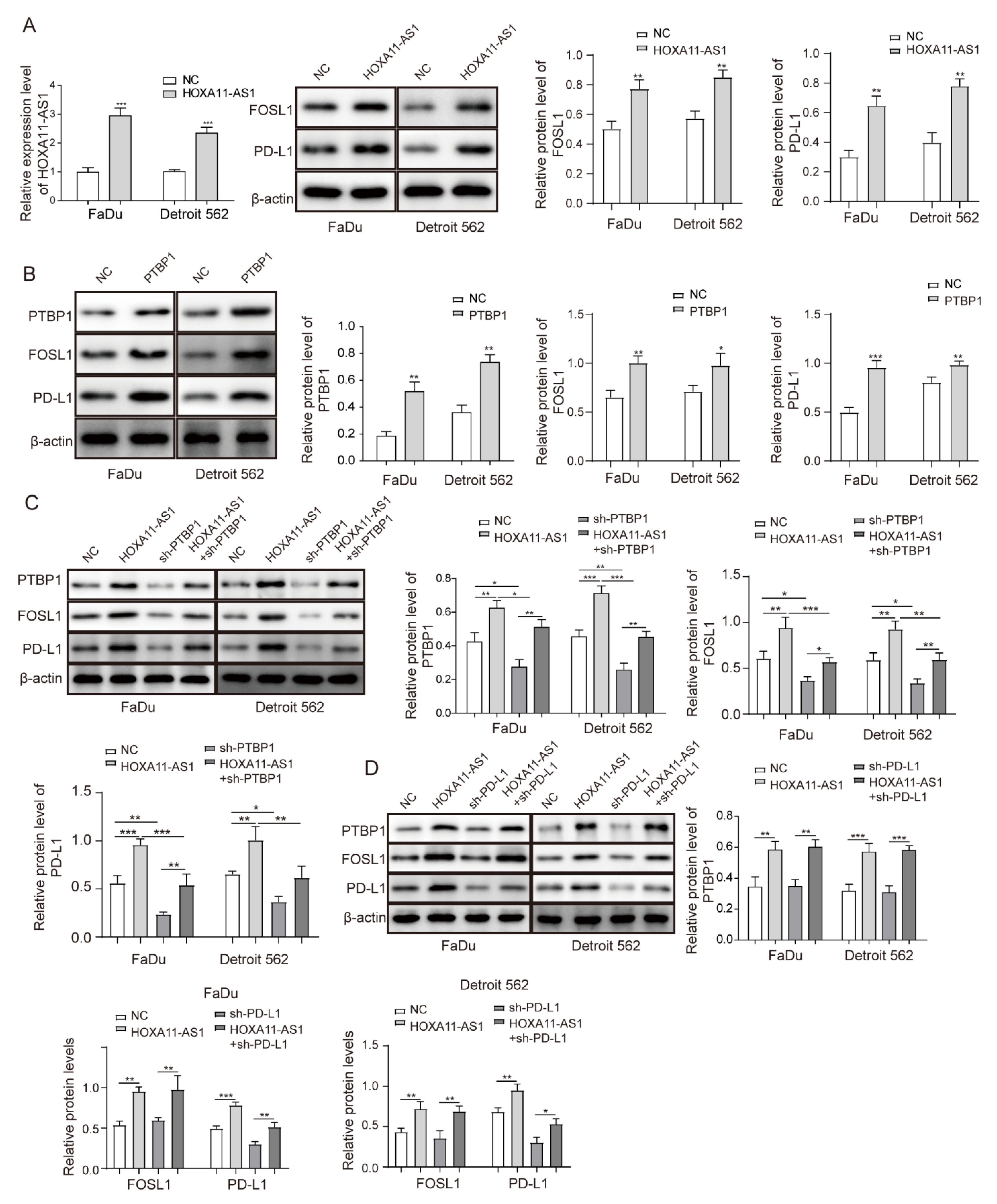
3.4. HOXA11-AS1 Enhanced FOSL1 mRNA Stability by Binding PTBP1
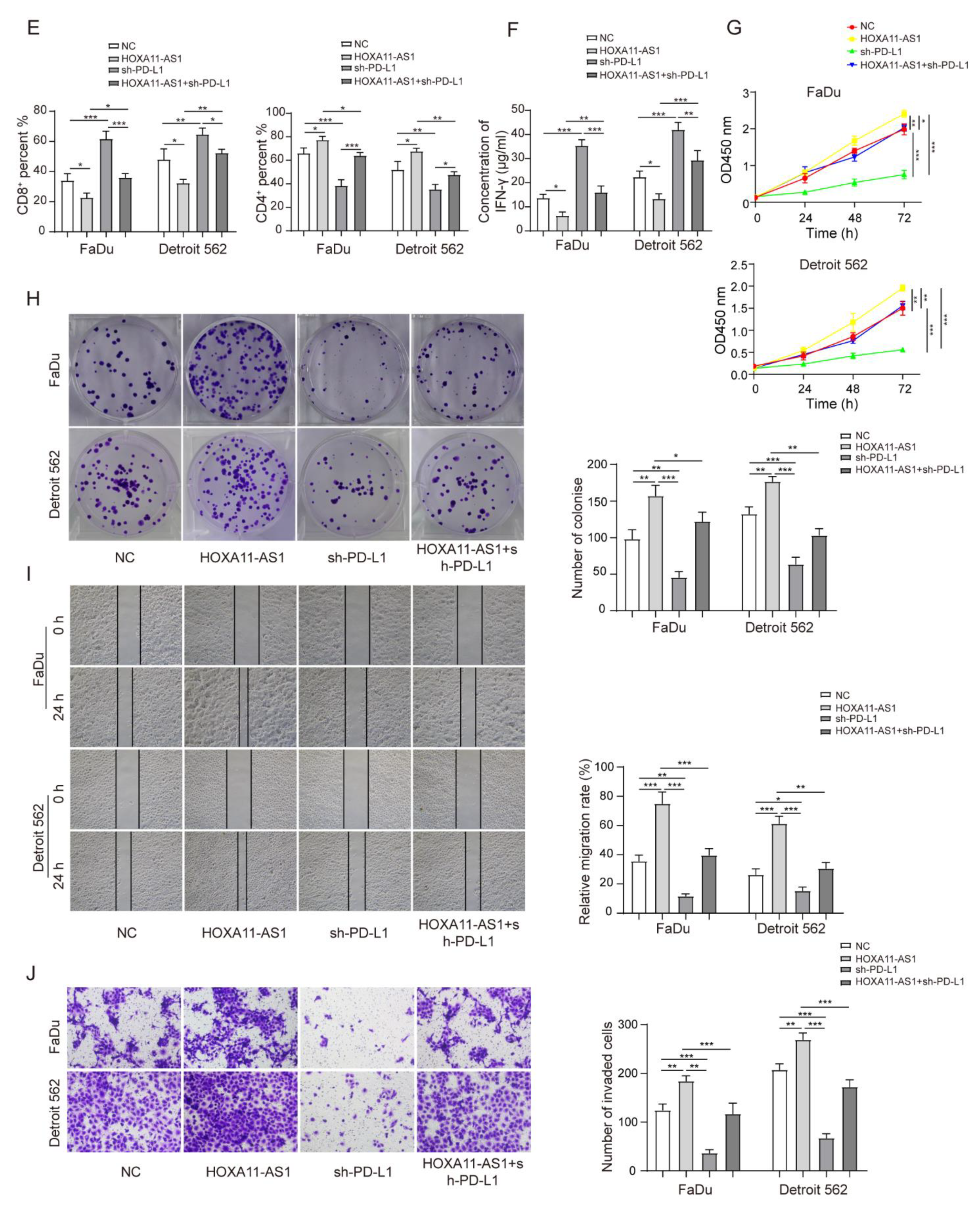
3.5. HOXA11-AS1 Promoted PD-L1 Expression by Upregulating FOSL1 Levels through PTBP1, Thereby Facilitating Cell Immune Escape, Growth, and Metastasis
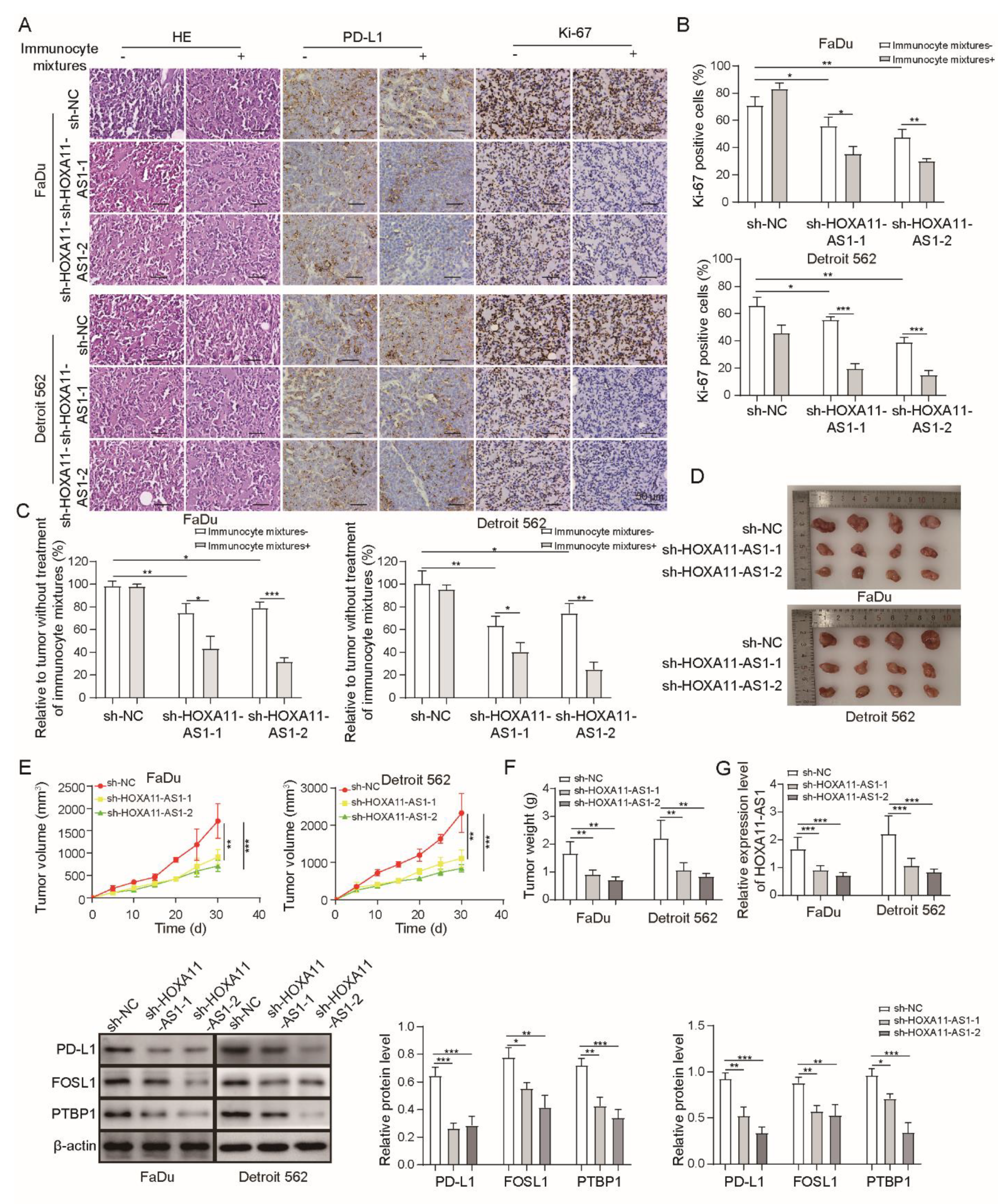
3.6. HOXA11-AS1 Knockdown Inhibited Immune Escape and Metastasis by Regulating PD-L1 and Downregulating FOSL1 In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cooper, J.S.; Porter, K.; Mallin, K.; Hoffman, H.T.; Weber, R.S.; Ang, K.K.; Gay, E.G.; Langer, C.J. National Cancer Database report on cancer of the head and neck: 10-year update. Head Neck 2009, 31, 748–758. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Sewnaik, A.; Hoorweg, J.J.; Knegt, P.P.; Wieringa, M.H.; van der Beek, J.M.; Kerrebijn, J.D. Treatment of hypopharyngeal carcinoma: Analysis of nationwide study in the Netherlands over a 10-year period. Clin. Otolaryngol. 2005, 30, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Tsou, Y.A.; Lin, M.H.; Hua, C.H.; Tseng, H.C.; Chen, S.W.; Yang, S.N.; Liang, J.A.; Tsai, M.H. Survival outcome by early chemoradiation therapy salvage or early surgical salvage for the treatment of hypopharyngeal cancer. Otolaryngol. Head Neck Surg. 2007, 137, 711–716. [Google Scholar] [CrossRef]
- Wycliffe, N.D.; Grover, R.S.; Kim, P.D.; Simental, A., Jr. Hypopharyngeal cancer. Top. Magn. Reson. Imaging 2007, 18, 243–258. [Google Scholar] [CrossRef]
- Topalian, S.L.; Drake, C.G.; Pardoll, D.M. Targeting the PD-1/B7-H1(PD-L1) pathway to activate anti-tumor immunity. Curr. Opin. Immunol. 2012, 24, 207–212. [Google Scholar] [CrossRef]
- Zou, W.; Chen, L. Inhibitory B7-family molecules in the tumour microenvironment. Nat. Rev. Immunol. 2008, 8, 467–477. [Google Scholar] [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar]
- Lenouvel, D.; Gonzalez-Moles, M.A.; Ruiz-Avila, I.; Chamorro-Santos, C.; Gonzalez-Ruiz, L.; Gonzalez-Ruiz, I.; Ramos-Garcia, P. Clinicopathological and prognostic significance of PD-L1 in oral cancer: A preliminary retrospective immunohistochemistry study. Oral Dis. 2021, 27, 173–182. [Google Scholar] [CrossRef]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef]
- Wintterle, S.; Schreiner, B.; Mitsdoerffer, M.; Schneider, D.; Chen, L.; Meyermann, R.; Weller, M.; Wiendl, H. Expression of the B7-related molecule B7-H1 by glioma cells: A potential mechanism of immune paralysis. Cancer Res. 2003, 63, 7462–7467. [Google Scholar]
- Hodgson, A.; Slodkowska, E.; Jungbluth, A.; Liu, S.K.; Vesprini, D.; Enepekides, D.; Higgins, K.; Katabi, N.; Xu, B.; Downes, M.R. PD-L1 Immunohistochemistry Assay Concordance in Urothelial Carcinoma of the Bladder and Hypopharyngeal Squamous Cell Carcinoma. Am. J. Surg. Pathol. 2018, 42, 1059–1066. [Google Scholar] [CrossRef]
- Cui, P.; Jing, P.; Liu, X.; Xu, W. Prognostic Significance of PD-L1 Expression and Its Tumor-Intrinsic Functions in Hypopharyngeal Squamous Cell Carcinoma. Cancer Manag. Res. 2020, 12, 5893–5902. [Google Scholar] [CrossRef]
- Hanna, G.J.; O’Neill, A.; Shin, K.Y.; Wong, K.; Jo, V.Y.; Quinn, C.T.; Cutler, J.M.; Flynn, M.; Lizotte, P.H.; Annino, D.J., Jr.; et al. Neoadjuvant and Adjuvant Nivolumab and Lirilumab in Patients with Recurrent, Resectable Squamous Cell Carcinoma of the Head and Neck. Clin. Cancer Res. 2022, 28, 468–478. [Google Scholar] [CrossRef]
- Desmet, C.J.; Gallenne, T.; Prieur, A.; Reyal, F.; Visser, N.L.; Wittner, B.S.; Smit, M.A.; Geiger, T.R.; Laoukili, J.; Iskit, S.; et al. Identification of a pharmacologically tractable Fra-1/ADORA2B axis promoting breast cancer metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 5139–5144. [Google Scholar] [CrossRef]
- Xu, L.; Hu, H.; Zheng, L.S.; Wang, M.Y.; Mei, Y.; Peng, L.X.; Qiang, Y.Y.; Li, C.Z.; Meng, D.F.; Wang, M.D.; et al. ETV4 is a theranostic target in clear cell renal cell carcinoma that promotes metastasis by activating the pro-metastatic gene FOSL1 in a PI3K-AKT dependent manner. Cancer Lett. 2020, 482, 74–89. [Google Scholar] [CrossRef]
- Cui, Y.P.; Xie, M.; Pan, W.X.; Zhang, Z.Y.; Li, W.F. HOXA10 promotes the development of bladder cancer through regulating FOSL1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2945–2954. [Google Scholar] [CrossRef]
- Chan, J.J.; Tay, Y. Noncoding RNA:RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310. [Google Scholar] [CrossRef]
- Ren, X. Genome-wide analysis reveals the emerging roles of long non-coding RNAs in cancer. Oncol. Lett. 2020, 19, 588–594. [Google Scholar] [CrossRef]
- Qian, Y.; Liu, D.; Cao, S.; Tao, Y.; Wei, D.; Li, W.; Li, G.; Pan, X.; Lei, D. Upregulation of the long noncoding RNA UCA1 affects the proliferation, invasion, and survival of hypopharyngeal carcinoma. Mol. Cancer 2017, 16, 68. [Google Scholar] [CrossRef]
- Cheng, Y.; Xiong, H.Y.; Li, Y.M.; Zuo, H.R.; Liu, Y.; Liao, G.L. LncRNA HOXA11-AS promotes cell growth by sponging miR-24-3p to regulate JPT1 in prostate cancer. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 4668–4677. [Google Scholar] [CrossRef]
- Bai, Y.; Lang, L.; Zhao, W.; Niu, R. Long Non-Coding RNA HOXA11-AS Promotes Non-Small Cell Lung Cancer Tumorigenesis Through microRNA-148a-3p/DNMT1 Regulatory Axis. Onco Targets Ther. 2019, 12, 11195–11206. [Google Scholar] [CrossRef]
- Xu, J.; Bo, Q.; Zhang, X.; Lei, D.; Wang, J.; Pan, X. lncRNA HOXA11-AS Promotes Proliferation and Migration via Sponging miR-155 in Hypopharyngeal Squamous Cell Carcinoma. Oncol. Res. 2020, 28, 311–319. [Google Scholar] [CrossRef]
- Zhu, W.; Zhou, B.L.; Rong, L.J.; Ye, L.; Xu, H.J.; Zhou, Y.; Yan, X.J.; Liu, W.D.; Zhu, B.; Wang, L.; et al. Roles of PTBP1 in alternative splicing, glycolysis, and oncogensis. J. Zhejiang Univ. Sci. B 2020, 21, 122–136. [Google Scholar] [CrossRef]
- Huan, L.; Guo, T.; Wu, Y.; Xu, L.; Huang, S.; Xu, Y.; Liang, L.; He, X. Hypoxia induced LUCAT1/PTBP1 axis modulates cancer cell viability and chemotherapy response. Mol. Cancer 2020, 19, 11. [Google Scholar] [CrossRef]
- Iwai, Y.; Ishida, M.; Tanaka, Y.; Okazaki, T.; Honjo, T.; Minato, N. Involvement of PD-L1 on tumor cells in the escape from host immune system and tumor immunotherapy by PD-L1 blockade. Proc. Natl. Acad. Sci. USA 2002, 99, 12293–12297. [Google Scholar] [CrossRef]
- Wahlberg, P.C.; Andersson, K.E.; Biorklund, A.T.; Moller, T.R. Carcinoma of the hypopharynx: Analysis of incidence and survival in Sweden over a 30-year period. Head Neck 1998, 20, 714–719. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Yu, X.; Sun, Y.; Miao, S.; Liu, S.; Li, Z.; Yi, J.; An, C. Different Primary Sites of Hypopharyngeal Cancer Have Different Lymph Node Metastasis Patterns: A Retrospective Analysis From Multi-Center Data. Front. Oncol. 2021, 11, 727991. [Google Scholar] [CrossRef]
- Prensner, J.R.; Chinnaiyan, A.M. The emergence of lncRNAs in cancer biology. Cancer Discov. 2011, 1, 391–407. [Google Scholar] [CrossRef]
- Gutschner, T.; Diederichs, S. The hallmarks of cancer: A long non-coding RNA point of view. RNA Biol. 2012, 9, 703–719. [Google Scholar] [CrossRef]
- Zhou, J.; Li, W.; Jin, T.; Xiang, X.; Li, M.; Wang, J.; Li, G.; Pan, X.; Lei, D. Gene microarray analysis of lncRNA and mRNA expression profiles in patients with hypopharyngeal squamous cell carcinoma. Int. J. Clin. Exp. Med. 2015, 8, 4862–4882. [Google Scholar] [PubMed]
- Zhou, J.; Li, M.; Yu, W.; Li, W.; Wang, J.; Xiang, X.; Li, G.; Pan, X.; Lei, D. AB209630, a long non-coding RNA decreased expression in hypopharyngeal squamous cell carcinoma, influences proliferation, invasion, metastasis, and survival. Oncotarget 2016, 7, 14628–14638. [Google Scholar] [CrossRef] [PubMed]
- Pauken, K.E.; Wherry, E.J. Overcoming T cell exhaustion in infection and cancer. Trends Immunol. 2015, 36, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Dermani, F.K.; Samadi, P.; Rahmani, G.; Kohlan, A.K.; Najafi, R. PD-1/PD-L1 immune checkpoint: Potential target for cancer therapy. J. Cell. Physiol. 2019, 234, 1313–1325. [Google Scholar] [CrossRef]
- Hu, C.; Tian, S.; Lin, L.; Zhang, J.; Ding, H. Prognostic and clinicopathological significance of PD-L1 and tumor infiltrating lymphocytes in hypopharyngeal squamous cell carcinoma. Oral Oncol. 2020, 102, 104560. [Google Scholar] [CrossRef]
- Zhao, M.; Sun, D.; Li, X.; Xu, Y.; Zhang, H.; Qin, Y.; Xia, M. Overexpression of long noncoding RNA PEG10 promotes proliferation, invasion and metastasis of hypopharyngeal squamous cell carcinoma. Oncol. Lett. 2017, 14, 2919–2925. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Zarrin, V.; Moghadam, E.R.; Zabolian, A.; Tavakol, S.; Samarghandian, S.; Najafi, M. PD-1/PD-L1 axis regulation in cancer therapy: The role of long non-coding RNAs and microRNAs. Life Sci. 2020, 256, 117899. [Google Scholar] [CrossRef]
- Mineo, M.; Lyons, S.M.; Zdioruk, M.; von Spreckelsen, N.; Ferrer-Luna, R.; Ito, H.; Alayo, Q.A.; Kharel, P.; Giantini Larsen, A.; Fan, W.Y.; et al. Tumor Interferon Signaling Is Regulated by a lncRNA INCR1 Transcribed from the PD-L1 Locus. Mol. Cell 2020, 78, 1207–1223. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, Y.; Zhang, J.; Liu, Y.; Qi, Q. LncRNA SNHG14/miR-5590-3p/ZEB1 positive feedback loop promoted diffuse large B cell lymphoma progression and immune evasion through regulating PD-1/PD-L1 checkpoint. Cell Death Dis. 2019, 10, 731. [Google Scholar] [CrossRef]
- Jiang, X.; Xie, H.; Dou, Y.; Yuan, J.; Zeng, D.; Xiao, S. Expression and function of FRA1 protein in tumors. Mol. Biol. Rep. 2020, 47, 737–752. [Google Scholar] [CrossRef]
- Elangovan, I.M.; Vaz, M.; Tamatam, C.R.; Potteti, H.R.; Reddy, N.M.; Reddy, S.P. FOSL1 Promotes Kras-induced Lung Cancer through Amphiregulin and Cell Survival Gene Regulation. Am. J. Respir. Cell Mol. Biol. 2018, 58, 625–635. [Google Scholar] [CrossRef]
- Knoch, K.P.; Bergert, H.; Borgonovo, B.; Saeger, H.D.; Altkruger, A.; Verkade, P.; Solimena, M. Polypyrimidine tract-binding protein promotes insulin secretory granule biogenesis. Nat. Cell Biol. 2004, 6, 207–214. [Google Scholar] [CrossRef]
- Sawicka, K.; Bushell, M.; Spriggs, K.A.; Willis, A.E. Polypyrimidine-tract-binding protein: A multifunctional RNA-binding protein. Biochem. Soc. Trans. 2008, 36, 641–647. [Google Scholar] [CrossRef]
- Li, J.; Yang, Y.; Fan, J.; Xu, H.; Fan, L.; Li, H.; Zhao, R.C. Long noncoding RNA ANCR inhibits the differentiation of mesenchymal stem cells toward definitive endoderm by facilitating the association of PTBP1 with ID2. Cell Death Dis. 2019, 10, 492. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Z.; Liu, Q.; Zhang, G.; Mohammed, D.; Amadou, S.; Tan, G.; Zhang, X. RETRACTED: HOXA11-AS1 Promotes PD-L1-Mediated Immune Escape and Metastasis of Hypopharyngeal Carcinoma by Facilitating PTBP1 and FOSL1 Association. Cancers 2022, 14, 3694. https://doi.org/10.3390/cancers14153694
Zhou Z, Liu Q, Zhang G, Mohammed D, Amadou S, Tan G, Zhang X. RETRACTED: HOXA11-AS1 Promotes PD-L1-Mediated Immune Escape and Metastasis of Hypopharyngeal Carcinoma by Facilitating PTBP1 and FOSL1 Association. Cancers. 2022; 14(15):3694. https://doi.org/10.3390/cancers14153694
Chicago/Turabian StyleZhou, Zheng, Qian Liu, Gehou Zhang, Diab Mohammed, Sani Amadou, Guolin Tan, and Xiaowei Zhang. 2022. "RETRACTED: HOXA11-AS1 Promotes PD-L1-Mediated Immune Escape and Metastasis of Hypopharyngeal Carcinoma by Facilitating PTBP1 and FOSL1 Association" Cancers 14, no. 15: 3694. https://doi.org/10.3390/cancers14153694
APA StyleZhou, Z., Liu, Q., Zhang, G., Mohammed, D., Amadou, S., Tan, G., & Zhang, X. (2022). RETRACTED: HOXA11-AS1 Promotes PD-L1-Mediated Immune Escape and Metastasis of Hypopharyngeal Carcinoma by Facilitating PTBP1 and FOSL1 Association. Cancers, 14(15), 3694. https://doi.org/10.3390/cancers14153694