
HOPX: A Unique Homeodomain Protein in Development and Tumor Suppression


Abstract
Simple Summary
Abstract
1. Introduction
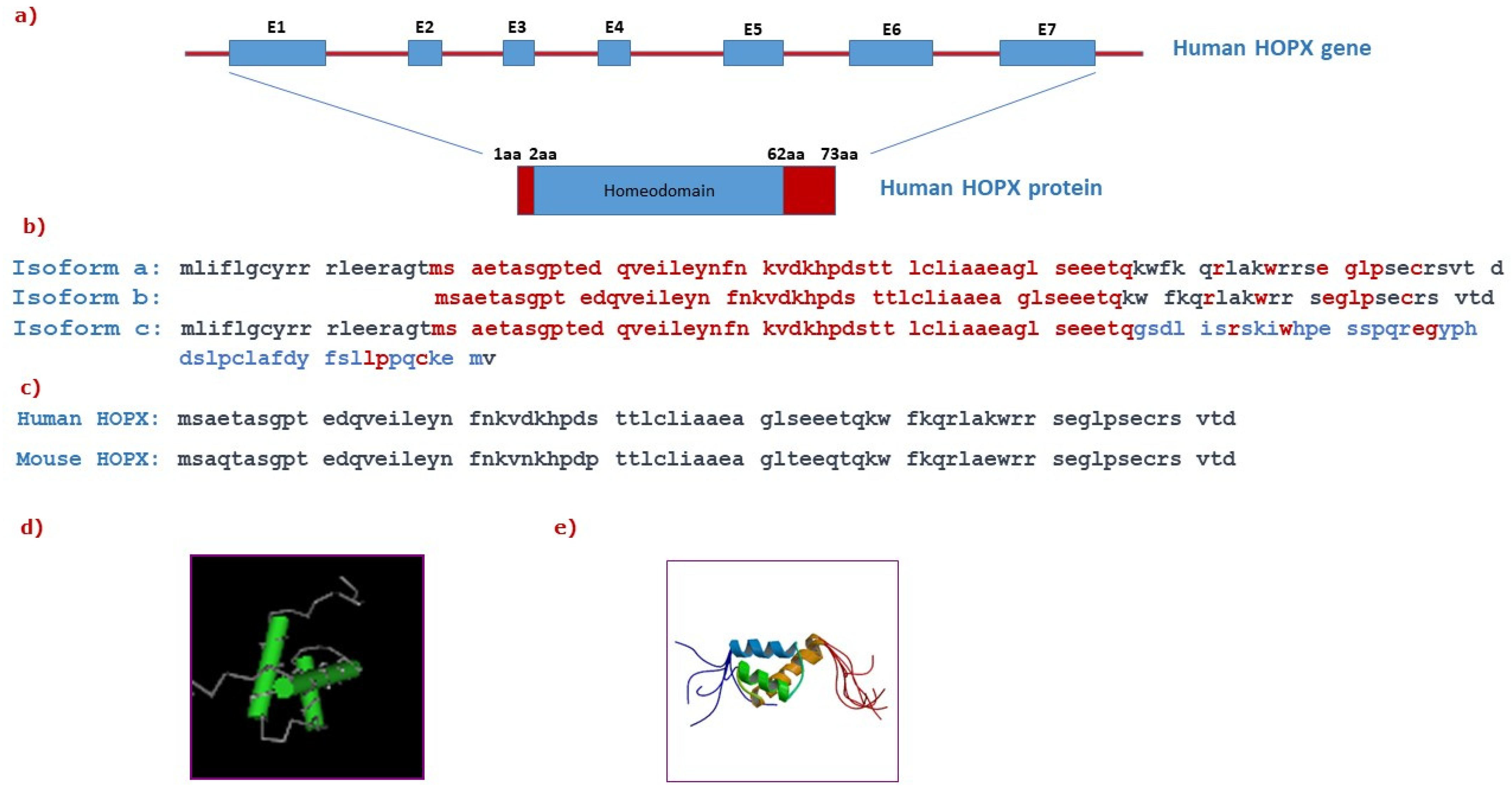
2. HOPX Structure
3. HOPX in Development and Differentiation
3.1. Embryonic Stem Cells
3.2. Adult Stem Cells
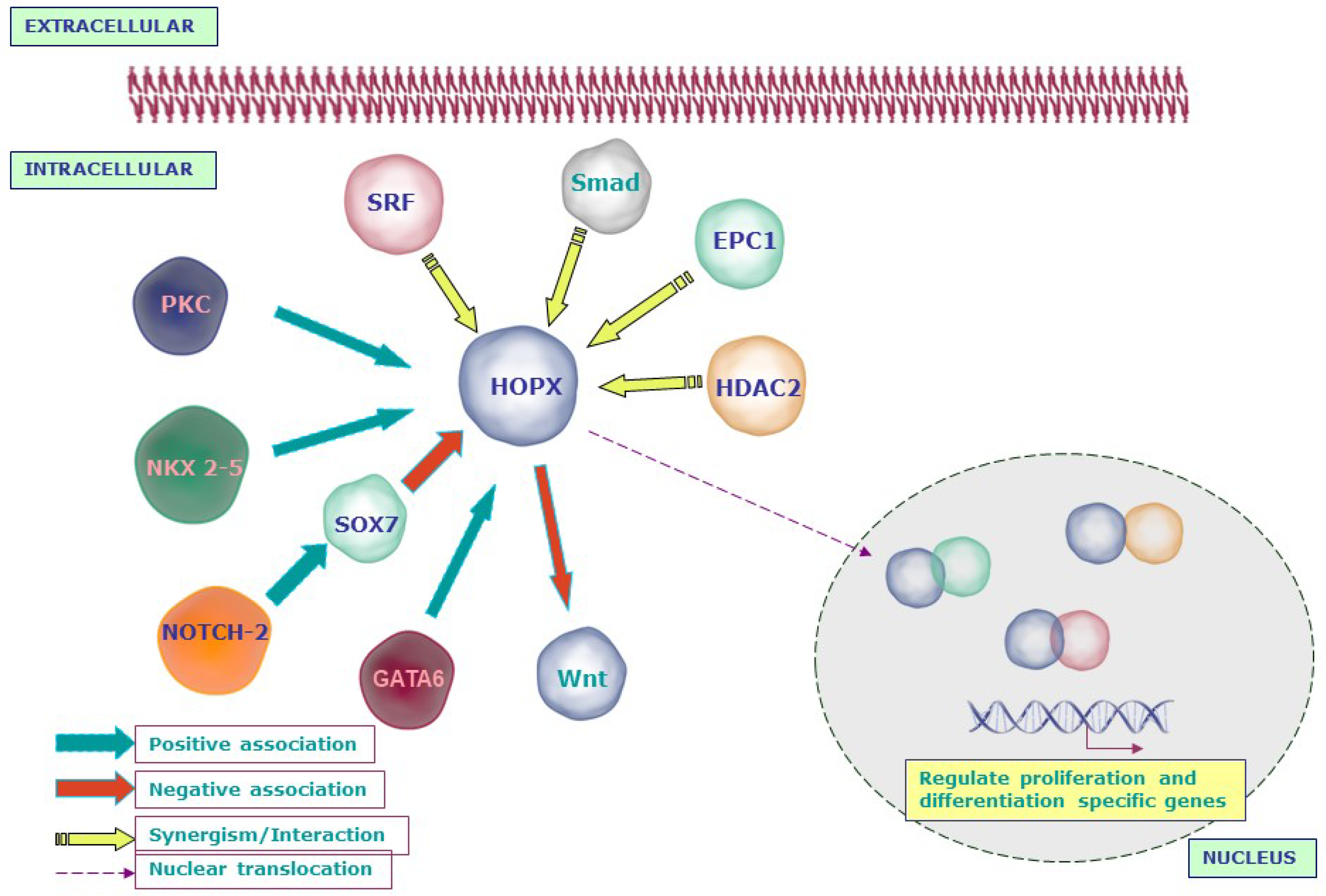
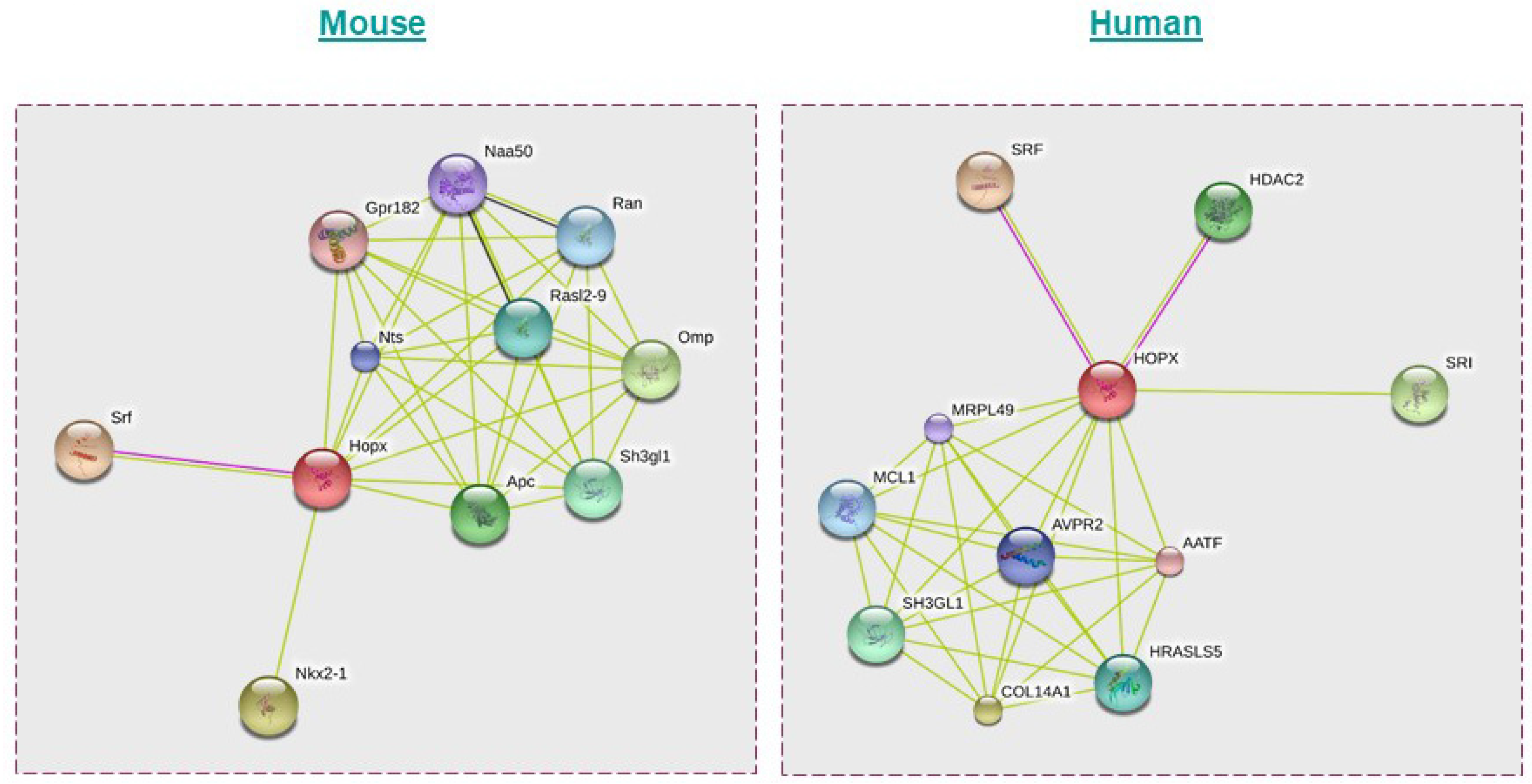
4. HOPX Regulation of Molecular Signaling Networks
5. HOPX in Carcinogenesis
5.1. HOPX as a Tumor Suppressor Gene
5.1.1. HOPX Expression in Normal and Malignant Tissues
5.1.2. HOPX Promoter Methylation
5.1.3. HOPX Affects Tumor Cell Behavior
5.1.4. HOPX and Senescence
5.1.5. HOPX in Cancer Cell Metastasis
5.2. Evidence for an Oncogenic Function for HOPX
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Svingen, T.; Tonissen, K.F. Hox transcription factors and their elusive mammalian gene targets. Heredity 2006, 97, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Desplan, C.; Theis, J.; O’Farrell, P.H. The sequence specificity of homeodomain-DNA interaction. Cell 1988, 54, 1081–1090. [Google Scholar] [CrossRef]
- Lewis, E.B. A gene complex controlling segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Kornberg, T.B. Understanding the homeodomain. J. Biol. Chem. 1993, 268, 26813–26816. [Google Scholar] [CrossRef]
- Boncinelli, E. Homeobox genes and disease. Curr. Opin. Genet. Dev. 1997, 7, 331–337. [Google Scholar] [CrossRef]
- Abate-Shen, C. Deregulated homeobox gene expression in cancer: Cause or consequence? Nat. Rev. Cancer 2002, 2, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Davidson, D. The function and evolution of Msx genes: Pointers and paradoxes. Trends Genet. 1995, 11, 405–411. [Google Scholar] [CrossRef]
- Del Bene, F.; Wittbrodt, J. Cell cycle control by homeobox genes in development and disease. Semin. Cell Dev. Biol. 2005, 16, 449–460. [Google Scholar] [CrossRef]
- Chiou, S.H.; Wang, M.L.; Chou, Y.T.; Chen, C.J.; Hong, C.F.; Hsieh, W.J.; Chang, H.T.; Chen, Y.S.; Lin, T.W.; Hsu, H.S.; et al. Coexpression of Oct4 and Nanog enhances malignancy in lung adenocarcinoma by inducing cancer stem cell-like properties and epithelial-mesenchymal transdifferentiation. Cancer Res. 2010, 70, 10433–10444. [Google Scholar] [CrossRef]
- Samuel, S.; Naora, H. Homeobox gene expression in cancer: Insights from developmental regulation and deregulation. Eur. J. Cancer 2005, 41, 2428–2437. [Google Scholar] [CrossRef]
- Chen, F.; Kook, H.; Milewski, R.; Gitler, A.D.; Lu, M.M.; Li, J.; Nazarian, R.; Schnepp, R.; Jen, K.; Biben, C.; et al. Hop is an unusual homeobox gene that modulates cardiac development. Cell 2002, 110, 713–723. [Google Scholar] [CrossRef]
- Shin, C.H.; Liu, Z.P.; Passier, R.; Zhang, C.L.; Wang, D.Z.; Harris, T.M.; Yamagishi, H.; Richardson, J.A.; Childs, G.; Olson, E.N. Modulation of cardiac growth and development by HOP, an unusual homeodomain protein. Cell 2002, 110, 725–735. [Google Scholar] [CrossRef]
- Adu, J.; Leong, F.T.; Smith, N.R.; Leek, J.P.; Markham, A.F.; Robinson, P.A.; Mighell, A.J. Expression of mOb1, a novel atypical 73 amino acid K50-homeodomain protein, during mouse development. Gene Expr. Patterns 2002, 2, 39–43. [Google Scholar] [CrossRef]
- Asanoma, K.; Matsuda, T.; Kondo, H.; Kato, K.; Kishino, T.; Niikawa, N.; Wake, N.; Kato, H. NECC1, a candidate choriocarcinoma suppressor gene that encodes a homeodomain consensus motif. Genomics 2003, 81, 15–25. [Google Scholar] [CrossRef]
- Chen, Y.; Petersen, S.; Pacyna-Gengelbach, M.; Pietas, A.; Petersen, I. Identification of a novel homeobox-containing gene, LAGY, which is downregulated in lung cancer. Oncology 2003, 64, 450–458. [Google Scholar] [CrossRef]
- Holland, P.W.; Booth, H.A.; Bruford, E.A. Classification and nomenclature of all human homeobox genes. BMC Biol. 2007, 5, 47. [Google Scholar] [CrossRef]
- Burglin, T.R.; Affolter, M. Homeodomain proteins: An update. Chromosoma 2016, 125, 497–521. [Google Scholar] [CrossRef]
- Kook, H.; Yung, W.W.; Simpson, R.J.; Kee, H.J.; Shin, S.; Lowry, J.A.; Loughlin, F.E.; Yin, Z.; Epstein, J.A.; Mackay, J.P. Analysis of the structure and function of the transcriptional coregulator HOP. Biochemistry 2006, 45, 10584–10590. [Google Scholar] [CrossRef]
- Mariotto, A.; Pavlova, O.; Park, H.S.; Huber, M.; Hohl, D. HOPX: The Unusual Homeodomain-Containing Protein. J. Investig. Dermatol. 2016, 136, 905–911. [Google Scholar] [CrossRef]
- Vasiliev, O.; Rhodes, S.J.; Beebe, D.C. Identification and expression of Hop, an atypical homeobox gene expressed late in lens fiber cell terminal differentiation. Mol. Vis. 2007, 13, 114–124. [Google Scholar]
- Asanoma, K.; Kato, H.; Yamaguchi, S.; Shin, C.H.; Liu, Z.P.; Kato, K.; Inoue, T.; Miyanari, Y.; Yoshikawa, K.; Sonoda, K.; et al. HOP/NECC1, a novel regulator of mouse trophoblast differentiation. J. Biol. Chem. 2007, 282, 24065–24074. [Google Scholar] [CrossRef] [PubMed]
- Palpant, N.J.; Wang, Y.; Hadland, B.; Zaunbrecher, R.J.; Redd, M.; Jones, D.; Pabon, L.; Jain, R.; Epstein, J.; Ruzzo, W.L.; et al. Chromatin and Transcriptional Analysis of Mesoderm Progenitor Cells Identifies HOPX as a Regulator of Primitive Hematopoiesis. Cell Rep. 2017, 20, 1597–1608. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Georgantas, R.W., 3rd; Tanadve, V.; Malehorn, M.; Heimfeld, S.; Chen, C.; Carr, L.; Martinez-Murillo, F.; Riggins, G.; Kowalski, J.; Civin, C.I. Microarray and serial analysis of gene expression analyses identify known and novel transcripts overexpressed in hematopoietic stem cells. Cancer Res. 2004, 64, 4434–4441. [Google Scholar] [CrossRef]
- Takeda, N.; Jain, R.; Leboeuf, M.R.; Padmanabhan, A.; Wang, Q.; Li, L.; Lu, M.M.; Millar, S.E.; Epstein, J.A. Hopx expression defines a subset of multipotent hair follicle stem cells and a progenitor population primed to give rise to K6+ niche cells. Development 2013, 140, 1655–1664. [Google Scholar] [CrossRef]
- Barkauskas, C.E.; Cronce, M.J.; Rackley, C.R.; Bowie, E.J.; Keene, D.R.; Stripp, B.R.; Randell, S.H.; Noble, P.W.; Hogan, B.L. Type 2 alveolar cells are stem cells in adult lung. J. Clin. Investig. 2013, 123, 3025–3036. [Google Scholar] [CrossRef] [PubMed]
- Takeda, N.; Jain, R.; LeBoeuf, M.R.; Wang, Q.; Lu, M.M.; Epstein, J.A. Interconversion between intestinal stem cell populations in distinct niches. Science 2011, 334, 1420–1424. [Google Scholar] [CrossRef]
- Roth, S.; Franken, P.; Sacchetti, A.; Kremer, A.; Anderson, K.; Sansom, O.; Fodde, R. Paneth cells in intestinal homeostasis and tissue injury. PLoS ONE 2012, 7, e38965. [Google Scholar] [CrossRef]
- Rees, W.D.; Tandun, R.; Yau, E.; Zachos, N.C.; Steiner, T.S. Regenerative Intestinal Stem Cells Induced by Acute and Chronic Injury: The Saving Grace of the Epithelium? Front. Cell Dev. Biol. 2020, 8, 583919. [Google Scholar] [CrossRef]
- Kawasaki, H.; Yoshida, T.; Horiguchi, K.; Ohama, T.; Sato, K. Characterization of anoikis-resistant cells in mouse colonic epithelium. J. Vet. Med. Sci. 2013, 75, 1173–1180. [Google Scholar] [CrossRef][Green Version]
- Gracz, A.D.; Fuller, M.K.; Wang, F.; Li, L.; Stelzner, M.; Dunn, J.C.; Martin, M.G.; Magness, S.T. Brief report: CD24 and CD44 mark human intestinal epithelial cell populations with characteristics of active and facultative stem cells. Stem Cells 2013, 31, 2024–2030. [Google Scholar] [CrossRef]
- Hng, C.H.; Camp, E.; Anderson, P.; Breen, J.; Zannettino, A.; Gronthos, S. HOPX regulates bone marrow-derived mesenchymal stromal cell fate determination via suppression of adipogenic gene pathways. Sci. Rep. 2020, 10, 11345. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Barkauskas, C.E.; Takeda, N.; Bowie, E.J.; Aghajanian, H.; Wang, Q.; Padmanabhan, A.; Manderfield, L.J.; Gupta, M.; Li, D.; et al. Plasticity of Hopx(+) type I alveolar cells to regenerate type II cells in the lung. Nat. Commun. 2015, 6, 6727. [Google Scholar] [CrossRef] [PubMed]
- Grego-Bessa, J.; Luna-Zurita, L.; del Monte, G.; Bolos, V.; Melgar, P.; Arandilla, A.; Garratt, A.N.; Zang, H.; Mukouyama, Y.S.; Chen, H.; et al. Notch signaling is essential for ventricular chamber development. Dev. Cell 2007, 12, 415–429. [Google Scholar] [CrossRef]
- Zhou, X.; Crow, A.L.; Hartiala, J.; Spindler, T.J.; Ghazalpour, A.; Barsky, L.W.; Bennett, B.J.; Parks, B.W.; Eskin, E.; Jain, R.; et al. The Genetic Landscape of Hematopoietic Stem Cell Frequency in Mice. Stem Cell Rep. 2015, 5, 125–138. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lin, C.C.; Yao, C.Y.; Hsu, Y.C.; Hou, H.A.; Yuan, C.T.; Li, Y.H.; Kao, C.J.; Chuang, P.H.; Chiu, Y.C.; Chen, Y.; et al. Knock-out of Hopx disrupts stemness and quiescence of hematopoietic stem cells in mice. Oncogene 2020, 39, 5112–5123. [Google Scholar] [CrossRef]
- Dorrell, C.; Schug, J.; Lin, C.F.; Canaday, P.S.; Fox, A.J.; Smirnova, O.; Bonnah, R.; Streeter, P.R.; Stoeckert, C.J., Jr.; Kaestner, K.H.; et al. Transcriptomes of the major human pancreatic cell types. Diabetologia 2011, 54, 2832–2844. [Google Scholar] [CrossRef]
- Muhlfriedel, S.; Kirsch, F.; Gruss, P.; Stoykova, A.; Chowdhury, K. A roof plate-dependent enhancer controls the expression of Homeodomain only protein in the developing cerebral cortex. Dev. Biol. 2005, 283, 522–534. [Google Scholar] [CrossRef]
- De Toni, A.; Zbinden, M.; Epstein, J.A.; Ruiz i Altaba, A.; Prochiantz, A.; Caille, I. Regulation of survival in adult hippocampal and glioblastoma stem cell lineages by the homeodomain-only protein HOP. Neural Dev. 2008, 3, 13. [Google Scholar] [CrossRef]
- Liu, F.; Ismat, F.A.; Patel, V.V. Role of homeodomain-only protein in the cardiac conduction system. Trends Cardiovasc. Med. 2006, 16, 193–198. [Google Scholar] [CrossRef][Green Version]
- Kook, H.; Lepore, J.J.; Gitler, A.D.; Lu, M.M.; Wing-Man Yung, W.; Mackay, J.; Zhou, R.; Ferrari, V.; Gruber, P.; Epstein, J.A. Cardiac hypertrophy and histone deacetylase-dependent transcriptional repression mediated by the atypical homeodomain protein Hop. J. Clin. Investig. 2003, 112, 863–871. [Google Scholar] [CrossRef]
- Yin, Z.; Gonzales, L.; Kolla, V.; Rath, N.; Zhang, Y.; Lu, M.M.; Kimura, S.; Ballard, P.L.; Beers, M.F.; Epstein, J.A.; et al. Hop functions downstream of Nkx2.1 and GATA6 to mediate HDAC-dependent negative regulation of pulmonary gene expression. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 291, L191–L199. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Li, D.; Gupta, M.; Manderfield, L.J.; Ifkovits, J.L.; Wang, Q.; Liu, F.; Liu, Y.; Poleshko, A.; Padmanabhan, A.; et al. HEART DEVELOPMENT. Integration of Bmp and Wnt signaling by Hopx specifies commitment of cardiomyoblasts. Science 2015, 348, aaa6071. [Google Scholar] [CrossRef] [PubMed]
- Kee, H.J.; Kim, J.R.; Nam, K.I.; Park, H.Y.; Shin, S.; Kim, J.C.; Shimono, Y.; Takahashi, M.; Jeong, M.H.; Kim, N.; et al. Enhancer of polycomb1, a novel homeodomain only protein-binding partner, induces skeletal muscle differentiation. J. Biol. Chem. 2007, 282, 7700–7709. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Lee, H.J.; Kamli, M.R.; Pokharel, S.; Bhat, A.R.; Lee, Y.H.; Choi, B.H.; Chun, T.; Kang, S.W.; Lee, Y.S.; et al. Depot-specific gene expression profiles during differentiation and transdifferentiation of bovine muscle satellite cells, and differentiation of preadipocytes. Genomics 2012, 100, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.M.; Sim, S.M.; Kim, H.Y.; Park, G.T. Expression of the homeobox gene, HOPX, is modulated by cell differentiation in human keratinocytes and is involved in the expression of differentiation markers. Eur. J. Cell Biol. 2010, 89, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Hawiger, D.; Wan, Y.Y.; Eynon, E.E.; Flavell, R.A. The transcription cofactor Hopx is required for regulatory T cell function in dendritic cell-mediated peripheral T cell unresponsiveness. Nat. Immunol. 2010, 11, 962–968. [Google Scholar] [CrossRef]
- Albrecht, I.; Niesner, U.; Janke, M.; Menning, A.; Loddenkemper, C.; Kuhl, A.A.; Lepenies, I.; Lexberg, M.H.; Westendorf, K.; Hradilkova, K.; et al. Persistence of effector memory Th1 cells is regulated by Hopx. Eur. J. Immunol. 2010, 40, 2993–3006. [Google Scholar] [CrossRef]
- Jones, A.; Opejin, A.; Henderson, J.G.; Gross, C.; Jain, R.; Epstein, J.A.; Flavell, R.A.; Hawiger, D. Peripherally Induced Tolerance Depends on Peripheral Regulatory T Cells That Require Hopx To Inhibit Intrinsic IL-2 Expression. J. Immunol. 2015, 195, 1489–1497. [Google Scholar] [CrossRef]
- Trivedi, C.M.; Cappola, T.P.; Margulies, K.B.; Epstein, J.A. Homeodomain only protein x is down-regulated in human heart failure. J. Mol. Cell Cardiol. 2011, 50, 1056–1058. [Google Scholar] [CrossRef]
- Liu, F.; Levin, M.D.; Petrenko, N.B.; Lu, M.M.; Wang, T.; Yuan, L.J.; Stout, A.L.; Epstein, J.A.; Patel, V.V. Histone-deacetylase inhibition reverses atrial arrhythmia inducibility and fibrosis in cardiac hypertrophy independent of angiotensin. J. Mol. Cell Cardiol. 2008, 45, 715–723. [Google Scholar] [CrossRef]
- Gulec, C.; Abaci, N.; Bayrak, F.; Komurcu Bayrak, E.; Kahveci, G.; Guven, C.; Erginel Unaltuna, N. Association between non-coding polymorphisms of HOPX gene and syncope in hypertrophic cardiomyopathy. Anadolu. Kardiyol. Derg. 2014, 14, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, F.; Millon, R.; Muller, D.; Rabouel, Y.; Bracco, L.; Abecassis, J.; Wasylyk, B. Loss of HOP tumour suppressor expression in head and neck squamous cell carcinoma. Br. J. Cancer 2004, 91, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K.; Kim, M.S.; Park, H.L.; Tokumaru, Y.; Osada, M.; Inoue, H.; Mori, M.; Sidransky, D. HOP/OB1/NECC1 promoter DNA is frequently hypermethylated and involved in tumorigenic ability in esophageal squamous cell carcinoma. Mol. Cancer Res. 2008, 6, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Toruner, G.A.; Ulger, C.; Alkan, M.; Galante, A.T.; Rinaggio, J.; Wilk, R.; Tian, B.; Soteropoulos, P.; Hameed, M.R.; Schwalb, M.N.; et al. Association between gene expression profile and tumor invasion in oral squamous cell carcinoma. Cancer Genet. Cytogenet 2004, 154, 27–35. [Google Scholar] [CrossRef]
- Ooki, A.; Yamashita, K.; Kikuchi, S.; Sakuramoto, S.; Katada, N.; Kokubo, K.; Kobayashi, H.; Kim, M.S.; Sidransky, D.; Watanabe, M. Potential utility of HOP homeobox gene promoter methylation as a marker of tumor aggressiveness in gastric cancer. Oncogene 2010, 29, 3263–3275. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, L.; Cui, T.; Pacyna-Gengelbach, M.; Petersen, I. HOPX is methylated and exerts tumour-suppressive function through Ras-induced senescence in human lung cancer. J. Pathol. 2015, 235, 397–407. [Google Scholar] [CrossRef]
- Cheung, W.K.; Zhao, M.; Liu, Z.; Stevens, L.E.; Cao, P.D.; Fang, J.E.; Westbrook, T.F.; Nguyen, D.X. Control of alveolar differentiation by the lineage transcription factors GATA6 and HOPX inhibits lung adenocarcinoma metastasis. Cancer Cell 2013, 23, 725–738. [Google Scholar] [CrossRef]
- Chen, Y.; Pacyna-Gengelbach, M.; Deutschmann, N.; Niesporek, S.; Petersen, I. Homeobox gene HOP has a potential tumor suppressive activity in human lung cancer. Int. J. Cancer J. Int. Cancer 2007, 121, 1021–1027. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Asanoma, K.; Takao, T.; Kato, K.; Wake, N. Homeobox gene HOPX is epigenetically silenced in human uterine endometrial cancer and suppresses estrogen-stimulated proliferation of cancer cells by inhibiting serum response factor. Int. J. Cancer 2009, 124, 2577–2588. [Google Scholar] [CrossRef]
- Harada, Y.; Kijima, K.; Shinmura, K.; Sakata, M.; Sakuraba, K.; Yokomizo, K.; Kitamura, Y.; Shirahata, A.; Goto, T.; Mizukami, H.; et al. Methylation of the homeobox gene, HOPX, is frequently detected in poorly differentiated colorectal cancer. Anticancer Res. 2011, 31, 2889–2892. [Google Scholar]
- Katoh, H.; Yamashita, K.; Waraya, M.; Margalit, O.; Ooki, A.; Tamaki, H.; Sakagami, H.; Kokubo, K.; Sidransky, D.; Watanabe, M. Epigenetic silencing of HOPX promotes cancer progression in colorectal cancer. Neoplasia 2012, 14, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Waraya, M.; Yamashita, K.; Katoh, H.; Ooki, A.; Kawamata, H.; Nishimiya, H.; Nakamura, K.; Ema, A.; Watanabe, M. Cancer specific promoter CpG Islands hypermethylation of HOP homeobox (HOPX) gene and its potential tumor suppressive role in pancreatic carcinogenesis. BMC Cancer 2012, 12, 397. [Google Scholar] [CrossRef] [PubMed]
- Cromer, A.; Carles, A.; Millon, R.; Ganguli, G.; Chalmel, F.; Lemaire, F.; Young, J.; Dembele, D.; Thibault, C.; Muller, D.; et al. Identification of genes associated with tumorigenesis and metastatic potential of hypopharyngeal cancer by microarray analysis. Oncogene 2004, 23, 2484–2498. [Google Scholar] [CrossRef] [PubMed]
- Roepman, P.; Wessels, L.F.; Kettelarij, N.; Kemmeren, P.; Miles, A.J.; Lijnzaad, P.; Tilanus, M.G.; Koole, R.; Hordijk, G.J.; van der Vliet, P.C.; et al. An expression profile for diagnosis of lymph node metastases from primary head and neck squamous cell carcinomas. Nat. Genet. 2005, 37, 182–186. [Google Scholar] [CrossRef]
- Hunter, K.D.; Thurlow, J.K.; Fleming, J.; Drake, P.J.; Vass, J.K.; Kalna, G.; Higham, D.J.; Herzyk, P.; Macdonald, D.G.; Parkinson, E.K.; et al. Divergent routes to oral cancer. Cancer Res. 2006, 66, 7405–7413. [Google Scholar] [CrossRef]
- Yap, L.F.; Lai, S.L.; Patmanathan, S.N.; Gokulan, R.; Robinson, C.M.; White, J.B.; Chai, S.J.; Rajadurai, P.; Prepageran, N.; Liew, Y.T.; et al. HOPX functions as a tumour suppressor in head and neck cancer. Sci. Rep. 2016, 6, 38758. [Google Scholar] [CrossRef]
- Liang, H.; Wang, C.; Gao, K.; Li, J.; Jia, R. MuicroRNA421 promotes the progression of nonsmall cell lung cancer by targeting HOPX and regulating the Wnt/betacatenin signaling pathway. Mol. Med. Rep. 2019, 20, 151–161. [Google Scholar] [CrossRef]
- Pavlova, O.; Lefort, K.; Mariotto, A.; Huber, M.; Hohl, D. HOPX Exhibits Oncogenic Activity during Squamous Skin Carcinogenesis. J. Investig. Dermatol. 2021, 141, 2354–2368. [Google Scholar] [CrossRef]
- Lin, C.C.; Hsu, Y.C.; Li, Y.H.; Kuo, Y.Y.; Hou, H.A.; Lan, K.H.; Chen, T.C.; Tzeng, Y.S.; Kuo, Y.Y.; Kao, C.J.; et al. Higher HOPX expression is associated with distinct clinical and biological features and predicts poor prognosis in de novo acute myeloid leukemia. Haematologica 2017, 102, 1044–1053. [Google Scholar] [CrossRef]
- Kazanets, A.; Shorstova, T.; Hilmi, K.; Marques, M.; Witcher, M. Epigenetic silencing of tumor suppressor genes: Paradigms, puzzles, and potential. Biochim. Biophys. Acta 2016, 1865, 275–288. [Google Scholar] [CrossRef]
- Ooizumi, Y.; Katoh, H.; Yokota, M.; Watanabe, M.; Yamashita, K. Epigenetic silencing of HOPX is critically involved in aggressive phenotypes and patient prognosis in papillary thyroid cancer. Oncotarget 2019, 10, 5906–5918. [Google Scholar] [CrossRef] [PubMed]
- You, Q.; Geng, Y.; Ye, H.; Zhu, G.; Gao, X.; Zhu, H. HOPX Is an Epigenetically Inactivated Tumor Suppressor and Overexpression of HOPX Induce Apoptosis and Cell Cycle Arrest in Breast Cancer. Onco. Targets Ther. 2020, 13, 5955–5965. [Google Scholar] [CrossRef] [PubMed]
- Minty, F.; Thurlow, J.K.; Harrison, P.R.; Parkinson, E.K. Telomere dysfunction in human keratinocytes elicits senescence and a novel transcription profile. Exp. Cell Res. 2008, 314, 2434–2447. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Pinedo, E.C.; Durham, A.C.; Stewart, K.M.; Goss, A.M.; Lu, M.M.; Demayo, F.J.; Morrisey, E.E. Wnt/beta-catenin signaling accelerates mouse lung tumorigenesis by imposing an embryonic distal progenitor phenotype on lung epithelium. J. Clin. Investig. 2011, 121, 1935–1945. [Google Scholar] [CrossRef] [PubMed]
- Kovarova, D.; Plachy, J.; Kosla, J.; Trejbalova, K.; Cermak, V.; Hejnar, J. Downregulation of HOPX controls metastatic behavior in sarcoma cells and identifies genes associated with metastasis. Mol. Cancer Res. 2013, 11, 1235–1247. [Google Scholar] [CrossRef]
- Ren, X.; Yang, X.; Cheng, B.; Chen, X.; Zhang, T.; He, Q.; Li, B.; Li, Y.; Tang, X.; Wen, X.; et al. HOPX hypermethylation promotes metastasis via activating SNAIL transcription in nasopharyngeal carcinoma. Nat. Commun. 2017, 8, 14053. [Google Scholar] [CrossRef]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; von Mering, C.; et al. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, D808–D815. [Google Scholar] [CrossRef]
- Greif, P.A.; Konstandin, N.P.; Metzeler, K.H.; Herold, T.; Pasalic, Z.; Ksienzyk, B.; Dufour, A.; Schneider, F.; Schneider, S.; Kakadia, P.M.; et al. RUNX1 mutations in cytogenetically normal acute myeloid leukemia are associated with a poor prognosis and up-regulation of lymphoid genes. Haematologica 2012, 97, 1909–1915. [Google Scholar] [CrossRef]
- Santin, A.D.; Zhan, F.; Bignotti, E.; Siegel, E.R.; Cane, S.; Bellone, S.; Palmieri, M.; Anfossi, S.; Thomas, M.; Burnett, A.; et al. Gene expression profiles of primary HPV16- and HPV18-infected early stage cervical cancers and normal cervical epithelium: Identification of novel candidate molecular markers for cervical cancer diagnosis and therapy. Virology 2005, 331, 269–291. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Cancer | Expression Pattern and Methylation Status | Functional Insights | References |
|---|---|---|---|
| Head and Neck cancer |
|
| [52,53,54] |
| Esophageal cancer |
|
| [53] |
| Gastric cancer |
|
| [55] |
| Lung Cancer |
|
| [15,56,57,58] |
| Uterine cancer |
|
| [14,59] |
| Colon cancer |
|
| [60,61] |
| Pancreaticcancer |
|
| [62] |
| Breast cancer |
|
| [59] |
| Placenta (Tropho) |
|
| [14] |
| Glioblastoma |
|
| [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caspa Gokulan, R.; Yap, L.F.; Paterson, I.C. HOPX: A Unique Homeodomain Protein in Development and Tumor Suppression. Cancers 2022, 14, 2764. https://doi.org/10.3390/cancers14112764
Caspa Gokulan R, Yap LF, Paterson IC. HOPX: A Unique Homeodomain Protein in Development and Tumor Suppression. Cancers. 2022; 14(11):2764. https://doi.org/10.3390/cancers14112764
Chicago/Turabian StyleCaspa Gokulan, Ravindran, Lee Fah Yap, and Ian C. Paterson. 2022. "HOPX: A Unique Homeodomain Protein in Development and Tumor Suppression" Cancers 14, no. 11: 2764. https://doi.org/10.3390/cancers14112764
APA StyleCaspa Gokulan, R., Yap, L. F., & Paterson, I. C. (2022). HOPX: A Unique Homeodomain Protein in Development and Tumor Suppression. Cancers, 14(11), 2764. https://doi.org/10.3390/cancers14112764