
Lymph Nodes as Anti-Tumor Immunotherapeutic Tools: Intranodal-Tumor-Specific Antigen-Pulsed Dendritic Cell Vaccine Immunotherapy


Abstract
:Simple Summary
Abstract
1. Introduction
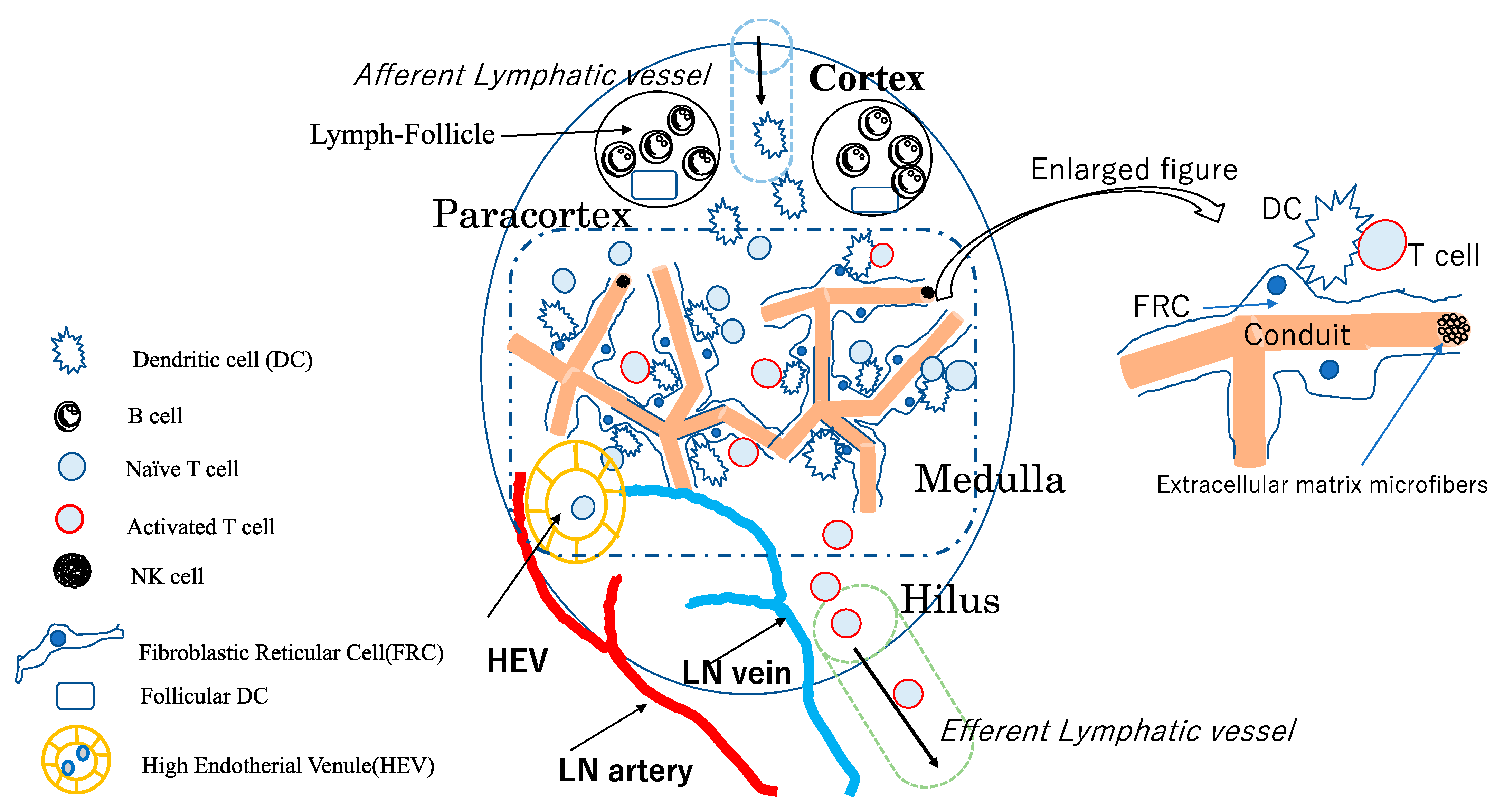
2. Structure of LNs and Their Constituent Cells
3. Tumor Immune Microenvironment and Local Immunotherapy in Tumor Drainage LNs
4. DC Vaccine Therapy
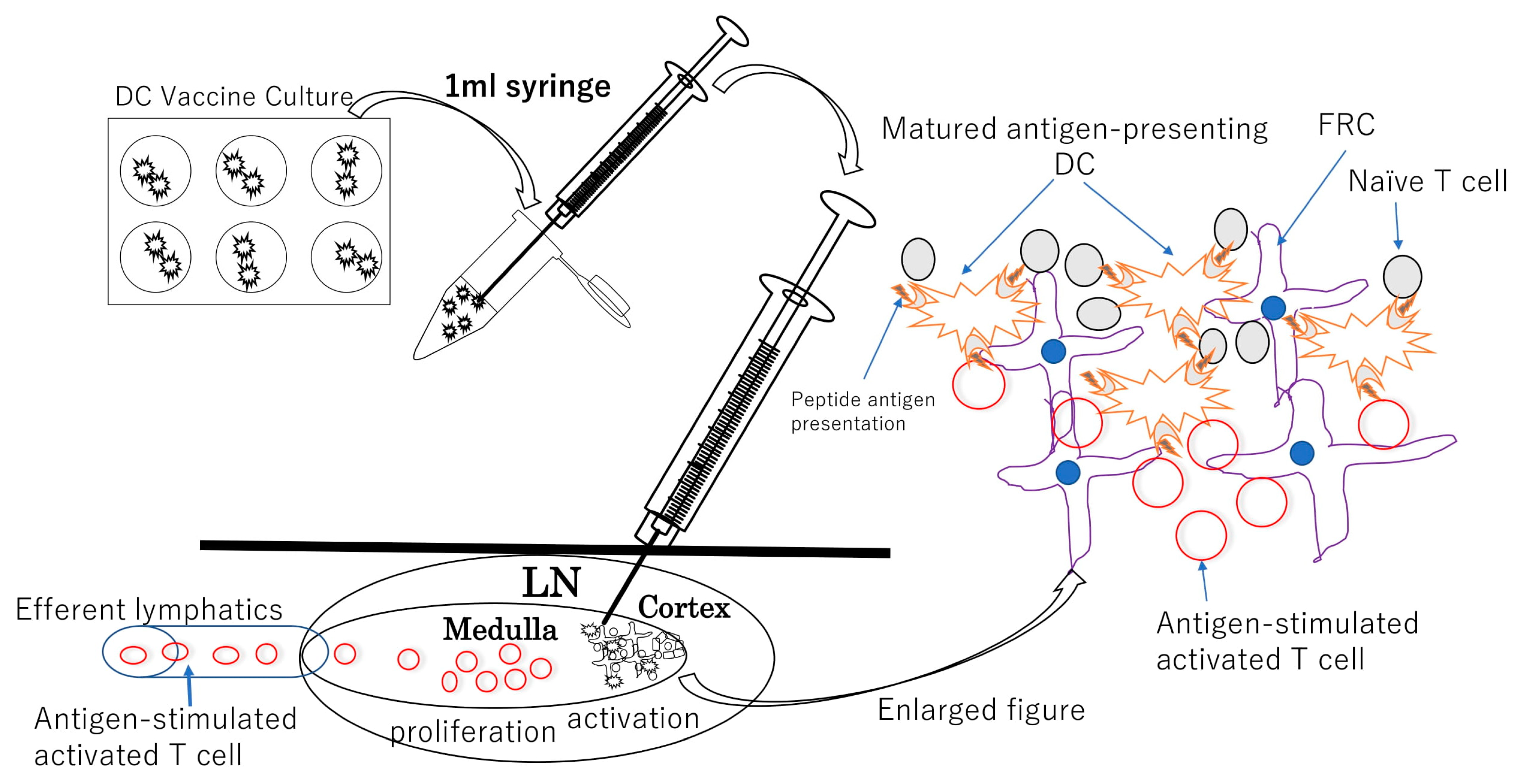
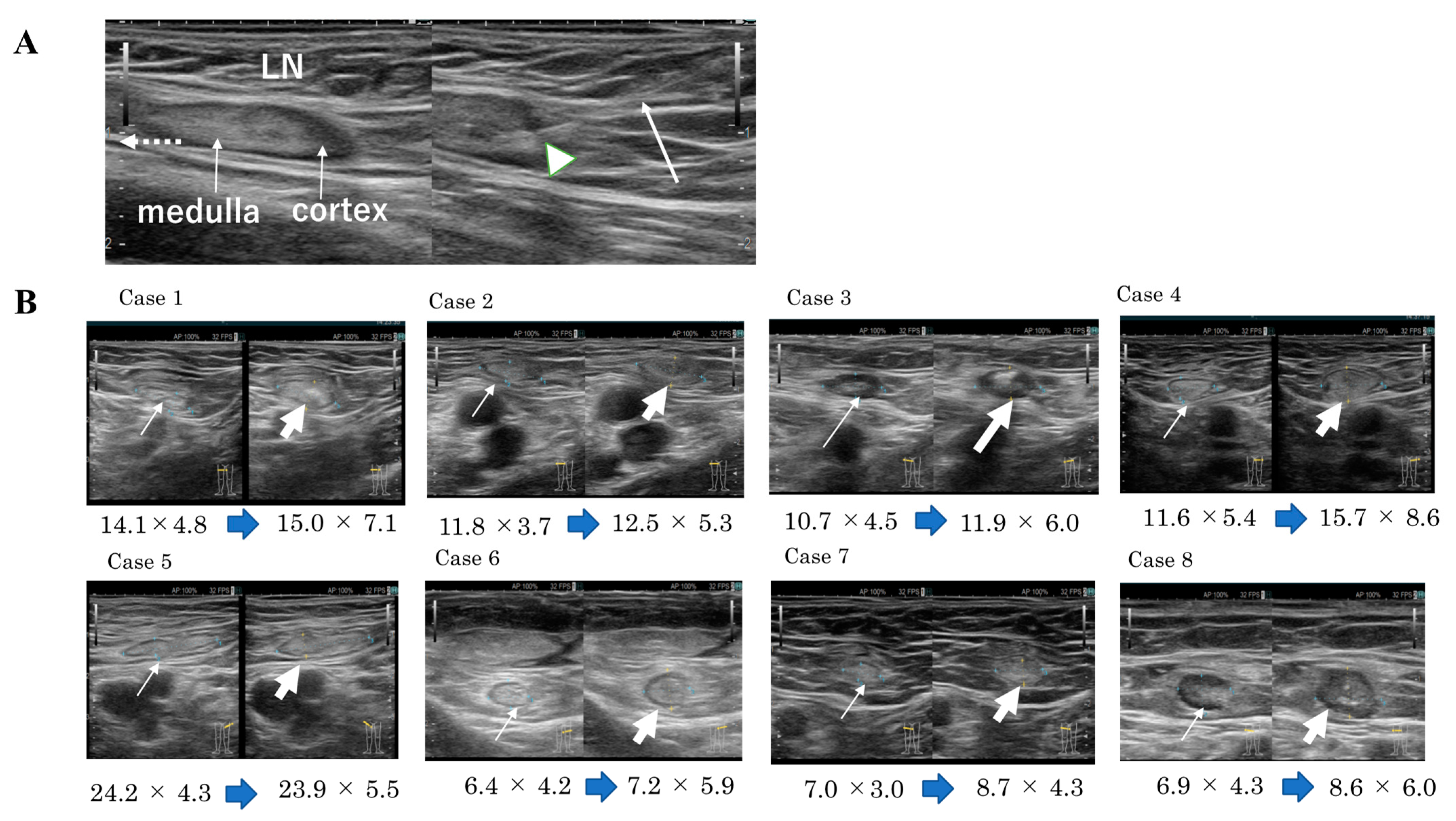
5. The Rationale for Intranodal Administration of DCs and Intranodal Contrast Injection
6. DC Vaccine Intranodal Infusion Therapy

{kind=link}
{kind=link}
{kind=link}
| Reference | Study Type | Tumor | Antigens | Adjuvant/Stimulant | Results | Reference |
|---|---|---|---|---|---|---|
| Nestle FO et al., 1998 | Clinical | Melanoma | Tumor Lysate. MelanA/Mart1, Mage1/Mage3 | KLH | 2 CR and 3 PR in 16 patients | [41] |
| Lambert LA et al., 2001 | Pre-clinical(mice) | Melanoma | Tumor Lysate | ( - ) | IN superior than ID in Th1 response | [50] |
| Bedrisian I et al., 2003 | Clinical | Melanoma | Peptieds for Mart1, gp100, Tyrosinase | ( - ) | IN superior than ID in Th1 response | [51] |
| Jonuleit H et al., 2001 | Clinical | Melanoma | MelanA/Mart1, Mage1/Mage3 | ( - ) | in case of matured DC CD4 response 7/8 CD8 response 5/7 | [42] |
| Gilliet M et al., 2003 | Clinical | Melanoma | MelanA/Mart1, Mage1/Mage3 | KLH | Long lasting CD4T-cell response with TH-1 cytokine response in all 5 patients | [43] |
| Maier T et al., 2003 | Clinical | Lymphoma | Tumor Lysate | KLH | 4 PR and 1 CR in 8 patients | [44] |
| Schwaab TS et al., 2009 | Clinical | Renal Cell Cancer | Tumor Lysate | IFN-a2a, IL-2 | 50% ORR and 3 CR in 18 patients | [45] |
| Yi Q et al., 2010 | Clinical | Multiple Myeloma | Idiotype protein | KLH, CD40 | SD in 6 of 9 patients | [49] |
| Barth RJ et al., 2010 | Clinical | Colorectal cancer | Tumor Lysate | KLH, CD40 | 61% DTH response in 24 patients. 5year recurrence free 63% | [46] |
| Aarntzen EHJG et al., 2012 | Clinical | Melanoma | mRNA for gp100/Tyrosinase | KLH | TAA specific Th1 responsein stage III: median OR 24.1 months | [47] |
| Bol KF et al., 2015 | Clinical | Melanoma | mRNA for gp100/Tyrosinase | CD40,/TLR4 mRNA | 1 MR and 2 durable SD in 8 patients | [48] |
| Morisaki T et al., 2020 | Clinical | Ovarian Cancer | Neoantigen peptides | ( - ) | 1 case report: durable SD | [55] |
| Morisaki T et al., 2021 | Clnical | Solid tumors | Neoantigen peptides | ( - ) | 1 CR, 3 PR, and 10 SD in 17 patietns | [56] |
7. Techniques of Intranodal Administration of Mature DCs
8. Intranodal Neoantigen Peptide-Pulsed DC Vaccine Therapy
9. Conclusions and Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Foster, R.S., Jr. The Biologic and Clinical Significance of Lymphatic Metastases in Breast Cancer. Surg. Oncol. Clin. N. Am. 1996, 5, 79–104. [Google Scholar] [CrossRef]
- McGuire, W.L. Prognostic Factors for Recurrence and Survival in Human Breast Cancer. Breast Cancer Res. Treat. 1987, 10, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Märkl, B. Stage migration vs immunology: The lymph node story in colon cancer. World J. Gastroenterol. 2015, 21, 12218–12233. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, S.; Stephenson, K. More is better: Lymph node harvesting in colorectal cancer. Am. J. Surg. 2017, 213, 926–930. [Google Scholar] [CrossRef]
- Harrison, J.C.; Dean, P.J.; el-Zeky, F.; Vander Zwaag, R. From Dukes through Jass: Pathological prognostic indicators in rectal cancer. Hum. Pathol. 1994, 25, 498–505. [Google Scholar] [CrossRef]
- Balch, C.M.; Soong, S.J.; Gershenwald, J.E.; Thompson, J.F.; Reintgen, D.S.; Cascinelli, N.; Urist, M.; McMasters, K.M.; Ross, M.I.; Kirkwood, J.M.; et al. Prognostic factors analysis of 17600 melanoma patients: Validation of the American Joint Committee on Cancer melanoma staging system. J. Clin. Oncol. 2001, 19, 3622–3634. [Google Scholar] [CrossRef]
- Pereira, E.R.; Jones, D.; Jung, K.; Padera, T.P. The Lymph Node Microenvironment and Its Role in the Progression of Metastatic Cancer. Semin. Cell Dev. Biol. 2015, 38, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, G.; Ataide, M.; Kastenmüller, W. Lymph Node—An Organ for T-Cell Activation and Pathogen Defense. Immunol. Rev. 2016, 271, 200–220. [Google Scholar] [CrossRef]
- Sainte-Marie, G. The lymph node revisited: Development, morphology, functioning, and role in triggering primary immune responses. Anat. Rec. 2010, 293, 320–337. [Google Scholar] [CrossRef]
- Fransen, M.F.; Schoonderwoerd, M.; Knopf, P.; Camps, M.G.; Hawinkels, L.J.; Kneilling, M.; van Hall, T.; Ossendorp, F. Tumor-Draining Lymph Nodes Are Pivotal in PD-1/PD-L1 Checkpoint Therapy. JCI Insight. 2018, 6, e124507. [Google Scholar] [CrossRef] [Green Version]
- Inamori, K.; Togashi, Y.; Fukuoka, S.; Akagi, K.; Ogasawara, K.; Irie, T.; Motooka, D.; Kobayashi, Y.; Sugiyama, D.; Kojima, M.; et al. Importance of Lymph Node Immune Responses in MSI-H/dMMR Colorectal Cancer. JCI Insight. 2021, 10, e137365. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.; Wiechers, C.; Huehn, J. Lymph Node Stromal Cell Subsets-Emerging Specialists for Tailored Tissue-Specific Immune Responses. Int. J. Med. Microbiol. 2021, 311, 151492. [Google Scholar] [CrossRef] [PubMed]
- Acton, S.E.; Farrugia, A.J.; Astarita, J.L.; Mourão-Sá, D.; Jenkins, R.P.; Nye, E.; Hooper, S.; van Blijswijk, J.; Rogers, N.C.; Snelgrove, K.J.; et al. Dendritic Cells Control Fibroblastic Reticular Network Tension and Lymph Node Expansion. Nature 2014, 514, 498–502. [Google Scholar] [CrossRef]
- Tay, B.Q.; Wright, Q.; Ladwa, R.; Perry, C.; Leggatt, G.; Simpson, F.; Wells, J.W.; Panizza, B.J.; Frazer, I.H.; Cruz, J.L.G. Evolution of Cancer Vaccines-Challenges, Achievements, and Future Directions. Vaccines 2021, 9, 535. [Google Scholar] [CrossRef] [PubMed]
- Willard-Mack, C.L. Normal Structure, Function, and Histology of lymph nodes. Toxicol. Pathol. 2006, 34, 409–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arasa, J.; Collado-Diaz, V.; Halin, C. Structure and Immune Function of Afferent Lymphatics and Their Mechanistic Contribution to Dendritic Cell and T Cell Trafficking. Cells 2021, 10, 1269. [Google Scholar] [CrossRef] [PubMed]
- Acton, S.E.; Onder, L.; Novkovic, M.; Martinez, V.G.; Ludewig, B. Communication, construction, and fluid control: Lymphoid organ fibroblastic reticular cell and conduit networks. Trends Immunol. 2021, 42, 782–794. [Google Scholar] [CrossRef]
- Onder, L.; Narang, P.; Scandella, E.; Chai, Q.; Iolyeva, M.; Hoorweg, K.; Halin, C.; Richie, E.; Kaye, P.; Westermann, J.; et al. IL-7-producing stromal cells are critical for lymph node remodeling. Blood 2012, 120, 4675–4683. [Google Scholar] [CrossRef]
- Ager, A.; May, M.J. Understanding high endothelial venules: Lessons for cancer immunology. Oncoimmunology 2015, 4, e1008791. [Google Scholar] [CrossRef] [Green Version]
- Acton, S.E.; Reise Sousa, C. Dendritic cells in remodeling of lymph nodes during immune responses. Immunol. Rev. 2016, 271, 221–229. [Google Scholar] [CrossRef]
- Arneth, B. Tumor microenvironment. Medicina 2019, 56, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, R.-C. Lymph Nodes and Cancer Metastasis: New Perspectives on the Role of Intranodal Lymphatic Sinuses. Int. J. Mol. Sci. 2016, 18, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Lei, P.J.; Padera, T.P. Progression of Metastasis through Lymphatic System. Cells 2021, 10, 627. [Google Scholar] [CrossRef] [PubMed]
- Goode, E.F.; Roussos Torres, E.T.; Irshad, S. Lymph Node Immune Profiles as Predictive Biomarkers for Immune Checkpoint Inhibitor Response. Front. Mol. Biosci. 2021, 8, 674558. [Google Scholar] [CrossRef]
- Jana, S.; Muscarella, R.A., Jr.; Jones, D. The Multifaceted Effects of Breast Cancer on Tumor-Draining Lymph Nodes. Am. J. Pathol. 2021, 191, 1353–1363. [Google Scholar] [CrossRef]
- Fransen, M.F.; van Hall, T.; Ossendorp, F. Immune Checkpoint Therapy: Tumor Draining Lymph Nodes in the Spotlights. Int. J. Mol. Sci. 2021, 22, 9401. [Google Scholar] [CrossRef]
- van Pul, K.M.; Fransen, M.F.; van de Ven, R.; de Gruijl, T.D. Immunotherapy Goes Local: The Central Role of Lymph Nodes in Driving Tumor Infiltration and Efficacy. Front. Immunol. 2021, 12, 643291. [Google Scholar] [CrossRef]
- Melero, I.; Castanon, E.; Alvarez, M.; Champiat, S.; Marabelle, A. Intratumoural Administration and Tumour Tissue Targeting of Cancer Immunotherapies. Nat. Rev. Clin. Oncol. 2021, 18, 558–576. [Google Scholar] [CrossRef]
- Liu, H.; Moynihan, K.D.; Zheng, Y.; Szeto, G.L.; Li, A.V.; Huang, B.; Van Egeren, D.S.; Park, C.; Irvine, D.J. Structure-Based Programming of Lymph-Node Targeting in Molecular Vaccines. Nature 2014, 507, 519–522. [Google Scholar] [CrossRef] [Green Version]
- Schudel, A.; Chapman, A.P.; Yau, M.K.; Higginson, C.J.; Francis, D.M.; Manspeaker, M.P.; Avecilla, A.R.C.; Rohner, N.A.; Finn, M.G.; Thomas, S.N. Programmable Multistage Drug Delivery to Lymph Nodes. Nat. Nanotechnol. 2020, 15, 491–499. [Google Scholar] [CrossRef]
- Palucka, K.; Banchereau, J. Cancer Immunotherapy via Dendritic Cells. Nat. Rev. Cancer 2012, 12, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.D.; Vara Perez, M.; Schaaf, M.; Agostinis, P.; Zitvogel, L.; Kroemer, G.; Galluzzi, L. Trial Watch: Dendritic Cell-Based Anticancer Immunotherapy. Oncoimmunology 2017, 6, e1328341. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; Bhardwaj, N. Re-emergence of dendritic cell vaccines for cancer treatment. Trends Cancer 2018, 4, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Postow, M.A.; Sidlow, R.; Hellmann, M.D. Immune-Related Adverse Events Associated with Immune Checkpoint Blockade. N. Engl. J. Med. 2018, 378, 158–168. [Google Scholar] [CrossRef]
- Adashek, J.J.; Kato, S.; Ferrara, R.; Lo Russo, G.; Kurzrock, R. Hyperprogression and Immune Checkpoint Inhibitors: Hype or Progress? Oncologist 2020, 25, 94–98. [Google Scholar] [CrossRef] [Green Version]
- Quillien, V.; Moisan, A.; Carsin, A.; Lesimple, T.; Lefeuvre, C.; Adamski, H.; Bertho, N.; Devillers, A.; Leberre, C.; Toujas. Biodistribution of Radiolabelled Human Dendritic Cells Injected by Various Routes. Eur. J. Nucl. Med. Mol. Imaging. 2005, 32, 731–741. [Google Scholar] [CrossRef]
- Rajebi, M.R.; Chaudry, G.; Padua, H.M.; Dillon, B.; Yilmaz, S.; Arnold, R.W.; Landrigan-Ossar, M.F.; Alomari, A.I. Intranodal Lymphangiography: Feasibility and Preliminary Experience in Children. J. Vasc. Interv. Radiol. 2011, 22, 1300–1305. [Google Scholar] [CrossRef]
- Nadolski, G.J.; Itkin, M. Feasibility of Ultrasound-Guided Intranodal Lymphangiogram for Thoracic Duct Embolization. J. Vasc. Interv. Radiol. 2012, 23, 613–616. [Google Scholar] [CrossRef]
- Sommer, C.M.; Pieper, C.C.; Itkin, M.; Nadolski, G.J.; Hur, S.; Kim, J.; Maleux, G.; Kauczor, H.U.; Richter, G.M. Conventional Lymphangiography (CL) in the Management of Postoperative Lymphatic Leakage (PLL): A Systematic Review. RöFo 2020, 192, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhou, F.; Bai, X.; Yuan, Q.; Zhang, M.; Ma, L.; Jin, Y. Application of Ultrasound-Guided Intranodal Lymphangiography and Embolisation in Cancer Patients with Postoperative Lymphatic Leakage. World J. Surg. Oncol. 2021, 19, 32. [Google Scholar] [CrossRef]
- Nestle, F.O.; Alijagic, S.; Gilliet, M.; Sun, Y.; Grabbe, S.; Dummer, R.; Burg, G.; Schadendorf, D. Vaccination of Melanoma Patients with Peptide- or Tumor Lysate-Pulsed Dendritic Cells. Nat. Med. 1998, 4, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Jonuleit, H.; Giesecke-Tuettenberg, A.; Tüting, T.; Thurner-Schuler, B.; Stuge, T.B.; Paragnik, L.; Kandemir, A.; Lee, P.P.; Schuler, G.; Knop, J.; et al. A Comparison of Two Types of Dendritic Cell as Adjuvants for the Induction of Melanoma-Specific T-Cell Responses in Humans Following Intranodal Injection. Int. J. Cancer. 2001, 93, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Gilliet, M.; Kleinhans, M.; Lantelme, E.; Schadendorf, D.; Burg, G.; Nestle, F.O. Intranodal Injection of Semimature Monocyte-Derived Dendritic Cells Induces T Helper Type 1 Responses to Protein Neoantigen. Blood 2003, 102, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Maier, T.; Tun-Kyi, A.; Tassis, A.; Jungius, K.P.; Burg, G.; Dummer, R.; Nestle, F.O. Vaccination of Patients with Cutaneous T-Cell Lymphoma Using Intranodal Injection of Autologous Tumor-Lysate-Pulsed Dendritic Cells. Blood 2003, 102, 2338–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwaab, T.; Schwarzer, A.; Wolf, B.; Crocenzi, T.S.; Seigne, J.D.; Crosby, N.A.; Cole, B.F.; Fisher, J.L.; Uhlenhake, J.C.; Mellinger, D.; et al. Clinical and Immunologic Effects of Intranodal Autologous Tumor Lysate-Dendritic Cell Vaccine with Aldesleukin (Interleukin 2) and IFN-{Alpha}2a Therapy in Metastatic Renal Cell Carcinoma Patients. Clin. Cancer Res. 2009, 15, 4986–4992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, R.J., Jr.; Fisher, D.A.; Wallace, P.K.; Channon, J.Y.; Noelle, R.J.; Gui, J.; Ernstoff, M.S. A Randomized Trial of Ex Vivo CD40L Activation of a Dendritic Cell Vaccine in Colorectal Cancer Patients: Tumor-Specific Immune Responses Are Associated with Improved Survival. Clin. Cancer Res. 2010, 16, 5548–5556. [Google Scholar] [CrossRef] [Green Version]
- Aarntzen, E.H.; Schreibelt, G.; Bol, K.; Lesterhuis, W.J.; Croockewit, A.J.; de Wilt, J.H.; van Rossum, M.M.; Blokx, W.A.; Jacobs, J.F.; Duiveman-de Boer, T.; et al. Vaccination with mRNA-Electroporated Dendritic Cells Induces Robust Tumor Antigen-Specific CD4+ and CD8+ T Cells Responses in stage III and IV Melanoma Patients. Clin. Cancer Res. 2012, 18, 5460–5470. [Google Scholar] [CrossRef] [Green Version]
- Bol, K.F.; Figdor, C.G.; Aarntzen, E.H.; Welzen, M.E.; van Rossum, M.M.; Blokx, W.A.; van de Rakt, M.W.; Scharenborg, N.M.; de Boer, A.J.; Pots, J.M.; et al. Intranodal Vaccination with mRNA-Optimized Dendritic Cells in Metastatic Melanoma Patients. Oncoimmunology 2015, 4, e1019197. [Google Scholar] [CrossRef]
- Yi, Q.; Szmania, S.; Freeman, J.; Qian, J.; Rosen, N.A.; Viswamitra, S.; Cottler-Fox, M.; Barlogie, B.; Tricot, G.; van Rhee, F. Optimizing Dendritic Cell-Based Immunotherapy in Multiple Myeloma: Intranodal Injections of Idiotype-Pulsed CD40 Ligand-Matured Vaccines Led to Induction of type-1 and Cytotoxic T-Cell Immune Responses in Patients. Br. J. Haematol. 2010, 150, 554–564. [Google Scholar] [CrossRef] [Green Version]
- Lambert, L.A.; Gibson, G.R.; Maloney, M.; Durell, B.; Noelle, R.J.; Barth, R.J., Jr. Intranodal Immunization with Tumor Lysate-Pulsed Dendritic Cells Enhances Protective Antitumor Immunity. Cancer Res. 2001, 61, 641–646. [Google Scholar]
- Bedrosian, I.; Mick, R.; Xu, S.; Nisenbaum, H.; Faries, M.; Zhang, P.; Cohen, P.A.; Koski, G.; Czerniecki, B.J. Intranodal Administration of Peptide-Pulsed Mature Dendritic Cell Vaccines Results in Superior CD8+ T-Cell Function in Melanoma Patients. J. Clin. Oncol. 2003, 21, 3826–3835. [Google Scholar] [CrossRef] [PubMed]
- Lesterhuis, W.J.; de Vries, I.J.; Schreibelt, G.; Lambeck, A.J.; Aarntzen, E.H.; Jacobs, J.F.; Scharenborg, N.M.; van de Rakt, M.W.; de Boer, A.J.; Croockewit, S.; et al. Route of Administration Modulates the Induction of Dendritic Cell Vaccine-Induced Antigen-Specific T Cells in Advanced Melanoma Patients. Clin. Cancer Res. 2011, 17, 5725–5735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyte, J.A.; Aamdal, S.; Dueland, S.; Sæbøe-Larsen, S.; Inderberg, E.M.; Madsbu, U.E.; Skovlund, E.; Gaudernack, G.; Kvalheim, G. Immune Response and Long-Term Clinical Outcome in Advanced Melanoma Patients Vaccinated with Tumor-mRNA-Transfected Dendritic Cells. Oncoimmunology 2016, 5, e1232237. [Google Scholar] [CrossRef] [PubMed]
- Lowenfeld, L.; Mick, R.; Datta, J.; Xu, S.; Fitzpatrick, E.; Fisher, C.S.; Fox, K.R.; DeMichele, A.; Zhang, P.J.; Weinstein, S.P.; et al. Dendritic Cell Vaccination Enhances Immune Responses and Induces Regression of HER2(Pos) DCIS Independent of Route: Results of Randomized Selection Design Trial. Clin. Cancer Res. 2017, 23, 2961–2971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morisaki, T.; Hikichi, T.; Onishi, H.; Morisaki, T.; Kubo, M.; Hirano, T.; Yoshimura, S.; Kiyotani, K.; Nakamura, Y. Intranodal Administration of Neoantigen Peptide-Loaded Dendritic Cell Vaccine Elicits Epitope-Specific T Cell Responses and Clinical Effects in a Patient with Chemorefractory Ovarian Cancer with Malignant Ascites. Immunol. Investig. 2021, 50, 562–579. [Google Scholar] [CrossRef] [PubMed]
- Morisaki, T.; Morisaki, T.; Kubo, M.; Onishi, H.; Hirano, T.; Morisaki, S.; Eto, M.; Monji, K.; Takeuchi, A.; Nakagawa, S.; et al. Efficacy of Intranodal Neoantigen Peptide-Pulsed Dendritic Cell Vaccine Monotherapy in Patients with Advanced Solid Tumors: A Retrospective Analysis. Anticancer Res. 2021, 41, 4101–4115. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, T.N.; Scheper, W.; Kvistborg, P. Cancer Neoantigens. Annu. Rev. Immunol. 2019, 37, 173–200. [Google Scholar] [CrossRef]
- Blass, E.; Ott, P.A. Advances in the development of personalized neoantigen-based therapeutic cancer vaccines. Nat. Rev. Clin. Oncol. 2021, 18, 215–229. [Google Scholar] [CrossRef]
- Ott, P.A.; Hu-Lieskovan, S.; Chmielowski, B.; Govindan, R.; Naing, A.; Bhardwaj, N.; Margolin, K.; Awad, M.M.; Hellmann, M.D.; Lin, J.J.; et al. A Phase Ib Trial of Personalized Neoantigen Therapy plus Anti-PD-1 in Patients with Advanced Melanoma, Non-Small Cell Lung Cancer, or Bladder Cancer. Cells 2020, 183, 347–362.e24. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morisaki, T.; Morisaki, T.; Kubo, M.; Morisaki, S.; Nakamura, Y.; Onishi, H. Lymph Nodes as Anti-Tumor Immunotherapeutic Tools: Intranodal-Tumor-Specific Antigen-Pulsed Dendritic Cell Vaccine Immunotherapy. Cancers 2022, 14, 2438. https://doi.org/10.3390/cancers14102438
Morisaki T, Morisaki T, Kubo M, Morisaki S, Nakamura Y, Onishi H. Lymph Nodes as Anti-Tumor Immunotherapeutic Tools: Intranodal-Tumor-Specific Antigen-Pulsed Dendritic Cell Vaccine Immunotherapy. Cancers. 2022; 14(10):2438. https://doi.org/10.3390/cancers14102438
Chicago/Turabian StyleMorisaki, Takashi, Takafumi Morisaki, Makoto Kubo, Shinji Morisaki, Yusuke Nakamura, and Hideya Onishi. 2022. "Lymph Nodes as Anti-Tumor Immunotherapeutic Tools: Intranodal-Tumor-Specific Antigen-Pulsed Dendritic Cell Vaccine Immunotherapy" Cancers 14, no. 10: 2438. https://doi.org/10.3390/cancers14102438
APA StyleMorisaki, T., Morisaki, T., Kubo, M., Morisaki, S., Nakamura, Y., & Onishi, H. (2022). Lymph Nodes as Anti-Tumor Immunotherapeutic Tools: Intranodal-Tumor-Specific Antigen-Pulsed Dendritic Cell Vaccine Immunotherapy. Cancers, 14(10), 2438. https://doi.org/10.3390/cancers14102438