
Can Schlafen 11 Help to Stratify Ovarian Cancer Patients Treated with DNA-Damaging Agents?

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
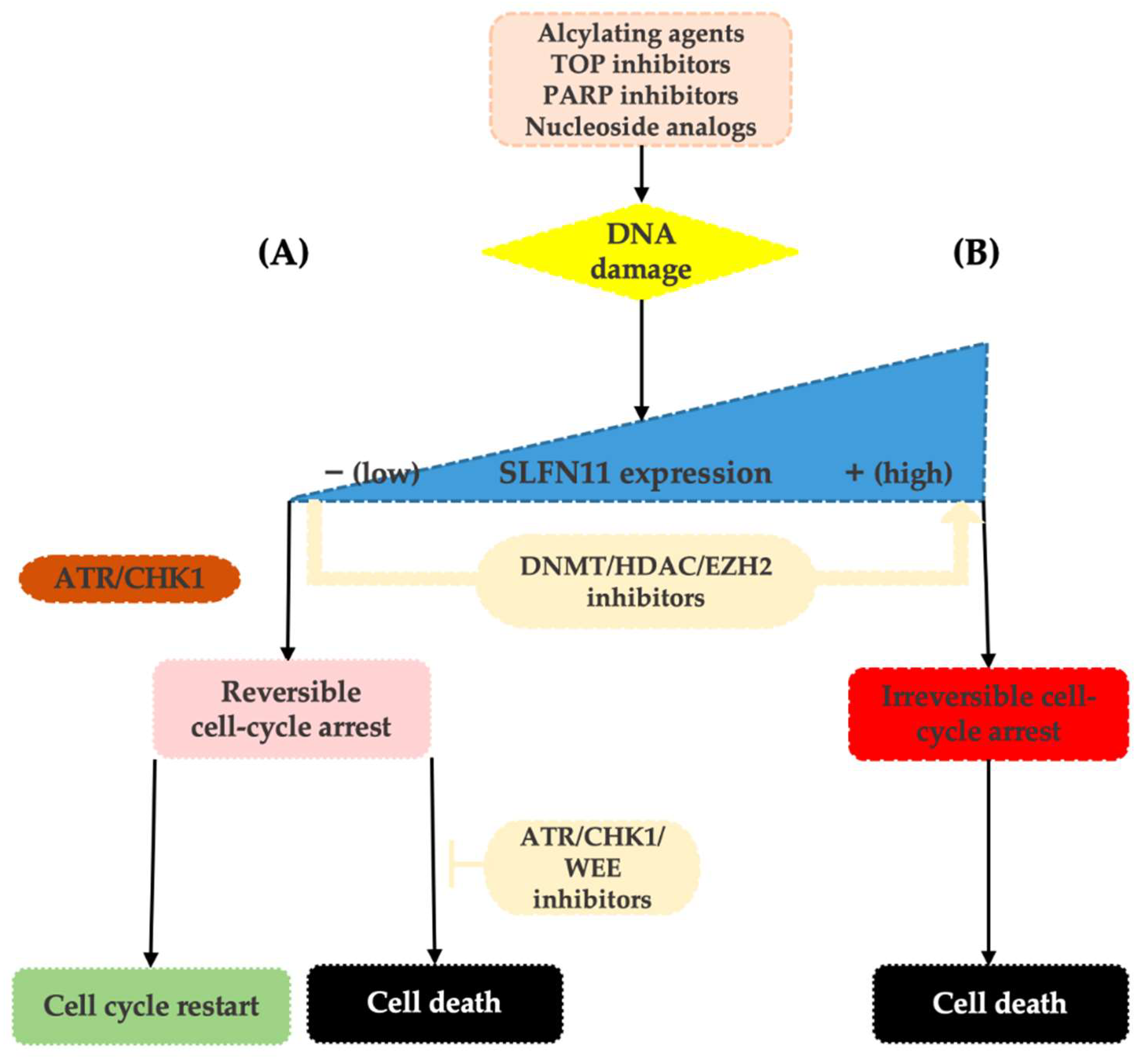
3. SLFN11 as a Guardian of the Genome in Response to Replication Stress
4. Clinical Evaluation of SLFN11 Status
5. Prevalence of SLFN11 in Ovarian Cancer
6. SLFN11 as a Predictive Biomarker for DDA or PARPi Response in Ovarian Cancer Patients
7. SLFN11 as a Prognostic Biomarker in Ovarian Cancer
8. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Cannistra, S.A. Cancer of the Ovary. N. Engl. J. Med. 2004, 351, 2519–2529. [Google Scholar] [CrossRef] [PubMed]
- Colombo, N.; Sessa, C.; du Bois, A.; Ledermann, J.; McCluggage, W.G.; McNeish, I.; Morice, P.; Pignata, S.; Ray-Coquard, I.; Vergote, I.; et al. ESMO–ESGO Consensus Conference Recommendations on Ovarian Cancer: Pathology and Molecular Biology, Early and Advanced Stages, Borderline Tumours and Recurrent Disease†. Ann. Oncol. 2019, 30, 672–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.K.; Pujade-Lauraine, E.; Aoki, D.; Mirza, M.R.; Lorusso, D.; Oza, A.M.; du Bois, A.; Vergote, I.; Reuss, A.; Bacon, M.; et al. Fifth Ovarian Cancer Consensus Conference of the Gynecologic Cancer InterGroup: Recurrent Disease. Ann. Oncol. 2017, 28, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Pujade-Lauraine, E.; Hilpert, F.; Weber, B.; Reuss, A.; Poveda, A.; Kristensen, G.; Sorio, R.; Vergote, I.; Witteveen, P.; Bamias, A.; et al. Bevacizumab Combined With Chemotherapy for Platinum-Resistant Recurrent Ovarian Cancer: The AURELIA Open-Label Randomized Phase III Trial. J. Clin. Oncol. 2014, 32, 1302–1308. [Google Scholar] [CrossRef]
- Burger, R.A.; Brady, M.F.; Bookman, M.A.; Fleming, G.F.; Monk, B.J.; Huang, H.; Mannel, R.S.; Homesley, H.D.; Fowler, J.; Greer, B.E.; et al. Incorporation of Bevacizumab in the Primary Treatment of Ovarian Cancer. N. Engl. J. Med. 2011, 365, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Oza, A.M.; Cook, A.D.; Pfisterer, J.; Embleton, A.; Ledermann, J.A.; Pujade-Lauraine, E.; Kristensen, G.; Carey, M.S.; Beale, P.; Cervantes, A.; et al. Standard Chemotherapy with or without Bevacizumab for Women with Newly Diagnosed Ovarian Cancer (ICON7): Overall Survival Results of a Phase 3 Randomised Trial. Lancet Oncol. 2015, 16, 928–936. [Google Scholar] [CrossRef]
- Aghajanian, C.; Blank, S.V.; Goff, B.A.; Judson, P.L.; Teneriello, M.G.; Husain, A.; Sovak, M.A.; Yi, J.; Nycum, L.R. OCEANS: A Randomized, Double-Blind, Placebo-Controlled Phase III Trial of Chemotherapy With or Without Bevacizumab in Patients With Platinum-Sensitive Recurrent Epithelial Ovarian, Primary Peritoneal, or Fallopian Tube Cancer. J. Clin. Oncol. 2012, 30, 2039–2045. [Google Scholar] [CrossRef] [Green Version]
- Colombo, I.; Kurnit, K.C.; Westin, S.N.; Oza, A.M. Moving from Mutation to Actionability. Am. Soc. Clin. Oncol. Educ. Book 2018, 38, 495–503. [Google Scholar] [CrossRef]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehár, J.; Kryukov, G.V.; Sonkin, D.; et al. The Cancer Cell Line Encyclopedia Enables Predictive Modelling of Anticancer Drug Sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef]
- Stewart, C.A.; Tong, P.; Cardnell, R.J.; Sen, T.; Li, L.; Gay, C.M.; Masrorpour, F.; Fan, Y.; Bara, R.O.; Feng, Y.; et al. Dynamic Variations in Epithelial-to-Mesenchymal Transition (EMT), ATM, and SLFN11 Govern Response to PARP Inhibitors and Cisplatin in Small Cell Lung Cancer. Oncotarget 2017, 8, 28575–28587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballestrero, A.; Bedognetti, D.; Ferraioli, D.; Franceschelli, P.; Labidi-Galy, S.I.; Leo, E.; Murai, J.; Pommier, Y.; Tsantoulis, P.; Vellone, V.G.; et al. Report on the First SLFN11 Monothematic Workshop: From Function to Role as a Biomarker in Cancer. J. Transl. Med. 2017, 15, 199. [Google Scholar] [CrossRef] [PubMed]
- Colombo, P.-E.; Fabbro, M.; Theillet, C.; Bibeau, F.; Rouanet, P.; Ray-Coquard, I. Sensitivity and Resistance to Treatment in the Primary Management of Epithelial Ovarian Cancer. Crit. Rev. Oncol. Hematol. 2014, 89, 207–216. [Google Scholar] [CrossRef] [PubMed]
- McMullen, M.; Madariaga, A.; Lheureux, S. New Approaches for Targeting Platinum-Resistant Ovarian Cancer. Semin. Cancer Biol. 2021, 77, 167–181. [Google Scholar] [CrossRef]
- The Australian Ovarian Cancer Study Group; Patch, A.-M.; Christie, E.L.; Etemadmoghadam, D.; Garsed, D.W.; George, J.; Fereday, S.; Nones, K.; Cowin, P.; Alsop, K.; et al. Whole–Genome Characterization of Chemoresistant Ovarian Cancer. Nature 2015, 521, 489–494. [Google Scholar] [CrossRef]
- Lloyd, K.L.; Cree, I.A.; Savage, R.S. Prediction of Resistance to Chemotherapy in Ovarian Cancer: A Systematic Review. BMC Cancer 2015, 15, 117. [Google Scholar] [CrossRef] [Green Version]
- Weberpals, J.I.; Pugh, T.J.; Marco-Casanova, P.; Goss, G.D.; Andrews Wright, N.; Rath, P.; Torchia, J.; Fortuna, A.; Jones, G.N.; Roudier, M.P.; et al. Tumor Genomic, Transcriptomic, and Immune Profiling Characterizes Differential Response to First-line Platinum Chemotherapy in High Grade Serous Ovarian Cancer. Cancer Med. 2021, 10, 3045–3058. [Google Scholar] [CrossRef]
- Tian, L.; Song, S.; Liu, X.; Wang, Y.; Xu, X.; Hu, Y.; Xu, J. Schlafen-11 Sensitizes Colorectal Carcinoma Cells to Irinotecan. Anticancer Drugs 2014, 25, 1175–1181. [Google Scholar] [CrossRef]
- Kang, M.H.; Wang, J.; Makena, M.R.; Lee, J.-S.; Paz, N.; Hall, C.P.; Song, M.M.; Calderon, R.I.; Cruz, R.E.; Hindle, A.; et al. Activity of MM-398, Nanoliposomal Irinotecan (Nal-IRI), in Ewing’s Family Tumor Xenografts Is Associated with High Exposure of Tumor to Drug and High SLFN11 Expression. Clin. Cancer Res. 2015, 21, 1139–1150. [Google Scholar] [CrossRef] [Green Version]
- Shee, K.; Wells, J.D.; Jiang, A.; Miller, T.W. Integrated Pan-Cancer Gene Expression and Drug Sensitivity Analysis Reveals SLFN11 MRNA as a Solid Tumor Biomarker Predictive of Sensitivity to DNA-Damaging Chemotherapy. PLoS ONE 2019, 14, e0224267. [Google Scholar] [CrossRef]
- Coussy, F.; El-Botty, R.; Château-Joubert, S.; Dahmani, A.; Montaudon, E.; Leboucher, S.; Morisset, L.; Painsec, P.; Sourd, L.; Huguet, L.; et al. BRCAness, SLFN11, and RB1 Loss Predict Response to Topoisomerase I Inhibitors in Triple-Negative Breast Cancers. Sci. Transl. Med. 2020, 12, eaax2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathkey, D.; Khanal, M.; Murai, J.; Zhang, J.; Sengupta, M.; Jiang, Q.; Morrow, B.; Evans, C.N.; Chari, R.; Fetsch, P.; et al. Sensitivity of Mesothelioma Cells to PARP Inhibitors Is Not Dependent on BAP1 but Is Enhanced by Temozolomide in Cells with High-Schlafen 11 and Low-O6-Methylguanine-DNA Methyltransferase Expression. J. Thorac. Oncol. 2020, 15, 843–859. [Google Scholar] [CrossRef] [PubMed]
- Takashima, T.; Taniyama, D.; Sakamoto, N.; Yasumoto, M.; Asai, R.; Hattori, T.; Honma, R.; Thang, P.Q.; Ukai, S.; Maruyama, R.; et al. Schlafen 11 Predicts Response to Platinum-Based Chemotherapy in Gastric Cancers. Br. J. Cancer 2021, 125, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Zoppoli, G.; Regairaz, M.; Leo, E.; Reinhold, W.C.; Varma, S.; Ballestrero, A.; Doroshow, J.H.; Pommier, Y. Putative DNA/RNA Helicase Schlafen-11 (SLFN11) Sensitizes Cancer Cells to DNA-Damaging Agents. Proc. Natl. Acad. Sci. USA 2012, 109, 15030–15035. [Google Scholar] [CrossRef] [Green Version]
- Nogales, V.; Reinhold, W.C.; Varma, S.; Martinez-Cardus, A.; Moutinho, C.; Moran, S.; Heyn, H.; Sebio, A.; Barnadas, A.; Pommier, Y.; et al. Epigenetic Inactivation of the Putative DNA/RNA Helicase SLFN11 in Human Cancer Confers Resistance to Platinum Drugs. Oncotarget 2016, 7, 3084–3097. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Cai, Y.; Huang, Y.; Yang, Z.; Bai, Y.; Liu, Y.; Deng, X.; Wang, J. High SLFN11 Expression Predicts Better Survival for Patients with KRAS Exon 2 Wild Type Colorectal Cancer after Treated with Adjuvant Oxaliplatin-Based Treatment. BMC Cancer 2015, 15, 833. [Google Scholar] [CrossRef] [Green Version]
- Lok, B.H.; Gardner, E.E.; Schneeberger, V.E.; Ni, A.; Desmeules, P.; Rekhtman, N.; de Stanchina, E.; Teicher, B.A.; Riaz, N.; Powell, S.N.; et al. PARP Inhibitor Activity Correlates with SLFN11 Expression and Demonstrates Synergy with Temozolomide in Small Cell Lung Cancer. Clin. Cancer Res. 2017, 23, 523–535. [Google Scholar] [CrossRef] [Green Version]
- Pietanza, M.C.; Waqar, S.N.; Krug, L.M.; Dowlati, A.; Hann, C.L.; Chiappori, A.; Owonikoko, T.K.; Woo, K.M.; Cardnell, R.J.; Fujimoto, J.; et al. Randomized, Double-Blind, Phase II Study of Temozolomide in Combination with Either Veliparib or Placebo in Patients with Relapsed-Sensitive or Refractory Small-Cell Lung Cancer. J. Clin. Oncol. 2018, 36, 2386–2394. [Google Scholar] [CrossRef]
- Takashima, T.; Sakamoto, N.; Murai, J.; Taniyama, D.; Honma, R.; Ukai, S.; Maruyama, R.; Kuraoka, K.; Rajapakse, V.N.; Pommier, Y.; et al. Immunohistochemical Analysis of SLFN11 Expression Uncovers Potential Non-Responders to DNA-Damaging Agents Overlooked by Tissue RNA-Seq. Virchows Arch. 2021, 478, 569–579. [Google Scholar] [CrossRef]
- Zhang, B.; Ramkumar, K.; Cardnell, R.J.; Gay, C.M.; Stewart, C.A.; Wang, W.-L.; Fujimoto, J.; Wistuba, I.I.; Byers, L.A. A Wake-up Call for Cancer DNA Damage: The Role of Schlafen 11 (SLFN11) across Multiple Cancers. Br. J. Cancer 2021, 125, 1333–1340. [Google Scholar] [CrossRef]
- Aladjem, M.I.; Redon, C.E. Order from Clutter: Selective Interactions at Mammalian Replication Origins. Nat. Rev. Genet. 2017, 18, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Murai, J.; Tang, S.-W.; Leo, E.; Baechler, S.A.; Redon, C.E.; Zhang, H.; Al Abo, M.; Rajapakse, V.N.; Nakamura, E.; Jenkins, L.M.M.; et al. SLFN11 Blocks Stressed Replication Forks Independently of ATR. Mol. Cell 2018, 69, 371–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, Y.; Lou, J.; Srivastava, M.; Zhao, B.; Feng, X.-H.; Liu, T.; Chen, J.; Huang, J. SLFN11 Inhibits Checkpoint Maintenance and Homologous Recombination Repair. EMBO Rep. 2016, 17, 94–109. [Google Scholar] [CrossRef] [Green Version]
- Jo, U.; Murai, Y.; Chakka, S.; Chen, L.; Cheng, K.; Murai, J.; Saha, L.K.; Miller Jenkins, L.M.; Pommier, Y. SLFN11 Promotes CDT1 Degradation by CUL4 in Response to Replicative DNA Damage, While Its Absence Leads to Synthetic Lethality with ATR/CHK1 Inhibitors. Proc. Natl. Acad. Sci. USA 2021, 118, e2015654118. [Google Scholar] [CrossRef] [PubMed]
- Jo, U.; Murai, Y.; Takebe, N.; Thomas, A.; Pommier, Y. Precision Oncology with Drugs Targeting the Replication Stress, ATR, and Schlafen 11. Cancers 2021, 13, 4601. [Google Scholar] [CrossRef] [PubMed]
- Murai, J.; Thomas, A.; Miettinen, M.; Pommier, Y. Schlafen 11 (SLFN11), a Restriction Factor for Replicative Stress Induced by DNA-Targeting Anti-Cancer Therapies. Pharmacol. Ther. 2019, 201, 94–102. [Google Scholar] [CrossRef]
- Forment, J.V.; O’Connor, M.J. Targeting the Replication Stress Response in Cancer. Pharmacol. Ther. 2018, 188, 155–167. [Google Scholar] [CrossRef]
- Rajapakse, V.N.; Luna, A.; Yamade, M.; Loman, L.; Varma, S.; Sunshine, M.; Iorio, F.; Sousa, F.G.; Elloumi, F.; Aladjem, M.I.; et al. CellMinerCDB for Integrative Cross-Database Genomics and Pharmacogenomics Analyses of Cancer Cell Lines. iScience 2018, 10, 247–264. [Google Scholar] [CrossRef] [Green Version]
- Moribe, F.; Nishikori, M.; Takashima, T.; Taniyama, D.; Onishi, N.; Arima, H.; Sasanuma, H.; Akagawa, R.; Elloumi, F.; Takeda, S.; et al. Epigenetic Suppression of SLFN11 in Germinal Center B-Cells during B-Cell Development. PLoS ONE 2021, 16, e0237554. [Google Scholar] [CrossRef]
- Reinhold, W.C.; Thomas, A.; Pommier, Y. DNA-Targeted Precision Medicine; Have We Been Caught Sleeping? Trends Cancer 2017, 3, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Reinhold, W.C.; Varma, S.; Sunshine, M.; Rajapakse, V.; Luna, A.; Kohn, K.W.; Stevenson, H.; Wang, Y.; Heyn, H.; Nogales, V.; et al. The NCI-60 Methylome and Its Integration into CellMiner. Cancer Res. 2017, 77, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.-W.; Thomas, A.; Murai, J.; Trepel, J.B.; Bates, S.E.; Rajapakse, V.N.; Pommier, Y. Overcoming Resistance to DNA-Targeted Agents by Epigenetic Activation of Schlafen 11 (SLFN11) Expression with Class I Histone Deacetylase Inhibitors. Clin. Cancer Res. 2018, 24, 1944–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, E.E.; Lok, B.H.; Schneeberger, V.E.; Desmeules, P.; Miles, L.A.; Arnold, P.K.; Ni, A.; Khodos, I.; de Stanchina, E.; Nguyen, T.; et al. Chemosensitive Relapse in Small Cell Lung Cancer Proceeds through an EZH2-SLFN11 Axis. Cancer Cell 2017, 31, 286–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lheureux, S.; Oaknin, A.; Garg, S.; Bruce, J.P.; Madariaga, A.; Dhani, N.C.; Bowering, V.; White, J.; Accardi, S.; Tan, Q.; et al. EVOLVE: A Multicenter Open-Label Single-Arm Clinical and Translational Phase II Trial of Cediranib Plus Olaparib for Ovarian Cancer after PARP Inhibition Progression. Clin. Cancer Res. 2020, 26, 4206–4215. [Google Scholar] [CrossRef]
- Murai, J.; Feng, Y.; Yu, G.K.; Ru, Y.; Tang, S.-W.; Shen, Y.; Pommier, Y. Resistance to PARP Inhibitors by SLFN11 Inactivation Can Be Overcome by ATR Inhibition. Oncotarget 2016, 7, 76534–76550. [Google Scholar] [CrossRef] [Green Version]
- Winkler, C.; Armenia, J.; Jones, G.N.; Tobalina, L.; Sale, M.J.; Petreus, T.; Baird, T.; Serra, V.; Wang, A.T.; Lau, A.; et al. SLFN11 Informs on Standard of Care and Novel Treatments in a Wide Range of Cancer Models. Br. J. Cancer 2021, 124, 951–962. [Google Scholar] [CrossRef]
- Zhou, C.; Liu, C.; Liu, W.; Chen, W.; Yin, Y.; Li, C.-W.; Hsu, J.L.; Sun, J.; Zhou, Q.; Li, H.; et al. SLFN11 Inhibits Hepatocellular Carcinoma Tumorigenesis and Metastasis by Targeting RPS4X via MTOR Pathway. Theranostics 2020, 10, 4627–4643. [Google Scholar] [CrossRef]
- Kagami, T.; Yamade, M.; Suzuki, T.; Uotani, T.; Tani, S.; Hamaya, Y.; Iwaizumi, M.; Osawa, S.; Sugimoto, K.; Miyajima, H.; et al. The First Evidence for SLFN11 Expression as an Independent Prognostic Factor for Patients with Esophageal Cancer after Chemoradiotherapy. BMC Cancer 2020, 20, 1123. [Google Scholar] [CrossRef]
- Winkler, C.; King, M.; Berthe, J.; Ferraioli, D.; Garuti, A.; Grillo, F.; Rodriguez-Canales, J.; Ferrando, L.; Chopin, N.; Ray-Coquard, I.; et al. SLFN11 Captures Cancer-Immunity Interactions Associated with Platinum Sensitivity in High-Grade Serous Ovarian Cancer. JCI Insight 2021, 6, e146098. [Google Scholar] [CrossRef]
- Gartrell, J.; Mellado-Largarde, M.; Clay, M.R.; Bahrami, A.; Sahr, N.A.; Sykes, A.; Blankenship, K.; Hoffmann, L.; Xie, J.; Cho, H.P.; et al. SLFN11 Is Widely Expressed in Pediatric Sarcoma and Induces Variable Sensitization to Replicative Stress Caused By DNA-Damaging Agents. Mol. Cancer Ther. 2021, 20, 2151–2165. [Google Scholar] [CrossRef]
- Taniyama, D.; Sakamoto, N.; Takashima, T.; Takeda, M.; Pham, Q.T.; Ukai, S.; Maruyama, R.; Harada, K.; Babasaki, T.; Sekino, Y.; et al. Prognostic Impact of Schlafen 11 in Bladder Cancer Patients Treated with Platinum-based Chemotherapy. Cancer Sci. 2022, 113, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Willis, S.E.; Winkler, C.; Roudier, M.P.; Baird, T.; Marco-Casanova, P.; Jones, E.V.; Rowe, P.; Rodriguez-Canales, J.; Angell, H.K.; Ng, F.S.L.; et al. Retrospective Analysis of Schlafen11 (SLFN11) to Predict the Outcomes to Therapies Affecting the DNA Damage Response. Br. J. Cancer 2021, 125, 1666–1676. [Google Scholar] [CrossRef] [PubMed]
- Ledermann, J.; Harter, P.; Gourley, C.; Friedlander, M.; Vergote, I.; Rustin, G.; Scott, C.; Meier, W.; Shapira-Frommer, R.; Safra, T.; et al. Olaparib Maintenance Therapy in Platinum-Sensitive Relapsed Ovarian Cancer. N. Engl. J. Med. 2012, 366, 1382–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byers, L.A.; Stewart, A.; Gay, C.; Heymach, J.; Fernandez, L.; Lu, D.; Rich, R.; Chu, L.; Wang, Y.; Dittamore, R. Abstract 2215: SLFN11 and EZH2 Protein Expression and Localization in Circulating Tumor Cells to Predict Response or Resistance to DNA Damaging Therapies in Small Cell Lung Cancer. Cancer Res. 2019, 79, 2215. [Google Scholar]
- Zhang, B.; Stewart, C.A.; Gay, C.M.; Wang, Q.; Cardnell, R.; Fujimoto, J.; Fernandez, L.; Jendrisak, A.; Gilbertson, C.; Schonhoft, J.; et al. Abstract 384: Detection of DNA Replication Blocker SLFN11 in Tumor Tissue and Circulating Tumor Cells to Predict Platinum Response in Small Cell Lung Cancer. Cancer Res. 2021, 81, 384. [Google Scholar] [CrossRef]
- Van Berckelaer, C.; Brouwers, A.J.; Peeters, D.J.E.; Tjalma, W.; Trinh, X.B.; van Dam, P.A. Current and Future Role of Circulating Tumor Cells in Patients with Epithelial Ovarian Cancer. Eur. J. Surg. Oncol. EJSO 2016, 42, 1772–1779. [Google Scholar] [CrossRef]
- Holcakova, J.; Bartosik, M.; Anton, M.; Minar, L.; Hausnerova, J.; Bednarikova, M.; Weinberger, V.; Hrstka, R. New Trends in the Detection of Gynecological Precancerous Lesions and Early-Stage Cancers. Cancers 2021, 13, 6339. [Google Scholar] [CrossRef]
- Federico, S.M.; Pappo, A.S.; Sahr, N.; Sykes, A.; Campagne, O.; Stewart, C.F.; Clay, M.R.; Bahrami, A.; McCarville, M.B.; Kaste, S.C.; et al. A Phase I Trial of Talazoparib and Irinotecan with and without Temozolomide in Children and Young Adults with Recurrent or Refractory Solid Malignancies. Eur. J. Cancer 2020, 137, 204–213. [Google Scholar] [CrossRef]
- Byers, L.A.; Bentsion, D.; Gans, S.; Penkov, K.; Son, C.; Sibille, A.; Owonikoko, T.K.; Groen, H.J.M.; Gay, C.M.; Fujimoto, J.; et al. Veliparib in Combination with Carboplatin and Etoposide in Patients with Treatment-Naïve Extensive-Stage Small Cell Lung Cancer: A Phase 2 Randomized Study. Clin. Cancer Res. 2021, 27, 3884–3895. [Google Scholar] [CrossRef]
- He, T.; Zhang, M.; Zheng, R.; Zheng, S.; Linghu, E.; Herman, J.G.; Guo, M. Methylation of SLFN11 Is a Marker of Poor Prognosis and Cisplatin Resistance in Colorectal Cancer. Epigenomics 2017, 9, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Hwang, W.-T.; Adams, S.F.; Tahirovic, E.; Hagemann, I.S.; Coukos, G. Prognostic Significance of Tumor-Infiltrating T Cells in Ovarian Cancer: A Meta-Analysis. Gynecol. Oncol. 2012, 124, 192–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, J.; Chen, R.; Bai, Y.; Lu, X. The Prognostic Value of Tumor-Infiltrating T Lymphocytes in Ovarian Cancer. Oncotarget 2017, 8, 15621–15631. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| DNA-Targeting Agents | Representative Drugs | Target | Mechanism of Action |
|---|---|---|---|
| Alkylating agents | Cisplatine Carboplatine Oxaliplatine | DNA template damage | Inter-strand crosslinks |
| Temozolomide | DNA template damage MGMT | O6-alkyl-guanine lesions on DNA | |
| TOP I and II inhibitors | Irinotecan Topotecan Etoposide Doxorubicin Mitoxantrone | DNA template damage | Block the re-ligation of the TOP-DNA cleavage complexes |
| PARP inhibitors | Olaparib Rucaparib Niraparib Talazoparib Veliparib | DNA template damage by defective single-strand breaks repair | Generating toxic PARP-DNA complexes |
| Nucleoside analogs | Gemcitabine Cytarabine 5-azacytidine | DNA elongation inhibition | Blocking DNA polymerase or reducing the pool of nucleotides |
| Tumor Origin (n) | Tissue | Antibody | Evaluation | Author [Reference] |
|---|---|---|---|---|
| CRC (261) | FFPE | Abcam; ab121731 | H-score; value 0–6 1 >4.5—high | Deng [26] |
| HGSC (75) | N/A | N/A | H-score; value 0–9 2 0—negative 1, 2—low 3, 4—intermediate 6, 9—high | Ballestrero [12] |
| SCLC (12) | PDX | Sigma-Aldrich; HPA023030 | H-score; value 0–300 3 | Stewart [11] |
| SCLC (7) | PDX | Sigma-Aldrich; HPA023030 | H-score; value 0–300 3 low v. high | Lok [27] |
| SCLC (48) | FFPE | Sigma-Aldrich; HPA023030 | H-score; value 0–300 3 >1—positive | Pietanza [28] |
| HCC (182/110) | FFPE | Sigma-Aldrich; HPA023030 | H-score 0/+—low ++/+++—high | Zhou [47] |
| TNBC (40) | PDX | Sigma-Aldrich; HPA023030 | H-score; value 0–300 3 0—negative 1–60—low >60—high | Coussy [21] |
| ESCC (73) | FFPE | Santa Cruz; #sc-515071 | H-score; value 0–300 3 ≥51—high | Kagami [48] |
| 16 human adult organs; malignant and adjacent non-tumor tissue (~ 700) | FFPE |
| IHC scoring system 1+ (1–10%) 2+ (11–50%) 3+ (51–100%) | Takashima [29] |
| Various cancer types | PDX | Abcam; ab121731 | H-score; value 0–300 3 ≥31—high | Winkler [46] |
| HGSC (28) | FFPE | Abcam; ab121731 | H-score; value 0–300 3 ≥60—high | Winkler [49] |
| Pediatric sarcoma (220) | FFPE | Sigma-Aldrich; HPA023030 | H-score; value 0–300 3 0—negative ≥1—positive | Gartrell [50] |
| Gastric (169) | FFPE | Santa Cruz; #sc-515071 | IHC scoring system >30%—positive | Takashima [23] |
| Bladder (120) | FFPE | Santa Cruz; #sc-515071 | IHC scoring system >5%—positive | Taniyama [51] |
| Non-tumor tissue (86) SCLC (124) HGSC (151) | TMA FFPE | Merck; MABF248 Abcam; ab121731 Novus; NBP2–57084 | H-score; value 0–300 3 >122—high >30—high | Willis [52] |
| Type | Regime | n | Method | Results | Author [Reference] |
|---|---|---|---|---|---|
| cystadeno-carcinoma | P-based | 110 | IB | high SLFN11 expression independently predicts better OS | Zoppoli [24] |
| serous OC | P-based | 41 | M | SLFN11 hypermethylation was significantly associated with shorter OS and PFS | Nogales [25] |
| OC | P-based | 110 (OS) 75 (DFS) | RNA-seq |
| Shee [20] |
| HGSC | P-based | 28 (PFI) 221 (OS) | IHC |
| Winkler [49] |
| HGSC |
| 34 110 | IHC |
| Willis [52] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bednarikova, M.; Hausnerova, J.; Ehrlichova, L.; Matulova, K.; Gazarkova, E.; Minar, L.; Weinberger, V. Can Schlafen 11 Help to Stratify Ovarian Cancer Patients Treated with DNA-Damaging Agents? Cancers 2022, 14, 2353. https://doi.org/10.3390/cancers14102353
Bednarikova M, Hausnerova J, Ehrlichova L, Matulova K, Gazarkova E, Minar L, Weinberger V. Can Schlafen 11 Help to Stratify Ovarian Cancer Patients Treated with DNA-Damaging Agents? Cancers. 2022; 14(10):2353. https://doi.org/10.3390/cancers14102353
Chicago/Turabian StyleBednarikova, Marketa, Jitka Hausnerova, Lucie Ehrlichova, Kvetoslava Matulova, Eliska Gazarkova, Lubos Minar, and Vit Weinberger. 2022. "Can Schlafen 11 Help to Stratify Ovarian Cancer Patients Treated with DNA-Damaging Agents?" Cancers 14, no. 10: 2353. https://doi.org/10.3390/cancers14102353
APA StyleBednarikova, M., Hausnerova, J., Ehrlichova, L., Matulova, K., Gazarkova, E., Minar, L., & Weinberger, V. (2022). Can Schlafen 11 Help to Stratify Ovarian Cancer Patients Treated with DNA-Damaging Agents? Cancers, 14(10), 2353. https://doi.org/10.3390/cancers14102353