
Aging Aggravates Cachexia in Tumor-Bearing Mice
 , , , ,
, , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mouse Models
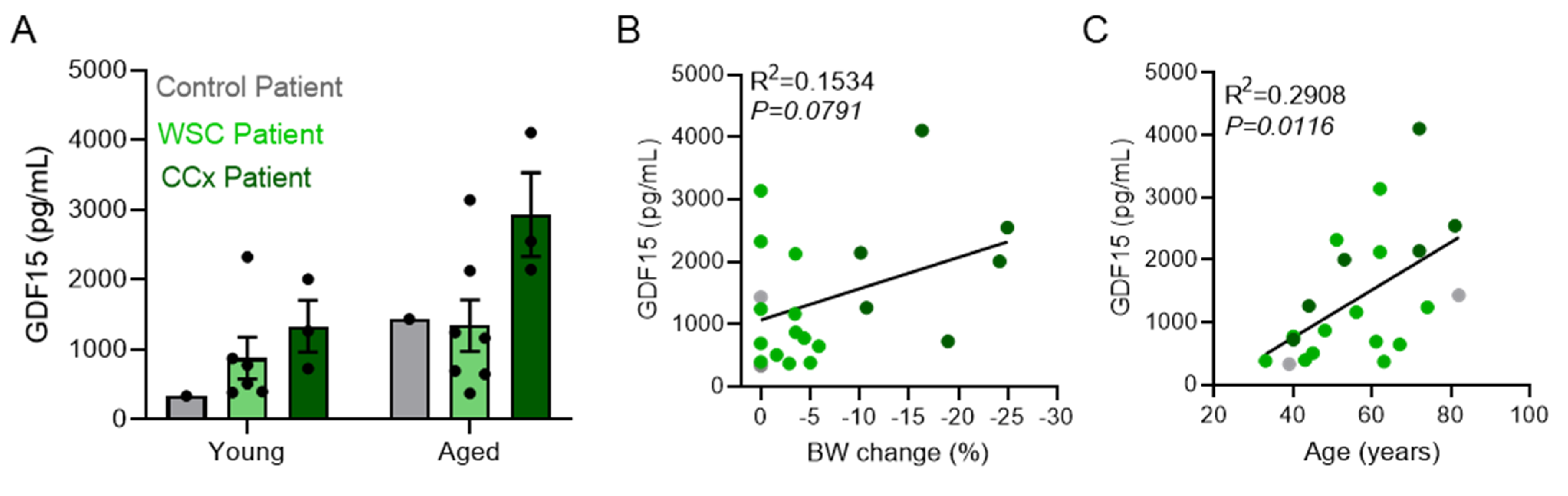
2.2. Patient Cohort and Samples
2.3. Plasma Analysis and Enzyme-Linked Immunosorbent Assays
2.4. RNA Extraction and Reverse Transcription
2.5. Quantitative PCR
2.6. Immunohistochemistry
2.7. Protein Extraction and Immunoblotting
2.8. Cell Staining for Flow Cytometry
2.9. Statistical Analysis
3. Results
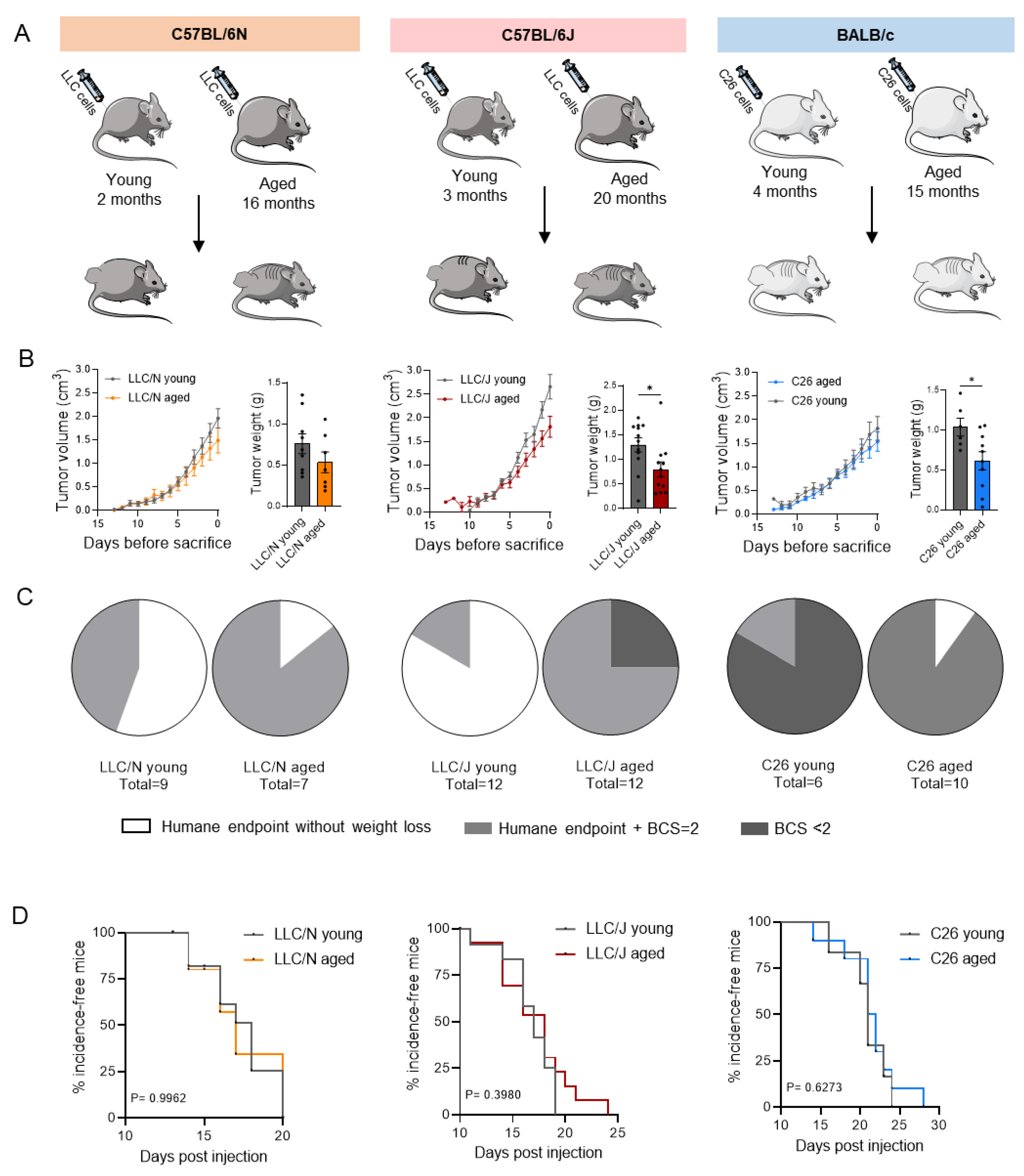
3.1. Tumor Growth Is Largely Unaffected by Age
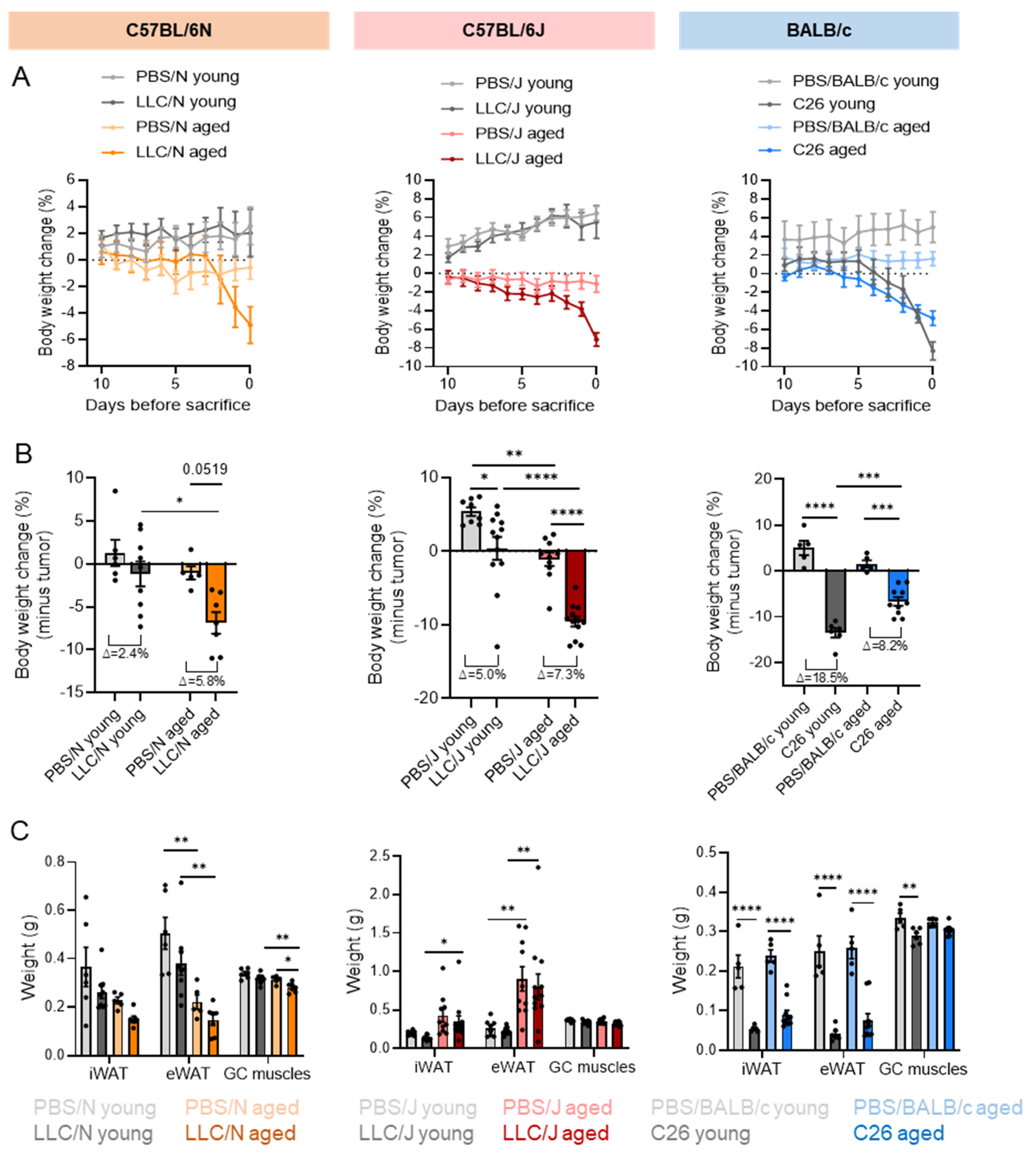
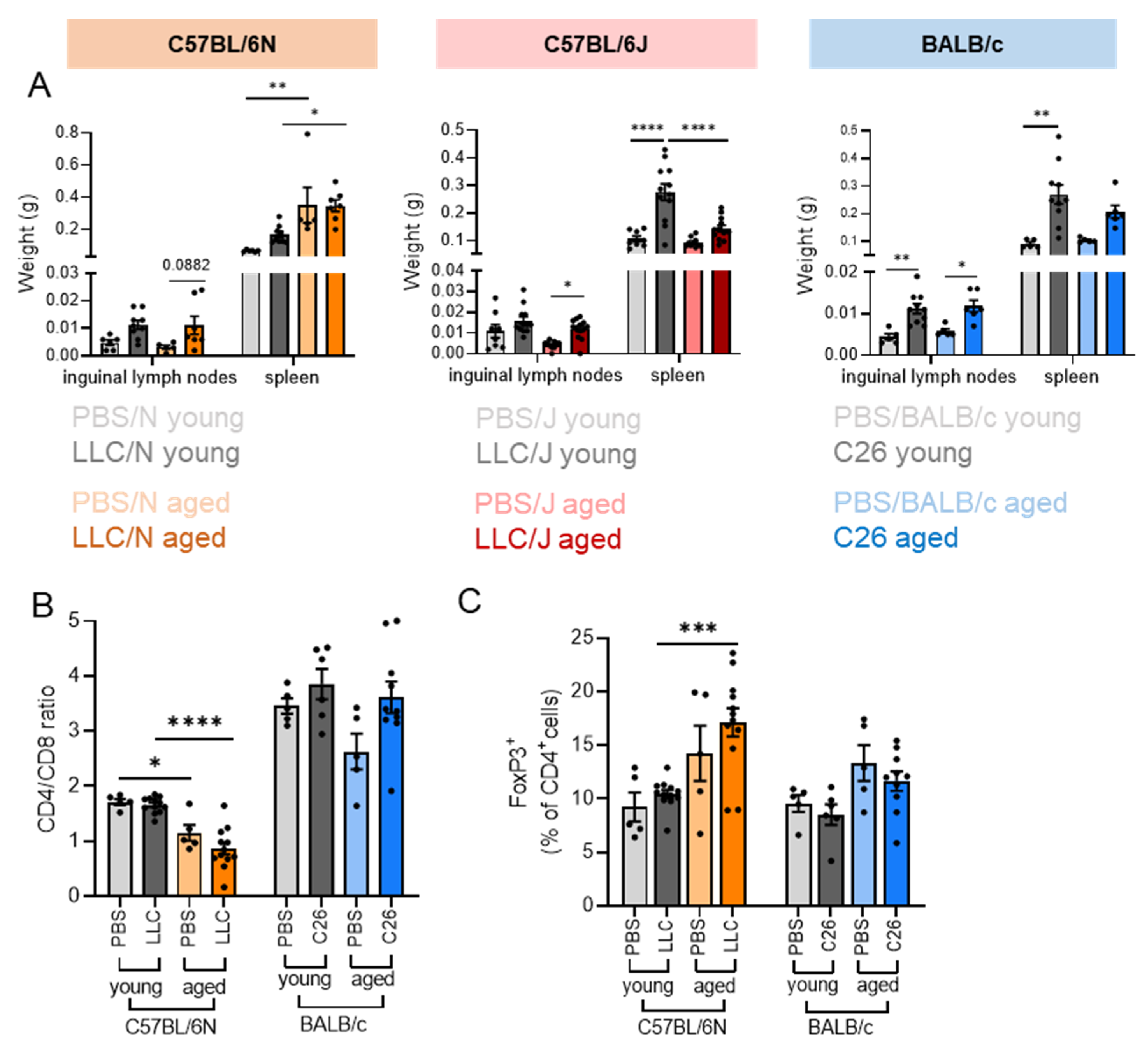
3.2. Strain-Dependent Differences in Cancer-Induced Loss of Body and Tissue Weights
3.3. Aging Increases the Tumor-Induced Expression of Atrogenes in Cachectic Muscle
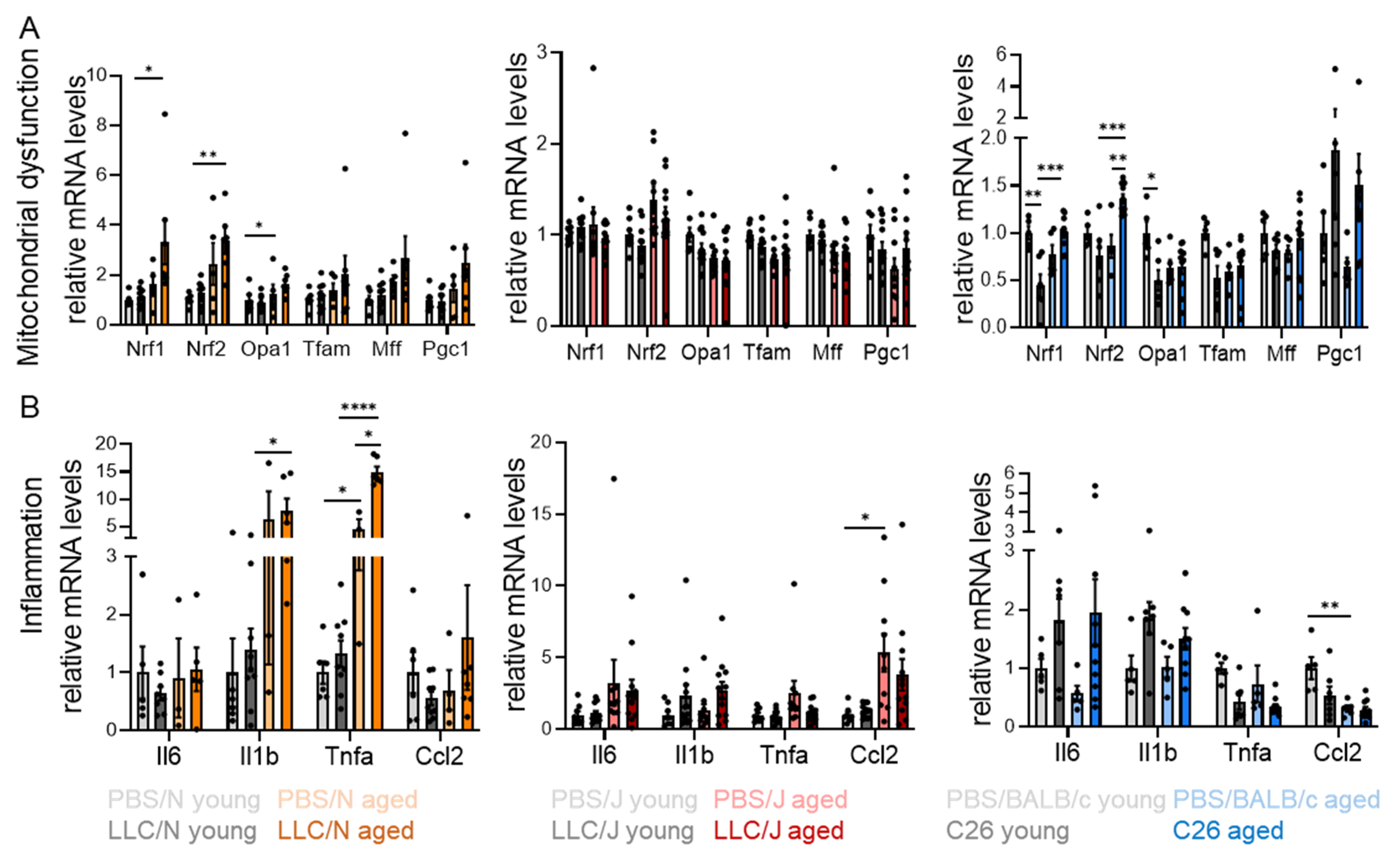
3.4. Age-Dependent Mitochondrial Stress and Inflammation in Adipose Tissue
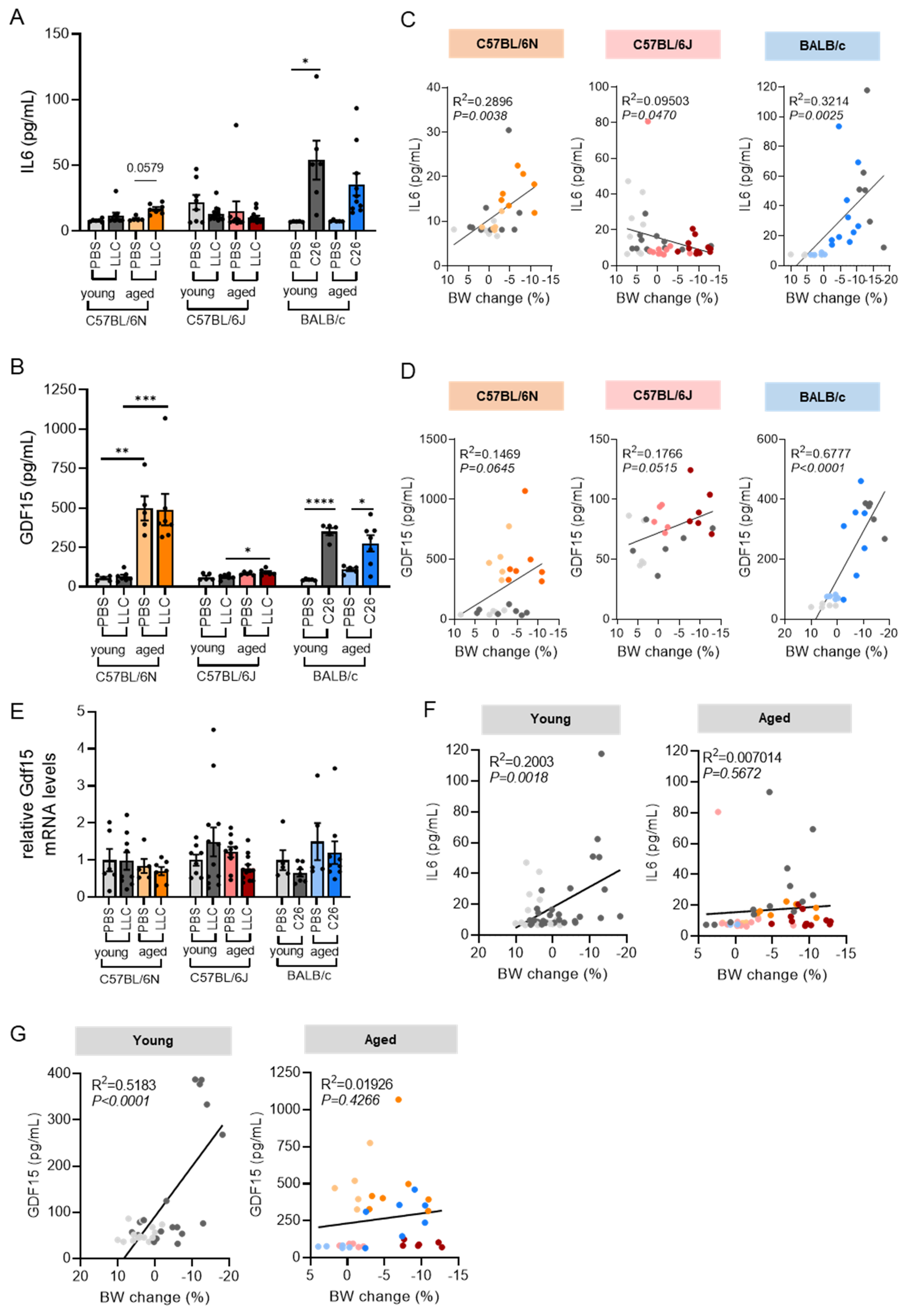
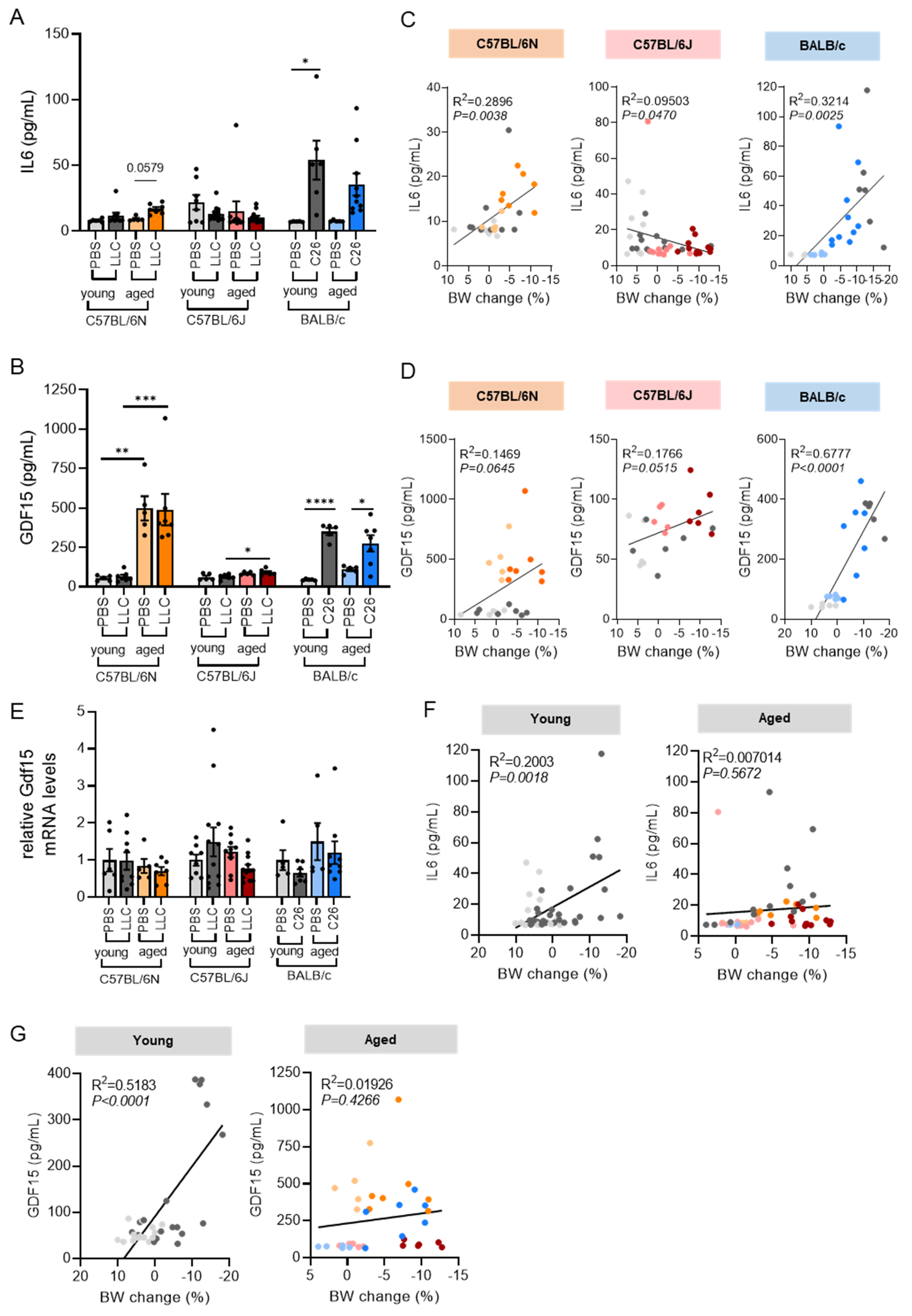
3.5. Circulating IL6 and GDF15 Levels Are Cachexia Markers in C26 Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Berben, L.; Floris, G.; Wildiers, H.; Hatse, S. Cancer and Aging: Two Tightly Interconnected Biological Processes. Cancers 2021, 13, 1400. [Google Scholar] [CrossRef] [PubMed]
- Dunne, R.F.; Loh, K.P.; Williams, G.R.; Jatoi, A.; Mustian, K.M.; Mohile, S.G. Cachexia and Sarcopenia in Older Adults with Cancer: A Comprehensive Review. Cancers 2019, 11, 1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuraguchi, M.; Wang, X.-P.; Bronson, R.T.; Rothenberg, R.; Ohene-Baah, N.Y.; Lund, J.J.; Kucherlapati, M.; Maas, R.L.; Kucherlapati, R. Adenomatous Polyposis Coli (APC) Is Required for Normal Development of Skin and Thymus. PLoS Genet. 2006, 2, e146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hingorani, S.R.; Wang, L.; Multani, A.S.; Combs, C.; Deramaudt, T.B.; Hruban, R.H.; Rustgi, A.K.; Chang, S.; Tuveson, D.A. Trp53R172H and KrasG12D cooperate to promote chromosomal instability and widely metastatic pancreatic ductal adenocarcinoma in mice. Cancer Cell 2005, 7, 469–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Eda, H.; Tanaka, T.; Udagawa, T.; Ishikawa, T.; Horii, I.; Ishitsuka, H.; Kataoka, T.; Taguchi, T. Experimental cancer cachexia induced by transplantable colon 26 adenocarcinoma in mice. Cancer Res. 1990, 50, 2290–2295. [Google Scholar] [PubMed]
- Sherry, B.A.; Gelin, J.; Fong, Y.; Marano, M.; Wei, H.; Cerami, A.; Lowry, S.F.; Lundholm, K.G.; Moldawer, L.L. Anticachectin/tumor necrosis factor-α antibodies attenuate development of cachexia in tumor models. FASEB J. 1989, 3, 1956–1962. [Google Scholar] [CrossRef]
- Wang, S.; Lai, X.; Deng, Y.; Song, Y. Correlation between mouse age and human age in anti-tumor research: Significance and method establishment. Life Sci. 2019, 242, 117242. [Google Scholar] [CrossRef]
- Beheshti, A.; Benzekry, S.; McDonald, J.; Ma, L.; Peluso, M.; Hahnfeldt, P.; Hlatky, L. Host Age Is a Systemic Regulator of Gene Expression Impacting Cancer Progression. Cancer Res. 2015, 75, 1134–1143. [Google Scholar] [CrossRef] [Green Version]
- Simon, M.M.; Greenaway, S.; White, J.K.; Fuchs, H.; Gailus-Durner, V.; Wells, S.; Sorg, T.; Wong, K.; Bedu, E.; Cartwright, E.J.; et al. A comparative phenotypic and genomic analysis of C57BL/6J and C57BL/6N mouse strains. Genome Biol. 2013, 14, R82. [Google Scholar] [CrossRef]
- Rashidi, B.; Yang, M.; Jiang, P.; Baranov, E.; An, Z.; Wang, X.; Moossa, A.; Hoffman, R. A highly metastatic Lewis lung carcinoma orthotopic green fluorescent protein model. Clin. Exp. Metastasis 2000, 18, 57–60. [Google Scholar] [CrossRef]
- Feldman, J.P.; Goldwasser, R.; Mark, S.; Schwartz, J.; Orion, I.A. A mathematical model for tumor volume evaluation using two-dimensions. J. Appl. Quant. Methods 2009, 4, 455–462. [Google Scholar]
- Ullman-Culleré, M.H.; Foltz, C.J. Body condition scoring: A rapid and accurate method for assessing health status in mice. Lab. Anim. Sci. 1999, 49, 319–323. [Google Scholar]
- Evans, W.J.; Morley, J.E.; Argilés, J.; Bales, C.; Baracos, V.; Guttridge, D.; Jatoi, A.; Kalantar-Zadeh, K.; Lochs, H.; Mantovani, G.; et al. Cachexia: A new definition. Clin. Nutr. 2008, 27, 793–799. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Guaraldi, G.; Zona, S.; Silva, A.R.; Menozzi, M.; Dolci, G.; Milic, J.; Carli, F.; Mussini, C. The dynamic association between Frailty, CD4 and CD4/CD8 ratio in people aging with HIV. PLoS ONE 2019, 14, e0212283. [Google Scholar] [CrossRef]
- Sim, B.-C.; Aftahi, N.; Reilly, C.; Bogen, B.; Schwartz, R.H.; Gascoigne, N.R.; Lo, D. Thymic skewing of the CD4/CD8 ratio maps with the T-cell receptor α-chain locus. Curr. Biol. 1998, 8, 701–704. [Google Scholar] [CrossRef] [Green Version]
- Garg, S.K.; Delaney, C.E.; Toubai, T.; Ghosh, A.; Reddy, P.; Banerjee, R.; Yung, R. Aging is associated with increased regulatory T-cell function. Aging Cell 2014, 13, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Rohm, M.; Zeigerer, A.; Machado, J.; Herzig, S. Energy metabolism in cachexia. EMBO Rep. 2019, 20. [Google Scholar] [CrossRef]
- Talbert, E.E.; Metzger, G.A.; He, W.A.; Guttridge, D.C. Modeling human cancer cachexia in colon 26 tumor-bearing adult mice. J. Cachex- Sarcopenia Muscle 2014, 5, 321–328. [Google Scholar] [CrossRef]
- Ebert, S.M.; Al-Zougbi, A.; Bodine, S.C.; Adams, C.M. Skeletal Muscle Atrophy: Discovery of Mechanisms and Potential Therapies. Physiology 2019, 34, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.T.; Ma, J.F.; Di Marco, S.; Gallouzi, I.-E. Inducible nitric oxide synthase (iNOS) in muscle wasting syndrome, sarcopenia, and cachexia. Aging 2011, 3, 702–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.M.; Kang, J. Prognostic impact of myosteatosis in patients with colorectal cancer: A systematic review and meta-analysis. J. Cachex- Sarcopenia Muscle 2020, 11, 1270–1282. [Google Scholar] [CrossRef] [PubMed]
- Rohm, M.; Schäfer, M.; Laurent, V.; Üstünel, B.E.; Niopek, K.; Algire, C.; Hautzinger, O.; Sijmonsma, T.P.; Zota, A.; Medrikova, D.; et al. An AMP-activated protein kinase–stabilizing peptide ameliorates adipose tissue wasting in cancer cachexia in mice. Nat. Med. 2016, 22, 1120–1130. [Google Scholar] [CrossRef]
- Das, S.K.; Eder, S.; Schauer, S.; Diwoky, C.; Temmel, H.; Guertl, B.; Gorkiewicz, G.; Tamilarasan, K.P.; Kumari, P.; Trauner, M.; et al. Adipose Triglyceride Lipase Contributes to Cancer-Associated Cachexia. Science 2011, 333, 233–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morigny, P.; Zuber, J.; Haid, M.; Kaltenecker, D.; Riols, F.; Lima, J.D.; Simoes, E.; Otoch, J.P.; Schmidt, S.F.; Herzig, S.; et al. High levels of modified ceramides are a defining feature of murine and human cancer cachexia. J. Cachex. Sarcopenia Muscle 2020, 11, 1459–1475. [Google Scholar] [CrossRef]
- Martignoni, M.E.; Kunze, P.; Hildebrandt, W.; Künzli, B.; Berberat, P.; Giese, T.; Klöters, O.; Hammer, J.; Büchler, M.W.; Giese, N.A.; et al. Role of Mononuclear Cells and Inflammatory Cytokines in Pancreatic Cancer-Related Cachexia. Clin. Cancer Res. 2005, 11, 5802–5808. [Google Scholar] [CrossRef] [Green Version]
- Suriben, R.; Chen, M.; Higbee, J.; Oeffinger, J.; Ventura, R.; Li, B.; Mondal, K.; Gao, Z.; Ayupova, D.; Taskar, P.; et al. Antibody-mediated inhibition of GDF15–GFRAL activity reverses cancer cachexia in mice. Nat. Med. 2020, 26, 1264–1270. [Google Scholar] [CrossRef]
- Rohm, M.; Herzig, S. An Antibody Attack against Body Wasting in Cancer. Cell Metab. 2020, 32, 331–333. [Google Scholar] [CrossRef]
- Coletti, D.; Berardi, E.; Aulino, P.; Rossi, E.; Moresi, V.; Li, Z.; Adamo, S. Substrains of Inbred Mice Differ in Their Physical Activity as a Behavior. Sci. World J. 2013, 2013, 237260. [Google Scholar] [CrossRef] [Green Version]
- Enríquez, J.A. Mind your mouse strain. Nat. Metab. 2019, 1, 5–7. [Google Scholar] [CrossRef]
- Sayer, A.A.; Robinson, S.M.; Patel, H.P.; Shavlakadze, T.; Cooper, C.; Grounds, M.D. New horizons in the pathogenesis, diagnosis and management of sarcopenia. Age Ageing 2013, 42, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Gaugler, M.; Brown, A.; Merrell, E.; DiSanto-Rose, M.; Rathmacher, J.A.; Reynolds, T.H. PKB signaling and atrogene expression in skeletal muscle of aged mice. J. Appl. Physiol. 2011, 111, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Niu, M.; Li, L.; Su, Z.; Wei, L.; Pu, W.; Zhao, C.; Ding, Y.; Wazir, J.; Cao, W.; Song, S.; et al. An integrative transcriptome study reveals Ddit4/Redd1 as a key regulator of cancer cachexia in rodent models. Cell Death Dis. 2021, 12, 652. [Google Scholar] [CrossRef]
- Aguilar-Cazares, D.; Chavez-Dominguez, R.; Carlos-Reyes, A.; Lopez-Camarillo, C.; de la Cruz, O.N.H.; Lopez-Gonzalez, J.S. Contribution of Angiogenesis to Inflammation and Cancer. Front. Oncol. 2019, 9, 1399. [Google Scholar] [CrossRef] [Green Version]
- Mohr, H.; Ballke, S.; Bechmann, N.; Gulde, S.; Malekzadeh-Najafabadi, J.; Peitzsch, M.; Ntziachristos, V.; Steiger, K.; Wiedemann, T.; Pellegata, N.S. Mutation of the Cell Cycle Regulator p27kip1 Drives Pseudohypoxic Pheochromocytoma Development. Cancers 2021, 13, 126. [Google Scholar] [CrossRef]
- Rupert, J.E.; Narasimhan, A.; Jengelley, D.H.; Jiang, Y.; Liu, J.; Au, E.; Silverman, L.M.; Sandusky, G.; Bonetto, A.; Cao, S.; et al. Tumor-derived IL-6 and trans-signaling among tumor, fat, and muscle mediate pancreatic cancer cachexia. J. Exp. Med. 2021, 218, e20190450. [Google Scholar] [CrossRef]
- Ershler, W.B.; Sun, W.H.; Binkley, N.; Gravenstein, S.; Volk, M.J.; Kamoske, G.; Klopp, R.G.; Roecker, E.B.; Daynes, R.A.; Weindruch, R. Interleukin-6 and aging: Blood levels and mononuclear cell production increase with advancing age and in vitro production is modifiable by dietary restriction. Lymphokine Cytokine Res. 1993, 12, 225–230. [Google Scholar]
- Osawa, Y.; Semba, R.D.; Fantoni, G.; Candia, J.; Biancotto, A.; Tanaka, T.; Bandinelli, S.; Ferrucci, L. Plasma proteomic signature of the risk of developing mobility disability: A 9-year follow-up. Aging Cell 2020, 19, e13132. [Google Scholar] [CrossRef]
- Laurens, C.; Parmar, A.; Murphy, E.; Carper, D.; Lair, B.; Maes, P.; Vion, J.; Boulet, N.; Fontaine, C.; Marquès, M.; et al. Growth and differentiation factor 15 is secreted by skeletal muscle during exercise and promotes lipolysis in humans. JCI Insight 2020, 5, e131870. [Google Scholar] [CrossRef] [Green Version]
- Gil, C.I.; Ost, M.; Kasch, J.; Schumann, S.; Heider, S.; Klaus, S. Role of GDF15 in active lifestyle induced metabolic adaptations and acute exercise response in mice. Sci. Rep. 2019, 9, 20120. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.C.; Glass, D.J.; Guttridge, D.C. Cancer Cachexia: Mediators, Signaling, and Metabolic Pathways. Cell Metab. 2012, 16, 153–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, H.; Numata, K.; Ito, T.; Takagi, K.; Matsukawa, A. Innate immune response in th1- and th2-dominant mouse strains. Shock 2004, 22, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Zhao, K.; Jose, I.; Hoogenraad, N.; Osellame, L. Biomarkers for Cancer Cachexia: A Mini Review. Int. J. Mol. Sci. 2021, 22, 4501. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | n | Cholesterol (mg/dL) | LDL (mg/dL) | NEFA (mmol/L) | Glucose (mg/dL) | Albumin (g/L) | Total Protein (g/L) | n | IGF-1 (ng/mL) |
|---|---|---|---|---|---|---|---|---|---|
| PBS/N young | 6 | 80.67 ± 2.99 | 9.53 ± 0.36 | 0.42 ± 0.04 | 319.5 ± 16.85 | 25.74 ± 0.35 | 45.37 ± 0.65 | 3 | 379.16 ± 28.45 |
| LLC/N young | 9 | 93.67 ± 3.80 | 21.37 ± 1.58 **** | 0.30 ± 0.04 | 288.7 ± 9.472 | 21.75 ± 0.82 ** | 42.80 ± 1.31 | 9 | 292.10 ± 20.64 |
| PBS/N aged | 5 | 88.00 ± 2.30 | 17.80 ± 0.37 ## | 0.27 ± 0.07 | 213.0 ± 11.34 #### | 24.19 ± 0.55 | 48.93 ± 1.15 | 5 | 664.42 ± 21.50 ### |
| LLC/N aged | 7 | 85.71 ± 3.91 | 22.56 ± 1.93 | 0.19 ± 0.03 | 185.0 ± 10.75 #### | 23.03 ± 0.78 | 47.10 ± 1.35 | 7 | 526.10 ± 43.70 *#### |
| PBS/J young | 8 | 69.38 ± 2.23 | 11.84 ± 0.95 | 0.36 ± 0.03 | 309.1 ± 18.62 | 21.82 ± 0.39 | 40.02 ± 0.51 | 5 | 236.88 ± 24.11 |
| LLC/J young | 12 | 82.25 ± 3.38 | 18.04 ± 1.33 ** | 0.41 ± 0.03 | 356.2 ± 20.91 | 21.98 ± 0.44 | 41.56 ± 0.91 | 6 | 290.43 ± 11.38 |
| PBS/J aged | 10 | 76.20 ± 5.89 | 11.18 ± 0.51 | 0.15 ± 0.02 #### | 269.8 ± 16.15 | 24.84 ± 0.88 # | 46.48 ± 1.04 ### | 5 | 322.30 ± 18.08 # |
| LLC/J aged | 12 | 85.08 ± 4.20 | 20.08 ± 1.26 **** | 0.26 ± 0.02 ## | 279.1 ± 14.96 # | 21.12 ± 0.53 *** | 42.15 ± 0.87 ** | 6 | 305.80 ± 10.53 |
| PBS/BALBc young | 5 | 109.60 ± 3.43 | 8.84 ± 0.34 | 0.30 ± 0.10 | 325.60 ± 15.69 | 21.05 ± 0.51 | 41.49 ± 2.19 | 5 | 459.30 ± 34.02 |
| C26 young | 6 | 123.50 ± 4.01 | 18.85 ± 1.34 **** | 0.14 ± 0.03 | 220.00 ± 11.69 *** | 22.68 ± 0.44 | 52.27 ± 0.90 **** | 5 | 389.18 ± 18.50 |
| PBS/BALBc aged | 5 | 126.80 ± 4.04 | 12.36 ± 0.52 | 0.22 ± 0.02 | 209.00 ± 17.641 ### | 22.52 ± 0.76 | 43.68 ± 0.99 | 5 | 510.96 ± 25.08 |
| C26 aged | 10 | 125.50 ± 4.68 | 18.64 ± 0.93 *** | 0.18 ± 0.01 | 184.80 ± 11.38 | 20.83 ± 0.42 | 46.88 ± 0.87 # | 7 | 475.60 ± 25.79 |
| Young (≤55y) | Aged (>55y) | |||||
|---|---|---|---|---|---|---|
| Plasma | n | R2 | p Value | n | R2 | p Value |
| IFNγ | 14 | 0.01020 | 0.7312 | 14 | 0.004835 | 0.8133 |
| IL10 | 14 | 0.5571 | 0.0022 | 14 | 0.003069 | 0.8508 |
| IL13 | 14 | 0.1103 | 0.2461 | 14 | 0.04233 | 0.4804 |
| IL1b | 14 | 0.4384 | 0.0099 | 14 | 0.006826 | 0.7789 |
| IL6 | 14 | 0.4499 | 0.0086 | 14 | 0.2428 | 0.0734 |
| IL8 | 14 | 0.02732 | 0.5722 | 14 | 0.006336 | 0.7868 |
| MCP1 | 14 | 0.2092 | 0.1001 | 14 | 0.04584 | 0.4623 |
| MIP1A | 14 | 0.001337 | 0.9012 | 14 | 6.88 × 10−7 | 0.9978 |
| MIP1B | 14 | 0.1588 | 0.1581 | 14 | 0.02744 | 0.5886 |
| Tnfa | 14 | 0.1056 | 0.2570 | 14 | 0.01546 | 0.6719 |
| Hemoglobin | 14 | 0.2449 | 0.1019 | 14 | 0.4042 | 0.0355 |
| CRP | 14 | 0.09554 | 0.2822 | 14 | 0.06038 | 0.3971 |
| Albumin | 14 | 0.1068 | 0.2542 | 14 | 0.1586 | 0.1584 |
| Cholesterol | 13 | 0.1093 | 0.2698 | 14 | 0.02556 | 0.5851 |
| HDL | 14 | 0.1124 | 0.2413 | 14 | 0.05848 | 0.4049 |
| LDL | 14 | 0.01040 | 0.7286 | 14 | 0.02746 | 0.5713 |
| Triglycerides | 14 | 0.06374 | 0.3839 | 14 | 0.1144 | 0.2369 |
| Clinical Parameters | Young Patients (≤55 years) | Aged Patients (>55 years) | p Value |
|---|---|---|---|
| Overall (n) | 14 | 14 | |
| Female/Male (n) | 7/7 | 8/6 | |
| Age (years) | 44.86 ± 1.565 | 68.21 ± 2.204 | <0.0001 |
| BW loss (%) | −8.010 ± 2.602 | −7.171 ± 2.216 | 0.8080 |
| BMI (kg/m2) | 24.51 ± 1.173 | 23.56 ± 1.016 | 0.5432 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geppert, J.; Walth, A.A.; Terrón Expósito, R.; Kaltenecker, D.; Morigny, P.; Machado, J.; Becker, M.; Simoes, E.; Lima, J.D.C.C.; Daniel, C.; et al. Aging Aggravates Cachexia in Tumor-Bearing Mice. Cancers 2022, 14, 90. https://doi.org/10.3390/cancers14010090
Geppert J, Walth AA, Terrón Expósito R, Kaltenecker D, Morigny P, Machado J, Becker M, Simoes E, Lima JDCC, Daniel C, et al. Aging Aggravates Cachexia in Tumor-Bearing Mice. Cancers. 2022; 14(1):90. https://doi.org/10.3390/cancers14010090
Chicago/Turabian StyleGeppert, Julia, Alina A. Walth, Raúl Terrón Expósito, Doris Kaltenecker, Pauline Morigny, Juliano Machado, Maike Becker, Estefania Simoes, Joanna D. C. C. Lima, Carolin Daniel, and et al. 2022. "Aging Aggravates Cachexia in Tumor-Bearing Mice" Cancers 14, no. 1: 90. https://doi.org/10.3390/cancers14010090
APA StyleGeppert, J., Walth, A. A., Terrón Expósito, R., Kaltenecker, D., Morigny, P., Machado, J., Becker, M., Simoes, E., Lima, J. D. C. C., Daniel, C., Berriel Diaz, M., Herzig, S., Seelaender, M., & Rohm, M. (2022). Aging Aggravates Cachexia in Tumor-Bearing Mice. Cancers, 14(1), 90. https://doi.org/10.3390/cancers14010090