
miRNA Expression Characterizes Histological Subtypes and Metastasis in Penile Squamous Cell Carcinoma

 , ,
, ,  ,
, 

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Patient Data
2.2. DNA and RNA Isolation
2.3. miRNA Microarray and Data Analysis
2.4. Real-Time RT-PCR Quantification of miRNAs
3. Results
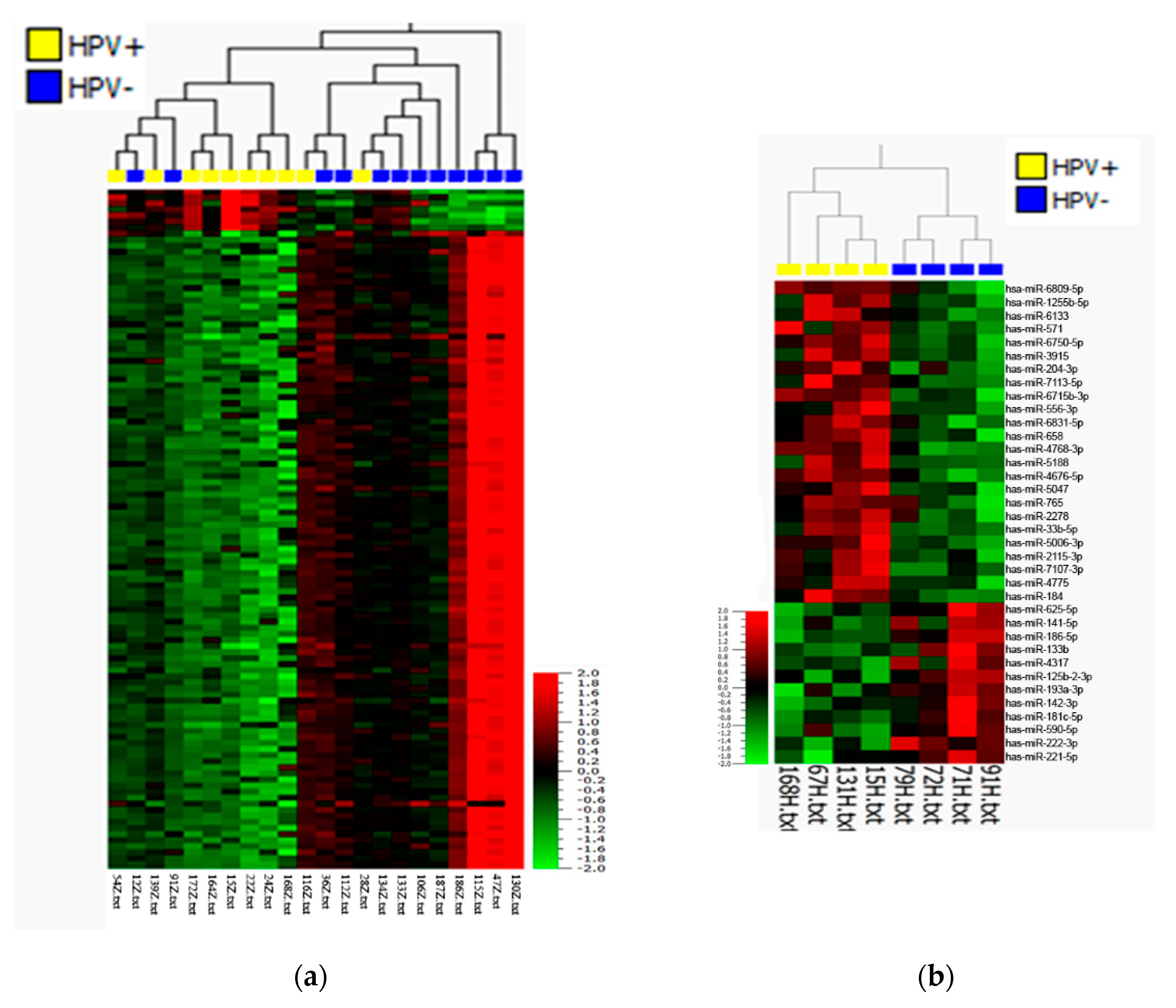
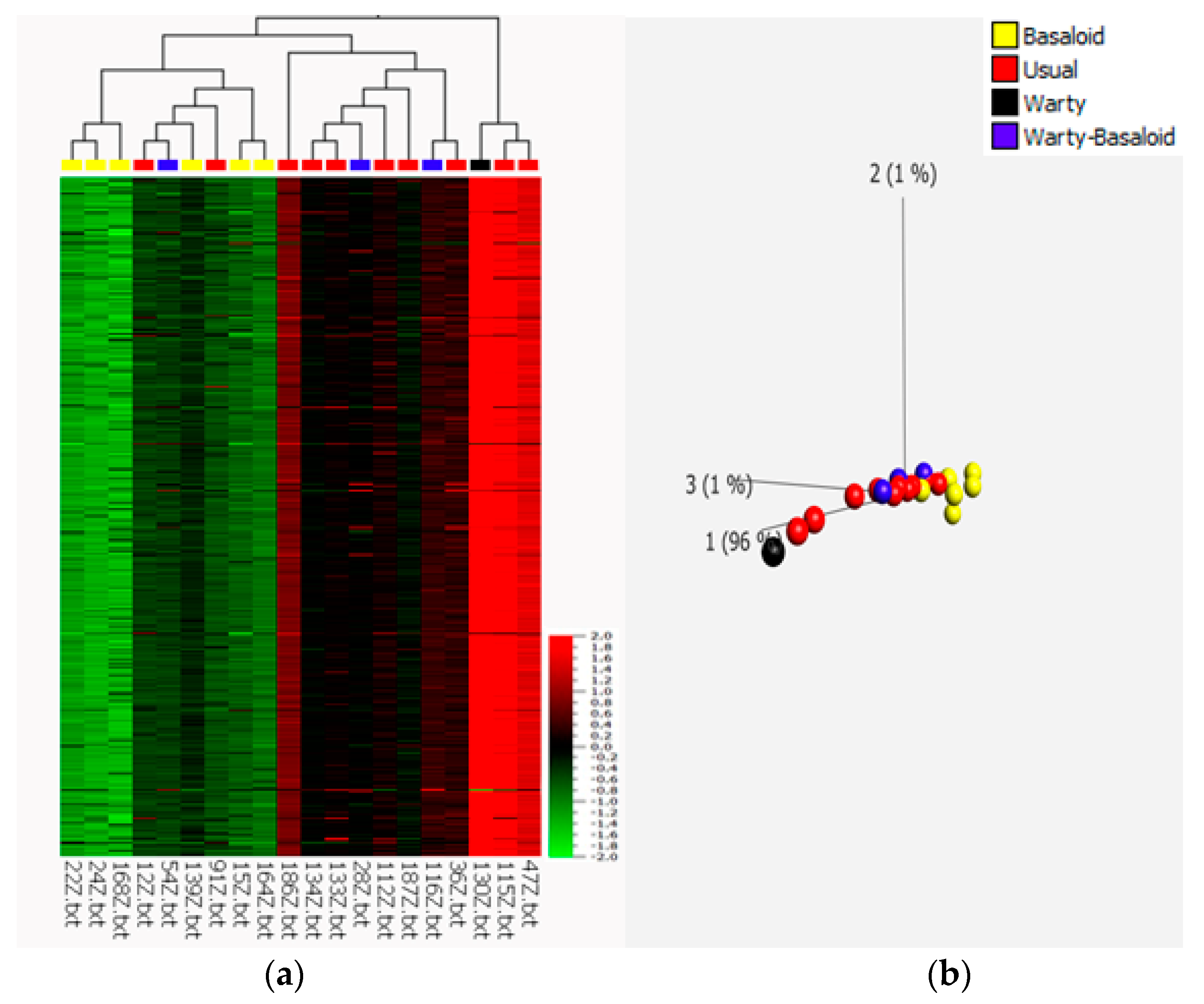
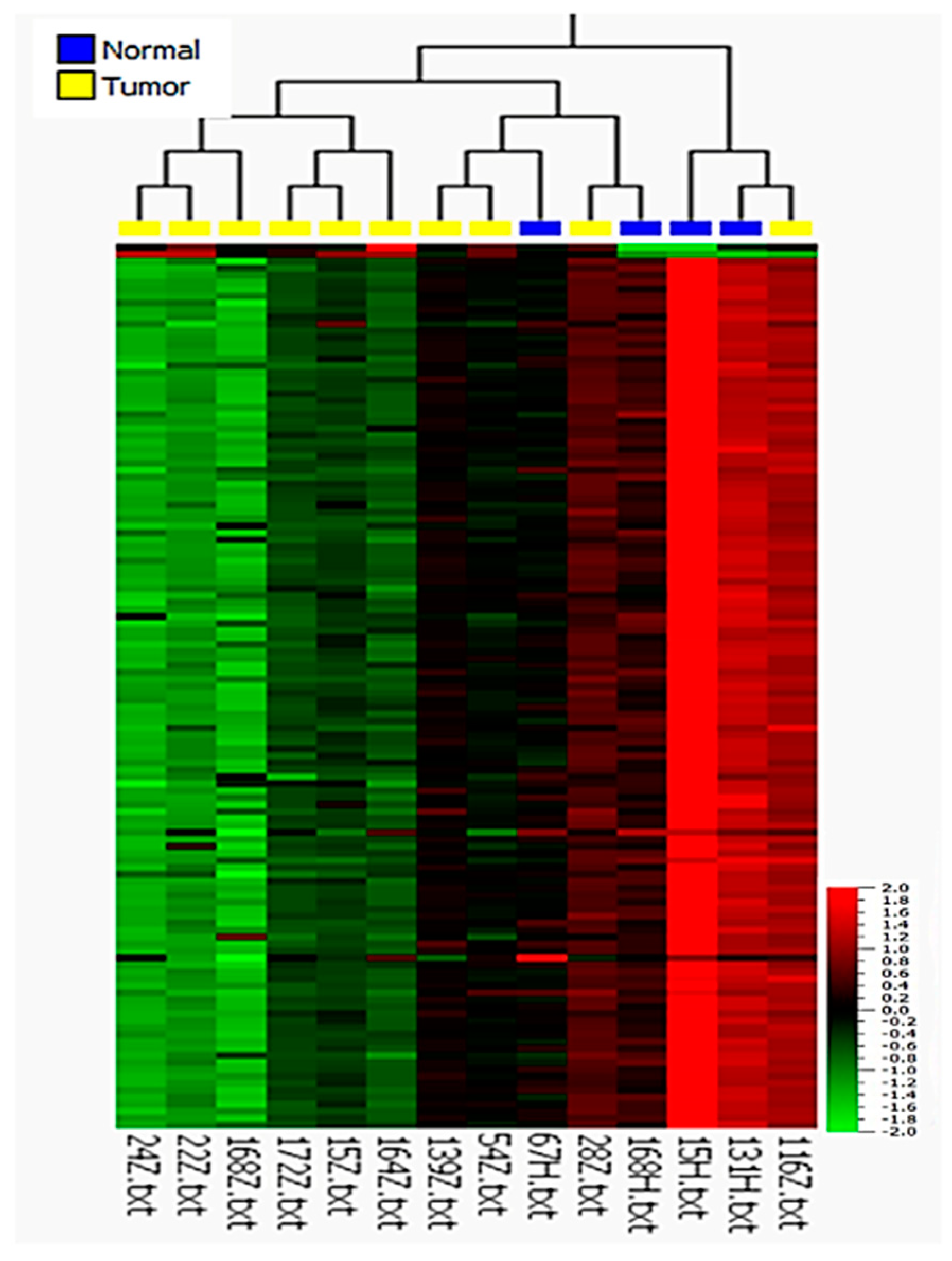
3.1. Microarray Analysis
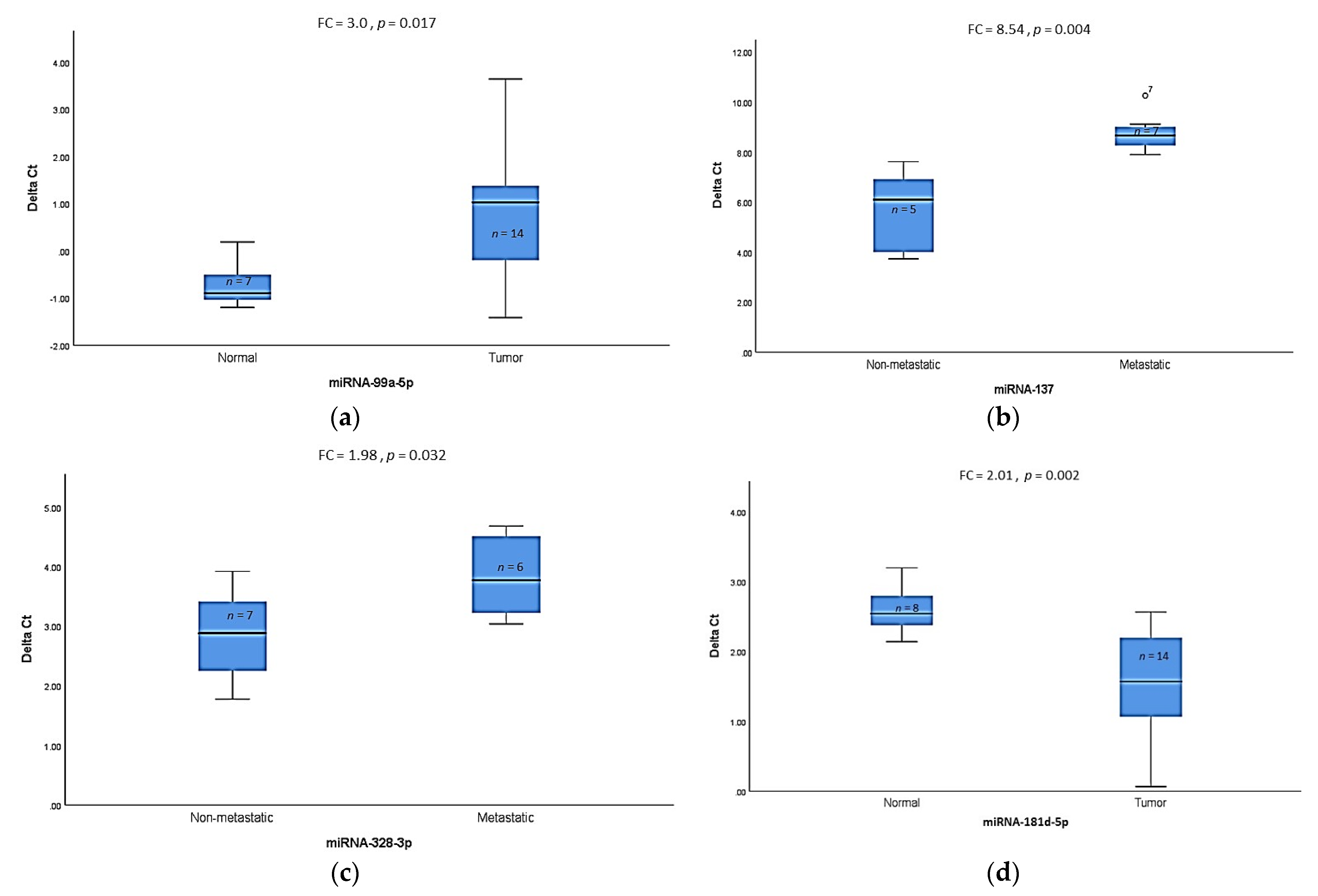
3.2. Validation of Differentially Expressed miRNAs by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bleeker, M.C.G.; Heideman, D.A.M.; Snijders, P.J.F.; Horenblas, S.; Dillner, J.; Meijer, C.J.L.M. Penile cancer: Epidemiology, pathogenesis and prevention. World J. Urol. 2009, 27, 141–150. [Google Scholar] [CrossRef]
- Jemal, A.; Siegel, R.; Ward, E.; Murray, T.; Xu, J.; Thun, M.J. Cancer Statistics. CA A Cancer J. Clin. 2007, 57, 43–66. [Google Scholar] [CrossRef]
- Parkin, D.M.; Muir, C.S. Cancer Incidence in Five Continents. Comparability and quality of data. IARC Sci. Publ. 1992, 120, 45–173. [Google Scholar]
- Olesen, T.B.; Sand, F.L.; Rasmussen, C.L.; Albieri, V.; Toft, B.G.; Norrild, B.; Munk, C.; Kjær, S.K. Prevalence of human papillomavirus DNA and p16INK4a in penile cancer and penile intraepithelial neoplasia: A systematic review and meta-analysis. Lancet Oncol. 2019, 20, 145–158. [Google Scholar] [CrossRef]
- Moch, H.; Cubilla, A.L.; Humphrey, P.A.; Reuter, V.E.; Ulbright, T.M. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs—Part A: Renal, Penile, and Testicular Tumours. Eur. Urol. 2016, 70, 93–105. [Google Scholar] [CrossRef]
- Miralles-Guri, C.; Bruni, L.; Cubilla, A.L.; Castellsagué, X.; Bosch, F.X.; De Sanjosé, S. Human papillomavirus prevalence and type distribution in penile carcinoma. J. Clin. Pathol. 2009, 62, 870–878. [Google Scholar] [CrossRef]
- Chaux, A.; Pfannl, R.; Rodríguez, I.M.; Barreto, J.E.; Velazquez, E.F.; Lezcano, C.; Piris, A.; Netto, G.J.; Cubilla, A.L. Distinctive Immunohistochemical Profile of Penile Intraepithelial Lesions. Am. J. Surg. Pathol. 2011, 35, 553–562. [Google Scholar] [CrossRef]
- Pinto, A.P.; Miron, A.; Yassin, Y.; Monte, N.; Woo, T.Y.C.; Mehra, K.K.; Medeiros, F.; Crum, C.P. Differentiated vulvar intraepithelial neoplasia contains Tp53 mutations and is genetically linked to vulvar squamous cell carcinoma. Mod. Pathol. 2010, 23, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Rocha, R.M.; Ignácio, J.A.; Jordán, J.; Carraro, D.M.; Lisboa, B.; Lopes, A.; Carvalho, K.C.; Da Cunha, I.W.; Cubilla, A.; Guimarães, G.C.; et al. A clinical, pathologic, and molecular study of p53 and murine double minute 2 in penile carcinogenesis and its relation to prognosis. Hum. Pathol. 2012, 43, 481–488. [Google Scholar] [CrossRef]
- Ferreux, E.; Lont, A.P.; Horenblas, S.; Gallee, M.P.W.; Raaphorst, F.M.; Doeberitz, M.V.K.; Meijer, C.J.L.M.; Snijders, P.J.F. Evidence for at least three alternative mechanisms targeting the p16INK4A/cyclin D/Rb pathway in penile carcinoma, one of which is mediated by high-risk human papillomavirus. J. Pathol. 2003, 201, 109–118. [Google Scholar] [CrossRef]
- Emmanuel, A.; Nettleton, J.; Watkin, N.; Berney, D.M. The molecular pathogenesis of penile carcinoma—current developments and understanding. Virchows Archiv. 2019, 475, 397–405. [Google Scholar] [CrossRef]
- Vanthoor, J.; Vos, G.; Albersen, M. Penile cancer: Potential target for immunotherapy? World J. Urol. 2020, 1–7. [Google Scholar] [CrossRef]
- Mannweiler, S.; Sygulla, S.; Winter, E.; Regauer, S. Two major pathways of penile carcinogenesis: HPV-induced penile cancers overexpress p16ink4a, HPV-negative cancers associated with dermatoses express p53, but lack p16ink4a overexpression. J. Am. Acad. Dermatol. 2013, 69, 73–81. [Google Scholar] [CrossRef]
- Afonso, L.A.; Carestiato, F.N.; Ornellas, A.A.; Ornellas, P.; Rocha, W.M.; Cordeiro, T.I.; Lisboa, D.C.; Alves, G.B.; Cavalcanti, S.M.B. Human papillomavirus, Epstein-Barr virus, and methylation status of p16ink4ain penile cancer. J. Med Virol. 2017, 89, 1837–1843. [Google Scholar] [CrossRef]
- Gage, J.R.; Meyers, C.; O Wettstein, F. The E7 proteins of the nononcogenic human papillomavirus type 6b (HPV-6b) and of the oncogenic HPV-16 differ in retinoblastoma protein binding and other properties. J. Virol. 1990, 64, 723–730. [Google Scholar] [CrossRef]
- Wang, F.; Li, B.; Xie, X. The roles and clinical significance of microRNAs in cervical cancer. Histol. Histopathol. 2015, 31. [Google Scholar]
- Zheng, Z.-M.; Wang, X. Regulation of cellular miRNA expression by human papillomaviruses. Biochim. Biophys. Acta (BBA) Bioenerg. 2011, 1809, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Snoek, B.C.; Babion, I.; Koppers-Lalic, D.; Pegtel, D.M.; Steenbergen, R.D. Altered microRNA processing proteins in HPV-induced cancers. Curr. Opin. Virol. 2019, 39, 23–32. [Google Scholar] [CrossRef]
- Hölters, S.; Khalmurzaev, O.; Pryalukhin, A.; Loertzer, P.; Janssen, M.; Heinzelbecker, J.; Ueberdiek, S.; Pfuhl, T.; Smola, S.; Agaimy, A.; et al. Challenging the prognostic impact of the new WHO and TNM classifications with special emphasis on HPV status in penile carcinoma. Virchows Archiv. 2019, 475, 211–221. [Google Scholar] [CrossRef]
- Sun, M.; Djajadiningrat, R.S.; Alnajjar, H.M.; Trinh, Q.-D.; Graafland, N.M.; Watkin, N.; Karakiewicz, P.I.; Horenblas, S. Development and external validation of a prognostic tool for prediction of cancer-specific mortality after complete loco-regional pathological staging for squamous cell carcinoma of the penis. BJU Int. 2014, 116, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Paner, G.P.; Stadler, W.M.; Hansel, D.E.; Montironi, R.; Lin, D.W.; Amin, M.B. Updates in the Eighth Edition of the Tumor-Node-Metastasis Staging Classification for Urologic Cancers. Eur. Urol. 2018, 73, 560–569. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef]
- Bala, S.; Marcos, M.; Szabo, G. Emerging role of microRNAs in liver diseases. World J. Gastroenterol. 2009, 15, 5633–5640. [Google Scholar] [CrossRef]
- Lee, K.T.-W.; Tan, J.-K.; Lam, A.K.-Y.; Gan, S.-Y. MicroRNAs serving as potential biomarkers and therapeutic targets in nasopharyngeal carcinoma: A critical review. Crit. Rev. Oncol. 2016, 103, 1–9. [Google Scholar] [CrossRef]
- Cho, W.C. MicroRNAs: Potential biomarkers for cancer diagnosis, prognosis and targets for therapy. Int. J. Biochem. Cell Biol. 2010, 42, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Wald, A.I.; Hoskins, E.E.; Wells, S.I.; Ferris, R.L.; Khan, S.A. Human papillomavirus alters microRNA profiles in squamous cell carcinoma of the head and neck (SCCHN) cell lines. Head Neck 2011, 33, 504–512. [Google Scholar] [CrossRef]
- Gao, D.; Zhang, Y.; Zhu, M.; Liu, S.; Wang, X. miRNA Expression Profiles of HPV-Infected Patients with Cervical Cancer in the Uyghur Population in China. PLoS ONE 2016, 11, e0164701. [Google Scholar] [CrossRef]
- Santos, J.M.; Da Silva, S.P.; Costa, N.R.; Gil Da Costa, R.M.; Medeiros, R. The Role of MicroRNAs in the Metastatic Process of High-Risk HPV-Induced Cancers. Cancers 2018, 10, 493. [Google Scholar] [CrossRef]
- Husman, A.-M.D.R.; Walboomers, J.M.M.; Brule, A.J.C.V.D.; Meijer, C.J.L.M.; Snijders, P.J.F. The use of general primers GP5 and GP6 elongated at their 3′ ends with adjacent highly conserved sequences improves human papillomavirus detection by PCR. J. Gen. Virol. 1995, 76, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Djajadiningrat, R.S.; Jordanova, E.S.; Kroon, B.K.; Van Werkhoven, E.; De Jong, J.; Pronk, D.T.; Snijders, P.J.; Horenblas, S.; Heideman, D.A. Human Papillomavirus Prevalence in Invasive Penile Cancer and Association with Clinical Outcome. J. Urol. 2015, 193, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, L.; Cubilla, A.L.; Reuter, V.E.; Haas, G.P.; Lancaster, W.D. Preferential Association of Human Papillomavirus With High-Grade Histologic Variants of Penile-Invasive Squamous Cell Carcinoma. J. Natl. Cancer Inst. 1995, 87, 1705–1709. [Google Scholar] [CrossRef] [PubMed]
- Lont, A.P.; Kroon, B.K.; Horenblas, S.; Gallee, M.P.W.; Berkhof, J.; Meijer, C.J.L.M.; Snijders, P.J.F. Presence of high-risk human papillomavirus DNA in penile carcinoma predicts favorable outcome in survival. Int. J. Cancer 2006, 119, 1078–1081. [Google Scholar] [CrossRef] [PubMed]
- Kuasne, H.; Barros-Filho, M.C.; Busso-Lopes, A.; Marchi, F.A.; Pinheiro, M.; Muñoz, J.J.M.; Scapulatempo-Neto, C.; Faria, E.F.; Guimarães, G.C.; Lopes, A.; et al. Integrative miRNA and mRNA analysis in penile carcinomas reveals markers and pathways with potential clinical impact. Oncotarget 2017, 8, 15294–15306. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Cappellesso, R.; Peta, E.; Militello, V.; Sinigaglia, A.; Fassan, M.; Simonato, F.; Guzzardo, V.; Ventura, L.; Blandamura, S.; et al. Profiling of Expression of Human Papillomavirus–Related Cancer miRNAs in Penile Squamous Cell Carcinomas. Am. J. Pathol. 2014, 184, 3376–3383. [Google Scholar] [CrossRef] [PubMed]
- Peta, E.; Cappellesso, R.; Masi, G.; Sinigaglia, A.; Trevisan, M.; Grassi, A.; Di Camillo, B.; Vassarotto, E.; Fassina, A.; Palù, G.; et al. Down-regulation of microRNA-146a is associated with high-risk human papillomavirus infection and epidermal growth factor receptor overexpression in penile squamous cell carcinoma. Hum. Pathol. 2017, 61, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.-F.; Lin, J.-F.; Chou, K.-Y.; Lin, Y.-C.; Chen, H.-E.; Hwang, T.I.-S. miR-99a-5p acts as tumor suppressor via targeting to mTOR and enhances RAD001-induced apoptosis in human urinary bladder urothelial carcinoma cells. OncoTargets Ther. 2018, 11, 239–252. [Google Scholar] [CrossRef]
- Chen, Y.-T.; Yao, J.-N.; Qin, Y.-T.; Hu, K.; Wu, F.; Fang, Y.-Y. Biological role and clinical value of miR-99a-5p in head and neck squamous cell carcinoma (HNSCC): A bioinformatics-based study. FEBS Open Bio 2018, 8, 1280–1298. [Google Scholar] [CrossRef]
- Saito, R.; Maruyama, S.; Kawaguchi, Y.; Akaike, H.; Shimizu, H.; Furuya, S.; Kawaida, H.; Ichikawa, D. miR-99a-5p as Possible Diagnostic and Prognostic Marker in Patients With Gastric Cancer. J. Surg. Res. 2020, 250, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wei, P.; Shen, X.; Zhang, Y.; Xu, B.; Zhou, J.; Fan, S.; Hao, Z.; Shi, H.; Zhang, X.; et al. MicroRNA Expression Profile in Penile Cancer Revealed by Next-Generation Small RNA Sequencing. PLoS ONE 2015, 10, e0131336. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, Z.; Zhang, X.; Liu, J.; Gui, J.; Cui, M.; Li, Y. miR-211-5p is down-regulated and a prognostic marker in bladder cancer. J. Gene Med. 2020, 22, 1–10. [Google Scholar] [CrossRef]
- Wang, K.; Jin, W.; Jin, P.; Fei, X.; Wang, X.; Chen, X. miR-211-5p Suppresses Metastatic Behavior by Targeting SNAI1 in Renal Cancer. Mol. Cancer Res. 2017, 15, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Pan, X.; He, T.; Lin, C.; Lai, Y.; Chen, P.; Zhang, Z.; Yang, S.; Wang, T.; Lai, Y. Tumor suppressor miR-211-5p is associated with cellular migration, proliferation and apoptosis in renal cell carcinoma. Exp. Ther. Med. 2018, 15, 4019–4028. [Google Scholar] [CrossRef]
- Chen, L.-L.; Zhang, Z.-J.; Yi, Z.-B.; Li, J.-J. MicroRNA-211-5p suppresses tumour cell proliferation, invasion, migration and metastasis in triple-negative breast cancer by directly targeting SETBP. Br. J. Cancer 2017, 117, 78–88. [Google Scholar] [CrossRef]
- Wang, B.; Hsu, S.-H.; Majumder, S.K.; Kutay, H.; Huang, W.; Jacob, S.T.; Ghoshal, K. TGFβ-mediated upregulation of hepatic miR-181b promotes hepatocarcinogenesis by targeting TIMP. Oncogene 2009, 29, 1787–1797. [Google Scholar] [CrossRef]
- Wang, H.; Wei, H.; Wang, J.; Li, L.; Chen, A.; Li, Z. MicroRNA-181d-5p-Containing Exosomes Derived from CAFs Promote EMT by Regulating CDX2/HOXA5 in Breast Cancer. Mol. Ther. Nucleic Acids 2020, 19, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Graule, J.; Uth, K.; Fischer, E.; Centeno, I.; Galván, J.A.; Eichmann, M.; Rau, T.T.; Langer, R.; Dawson, H.; Nitsche, U.; et al. CDX2 in colorectal cancer is an independent prognostic factor and regulated by promoter methylation and histone deacetylation in tumors of the serrated pathway. Clin. Epigenetics 2018, 10, 120. [Google Scholar] [CrossRef]
- Zhang, B.Y.; Jones, J.C.; Briggler, A.M.; Hubbard, J.M.; Kipp, B.R.; Sargent, D.J.; Dixon, J.G.; Grothey, A. Lack of Caudal-Type Homeobox Transcription Factor 2 Expression as a Prognostic Biomarker in Metastatic Colorectal Cancer. Clin. Color. Cancer 2017, 16, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-L.; Lu, S.-X.; Li, M.; Li, L.-Z.; Fu, J.; Hu, W.; Yang, Y.-Z.; Luo, R.-Z.; Zhang, C.Z.; Yun, J.-P. FoxD3-regulated microRNA-137 suppresses tumour growth and metastasis in human hepatocellular carcinoma by targeting AKT. Oncotarget 2014, 5, 5113–5124. [Google Scholar] [CrossRef]
- Zhu, L.-L.; Chen, W.; Wang, J.; Gan, T.; Wang, Y.-P.; Yang, J.-L. Expression Difference of MicroRNAs in Colorectal Adenoma and Colorectal Cancer. Sichuan Da Xue Xue Bao Yi Xue Ban J. Sichuan Univ. Med Sci. Ed. 2015, 46, 851–855. [Google Scholar]
- Chen, L.; Wang, X.; Wang, H.; Li, Y.; Yan, W.; Han, L.; Zhang, K.; Zhang, J.; Wang, Y.; Feng, Y.; et al. miR-137 is frequently down-regulated in glioblastoma and is a negative regulator of Cox. Eur. J. Cancer 2012, 48, 3104–3111. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Wang, L.; Ge, X.; Jiang, C.-F.; Shi, Z.-M.; Li, D.-M.; Liu, W.-T.; Yu, X.; Shu, Y.-Q. MicroRNA-137 inhibits tumor growth and sensitizes chemosensitivity to paclitaxel and cisplatin in lung cancer. Oncotarget 2016, 7, 20728–20742. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, J.; Zhou, Q. Elevated expression of microRNA-328-3p suppresses aggressive malignant behaviors via targeting matrix metalloprotease 16 in osteosarcoma. OncoTargets Ther. 2019, 12, 2063–2070. [Google Scholar] [CrossRef]
- Yang, S.-F.; Lee, W.-J.; Tan, P.; Tang, C.-H.; Hsiao, M.; Hsieh, F.-K.; Chien, M.-H. Upregulation of miR-328 and inhibition of CREB-DNA-binding activity are critical for resveratrol-mediated suppression of matrix metalloproteinase-2 and subsequent metastatic ability in human osteosarcomas. Oncotarget 2014, 6, 2736–2753. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-Z.; Li, J.; Liu, B.-Z. MicroRNA-328-3p inhibits malignant progression of hepatocellular carcinoma by regulating MMP-9 level. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 9331–9340. [Google Scholar]
- Pan, S.; Ren, F.; Li, L.; Liu, D.; Li, Y.; Wang, A.; Li, W.; Dong, Y.; Guo, W. MiR-328-3p inhibits cell proliferation and metastasis in colorectal cancer by targeting Girdin and inhibiting the PI3K/Akt signaling pathway. Exp. Cell Res. 2020, 390, 111939. [Google Scholar] [CrossRef]
- Lu, H.; Hu, J.; Li, J.; Lu, W.; Deng, X.; Wang, X. miR-328-3p overexpression attenuates the malignant proliferation and invasion of liver cancer via targeting Endoplasmic Reticulum Metallo Protease 1 to inhibit AKT phosphorylation. Ann. Transl. Med. 2020, 8, 754. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lin, W.; Tang, X.; Li, S.; Guo, L.; Lin, Y.; Kwok, H.F. The Roles of microRNAs in Regulating the Expression of PD-1/PD-L1 Immune Checkpoint. Int. J. Mol. Sci. 2017, 18, 2540. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Tang, J.; Sun, H.; Zheng, X.; Li, Z.; Sun, T.; Li, J.; Wang, S.; Zhou, X.; Sun, H.; et al. PD-L1 expression as poor prognostic factor in patients with non-squamous non-small cell lung cancer. Oncotarget 2017, 8, 58457–58468. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-M.; Sung, W.-W.; Hsieh, M.-J.; Tsai, S.-C.; Lai, H.-W.; Yang, S.-M.; Shen, K.-H.; Chen, M.-K.; Lee, H.; Yeh, K.-T.; et al. High PD-L1 Expression Correlates with Metastasis and Poor Prognosis in Oral Squamous Cell Carcinoma. PLoS ONE 2015, 10, e0142656. [Google Scholar] [CrossRef]
- Gu, X.; Dong, M.; Liu, Z.; Mi, Y.; Yang, J.; Zhang, Z.; Liu, K.; Jiang, L.; Zhang, Y.; Dong, S.; et al. Elevated PD-L1 expression predicts poor survival outcomes in patients with cervical cancer. Cancer Cell Int. 2019, 19, 146. [Google Scholar] [CrossRef] [PubMed]
- Ottenhof, S.R.; Djajadiningrat, R.S.; De Jong, J.; Thygesen, H.H.; Horenblas, S.; Jordanova, E.S. Expression of Programmed Death Ligand 1 in Penile Cancer is of Prognostic Value and Associated with HPV Status. J. Urol. 2017, 197, 690–697. [Google Scholar] [CrossRef]
- Udager, A.M.; Liu, T.-Y.; Skala, S.L.; Magers, M.J.; McDaniel, A.S.; Spratt, D.E.; Feng, F.Y.; Siddiqui, J.; Cao, X.; Fields, K.L.; et al. Frequent PD-L1 expression in primary and metastatic penile squamous cell carcinoma: Potential opportunities for immunotherapeutic approaches. Ann. Oncol. 2016, 27, 1706–1712. [Google Scholar] [CrossRef]
- Kao, S.C.; Cheng, Y.Y.; Williams, M.; Kirschner, M.B.; Madore, J.; Lum, T.; Sarun, K.H.; Linton, A.; McCaughan, B.; Klebe, S.; et al. Tumor Suppressor microRNAs Contribute to the Regulation of PD-L1 Expression in Malignant Pleural Mesothelioma. J. Thorac. Oncol. 2017, 12, 1421–1433. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, J.; Dong, K.; Lin, F.; Long, M.; Ouyang, Y.; Wei, J.; Chen, X.; Weng, Y.; He, T.; et al. Tumor suppressor miR-34a targets PD-L1 and functions as a potential immunotherapeutic target in acute myeloid leukemia. Cell. Signal. 2015, 27, 443–452. [Google Scholar] [CrossRef]
- Jia, L.; Xi, Q.; Wang, H.; Zhang, Z.; Liu, H.; Cheng, Y.; Guo, X.; Zhang, J.; Zhang, Q.; Zhang, L.; et al. miR-142-5p regulates tumor cell PD-L1 expression and enhances anti-tumor immunity. Biochem. Biophys. Res. Commun. 2017, 488, 425–431. [Google Scholar] [CrossRef]
- Chen, L.; Gibbons, D.L.; Goswami, S.; Cortez, M.A.; Ahn, Y.-H.; Byers, L.A.; Zhang, X.; Yi, X.; Dwyer, D.; Lin, W.; et al. Metastasis is regulated via microRNA-200/ZEB1 axis control of tumour cell PD-L1 expression and intratumoral immunosuppression. Nat. Commun. 2014, 5, 5241. [Google Scholar] [CrossRef]
- Sha, H.-H.; Wang, D.-D.; Chen, D.; Liu, S.-W.; Wang, Z.; Yan, D.-L.; Dong, S.-C.; Feng, J.-F. MiR-138: A promising therapeutic target for cancer. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef]
- Yeh, M.; Oh, C.S.; Yoo, J.Y.; Kaur, B.; Lee, T.J. Pivotal role of microRNA-138 in human cancers. Am. J. Cancer Res 2019, 9, 1118–1126. [Google Scholar] [PubMed]
- Song, N.; Li, P.; Song, P.; Li, Y.; Zhou, S.; Su, Q.; Li, X.; Yu, Y.; Li, P.; Feng, M.; et al. MicroRNA-138-5p Suppresses Non-small Cell Lung Cancer Cells by Targeting PD-L1/PD-1 to Regulate Tumor Microenvironment. Front. Cell Dev. Biol. 2020, 8, 540. [Google Scholar] [CrossRef] [PubMed]
- Pinho, J.D.; Silva, G.E.B.; Júnior, A.A.L.T.; Belfort, M.R.D.C.; Macedo, J.M.; Da Cunha, I.W.; Quintana, L.G.; Calixto, J.D.R.R.; Nogueira, L.R.; Coelho, R.W.P.; et al. MIR-107, MIR-223-3P and MIR-21-5P Reveals Potential Biomarkers in Penile Cancer. Asian Pac. J. Cancer Prev. 2020, 21, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Hartz, J.M.; Engelmann, D.; Fürst, K.; Marquardt, S.; Spitschak, A.; Goody, D.; Protzel, C.; Hakenberg, O.W.; Pützer, B.M. Integrated Loss of miR-1/miR-101/miR-204 Discriminates Metastatic from Nonmetastatic Penile Carcinomas and Can Predict Patient Outcome. J. Urol. 2016, 196, 570–578. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PATIENT CHARACTERISTICS | N | % | |
|---|---|---|---|
| HISTOLOGICAL SUBTYPE | usual | 18 | 51.4 |
| warty–basaloid | 6 | 17.1 | |
| basaloid | 8 | 22.9 | |
| warty | 1 | 2.9 | |
| clear cell | 1 | 2.9 | |
| verrucous | 1 | 2.9 | |
| PRIMARY TUMOR | pT1a | 8 | 22.9 |
| pT1b | 4 | 11.4 | |
| pT2 | 11 | 31.4 | |
| pT3 | 10 | 28.6 | |
| n/a | 2 | 5.7 | |
| REGIONAL LYMPH NODES | N0 | 17 | 48.6 |
| N1 | 6 | 17.1 | |
| N2 | 1 | 2.9 | |
| N3 | 9 | 25.7 | |
| n/a | 2 | 5.7 | |
| HISTOLOGIC GRADE | G1 | 2 | 5.7 |
| G2 | 16 | 45.7 | |
| G3 | 15 | 42.9 | |
| n/a | 2 | 5.7 | |
| METASTASIS STATUS | Non-metastatic | 18 | 51.4 |
| Metastatic | 17 | 48.6 | |
| HPV STATUS | HPV+ | 17 | 48.6 |
| HPV- | 18 | 51.4 |
| miRNAs | Fold Change in Microarray, p-Value | Fold Change in RT-qPCR, p-Value | HPV Status | Comparison | Histological Subtype |
|---|---|---|---|---|---|
| miR-99a-5p | −8.0 (p = 0.037) | −3.5 (p = 0.017) | HPV-negative | Tumor vs. Normal | Usual |
| miR-137 | −3.7 (p = 0.004) | −8.54 (p = 0.004) | HPV-negative | Metastatic vs. Non-metastatic | |
| miR-328-3p | −2.7 (p = 0.007) | −1.98 (p = 0.032) | HPV-negative | Metastatic vs. Non-metastatic | |
| miR-181d-5p | +3.4 (p = 0.004) | +2.01 (p = 0.002) | HPV-positive | Tumor vs. Normal | Basaloid, Warty, and Warty–basaloid |
| miR-211-5p | −16.9 (p = 0.046) | Not defined | HPV-positive | Tumor vs. Normal |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayoubian, H.; Heinzelmann, J.; Hölters, S.; Khalmurzaev, O.; Pryalukhin, A.; Loertzer, P.; Heinzelbecker, J.; Lohse, S.; Geppert, C.; Loertzer, H.; et al. miRNA Expression Characterizes Histological Subtypes and Metastasis in Penile Squamous Cell Carcinoma. Cancers 2021, 13, 1480. https://doi.org/10.3390/cancers13061480
Ayoubian H, Heinzelmann J, Hölters S, Khalmurzaev O, Pryalukhin A, Loertzer P, Heinzelbecker J, Lohse S, Geppert C, Loertzer H, et al. miRNA Expression Characterizes Histological Subtypes and Metastasis in Penile Squamous Cell Carcinoma. Cancers. 2021; 13(6):1480. https://doi.org/10.3390/cancers13061480
Chicago/Turabian StyleAyoubian, Hiresh, Joana Heinzelmann, Sebastian Hölters, Oybek Khalmurzaev, Alexey Pryalukhin, Philine Loertzer, Julia Heinzelbecker, Stefan Lohse, Carol Geppert, Hagen Loertzer, and et al. 2021. "miRNA Expression Characterizes Histological Subtypes and Metastasis in Penile Squamous Cell Carcinoma" Cancers 13, no. 6: 1480. https://doi.org/10.3390/cancers13061480
APA StyleAyoubian, H., Heinzelmann, J., Hölters, S., Khalmurzaev, O., Pryalukhin, A., Loertzer, P., Heinzelbecker, J., Lohse, S., Geppert, C., Loertzer, H., Wunderlich, H., Bohle, R. M., Stöckle, M., Matveev, V. B., Hartmann, A., & Junker, K. (2021). miRNA Expression Characterizes Histological Subtypes and Metastasis in Penile Squamous Cell Carcinoma. Cancers, 13(6), 1480. https://doi.org/10.3390/cancers13061480