
Cross-Reactivity and Functionality of Approved Human Immune Checkpoint Blockers in Dogs

 , , , , , , and
, , , , , , and
Abstract
Simple Summary
Abstract
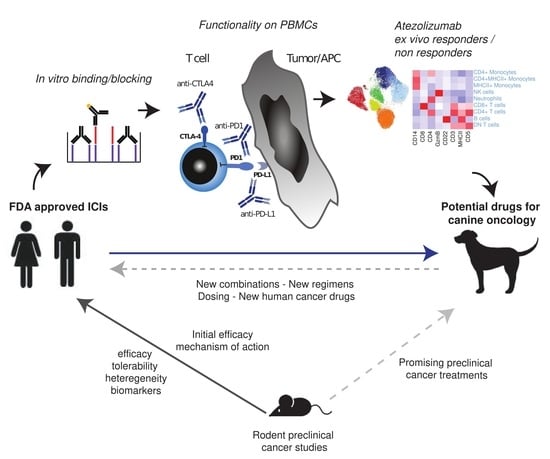
1. Introduction
2. Materials and Methods
2.1. Human Immune Checkpoint Inhibitor (ICI) Antibodies
2.2. Canine and Human Cell Line Culture
2.3. Canine Soluble Recombinant Proteins
2.4. Sequence Alignments of Canine and Human CTLA-4, PD-1 and PD-L1
2.5. Dog PBMC Isolation, Thawing, Stimulation and Suppression
2.6. ELISA and Binding Assays
2.7. Blocking of cPD-1 and cPD-L1 Binding Assay
2.8. Flow Cytometry
2.9. Statistics and Curve Fitting
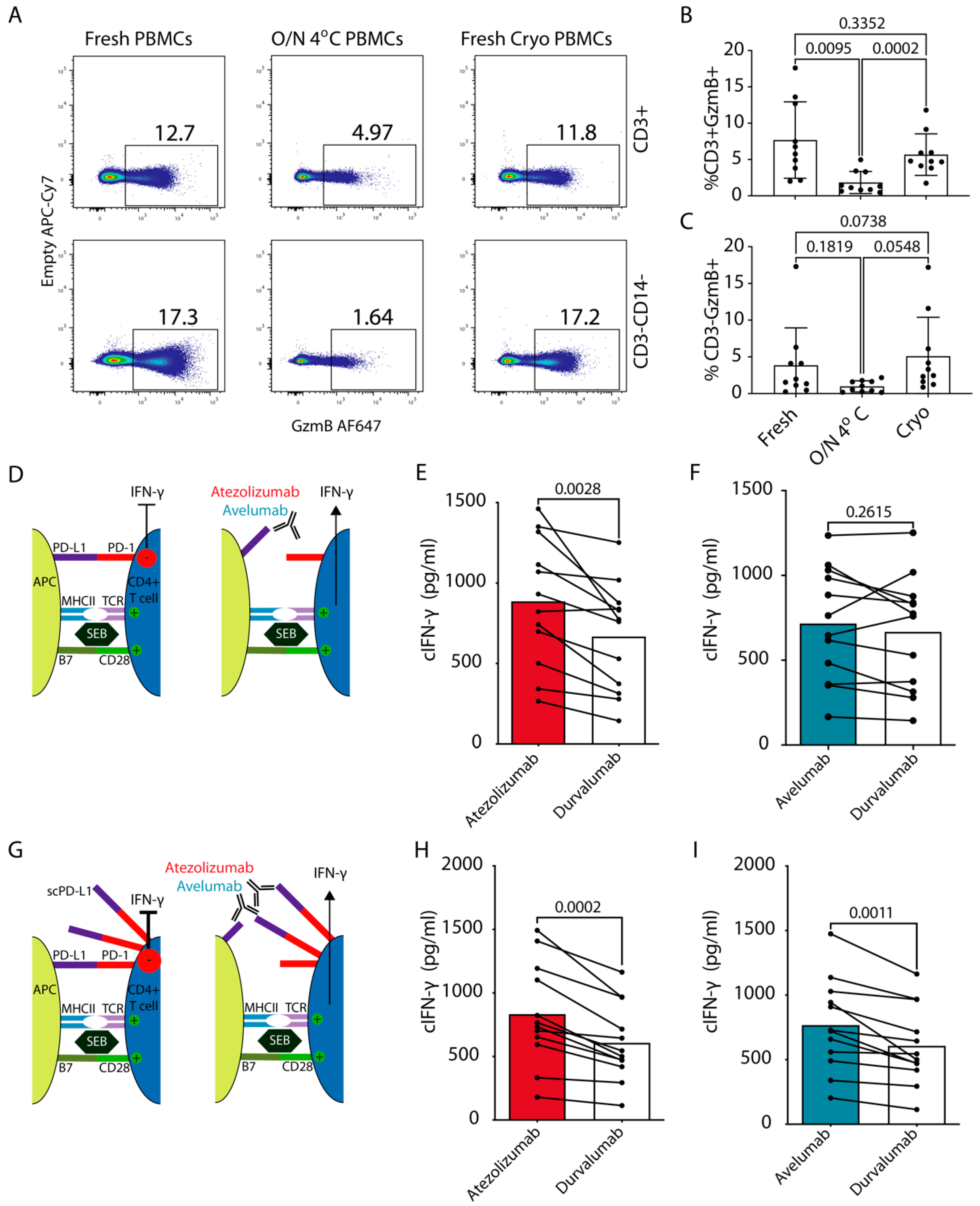
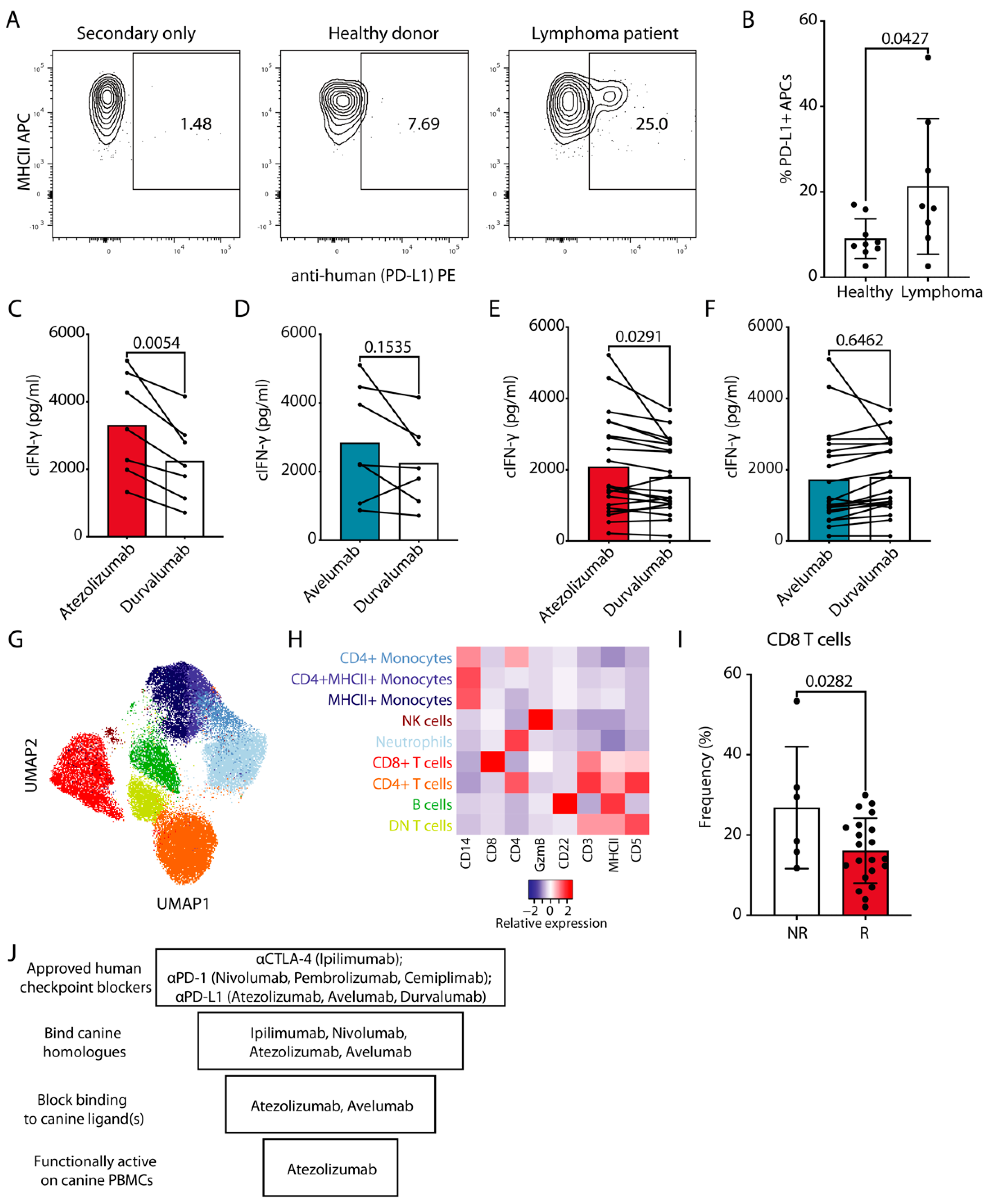
3. Results
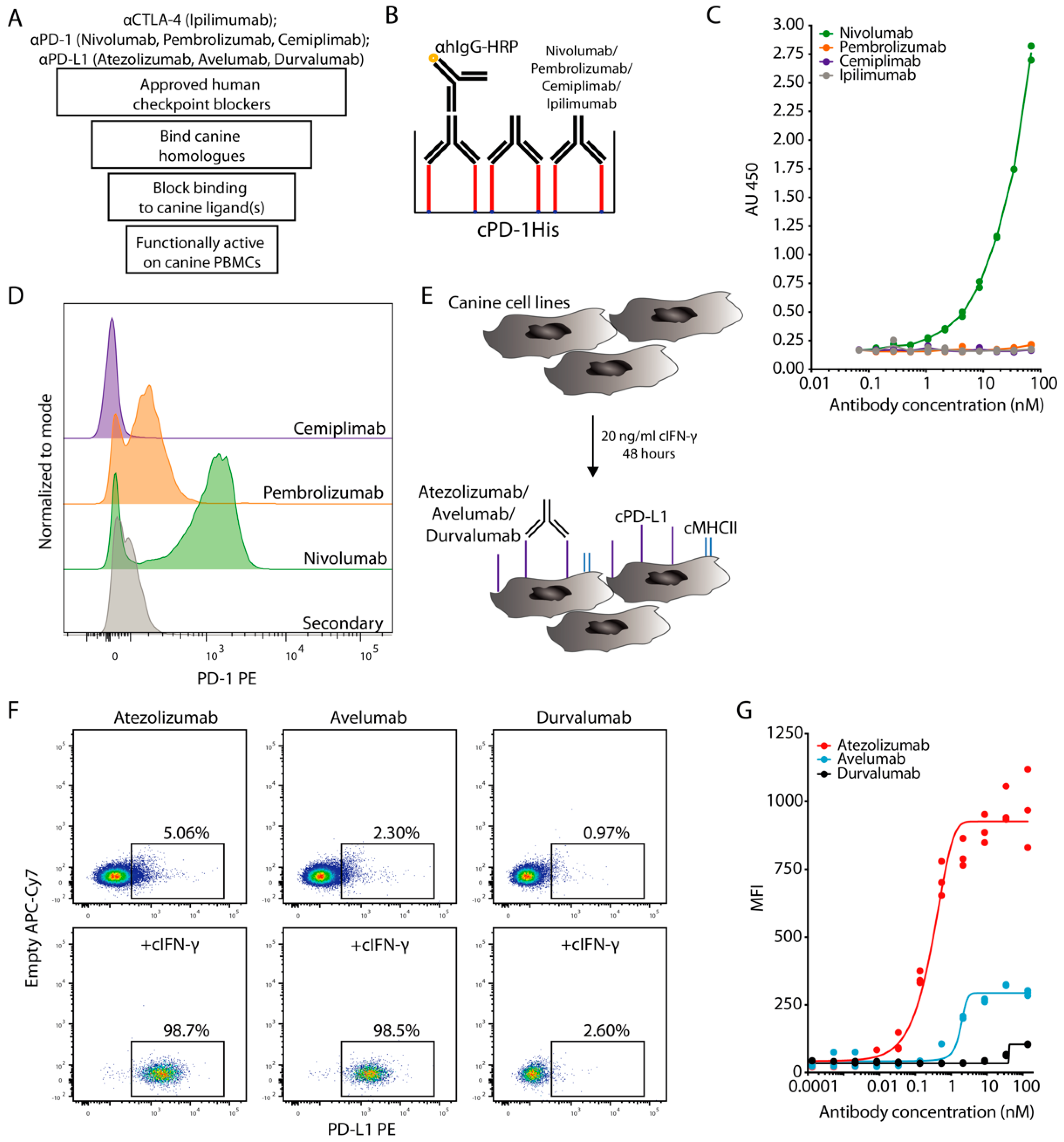
3.1. Evaluation of ICIs Binding to Canine PD-1
3.2. Evaluation of ICIs Binding to Canine PD-L1
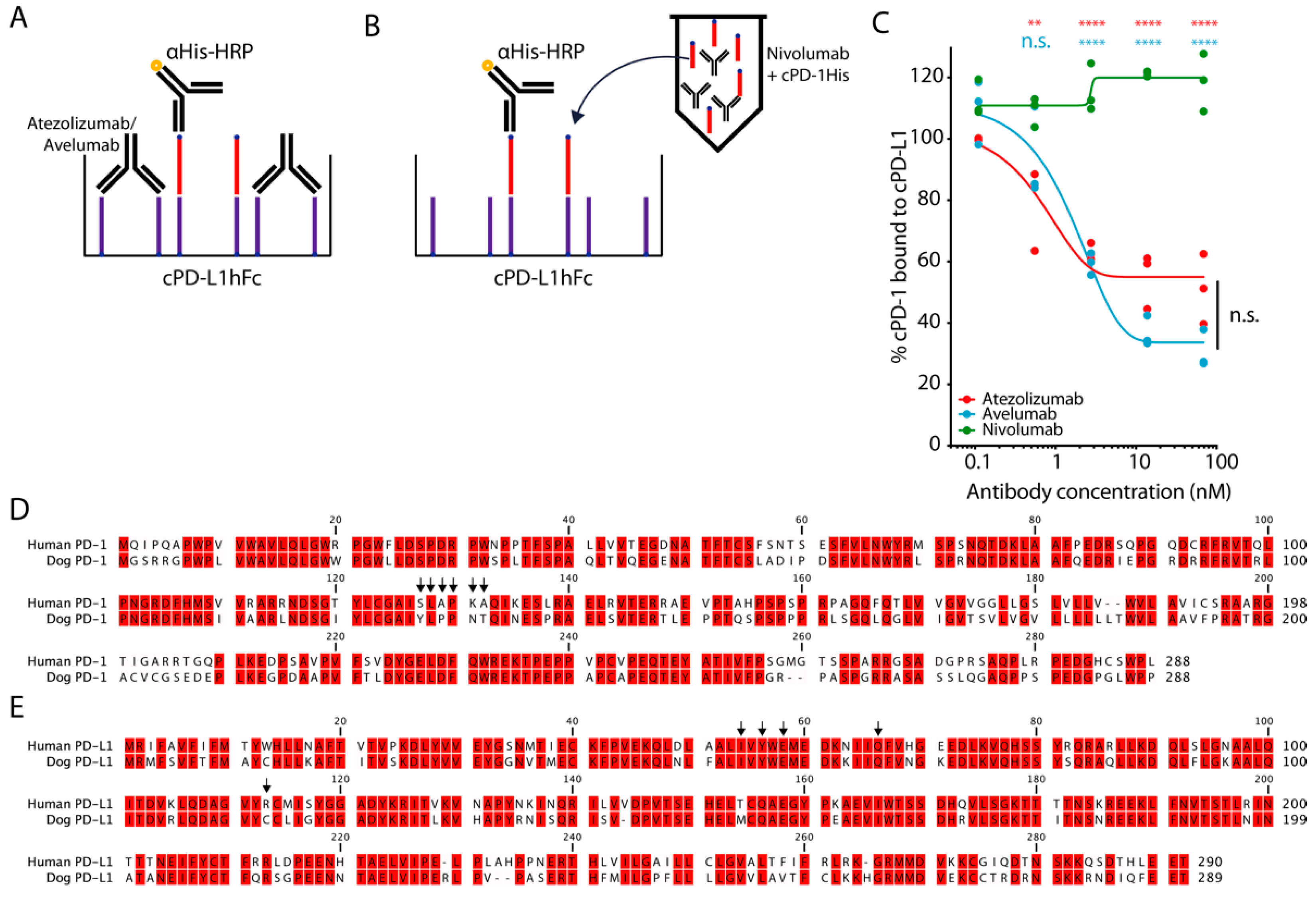
3.3. Atezolizumab and Avelumab Block the Binding of Canine PD-1 and PD-L1
3.4. Functional Effects of Blockade of cPD-1/PD-L1 Interaction
3.5. In Vitro Response of Cancer Patient cPBMCs to Atezolizumab-Mediated PD-L1 Blockade
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murciano-Goroff, Y.R.; Warner, A.B.; Wolchok, J.D. The future of cancer immunotherapy: Microenvironment-targeting combinations. Cell Res. 2020, 30, 507–519. [Google Scholar] [CrossRef]
- Mazzarella, L.; Duso, B.A.; Trapani, D.; Belli, C.; D’Amico, P.; Ferraro, E.; Viale, G.; Curigliano, G. The evolving landscape of ‘next-generation’ immune checkpoint inhibitors: A review. Eur. J. Cancer 2019, 117, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Shalabi, A.; Hubbard-Lucey, V. Comprehensive analysis of the clinical immuno-oncology landscape. Ann. Oncol. 2018, 29, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Withers, S.S.; Modiano, J.F.; Kent, M.S.; Chen, M.; Luna, J.I.; Culp, W.T.N.; Sparger, E.E.; Rebhun, R.B.; Monjazeb, A.M.; et al. Canine cancer immunotherapy studies: Linking mouse and human. J. Immunother. Cancer 2016, 4, 97. [Google Scholar] [CrossRef]
- Leblanc, A.K.; Mazcko, C.N. Improving human cancer therapy through the evaluation of pet dogs. Nat. Rev. Cancer 2020, 20, 727–742. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Withers, S.S.; Chang, H.; Spanier, J.A.; De La Trinidad, V.L.; Panesar, H.; Fife, B.T.; Sciammas, R.; Sparger, E.E.; Moore, P.F.; et al. Development of canine PD-1/PD-L1 specific monoclonal antibodies and amplification of canine T cell function. PLoS ONE 2020, 15, e0235518. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, N.; Konnai, S.; Ikebuchi, R.; Okagawa, T.; Adachi, M.; Takagi, S.; Kagawa, Y.; Nakajima, C.; Suzuki, Y.; Murata, S.; et al. Expression of PD-L1 on Canine Tumor Cells and Enhancement of IFN-γ Production from Tumor-Infiltrating Cells by PD-L1 Blockade. PLoS ONE 2014, 9, e98415. [Google Scholar] [CrossRef]
- Coy, J.; Caldwell, A.; Chow, L.; Guth, A.; Dow, S. PD-1 expression by canine T cells and functional effects of PD-1 blockade. Veter. Comp. Oncol. 2017, 15, 1487–1502. [Google Scholar] [CrossRef] [PubMed]
- Hartley, G.; Faulhaber, E.; Caldwell, A.; Coy, J.; Kurihara, J.; Guth, A.; Regan, D.; Dow, S. Immune regulation of canine tumour and macrophage PD-L1 expression. Veter. Comp. Oncol. 2017, 15, 534–549. [Google Scholar] [CrossRef] [PubMed]
- Hartley, A.N.; Cooley, G.; Gwyn, S.; Orozco, M.M.; Tarleton, R.L. Frequency of IFNγ-producing T cells correlates with seroreactivity and activated T cells during canine Trypanosoma cruzi infection. Veter. Res. 2014, 45, 6. [Google Scholar] [CrossRef] [PubMed]
- Hartley, G.; Elmslie, R.; Dow, S.; Guth, A. Checkpoint molecule expression by B and T cell lymphomas in dogs. Veter. Comp. Oncol. 2018, 16, 352–360. [Google Scholar] [CrossRef]
- Shosu, K.; Sakurai, M.; Inoue, K.; Nakagawa, T.; Sakai, H.; Morimoto, M.; Okuda, M.; Noguchi, S.; Mizuno, T. Programmed Cell Death Ligand 1 Expression in Canine Cancer. Vivo 2016, 30, 195–204. [Google Scholar]
- Tagawa, M.; Kurashima, C.; Takagi, S.; Maekawa, N.; Konnai, S.; Shimbo, G.; Matsumoto, K.; Inokuma, H.; Kawamoto, K.; Miyahara, K. Evaluation of costimulatory molecules in dogs with B cell high grade lymphoma. PLoS ONE 2018, 13, e0201222. [Google Scholar] [CrossRef]
- Guscetti, F.; Nassiri, S.; Beebe, E.; Brandao, I.R.; Graf, R.; Markkanen, E. Molecular homology between canine spontaneous oral squamous cell carcinomas and human head-and-neck squamous cell carcinomas reveals disease drivers and therapeutic vulnerabilities. Neoplasia 2020, 22, 778–788. [Google Scholar] [CrossRef]
- Igase, M.; Nemoto, Y.; Itamoto, K.; Tani, K.; Nakaichi, M.; Sakurai, M.; Sakai, Y.; Noguchi, S.; Kato, M.; Tsukui, T.; et al. A pilot clinical study of the therapeutic antibody against canine PD-1 for advanced spontaneous cancers in dogs. Sci. Rep. 2020, 10, 18311. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, N.; Konnai, S.; Takagi, S.; Kagawa, Y.; Okagawa, T.; Nishimori, A.; Ikebuchi, R.; Izumi, Y.; Deguchi, T.; Nakajima, C.; et al. A canine chimeric monoclonal antibody targeting PD-L1 and its clinical efficacy in canine oral malignant melanoma or undifferentiated sarcoma. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Klingemann, H. Immunotherapy for Dogs: Running Behind Humans. Front. Immunol. 2018, 9, 133. [Google Scholar] [CrossRef] [PubMed]
- Tomar, D.S.; Kumar, S.; Singh, S.K.; Goswami, S.; Li, L. Molecular basis of high viscosity in concentrated antibody solutions: Strategies for high concentration drug product development. mAbs 2016, 8, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Jain, T.; Sun, T.; Durand, S.; Hall, A.; Houston, N.R.; Nett, J.H.; Sharkey, B.; Bobrowicz, B.; Caffry, I.; Yu, Y.; et al. Biophysical properties of the clinical-stage antibody landscape. Proc. Natl. Acad. Sci. USA 2017, 114, 944–949. [Google Scholar] [CrossRef]
- Dow, S. A Role for Dogs in Advancing Cancer Immunotherapy Research. Front. Immunol. 2020, 10, 2935. [Google Scholar] [CrossRef]
- Wang, C.; Thudium, K.B.; Han, M.; Wang, X.-T.; Huang, H.; Feingersh, D.; Garcia, C.; Wu, Y.; Kuhne, M.; Srinivasan, M.; et al. In Vitro Characterization of the Anti-PD-1 Antibody Nivolumab, BMS-936558, and In Vivo Toxicology in Non-Human Primates. Cancer Immunol. Res. 2014, 2, 846–856. [Google Scholar] [CrossRef]
- Selby, M.J.; Engelhardt, J.J.; Johnston, R.J.; Lu, L.-S.; Han, M.; Thudium, K.; Yao, D.; Quigley, M.; Valle, J.; Wang, C.; et al. Preclinical Development of Ipilimumab and Nivolumab Combination Immunotherapy: Mouse Tumor Models, In Vitro Functional Studies, and Cynomolgus Macaque Toxicology. PLoS ONE 2016, 11, e0161779. [Google Scholar] [CrossRef]
- Suter, M.M.; Pantano, D.M.; Flanders, J.A.; Augustin-Voss, H.G.; Dougherty, E.P.; Varvayanis, M. Comparison of Growth and Differentiation of Normal and Neoplastic Canine Keratinocyte Cultures. Veter Pathol. 1991, 28, 131–138. [Google Scholar] [CrossRef]
- Graubner, F.R.; Reichler, I.M.; A Rahman, N.; Payan-Carreira, R.; Boos, A.; Kowalewski, M.P. Decidualization of the canine uterus: From early until late gestational in vivo morphological observations, and functional characterization of immortalized canine uterine stromal cell lines. Reprod. Domest. Anim. 2017, 52, 137–147. [Google Scholar] [CrossRef]
- Pantelyushin, S.; Ranninger, E.; Bettschart-Wolfensberger, R.; Berg, J.V. OMIP-065: Dog Immunophenotyping and T-Cell Activity Evaluation with a 14-Color Panel. Cytom. Part A 2020, 97, 1024–1027. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Diaz, A.; Shin, D.S.; Moreno, B.H.; Saco, J.; Escuin-Ordinas, H.; Rodriguez, G.A.; Zaretsky, J.M.; Sun, L.; Hugo, W.; Wang, X.; et al. Interferon Receptor Signaling Pathways Regulating PD-L1 and PD-L2 Expression. Cell Rep. 2017, 19, 1189–1201. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.T.; Lee, S.H.; Heo, Y.-S. Molecular Interactions of Antibody Drugs Targeting PD-1, PD-L1, and CTLA-4 in Immuno-Oncology. Molecules 2019, 24, 1190. [Google Scholar] [CrossRef]
- Pinchuk, I.V.; Beswick, E.J.; Reyes, V.E. Staphylococcal Enterotoxins. Toxins 2010, 2, 2177–2197. [Google Scholar] [CrossRef]
- Nixon, A.B.; Schalper, K.A.; Jacobs, I.; Potluri, S.; Wang, I.-M.; Fleener, C. Peripheral immune-based biomarkers in cancer immunotherapy: Can we realize their predictive potential? J. Immunother. Cancer 2019, 7, 325. [Google Scholar] [CrossRef] [PubMed]
- Mayoux, M.; Roller, A.; Pulko, V.; Sammicheli, S.; Chen, S.; Sum, E.; Jost, C.; Fransen, M.F.; Buser, R.B.; Kowanetz, M.; et al. Dendritic cells dictate responses to PD-L1 blockade cancer immunotherapy. Sci. Transl. Med. 2020, 12, eaav7431. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.A.; Wu, D.-C.; Cheung, J.; Navarro, A.; Xiong, H.; Cubas, R.; Totpal, K.; Chiu, H.; Wu, Y.; Comps-Agrar, L.; et al. PD-L1 expression by dendritic cells is a key regulator of T-cell immunity in cancer. Nat. Rev. Cancer 2020, 1, 681–691. [Google Scholar] [CrossRef]
- Akinleye, A.; Rasool, Z. Immune checkpoint inhibitors of PD-L1 as cancer therapeutics. J. Hematol. Oncol. 2019, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, L.M.; McCandless, E.E.; Dunham, S.; Dunkle, B.; Zhu, Y.; Shelly, J.; Lightle, S.; Gonzales, A.; Bainbridge, G. Comparative functional characterization of canine IgG subclasses. Veter Immunol. Immunopathol. 2014, 157, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Foltz, J.A.; Somanchi, S.S.; Yang, Y.; Aquino-Lopez, A.; Bishop, E.E.; Lee, D.A. NCR1 Expression Identifies Canine Natural Killer Cell Subsets with Phenotypic Similarity to Human Natural Killer Cells. Front. Immunol. 2016, 7, 521. [Google Scholar] [CrossRef]
- Lee, S.-H.; Shin, D.-J.; Kim, Y.; Kim, C.-J.; Lee, J.-J.; Yoon, M.S.; Uong, T.N.T.; Yu, D.; Jung, J.-Y.; Cho, D.; et al. Comparison of Phenotypic and Functional Characteristics Between Canine Non-B, Non-T Natural Killer Lymphocytes and CD3+CD5dimCD21− Cytotoxic Large Granular Lymphocytes. Front. Immunol. 2018, 9, 841. [Google Scholar] [CrossRef] [PubMed]
- Tabrizi, M.; Neupane, D.; Elie, S.E.; Shankaran, H.; Juan, V.; Zhang, S.; Hseih, S.; Fayadat-Dilman, L.; Zhang, D.; Song, Y.; et al. Pharmacokinetic Properties of Humanized IgG1 and IgG4 Antibodies in Preclinical Species: Translational Evaluation. AAPS J. 2019, 21, 39. [Google Scholar] [CrossRef]
- Maeda, S.; Murakami, K.; Inoue, A.; Yonezawa, T.; Matsuki, N. CCR4 Blockade Depletes Regulatory T Cells and Prolongs Survival in a Canine Model of Bladder Cancer. Cancer Immunol. Res. 2019, 7, 1175–1187. [Google Scholar] [CrossRef]
- Krieg, C.; Nowicka, M.; Guglietta, S.; Schindler, S.; Hartmann, F.J.; Weber, L.M.; Dummer, R.; Robinson, M.D.; Levesque, M.P.; Becher, B. Author Correction: High-dimensional single-cell analysis predicts response to anti-PD-1 immunotherapy. Nat. Med. 2018, 24, 1773–1775. [Google Scholar] [CrossRef] [PubMed]
- Hegde, P.S.; Chen, D.S. Top 10 Challenges in Cancer Immunotherapy. Immunity 2020, 52, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Kobold, S.; Pantelyushin, S.; Rataj, F.; Berg, J.V. Rationale for Combining Bispecific T Cell Activating Antibodies With Checkpoint Blockade for Cancer Therapy. Front. Oncol. 2018, 8, 285. [Google Scholar] [CrossRef] [PubMed]
- Boshuizen, J.; Peeper, D.S. Rational Cancer Treatment Combinations: An Urgent Clinical Need. Mol. Cell 2020, 78, 1002–1018. [Google Scholar] [CrossRef]
- Algazi, A.P.; Twitty, C.G.; Tsai, K.K.; Le, M.; Pierce, R.; Browning, E.; Hermiz, R.; Canton, D.A.; Bannavong, D.; Oglesby, A.; et al. Phase II Trial of IL-12 Plasmid Transfection and PD-1 Blockade in Immunologically Quiescent Melanoma. Clin. Cancer Res. 2020, 26, 2827–2837. [Google Scholar] [CrossRef] [PubMed]
- Tugues, S.; Burkhard, S.H.; Ohs, I.; Vrohlings, M.; Nussbaum, K.; vom Berg, J.; Kulig, P.; Becher, B. New insights into IL-12-mediated tumor suppression. Cell Death Differ. 2015, 22, 237–246. [Google Scholar] [CrossRef]
- Pavlin, D.; Cemazar, M.; Sersa, G.; Tozon, N. IL-12 based gene therapy in veterinary medicine. J. Transl. Med. 2012, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Paoloni, M.; Mazcko, C.; Selting, K.; Lana, S.; Barber, L.; Phillips, J.; Skorupski, K.; Vail, D.; Wilson, H.; Biller, B.; et al. Defining the Pharmacodynamic Profile and Therapeutic Index of NHS-IL12 Immunocytokine in Dogs with Malignant Melanoma. PLoS ONE 2015, 10, e0129954. [Google Scholar] [CrossRef]
- Omar, N.B.; Bentley, R.T.; Crossman, D.K.; Foote, J.B.; Koehler, J.W.; Markert, J.M.; Platt, S.R.; Rissi, D.R.; Shores, A.; Sorjonen, D.; et al. Safety and interim survival data after intracranial administration of M032, a genetically engineered oncolytic HSV-1 expressing IL-12, in pet dogs with sporadic gliomas. Neurosurg. Focus 2021, 50, E5. [Google Scholar] [CrossRef]
- Fallon, J.K.; Vandeveer, A.J.; Schlom, J.; Greiner, J.W. Enhanced antitumor effects by combining an IL-12/anti-DNA fusion protein with avelumab, an anti-PD-L1 antibody. Oncotarget 2017, 8, 20558–20571. [Google Scholar] [CrossRef]
- Green, M.R.; Monti, S.; Rodig, S.J.; Juszczynski, P.; Currie, T.; O’Donnell, E.; Chapuy, B.; Takeyama, K.; Neuberg, D.; Golub, T.R.; et al. Integrative analysis reveals selective 9p24.1 amplification, increased PD-1 ligand expression, and further induction via JAK2 in nodular sclerosing Hodgkin lymphoma and primary mediastinal large B-cell lymphoma. Blood 2010, 116, 3268–3277. [Google Scholar] [CrossRef]
- Ansell, S.M.; Lesokhin, A.M.; Borrello, I.; Halwani, A.; Scott, E.C.; Gutierrez, M.; Schuster, S.J.; Millenson, M.M.; Cattry, D.; Freeman, G.J.; et al. PD-1 Blockade with Nivolumab in Relapsed or Refractory Hodgkin’s Lymphoma. N. Engl. J. Med. 2015, 372, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Armand, P.; Chen, Y.-B.; Redd, R.A.; Joyce, R.M.; Bsat, J.; Jeter, E.; Merryman, R.W.; Coleman, K.C.; Dahi, P.B.; Nieto, Y.; et al. PD-1 blockade with pembrolizumab for classical Hodgkin lymphoma after autologous stem cell transplantation. Blood 2019, 134, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Rauch, D.A.; Conlon, K.C.; Janakiram, M.; Brammer, J.E.; Harding, J.C.; Ye, B.H.; Zang, X.; Ren, X.; Olson, S.; Cheng, X.; et al. Rapid progression of adult T-cell leukemia/lymphoma as tumor-infiltrating Tregs after PD-1 blockade. Blood 2019, 134, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Ratner, L.; Waldmann, T.A.; Janakiram, M.; Brammer, J.E. Rapid Progression of Adult T-Cell Leukemia–Lymphoma after PD-1 Inhibitor Therapy. N. Engl. J. Med. 2018, 378, 1947–1948. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Specificity | Fluorochrome | Clone | Dilution | Staining | Manufacturer | Product | Target | Host |
|---|---|---|---|---|---|---|---|---|---|
| 1 | CD45 | eFluor 450 | YKIX716.13 | 1:50 | Surface | Thermo Fisher | 48-5450-41 | Dog | Rat |
| 2 | Dead cells | Aqua | N/A | 1:500 | Surface | Biolegend | 423102 | N/A | N/A |
| 3 | CD25 | SB600 | P4A10 | 1:50 | Surface | Thermo Fisher | 63-0250-41 | Dog | Mouse |
| 4 | CD4 | SB645 | YKIX302.9 | 1:10 | Surface | Thermo Fisher | 64-5040-42 | Dog | Rat |
| 5 | CD8 | SB702 | YCATE55.9 | 1:25 | Surface | Thermo Fisher | 67-5080-42 | Dog | Rat |
| 6 | CD14 | BV785 | M5E2 | 1:100 | Surface | Biolegend | 301840 | Human | Mouse |
| 7 | CD3 | FITC | CA17.2A12 | 1:10 | Surface | BioRad | MCA1774F | Dog | Mouse |
| 8 | Eomes | PerCP-eFluor710 | WD1928 | 1:100 | IC | Thermo Fisher | 46-4877-42 | Human | Mouse |
| 9 | CD22 | PE | RFB-4 | 1:50 | Surface | Thermo Fisher | MHCD2204 | Human | Mouse |
| 10 | Granzyme B | PE-CF594 | GB11 | 1:1600 | IC | BD | 562462 | Human | Mouse |
| 11 | Granzyme B | Alexa Fluor 647 | GB11 | 1:800 | IC | BD | 560212 | Human | Mouse |
| 12 | FoxP3 | PE-Cy7 | FJK-16s | 1:800 | IC | Thermo Fisher | 25-5773-80 | Human | Rat |
| 13 | MHCII | APC | YKIX334.2 | 1:25 | Surface | Thermo Fisher | 17-5909-42 | Dog | Rat |
| 14 | Ki67 | Alexa Fluor 700 | SolA15 | 1:6400 | IC | Thermo Fisher | 56-5698-82 | Rat | Rat |
| 15 | CD5 | APC-eFluor780 | YKIX322.3 | 1:50 | Surface | Thermo Fisher | 47-5050-42 | Dog | Rat |
| 16 | human Fc | PE | HP6017 | 1:200 | Secondary | Biolegend | 409304 | Human | Mouse |
| # | Sex/Castrated | Age | Breed | R/NR | Atezolizumab/Durvalumab Response (%) | Atezolizumab/ Durvalumab Response with scPD-L1 (%) |
|---|---|---|---|---|---|---|
| 1 | Female | 5 | Mixed Breed | NR | −1.90 | 21.01 |
| 2 | Female spayed | 3 | Berger Blanc Suisse | R | 5.08 | 23.23 |
| 3 | Female | 7 | Barsoi | R | 22.64 | 54.51 |
| 4 | Female | 9 | Flat Coated Retriever | R | 88.70 | 64.77 |
| 5 | Female spayed | 6 | Mixed breed | R | 59.71 | 37.99 |
| 6 | Female | 7 | Continental Bulldog | R | 7.89 | 54.07 |
| 7 | Female | 7 | Boxer | R | 50.31 | 41.58 |
| 8 | Male | 4 | Golden Retriever | R | 98.52 | 40.46 |
| 9 | Male castrated | 6 | Vizla | R | 34.16 | 56.02 |
| 10 | Female spayed | 4 | Labrador Retriever | R | 22.69 | 13.16 |
| 11 | Female | 8 | Boxer | R | 84.60 | 57.57 |
| 12 | Female | 1 | Golden Retriever | R | 32.06 | 9.09 |
| # | Sex/Castrated | Age | Breed | Diagnosis | Leukaemic | R/NR | Atezolizumab/ Durvalumab Response (%) |
|---|---|---|---|---|---|---|---|
| 1 | Female spayed | 10 | Czechoslovakian Wolfdog | Mast cell tumor | R | 19.09 | |
| 2 | Female | 9 | German Shepherd | Oral malignant melanoma | R | 17.96 | |
| 3 | Male castrated | 10 | Mixed Breed | Malignant B cell lymphoma | R | 51.77 | |
| 4 | Female spayed | 8 | Labrador Retriever | Mast cell tumor | NR | −24.41 | |
| 5 | Female spayed | 12 | Mixed Breed | Mast cell tumor, meningioma | R | 15.79 | |
| 6 | Female | 3 | German Boxer | Mast cell tumor | R | 28.01 | |
| 7 | Female spayed | 11 | Mixed Breed | Anal sac adenocarcinoma | NR | −29.37 | |
| 8 | Female spayed | 12 | French Bulldog | Mast cell tumor | NR | −9.66 | |
| 9 | Male castrated | 8 | Mixed Breed | Prostate carcinoma | R | 24.21 | |
| 10 | Male castrated | 14 | Mixed Breed | Malignant B cell lymphoma | R | 74.79 | |
| 11 | Male castrated | 8 | Golden Retriever | Mast cell tumor | R | 21.80 | |
| 12 | Male castrated | 11 | Beagle | Mast cell tumor | R | 7.99 | |
| 13 | Male | 6 | Bernese Mountain Dog | Malignant B cell lymphoma | R | 41.88 | |
| 14 | Female | 10 | Golden Retriever | Soft tissue sarcoma | R | 8.65 | |
| 15 | Female spayed | 9 | Mixed Breed | Mast cell tumor | R | 37.05 | |
| 16 | Female spayed | 12 | Mixed Breed | Hepatocellular carcinoma | NR | −12.41 | |
| 17 | MK | 11 | Italian Segugio | Malignant T cell lymphoma | x | R | 16.76 |
| 18 | Female spayed | 10 | Mixed Breed | Sinonasal carcinoma, pulmonary carcinoma | R | 52.09 | |
| 19 | Female spayed | 8 | Mixed Breed | Meningioma | R | 14.59 | |
| 20 | Male | 6 | Rhodesian Ridgeback | Mast cell tumor | R | 40.02 | |
| 21 | Male castrated | 10 | Golden Retriever | Oral malignant melanoma | NR | 2.89 | |
| 22 | Female spayed | 6 | Rhodesian Ridgeback | Malignant T cell lymphoma | R | 85.73 | |
| 23 | Male castrated | 5 | Bernese Mountain Dog | Malignant B cell lymphoma | R | 26.58 | |
| 24 | Male | 9 | Labrador Retriever | Mast cell tumor | R | 8.88 | |
| 25 | Female spayed | 13 | Hungarian Viszla | Sinonasal carcinoma, pulmonary carcinoma | NR | −15.54 | |
| 26 | Male | 12 | Golden Retriever | Malignant lymphoma | x | R | 86.67 |
| 27 | Male castrated | 8 | Golden Retriever | Mast cell tumor | R | 16.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pantelyushin, S.; Ranninger, E.; Guerrera, D.; Hutter, G.; Maake, C.; Markkanen, E.; Bettschart-Wolfensberger, R.; Rohrer Bley, C.; Läubli, H.; vom Berg, J. Cross-Reactivity and Functionality of Approved Human Immune Checkpoint Blockers in Dogs. Cancers 2021, 13, 785. https://doi.org/10.3390/cancers13040785
Pantelyushin S, Ranninger E, Guerrera D, Hutter G, Maake C, Markkanen E, Bettschart-Wolfensberger R, Rohrer Bley C, Läubli H, vom Berg J. Cross-Reactivity and Functionality of Approved Human Immune Checkpoint Blockers in Dogs. Cancers. 2021; 13(4):785. https://doi.org/10.3390/cancers13040785
Chicago/Turabian StylePantelyushin, Stanislav, Elisabeth Ranninger, Diego Guerrera, Gregor Hutter, Caroline Maake, Enni Markkanen, Regula Bettschart-Wolfensberger, Carla Rohrer Bley, Heinz Läubli, and Johannes vom Berg. 2021. "Cross-Reactivity and Functionality of Approved Human Immune Checkpoint Blockers in Dogs" Cancers 13, no. 4: 785. https://doi.org/10.3390/cancers13040785
APA StylePantelyushin, S., Ranninger, E., Guerrera, D., Hutter, G., Maake, C., Markkanen, E., Bettschart-Wolfensberger, R., Rohrer Bley, C., Läubli, H., & vom Berg, J. (2021). Cross-Reactivity and Functionality of Approved Human Immune Checkpoint Blockers in Dogs. Cancers, 13(4), 785. https://doi.org/10.3390/cancers13040785