
PD-L1 Is an Independent Prognostic Marker in Middle Eastern PTC and Its Expression Is Upregulated by BRAFV600E Mutation


Abstract
Simple Summary
Abstract
1. Introduction
2. Results
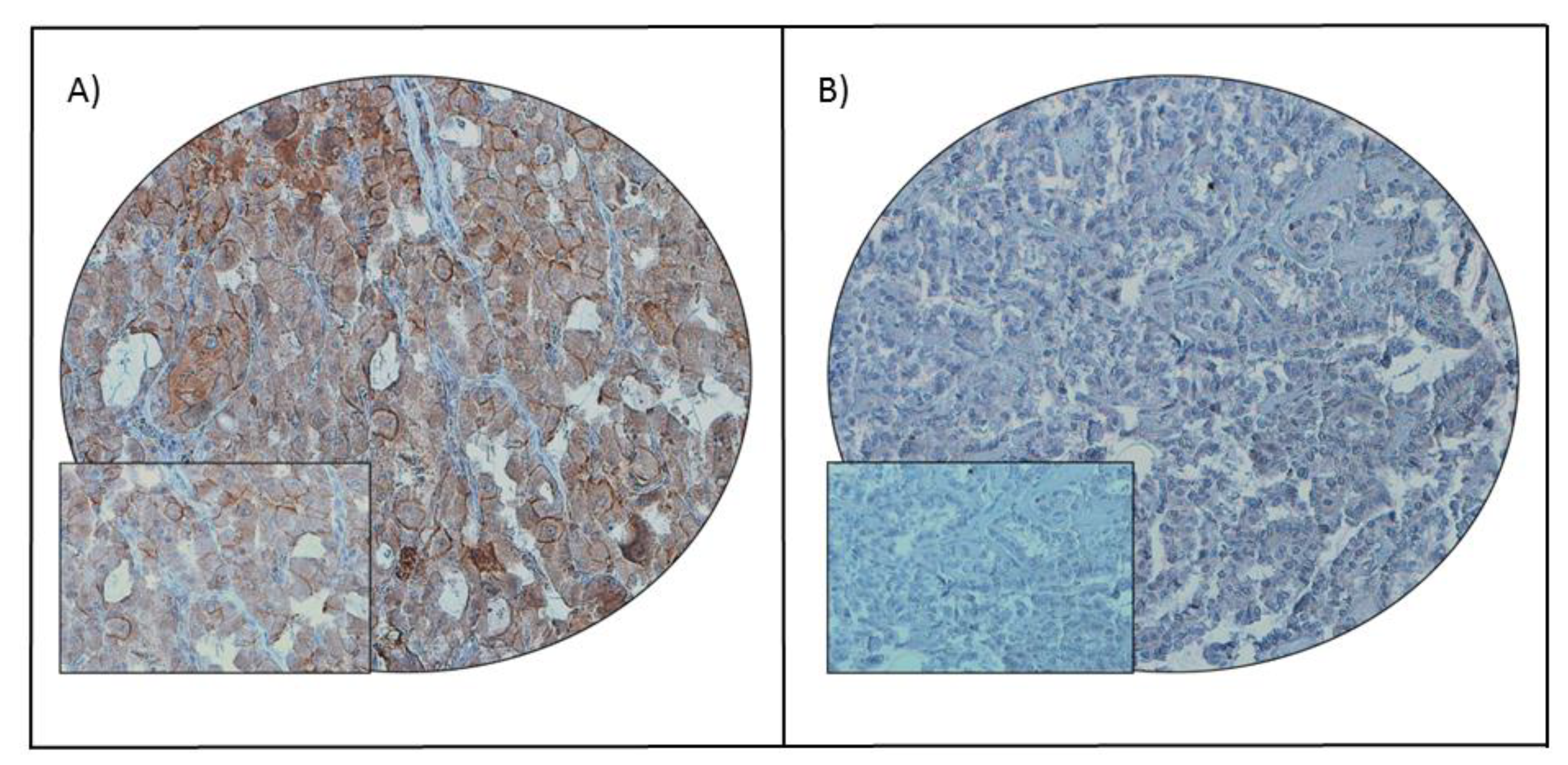
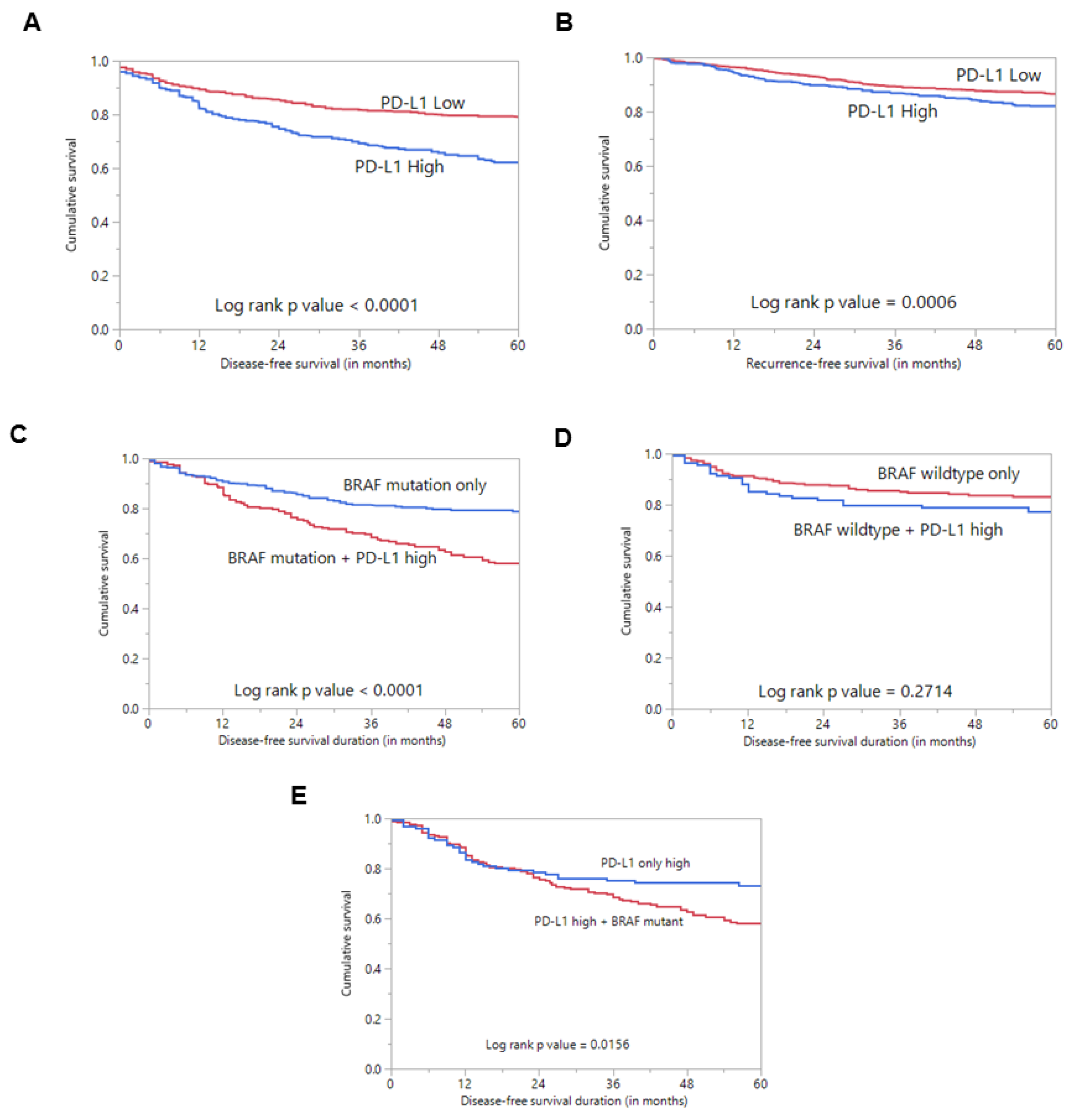
2.1. Programmed Cell Death Ligand 1 (PD-L1) Expression in Papillary Thyroid Carcinoma (PTC) and Its Clinico-Pathological Associations
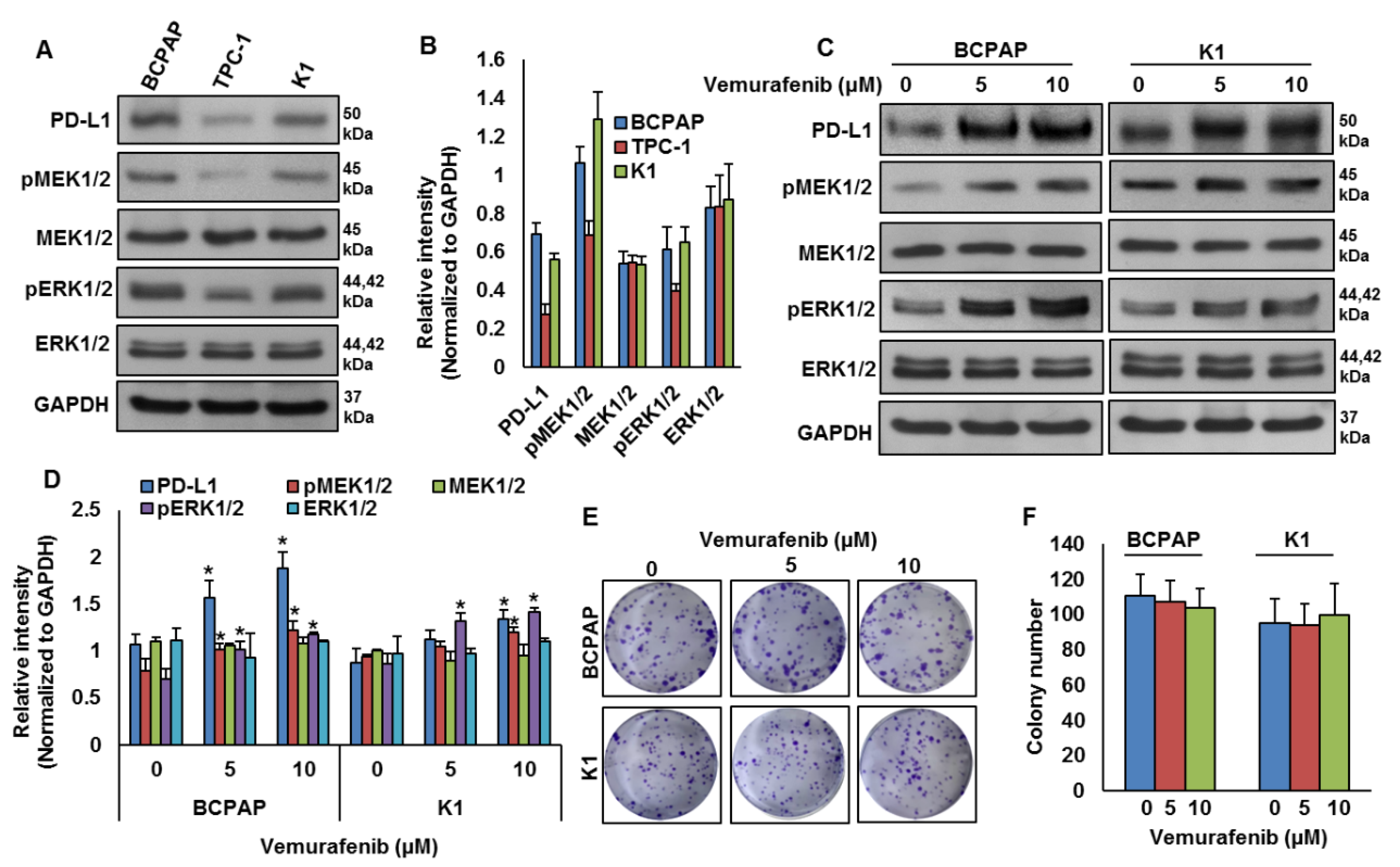
2.2. BRAF Mutation and Its Association with PD-L1 in PTC In Vitro
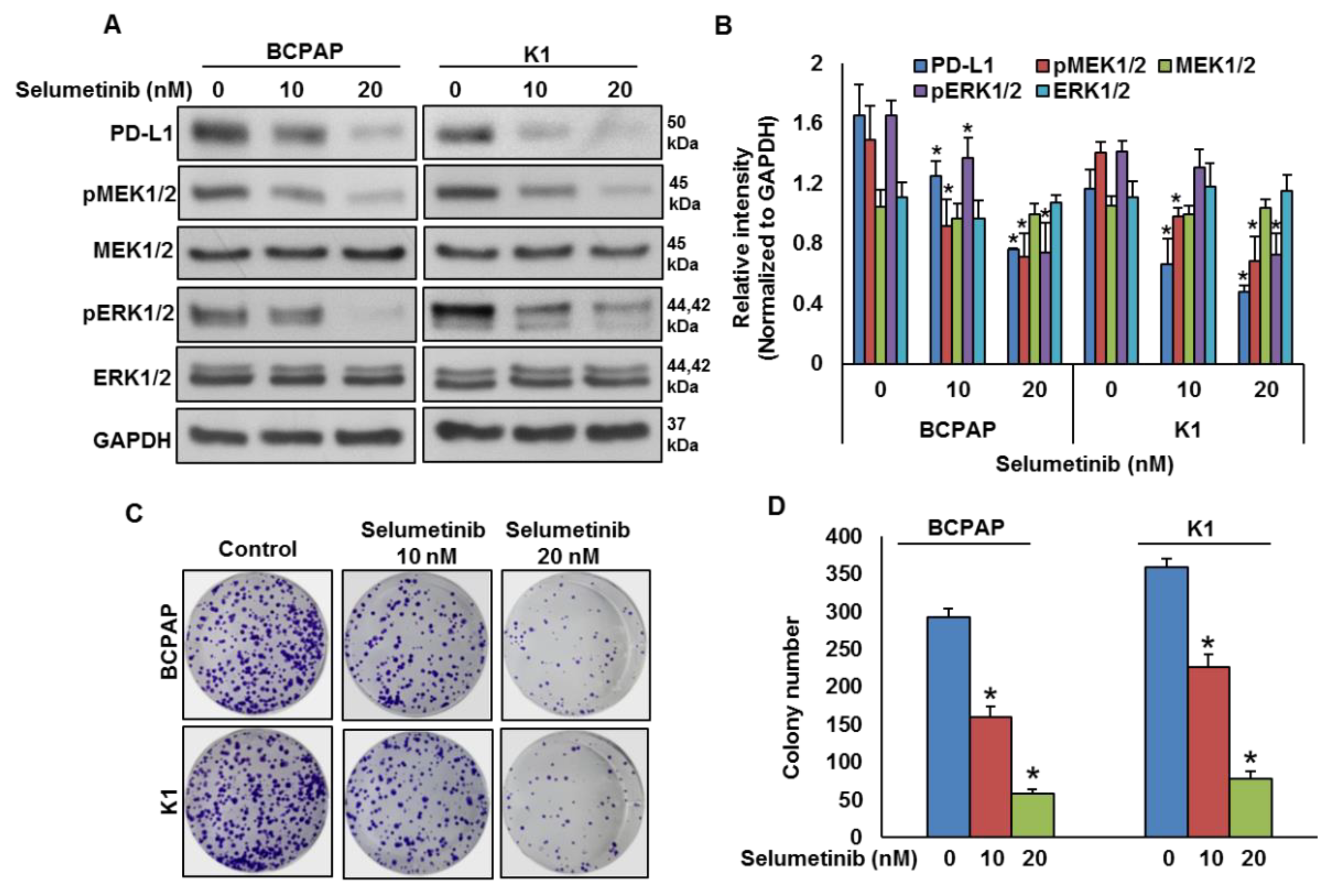
2.3. Mitogen-Activated Protein Kinase Kinase (MEK) Inhibition Decreases PD-L1 Expression
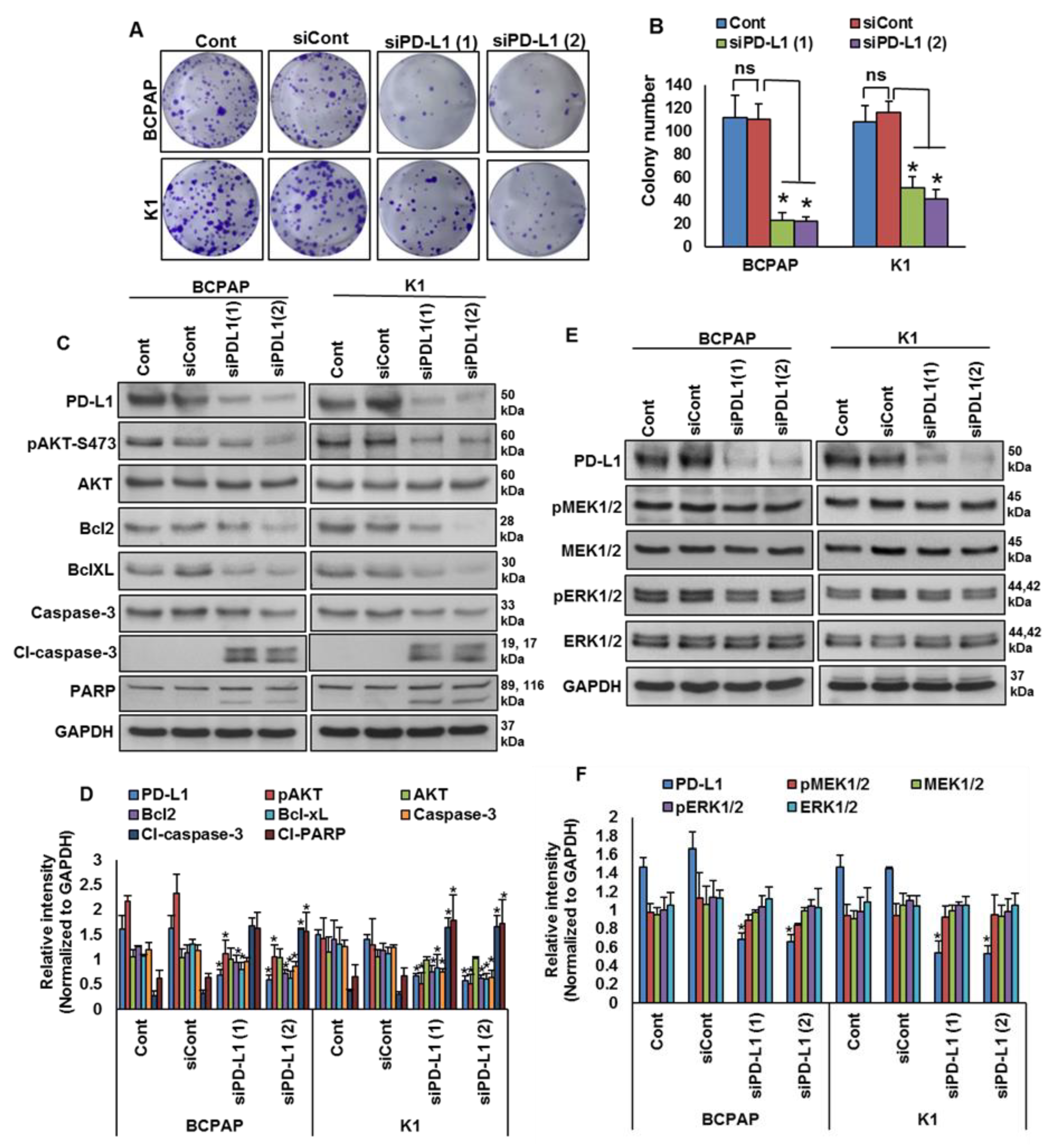
2.4. Downregulation of PD-L1 Decreases Cell Growth of BRAF-Mutated Cell Lines
3. Discussion
4. Materials and Methods
4.1. Sample Selection
4.2. DNA Isolation
4.3. Sanger Sequencing Analysis
4.4. Tissue Microarray (TMA) Construction and Immunohistochemistry (IHC)
4.5. Cell Culture
4.6. Reagents and Antibodies
4.7. Clonogenic Assay
4.8. Gene Silencing Using siRNA
4.9. Cell Lysis and Immuno-Blotting
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howlader, N.; Krapcho, M.; Miller, D.; Bishop, K.; Kosary, C.; Yu, M.; Cronin, K.A.; Noone, A.M.; Ruhl, J.; Tatalovich, Z.; et al. (Eds.) SEER Cancer Statistics Review, 1975–2014; National Cancer Institute: Bethesda, MD, USA, 2017. [Google Scholar]
- Schneider, D.F.; Chen, H. New developments in the diagnosis and treatment of thyroid cancer. CA Cancer J. Clin. 2013, 63, 373–394. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Alzahrani, A.S.; Carson, K.A.; Shong, Y.K.; Kim, T.Y.; Viola, D.; Elisei, R.; Bendlová, B.; Yip, L.; Mian, C.; et al. Association between BRAF V600E mutation and recurrence of papillary thyroid cancer. J. Clin. Oncol. 2015, 33, 42. [Google Scholar] [CrossRef] [PubMed]
- Ritter, A.; Mizrachi, A.; Bachar, G.; Vainer, I.; Shimon, I.; Hirsch, D.; Diker-Cohen, T.; Duskin-Bitan, H.; Robenshtok, E. Detecting recurrence following lobectomy for thyroid cancer: Role of thyroglobulin and thyroglobulin antibodies. J. Clin. Endocrinol. Metab. 2020, 105, e2145–e2151. [Google Scholar] [CrossRef] [PubMed]
- Tumino, D.; Frasca, F.; Newbold, K. Updates on the management of advanced, metastatic, and radioiodine refractory differentiated thyroid cancer. Front. Endocrinol. 2017, 8, 312. [Google Scholar] [CrossRef] [PubMed]
- Darvin, P.; Toor, S.M.; Nair, V.S.; Elkord, E. Immune checkpoint inhibitors: Recent progress and potential biomarkers. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Schadendorf, D.; Dummer, R.; Smylie, M.; Rutkowski, P.; et al. Combined nivolumab and ipilimumab or monotherapy in untreated melanoma. N. Engl. J. Med. 2015, 373, 23–34. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, C.; Jin, L.; Zhang, X. Regulation of PD-L1: A novel role of pro-survival signalling in cancer. Ann. Oncol. 2016, 27, 409–416. [Google Scholar] [CrossRef]
- Chen, N.; Fang, W.; Zhan, J.; Hong, S.; Tang, Y.; Kang, S.; Zhang, Y.; He, X.; Zhou, T.; Qin, T.; et al. Upregulation of PD-L1 by EGFR activation mediates the immune escape in EGFR-driven NSCLC: Implication for optional immune targeted therapy for NSCLC patients with EGFR mutation. J. Thorac. Oncol. 2015, 10, 910–923. [Google Scholar] [CrossRef]
- Zou, W.; Wolchok, J.D.; Chen, L. PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: Mechanisms, response biomarkers, and combinations. Sci. Transl. Med. 2016, 8, rv324–rv328. [Google Scholar] [CrossRef]
- Disis, M.L.; Taylor, M.H.; Kelly, K.; Beck, J.T.; Gordon, M.; Moore, K.M.; Patel, M.R.; Chaves, J.; Park, H.; Mita, A.C.; et al. Efficacy and safety of avelumab for patients with recurrent or refractory ovarian cancer: Phase 1b results from the JAVELIN solid tumor trial. JAMA Oncol. 2019, 5, 393–401. [Google Scholar] [CrossRef]
- Sheng, X.; Yan, X.; Chi, Z.; Si, L.; Cui, C.; Tang, B.; Li, S.; Mao, L.; Lian, B.; Wang, X.; et al. Axitinib in combination with toripalimab, a humanized immunoglobulin G4 monoclonal antibody against programmed cell death-1, in patients with metastatic mucosal melanoma: An open-label phase IB trial. J. Clin. Oncol. 2019, 37, 2987–2999. [Google Scholar] [CrossRef] [PubMed]
- Schmid, P.; Adams, S.; Rugo, H.S.; Schneeweiss, A.; Barrios, C.H.; Iwata, H.; Diéras, V.; Hegg, R.; Im, S.-A.; Shaw Wright, G.; et al. Atezolizumab and nab-paclitaxel in advanced triple-negative breast cancer. N. Engl. J. Med. 2018, 379, 2108–2121. [Google Scholar] [CrossRef] [PubMed]
- Dang, T.O.; Ogunniyi, A.; Barbee, M.S.; Drilon, A. Pembrolizumab for the treatment of PD-L1 positive advanced or metastatic non-small cell lung cancer. Expert Rev. Anticancer. Ther. 2016, 16, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Koemans, W.J.; Chalabi, M.; van Sandick, J.W.; van Dieren, J.M.; Kodach, L.L. Beyond the PD-L1 horizon: In search for a good biomarker to predict success of immunotherapy in gastric and esophageal adenocarcinoma. Cancer Lett. 2019, 442, 279–286. [Google Scholar] [CrossRef]
- Taube, J.M.; Klein, A.; Brahmer, J.R.; Xu, H.; Pan, X.; Kim, J.H.; Chen, L.; Pardoll, D.M.; Topalian, S.L.; Anders, R.A. Association of PD-1, PD-1 ligands, and other features of the tumor immune microenvironment with response to anti–PD-1 therapy. Clin. Cancer Res. 2014, 20, 5064–5074. [Google Scholar]
- Doroshow, D.B.; Sanmamed, M.F.; Hastings, K.; Politi, K.; Rimm, D.L.; Chen, L.; Melero, I.; Schalper, K.A.; Herbst, R.S. Immunotherapy in non–small cell lung cancer: Facts and hopes. Clin. Cancer Res. 2019, 25, 4592–4602. [Google Scholar] [CrossRef]
- Mehnert, J.M.; Varga, A.; Brose, M.S.; Aggarwal, R.R.; Lin, C.-C.; Prawira, A.; De Braud, F.; Tamura, K.; Doi, T.; Piha-Paul, S.A.; et al. Safety and antitumor activity of the anti–PD-1 antibody pembrolizumab in patients with advanced, PD-L1–positive papillary or follicular thyroid cancer. BMC Cancer 2019, 19, 196. [Google Scholar] [CrossRef]
- Azuma, T.; Yao, S.; Zhu, G.; Flies, A.S.; Flies, S.J.; Chen, L. B7-H1 is a ubiquitous antiapoptotic receptor on cancer cells. Blood J. Am. Soc. Hematol. 2008, 111, 3635–3643. [Google Scholar] [CrossRef]
- Clark, C.A.; Gupta, H.B.; Sareddy, G.; Pandeswara, S.; Lao, S.; Yuan, B.; Drerup, J.M.; Padron, A.; Conejo-Garcia, J.; Murthy, K.; et al. Tumor-intrinsic PD-L1 signals regulate cell growth, pathogenesis, and autophagy in ovarian cancer and melanoma. Cancer Res. 2016, 76, 6964–6974. [Google Scholar] [CrossRef]
- Lastwika, K.J.; Wilson, W.; Li, Q.K.; Norris, J.; Xu, H.; Ghazarian, S.R.; Kitagawa, H.; Kawabata, S.; Taube, J.M.; Yao, S.; et al. Control of PD-L1 expression by oncogenic activation of the AKT–mTOR pathway in non–small cell lung cancer. Cancer Res. 2016, 76, 227–238. [Google Scholar] [CrossRef]
- Kleffel, S.; Posch, C.; Barthel, S.R.; Mueller, H.; Schlapbach, C.; Guenova, E.; Elco, C.P.; Lee, N.; Juneja, V.R.; Zhan, Q.; et al. Melanoma cell-intrinsic PD-1 receptor functions promote tumor growth. Cell 2015, 162, 1242–1256. [Google Scholar] [CrossRef] [PubMed]
- Orzechowski, A.; Dong, H. B7-H1 confers tumor chemoresistance by regulating MAPK/ERK activation. In Proceedings of the AACR Annual Meeting, San Diego, CA, USA, 5–9 April 2014. [Google Scholar]
- Mohanty, A.; Afkhami, M.; Pharaon, R.; Yin, H.; Sun, J.; Nam, A.; Chang, S.; Gernon, T.; Kang, R.; Amini, A.; et al. Inhibition of BRAF induces PD-L1 expression in BRAF-mutated papillary thyroid carcinoma. Int. J. Radiat. Oncol. Biol. Phys. 2020, 106, 1192–1193. [Google Scholar] [CrossRef]
- Zaballos, M.A.; Santisteban, P. Key signaling pathways in thyroid cancer. J. Endocrinol. 2017, 235, R43–R61. [Google Scholar] [CrossRef] [PubMed]
- Gan, X.; Shen, F.; Deng, X.; Feng, J.; Lu, J.; Cai, W.; Peng, L.; Zheng, W.; Wang, W.; Huang, P.; et al. Prognostic implications of the BRAF-V600E mutation in papillary thyroid carcinoma based on a new cut-off age stratification. Oncol. Lett. 2020, 19, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Katano, T.; Oda, T.; Sekine, A.; Sato, M.; Yamaya, T.; Sato, Y.; Okudela, K.; Hagiwara, E.; Ogura, T. Five cases of BRAF V600E-mutant lung adenocarcinoma with high expression of programmed death ligand 1. Respir. Med. Case Rep. 2020, 30, 101071. [Google Scholar] [CrossRef] [PubMed]
- Dudnik, E.; Peled, N.; Nechushtan, H.; Wollner, M.; Onn, A.; Agbarya, A.; Moskovitz, M.; Keren, S.; Popovits-Hadari, N.; Urban, D.; et al. BRAF mutant lung cancer: Programmed death ligand 1 expression, tumor mutational burden, microsatellite instability status, and response to immune check-point inhibitors. J. Thorac. Oncol. 2018, 13, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Angell, T.E.; Lechner, M.G.; Jang, J.K.; Correa, A.J.; LoPresti, J.S.; Epstein, A.L. BRAFV600E in papillary thyroid carcinoma is associated with increased programmed death ligand 1 expression and suppressive immune cell infiltration. Thyroid 2014, 24, 1385–1393. [Google Scholar] [CrossRef]
- Feng, D.; Qin, B.; Pal, K.; Sun, L.; Dutta, S.; Dong, H.; Liu, X.; Mukhopadhyay, D.; Huang, S.; Sinicrope, F.A. BRAF V600E-induced, tumor intrinsic PD-L1 can regulate chemotherapy-induced apoptosis in human colon cancer cells and in tumor xenografts. Oncogene 2019, 38, 6752–6766. [Google Scholar]
- Shi, R.-l.; Qu, N.; Luo, T.-x.; Xiang, J.; Liao, T.; Sun, G.-h.; Wang, Y.; Wang, Y.-l.; Huang, C.-p.; Ji, Q.-h. Programmed death-ligand 1 expression in papillary thyroid cancer and its correlation with clinicopathologic factors and recurrence. Thyroid 2017, 27, 537–545. [Google Scholar] [CrossRef]
- Ahn, S.; Kim, T.H.; Kim, S.W.; Ki, C.S.; Jang, H.W.; Kim, J.S.; Kim, J.H.; Choe, J.-H.; Shin, J.H.; Hahn, S.Y.; et al. Comprehensive screening for PD-L1 expression in thyroid cancer. Endocr. Relat. Cancer 2017, 24, 97–106. [Google Scholar] [CrossRef]
- Chowdhury, S.; Veyhl, J.; Jessa, F.; Polyakova, O.; Alenzi, A.; MacMillan, C.; Ralhan, R.; Walfish, P.G. Programmed death-ligand 1 overexpression is a prognostic marker for aggressive papillary thyroid cancer and its variants. Oncotarget 2016, 7, 32318. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Devesa, S.S.; Sosa, J.A.; Check, D.; Kitahara, C.M. Trends in thyroid cancer incidence and mortality in the United States, 1974–2013. JAMA 2017, 317, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Alzahrani, A.S.; Carson, K.A.; Viola, D.; Elisei, R.; Bendlova, B.; Yip, L.; Mian, C.; Vianello, F.; Tuttle, R.M.; et al. Association between BRAF V600E mutation and mortality in patients with papillary thyroid cancer. JAMA 2013, 309, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Howlader, N.; Noone, A.; Krapcho, M.; Garshell, J.; Miller, D.; Altekruse, S.; Kosary, C.; Yu, M.; Ruhl, J.; Tatalovich, Z.; et al. (Eds.) SEER Cancer Statistics Review, 1975–2012; National Cancer Institute: Bethesda, MD, USA, 2015. [Google Scholar]
- Fallahi, P.; Mazzi, V.; Vita, R.; Ferrari, S.M.; Materazzi, G.; Galleri, D.; Benvenga, S.; Miccoli, P.; Antonelli, A. New therapies for dedifferentiated papillary thyroid cancer. Int. J. Mol. Sci. 2015, 16, 6153–6182. [Google Scholar] [CrossRef]
- Fugazzola, L.; Elisei, R.; Fuhrer, D.; Jarzab, B.; Leboulleux, S.; Newbold, K.; Smit, J. 2019 European Thyroid Association Guidelines for the Treatment and Follow-Up of Advanced Radioiodine-Refractory Thyroid Cancer. Eur. Thyroid. J. 2019, 8, 227–245. [Google Scholar] [CrossRef]
- Yi, M.; Jiao, D.; Xu, H.; Liu, Q.; Zhao, W.; Han, X.; Wu, K. Biomarkers for predicting efficacy of PD-1/PD-L1 inhibitors. Mol. Cancer 2018, 17, 1–14. [Google Scholar] [CrossRef]
- Davis, A.A.; Patel, V.G. The role of PD-L1 expression as a predictive biomarker: An analysis of all US Food and Drug Administration (FDA) approvals of immune checkpoint inhibitors. J. Immunother. Cancer 2019, 7, 278. [Google Scholar] [CrossRef]
- Li, Y.; Liang, L.; Dai, W.; Cai, G.; Xu, Y.; Li, X.; Li, Q.; Cai, S. Prognostic impact of programed cell death-1 (PD-1) and PD-ligand 1 (PD-L1) expression in cancer cells and tumor infiltrating lymphocytes in colorectal cancer. Mol. Cancer 2016, 15, 55. [Google Scholar] [CrossRef]
- Qin, T.; Zeng, Y.-D.; Qin, G.; Xu, F.; Lu, J.-B.; Fang, W.-F.; Xue, C.; Zhan, J.-H.; Zhang, X.-K.; Zheng, Q.-F.; et al. High PD-L1 expression was associated with poor prognosis in 870 Chinese patients with breast cancer. Oncotarget 2015, 6, 33972. [Google Scholar] [CrossRef]
- Wang, Q.; Lou, W.; Di, W.; Wu, X. Prognostic value of tumor PD-L1 expression combined with CD8+ tumor infiltrating lymphocytes in high grade serous ovarian cancer. Int. Immunopharmacol. 2017, 52, 7–14. [Google Scholar] [CrossRef]
- Pawelczyk, K.; Piotrowska, A.; Ciesielska, U.; Jablonska, K.; Glatzel-Plucinska, N.; Grzegrzolka, J.; Podhorska-Okolow, M.; Dziegiel, P.; Nowinska, K. Role of PD-L1 expression in non-small cell lung cancer and their prognostic significance according to clinicopathological factors and diagnostic markers. Int. J. Mol. Sci. 2019, 20, 824. [Google Scholar] [CrossRef] [PubMed]
- Aghajani, M.J.; Yang, T.; McCafferty, C.E.; Graham, S.; Wu, X.; Niles, N. Predictive relevance of programmed cell death protein 1 and tumor-infiltrating lymphocyte expression in papillary thyroid cancer. Surgery 2018, 163, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Sumimoto, H.; Takano, A.; Teramoto, K.; Daigo, Y. RAS–mitogen-activated protein kinase signal is required for enhanced PD-L1 expression in human lung cancers. PLoS ONE 2016, 11, e0166626. [Google Scholar] [CrossRef]
- Della Corte, C.M.; Barra, G.; Ciaramella, V.; Di Liello, R.; Vicidomini, G.; Zappavigna, S.; Luce, A.; Abate, M.; Fiorelli, A.; Caraglia, M.; et al. Antitumor activity of dual blockade of PD-L1 and MEK in NSCLC patients derived three-dimensional spheroid cultures. J. Exp. Clin. Cancer Res. 2019, 38, 253. [Google Scholar] [CrossRef] [PubMed]
- Stutvoet, T.S.; Kol, A.; de Vries, E.G.; de Bruyn, M.; Fehrmann, R.S.; Terwisscha van Scheltinga, A.G.; de Jong, S. MAPK pathway activity plays a key role in PD-L1 expression of lung adenocarcinoma cells. J. Pathol. 2019, 249, 52–64. [Google Scholar] [CrossRef]
- Ghebeh, H.; Lehe, C.; Barhoush, E.; Al-Romaih, K.; Tulbah, A.; Al-Alwan, M.; Hendrayani, S.-F.; Manogaran, P.; Alaiya, A.; Al-Tweigeri, T.; et al. Doxorubicin downregulates cell surface B7-H1 expression and upregulates its nuclear expression in breast cancer cells: Role of B7-H1 as an anti-apoptotic molecule. Breast Cancer Res. 2010, 12, R48. [Google Scholar] [CrossRef]
- Liu, J.; Quan, L.; Zhang, C.; Liu, A.; Tong, D.; Wang, J. Over-activated PD-1/PD-L1 axis facilitates the chemoresistance of diffuse large B-cell lymphoma cells to the CHOP regimen. Oncol. Lett. 2018, 15, 3321–3328. [Google Scholar] [CrossRef]
- Yan, F.; Pang, J.; Peng, Y.; Molina, J.R.; Yang, P.; Liu, S. Elevated cellular PD1/PD-L1 expression confers acquired resistance to cisplatin in small cell lung cancer cells. PLoS ONE 2016, 11, e0162925. [Google Scholar] [CrossRef]
- Abubaker, J.; Jehan, Z.; Bavi, P.; Sultana, M.; Al-Harbi, S.; Ibrahim, M.; Al-Nuaim, A.; Ahmed, M.; Amin, T.; Al-Fehaily, M.; et al. Clinicopathological analysis of papillary thyroid cancer with PIK3CA alterations in a Middle Eastern population. J. Clin. Endocrinol. Metab. 2008, 93, 611–618. [Google Scholar] [CrossRef]
- Bu, R.; Siraj, A.K.; Al-Obaisi, K.A.; Beg, S.; Al Hazmi, M.; Ajarim, D.; Tulbah, A.; Al-Dayel, F.; Al-Kuraya, K.S. Identification of novel BRCA founder mutations in Middle Eastern breast cancer patients using capture and Sanger sequencing analysis. Int. J. Cancer 2016, 139, 1091–1097. [Google Scholar] [CrossRef]
- Siraj, A.; Bavi, P.; Abubaker, J.; Jehan, Z.; Sultana, M.; Al-Dayel, F.; Al-Nuaim, A.; Alzahrani, A.; Ahmed, M.; Al-Sanea, O.; et al. Genome-wide expression analysis of Middle Eastern papillary thyroid cancer reveals c-MET as a novel target for cancer therapy. J. Pathol. 2007, 213, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Bavi, P.; Jehan, Z.; Atizado, V.; Al-Dossari, H.; Al-Dayel, F.; Tulbah, A.; Amr, S.S.; Sheikh, S.S.; Ezzat, A.; El-Solh, H.; et al. Prevalence of fragile histidine triad expression in tumors from Saudi Arabia: A tissue microarray analysis. Cancer Epidemiol. Prev. Biomark. 2006, 15, 1708–1718. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mesnage, S.; Auguste, A.; Genestie, C.; Dunant, A.; Pain, E.; Drusch, F.; Gouy, S.; Morice, P.; Bentivegna, E.; Lhomme, C.; et al. Neoadjuvant chemotherapy (NACT) increases immune infiltration and programmed death-ligand 1 (PD-L1) expression in epithelial ovarian cancer (EOC). Ann. Oncol. 2017, 28, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Sano, D.; Pickering, C.R.; Jasser, S.A.; Henderson, Y.C.; Clayman, G.L.; Sturgis, E.M.; Ow, T.J.; Lotan, R.; Carey, T.E.; et al. Assembly and initial characterization of a panel of 85 genomically validated cell lines from diverse head and neck tumor sites. Clin. Cancer Res. 2011, 17, 7248–7264. [Google Scholar] [CrossRef] [PubMed]
- Bu, R.; Siraj, A.K.; Divya, S.P.; Kong, Y.; Parvathareddy, S.K.; Al-Rasheed, M.; Al-Obaisi, K.A.; Victoria, I.G.; Al-Sobhi, S.S.; Al-Dawish, M.; et al. Telomerase reverse transcriptase mutations are independent predictor of disease-free survival in Middle Eastern papillary thyroid cancer. Int. J. Cancer 2018, 142, 2028–2039. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | PD-L1 High | PD-L1 Low | p Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No. | % | No. | % | No. | % | |||||
| No. of Patients | 1458 | 473 | 32.4 | 985 | 67.6 | |||||
| Age (Years) | ||||||||||
| <55 | 1189 | 81.5 | 389 | 32.7 | 800 | 67.3 | 0.6365 | |||
| ≥55 | 269 | 18.5 | 84 | 31.2 | 185 | 68.8 | ||||
| Sex | ||||||||||
| Female | 1102 | 75.6 | 370 | 33.6 | 732 | 66.4 | 0.1014 | |||
| Male | 356 | 24.4 | 103 | 28.9 | 253 | 71.1 | ||||
| Extrathyroidal Extension | ||||||||||
| Absent | 834 | 57.2 | 250 | 30.0 | 584 | 70.0 | 0.0203 (0.0271) * | |||
| Present | 624 | 42.8 | 223 | 35.7 | 401 | 64.3 | ||||
| pT | ||||||||||
| pT1 | 400 | 28.5 | 130 | 32.5 | 270 | 67.5 | 0.5456 | |||
| pT2 | 298 | 21.2 | 89 | 29.9 | 209 | 70.1 | ||||
| pT3 | 595 | 42.4 | 204 | 34.3 | 391 | 65.7 | ||||
| pT4 | 110 | 7.8 | 33 | 30.0 | 77 | 70.0 | ||||
| pN | ||||||||||
| pN0 | 582 | 44.4 | 176 | 30.2 | 406 | 69.8 | 0.0466 (0.0466) * | |||
| pN1 | 728 | 55.6 | 258 | 35.4 | 470 | 64.6 | ||||
| pM | ||||||||||
| pM0 | 1401 | 96.1 | 454 | 32.4 | 947 | 67.6 | 0.8836 | |||
| pM1 | 57 | 3.9 | 19 | 33.3 | 38 | 66.7 | ||||
| Stage | ||||||||||
| I | 1188 | 84.3 | 380 | 32.0 | 808 | 68.0 | 0.9322 | |||
| II | 152 | 10.8 | 53 | 34.9 | 99 | 65.1 | ||||
| III | 19 | 1.4 | 6 | 31.6 | 13 | 68.4 | ||||
| IVA | 19 | 1.4 | 5 | 26.3 | 14 | 73.7 | ||||
| IVB | 30 | 2.1 | 10 | 33.3 | 20 | 66.7 | ||||
| Histology Type | ||||||||||
| Classical Variant | 955 | 65.5 | 334 | 35.0 | 621 | 65.0 | <0.0001 (<0.0001) * | |||
| Follicular Variant | 258 | 17.7 | 54 | 20.9 | 204 | 79.1 | ||||
| Tall Cell Variant | 135 | 9.3 | 54 | 40.0 | 81 | 60.0 | ||||
| Other Variants | 110 | 7.5 | 31 | 28.2 | 79 | 71.8 | ||||
| BRAF Mutation | ||||||||||
| Yes | 707 | 56.2 | 250 | 35.4 | 457 | 64.6 | 0.0183 (0.0271) * | |||
| No | 550 | 43.8 | 160 | 29.1 | 390 | 70.9 | ||||
| Disease-Free Survival | ||||||||||
| 5 years | 266 | 62.2 | 751 | 79.2 | <0.0001 | |||||
| Recurrence-Free Survival | ||||||||||
| 5 years | 354 | 82.0 | 806 | 86.5 | 0.0006 | |||||
| Disease-Free Survival | Recurrence-Free Survival | |||||||
|---|---|---|---|---|---|---|---|---|
| Univariate | Multivariate | Univariate | Multivariate | |||||
| Clinico-pathological Variables | Risk Ratio (95% CI) | p Value | Risk Ratio (95% CI) | p Value | Risk Ratio (95% CI) | p Value | Risk Ratio (95% CI) | p Value |
| Age Above ≥55 years (vs. <55 years) | 2.54 (2.03–3.16) | <0.0001 * | 2.13 (1.59–2.87) | <0.0001 * | 2.97 (2.31–3.80) | <0.0001 * | 2.65 (1.90–3.70) | <0.0001 * |
| Sex Male (vs. Female) | 0.59 (0.48–0.73) | <0.0001 * | 0.67 (0.52–0.86) | 0.0016 * | 0.56 (0.44–0.71) | <0.0001 * | 0.71 (0.53–0.95) | 0.0224 * |
| Histology Tall Cell Variant (vs. Other Variants) | 1.94 (1.41–2.61) | <0.0001 * | 1.48 (1.04–2.05) | 0.0236 * | 1.27 (0.84–1.83) | 0.2528 | 0.93 (0.59–1.40) | 0.7422 |
| Extrathyroidal Extension Present (vs. Absent) | 2.27 (1.82–2.86) | <0.0001 * | 1.40 (1.07–1.82) | 0.0149 * | 2.92 (2.24–3.85) | <0.0001 * | 1.72 (1.25–2.36) | 0.0008 * |
| Lymph Node Metastasis N1 (vs. N0) | 2.33 (1.84–2.96) | <0.0001 * | 1.72 (1.30–2.26) | 0.0001 * | 2.74 (2.09–3.63) | <0.0001 * | 2.31 (1.65–3.23) | <0.0001 * |
| Distant Metastasis Present (vs. Absent) | 3.99 (2.83–5.62) | <0.0001 * | 2.29 (1.43–3.69) | 0.0006 * | 6.14 (4.25–8.62) | <0.0001 * | 2.78 (1.68–4.60) | <0.0001 * |
| Stage IV (vs. I-III) | 3.62 (2.39–5.27) | <0.0001 * | 0.83 (0.46–1.49) | 0.5305 | 6.25 (4.12–9.10) | <0.0001 * | 0.81 (0.44–1.52) | 0.5164 |
| PD-L1 High (vs. Low) | 2.04 (1.66–2.52) | <0.0001 * | 2.08 (1.65–2.62) | <0.0001 * | 1.51 (1.19–1.92) | 0.0008 * | 1.54 (1.18–2.00) | 0.0013 * |
| Clinico-Pathological Variables | n (%) |
|---|---|
| Age | |
| Median | 38.0 |
| Range (IQR) ^ | 29.0–50.0 |
| <55 years | 1189 (81.5) |
| ≥55 years | 269 (18.5) |
| Gender | |
| Female | 1102 (75.6) |
| Male | 356 (24.4) |
| Histopathology | |
| Classical Variant | 955 (65.5) |
| Follicular Variant | 258 (17.7) |
| Tall Cell Variant | 135 (9.3) |
| Others | 110 (7.5) |
| Extrathyroidal Extension | |
| Absent | 834 (57.2) |
| Present | 624 (42.8) |
| pT | |
| T1 | 400 (27.4) |
| T2 | 298 (20.4) |
| T3 | 595 (40.9) |
| T4 | 110 (7.5) |
| Unknown | 55 (3.8) |
| pN | |
| N0 | 582 (39.9) |
| N1 | 728 (49.9) |
| Nx | 148 (10.2) |
| pM | |
| M0 | 1401 (96.1) |
| M1 | 57 (3.9) |
| Stage | |
| I | 1188 (81.5) |
| II | 152 (10.4) |
| III | 19 (1.3) |
| IVA | 19 (1.3) |
| IVB | 30 (2.1) |
| Unknown | 50 (3.4) |
| BRAF Mutation | |
| Present | 707 (48.5) |
| Absent | 550 (37.7) |
| Unknown | 201 (13.8) |
| RAI Therapy | |
| Yes | 1113 (76.3%) |
| No | 345 (23.7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siraj, A.K.; Parvathareddy, S.K.; Pratheeshkumar, P.; Divya, S.P.; Al-Sobhi, S.S.; Al-Dayel, F.; Al-Kuraya, K.S. PD-L1 Is an Independent Prognostic Marker in Middle Eastern PTC and Its Expression Is Upregulated by BRAFV600E Mutation. Cancers 2021, 13, 555. https://doi.org/10.3390/cancers13030555
Siraj AK, Parvathareddy SK, Pratheeshkumar P, Divya SP, Al-Sobhi SS, Al-Dayel F, Al-Kuraya KS. PD-L1 Is an Independent Prognostic Marker in Middle Eastern PTC and Its Expression Is Upregulated by BRAFV600E Mutation. Cancers. 2021; 13(3):555. https://doi.org/10.3390/cancers13030555
Chicago/Turabian StyleSiraj, Abdul K., Sandeep Kumar Parvathareddy, Poyil Pratheeshkumar, Sasidharan Padmaja Divya, Saif S. Al-Sobhi, Fouad Al-Dayel, and Khawla S. Al-Kuraya. 2021. "PD-L1 Is an Independent Prognostic Marker in Middle Eastern PTC and Its Expression Is Upregulated by BRAFV600E Mutation" Cancers 13, no. 3: 555. https://doi.org/10.3390/cancers13030555
APA StyleSiraj, A. K., Parvathareddy, S. K., Pratheeshkumar, P., Divya, S. P., Al-Sobhi, S. S., Al-Dayel, F., & Al-Kuraya, K. S. (2021). PD-L1 Is an Independent Prognostic Marker in Middle Eastern PTC and Its Expression Is Upregulated by BRAFV600E Mutation. Cancers, 13(3), 555. https://doi.org/10.3390/cancers13030555