Simple Summary
Endemic Burkitt lymphoma (eBL) is a common pediatric cancer in sub-Saharan Africa. The incidence of this aggressive B-cell cancer is linked to Plasmodium falciparum (Pf) malaria and Epstein–Barr virus (EBV) co-infections during childhood. Most eBL tumors contain EBV and are characterized by the Epstein–Barr Nuclear Antigen 1 (EBNA1) latency I pattern of viral gene expression. The aim of our study was to compare the phenotypes and functions of CD4+ and CD8+ T cell responses to EBNA1 in children diagnosed with eBL and in healthy EBV-seropositive children to highlight differences that contribute to the balance between anti-viral immunity and eBL pathogenesis.
Abstract
Children diagnosed with endemic Burkitt lymphoma (eBL) are deficient in interferon-γ (IFN-γ) responses to Epstein–Barr Nuclear Antigen1 (EBNA1), the viral protein that defines the latency I pattern in this B cell tumor. However, the contributions of immune-regulatory cytokines and phenotypes of the EBNA1-specific T cells have not been characterized for eBL. Using a bespoke flow cytometry assay we measured intracellular IFN-γ, IL-10, IL-17A expression and phenotyped CD4+ and CD8+ T cell effector memory subsets specific to EBNA1 for eBL patients compared to two groups of healthy children with divergent malaria exposures. In response to EBNA1 and a malaria antigen (PfSEA-1A), the three study groups exhibited strikingly different cytokine expression and T cell memory profiles. EBNA1-specific IFN-γ-producing CD4+ T cell response rates were lowest in eBL (40%) compared to children with high malaria (84%) and low malaria (66%) exposures (p < 0.0001 and p = 0.0004, respectively). However, eBL patients did not differ in CD8+ T cell response rates or the magnitude of IFN-γ expression. In contrast, eBL children were more likely to have EBNA1-specific CD4+ T cells expressing IL-10, and less likely to have polyfunctional IFN-γ+IL-10+ CD4+ T cells (p = 0.02). They were also more likely to have IFN-γ+IL-17A+, IFN-γ+ and IL-17A+ CD8+ T cell subsets compared to healthy children. Cytokine-producing T cell subsets were predominantly CD45RA+CCR7+ TNAIVE-LIKE cells, yet PD-1, a marker of persistent activation/exhaustion, was more highly expressed by the central memory (TCM) and effector memory (TEM) T cell subsets. In summary, our study suggests that IL-10 mediated immune regulation and depletion of IFN-γ+ EBNA1-specific CD4+ T cells are complementary mechanisms that contribute to impaired T cell cytotoxicity in eBL pathogenesis.
1. Introduction
Endemic Burkitt lymphoma (eBL), the most common pediatric cancer in equatorial Africa [1,2] has long been associated with early-age infections by two pathogens: Epstein–Barr virus (EBV) and Plasmodium falciparum [3,4]. Children at risk of eBL experience primary EBV infection before 2 years of age [5,6,7] and have malaria-associated episodes of viral reactivation resulting in cumulatively higher EBV loads [6,7,8] and IFN-γ deficiencies against EBV lytic and latent antigens [9,10]. Recent studies have shown that the EBV status of eBL tumors is associated with distinct mutational landscapes [11,12,13], supporting the premise that EBV plays a role that transcends tumorigenesis. During early childhood, repeated P. falciparum (Pf) infections are also common for those residing in holoendemic areas and severe disease is the leading cause of morbidity and mortality in children under 5 years of age [14]. However, eBL is most often diagnosed between 5 and 9 years of age [2], after the development of premunition which is characterized by the ability to harbor asymptomatic and, therefore, untreated persistent Pf-malaria infections. Thus, prolonged malaria exposure has been postulated to diminish EBV-specific T cell immune surveillance as a risk factor in eBL pathogenesis [15,16,17,18]. Despite the long-standing epidemiological association, our understanding of how malaria disrupts the balance between antiviral immunity, which is insufficient to eliminate EBV yet sufficient to limit eBL oncogenesis, is incomplete. Furthering our understanding of the underlying mechanisms involved in the etiology of eBL will help identify alterations in immune cells that are important to control EBV.
Our knowledge of EBV-specific T cell immunity that minimizes its pathologic impact has been gained primarily from studying young adults residing in non-malarious areas, where robust immunity to EBV lytic and latent antigens decreases viral loads to undetectable levels [19,20]. Once this persistent herpesvirus infection is established, EBV evades immune detection by down-regulating viral protein expression during latency [21]. Of particular interest as an immunotherapeutic target is the Epstein–Barr nuclear antigen 1 (EBNA1), which is the viral antigen consistently expressed by all EBV-associated B cell tumors [22], as well as during the lytic and latent cycles of all EBV infections [23]. EBNA1 is recognized by CD4+ T cells via macroautophagy [24] but decreases translation and proteasomal processing by major histocompatibility (MHC) class I molecules due to a glycine-alanine repeat domain [25,26] thereby limiting the development of CD8+ T cell responses, which are engaged by non-classical MHC class I cross-presentation [27,28,29]. Our studies of children diagnosed with eBL were the first to describe a selective deficiency in IFN-γ responses to EBNA1 [18]. However, this study was limited to measuring IFN-γ by ELISPOT assay which did not determine the phenotype of EBNA1-specific cells or alternative cytokine expression profiles.
Based on these observations, we set forth to explore the influence of prolonged exposure to Pf during childhood on the quality of EBNA1-specific T cell immunity. First, we sought to determine if Pf co-infections led to the polarization of EBNA1-specific T cell immunity away from Th1 (IFN-γ-producing) towards anti-inflammatory, immune-regulatory T cells (i.e., IL-10 expressing) [30], or the generation of an alternative proinflammatory Th17 (IL-17A) phenotype [31]. For those with EBNA1-specific T cell responses, we compared the phenotype of the cytokine-producing cells to determine if young age or exposure to malaria were associated with distinct T cell subsets. We further explored the potential heterogeneity in human T cell responses, which has been described for many infectious diseases, including malaria [32], and characterized based on the expression of CD45RA and CCR7 to define central memory, effector memory, terminally differentiated RA re-expressing effector memory and naïve-like cell subsets [33]. Finally, other studies reported the upregulated expression of programmed cell-death 1 (PD-1) in virus-specific T cells [34], during malaria infections [35] and other parasitic infections [36,37] as an indicator of T cell exhaustion or persistent antigen stimulation. We speculated that EBNA1-specific T cell exhaustion could be present in children after a prolonged exposure to Pf and concomitantly higher EBV loads as a precursor to eBL tumorigenesis.
We conducted a case–control study of children diagnosed with eBL compared to two groups of healthy children residing in areas of Kenya with divergent malaria endemicity. All three cohorts experienced their primary EBV infection before 2 years of age [7]. We determined the frequency and phenotype of CD4+ and CD8+ T cell subsets across our three study populations, defined by multiparameter flow cytometry (CCR7, CD45RA, PD-1) and intracellular cytokine (IFN-γ, IL-10 and IL-17A) expression performed on peripheral blood mononuclear cells (PBMCs) after the ex vivo stimulation with EBNA1. We also compared EBNA1 responses to T cell responses against a malaria-specific Pf schizont egress antigen-1A (PfSEA-1A) [38]. Through this study, we show different pathogen-dependent (i.e., EBNA1 or PfSEA-1A) cytokine expressions and T cell memory profiles between our three groups of children.
2. Materials and Methods
2.1. Study Populations
Children diagnosed with eBL were enrolled at Jaramogi Oginga Odinga Teaching and Referral Hospital, a cancer referral center in Kisumu, Kenya. Diagnosis was confirmed by two independent pathologists and molecular methods [11,39] and blood samples were collected prior to chemotherapy. Healthy children were enrolled from two counties in western Kenya that had divergent Pf transmission patterns and, accordingly, differed in their incidence of eBL [40]. Kisumu County, a malaria holoendemic region (i.e., a region with recurrent or persistent malaria infections during childhood as previously described, is referred to as “chronic” malaria exposure) and Nandi County, a hypo-endemic region for malaria (i.e., few “acute” malaria infections during childhood) [41]. Adults from Nandi (n = 3) and Kisumu (n = 4) were enrolled as controls. Study participants did not have blood stage malaria infections when venous blood samples were collected (diagnosed by blood smear). To confirm differences in EBV load between study groups, viral copies were quantified as previously described [7]. PBMCs were processed within 3 h by Ficoll-Hypaque density centrifugation followed by cryopreservation.
Written informed consent was obtained from adult study participants and the parents or guardians prior to their children’s enrollment in this study. Ethical approval was obtained from the Institutional Review Board at the University of Massachusetts Chan Medical School, Worcester, MA, USA and the Scientific and Ethics Review Unit at the Kenya Medical Research Institute, Nairobi, Kenya.
2.2. Characteristics of Study Populations
Study participant characteristics are summarized in Supplementary Table S1. Children in the three groups had similar male-to-female ratios and ages (ranging from 2 to 14 years, with a median age of 8). As previously published [6], EBV copies were significantly higher for eBL children compared to detectable viremia in healthy Kisumu (high malaria) and Nandi (low malaria) children (p < 0.0001 and p < 0.0001, respectively). Resting CD4/CD8 ratios did not differ between study groups; however, Nandi children had fewer CD19−CD14−CD3+ lymphocytes compared to Kisumu and eBL children (p = 0.002 and p = 0.01, respectively), suggesting a higher number of natural killer (NK) cells, consistent with our previously published study [42]. Finally, antibodies to EBNA1 and viral capsid antigen (VCA) demonstrate that all children enrolled in this study were EBV-seropositive. As expected, Nandi children from the low malaria exposure area had significantly lower antibody levels against malaria antigens, AMA1 (apical membrane antigen 1) and MSP1 (merozoite surface protein 1) compared to the two groups of children who resided in high malaria exposure areas; Kisumu (p < 0.0001 and p = 0.01, respectively) and eBL children (p < 0.0001 and p = 0.007, respectively). AMA1 and MSP1 proteins were gifts from Evelina Angov (Walter Reed Army Institute of Research, Silver Spring, MD, USA).
2.3. T Cell Immunophenotyping by Flow Cytometry
PBMCs were thawed and washed twice in complete media (RPMI 1640, GIBCO with 10% heat-inactivated and filter-sterilized fetal bovine serum, Glutamine 1X GIBCO 100 U/mL, Penicillin 100 mg/mL, Streptomycin 1X GIBCO, Hepes BioWhittaker 10 mM) at 37 °C. Live cells were counted using Propidium Iodide (PI) and MACSquant Flow cytometer before overnight incubation (37 °C, 5% CO2). Cells were re-counted the next morning, stimulating an average of 250,000 viable cells (minimum of 85% viability) with each one of the following conditions at final concentrations indicated in parenthesis: overlapping EBNA1 peptides (5.76 μM), EBNA2 (10 μg/mL), EBNA3A (10 μg/mL) [43], DMSO (0.13% as a negative control to assess background signal for each individual), or recombinant malaria antigen, PfSEA-1A [38] (5 μg/mL) for 24 h at 37 °C in 5% CO2. Cell viability did not differ after stimulation compared to unstimulated cells. We defined individuals as responders when they had higher cytokine expression after antigen stimulation compared to their DMSO/negative control.
Cells were gated on SSC-A vs. FSC-A and single-cell cytoplots (Figure S1A). Dump channel applied: CD14/CD19-Pacific Blue (RRID:AB_10373537 and RRID:AB_10373689, respectively) and the Live/dead Zombie Violet (BioLegend cat#423114) for selection of live CD14−CD19− cells. T cell subsets and phenotypes were defined by the following monoclonal antibodies: CD3 BV605 (RRID:AB_256437), CD4 PerCP (RRID:AB_2028492), CD8 FITC (RRID:AB_314124), CD45RA Alexa Fluor 700 (RRID:AB_493763), PD-1 PE-Cy7 (RRID:AB_2159324), CCR7 APC-eFluor780 (RRID:AB_1518794), IFN-γ PE-Dazzle 594 (RRID:AB_2563627), IL-10 APC (RRID:AB_398582) and IL-17A PE (RRID:AB_961395). Data were acquired on a 5 laser BD LSRII flow cytometer (UMMS Flow Core Facility) with DIVA software, compensation and FMO controls. Using Live/Dead staining, we obtained an average and median of 77.45% and 86.45% live lymphocytes, respectively. Since IL-10 was measured by intracellular staining of T cells, this was assumed to be human IL-10 and not the viral IL-10 homologue. We analyzed PD-1-positive T cells compared to the unstimulated DMSO control for each condition. T cells subsets were defined by expression of CD45RA and CCR7 as follows: terminal effector (TEMRA, CD45RA+CCR7−), naïve-like (TNAIVE-LIKE, CD45RA+CCR7+), central memory (TCM, CD45RA−CCR7+) and effector memory (TEM, CD45RA−CCR7−) [44]. The expected age-dependent biological differences across T cell subsets showed that children had more TNAIVE-LIKE cells compared to adults with more TEMRA and TEM cells (Figure S1B).
2.4. Statistical Analysis
Flow cytometry data were analyzed with FlowJo software, version 10.7.1 and GraphPad Prism software, version 9.1.2. Assessment of T cell polyfunctionality was determined using Boolean gating in FlowJo including 3 parameters: IFN-γ, IL-10 and IL-17A and analyzed with SPICE v6.0 [45]. Histograms from SPICE 6 showed the mean of mono/poly-functional expression across study groups and each dot represents one sample, a Wilcoxon Rank Sum Test was applied. Radar (also known as flower) plots were generated using the ggplot2 package in R program (v4.1.0, https://cran.r-project.org/web/packages/ggplot2, accessed on 30 June 2021). Categorical data were analyzed using a chi-square test. Because the number of responders within each group was too low to verify the normality of the underlying distributions, we chose to use non-parametric tests, including Mann–Whitney test (for unpaired analysis), Wilcoxon signed-rank test (for paired analysis), or ANOVA (Friedman for paired data and Kruskal–Wallis for unpaired data). All tests were two-tailed and p = 0.05 was set as the level of significance. Because of the exploratory nature of the analysis, we did not use any adjustment of the p value for multiple comparisons. The test used is indicated in the legend of each figure. Statistical analysis was performed in GraphPad Prism. Results were expressed using the mean with standard deviation (SD) for dot plots, the mean for the diagrams, and exact p-values.
3. Results
3.1. Heterogeneity of EBNA1 and PfSEA-1A Specific T Cell Responses
To determine if a history of chronic malaria exposure or the development of eBL was associated with T cell polarization, the frequency of IFN-γ, IL-10 and IL-17A CD4+ and CD8+ T cell responders to EBNA1 were compared across our three study groups. The frequencies of each cytokine response and the significance of the comparisons between the different groups of children are summarized in Table 1. Malaria-exposed Kisumu children were more likely to have IFN-γ+ CD4+ T cell responses (84%) to EBNA1 compared to Nandi (66%, p = 0.005) and eBL (40%, p < 0.0001) children; however, the frequency of IFN-γ+ CD8+ T cell responders did not differ (63%, 58% and 50%, respectively). In contrast, EBNA1 specific CD4+ T cell IL-10 responders were more frequent for eBL (73%) patients compared to Kisumu (38%) and Nandi (33%) children (p < 0.0001 for both) but did not differ for CD8+ T cells (56%, 54% and 52%, respectively). For EBNA1-specific CD4+ T cells expressing IL-17A, eBL and Nandi children had similar response rates (66% and 73%, respectively) which were significantly more prevalent than for Kisumu children (46%, p = 0.006 and p = 0.0002, respectively). However, IL-17A expressing CD8+ T cell response rates to EBNA1 were significantly different across the study groups, with eBL (68%) and Nandi (52%) having significantly more responders than the Kisumu children (18%, p < 0.0001 for both). Overall, differences in IFN-γ and IL-10 responders between our study groups involved CD4+ T cells to EBNA1, except for IL-17A, which differed for both CD4+ and CD8+ T cells.

Table 1.
The frequency of cytokine responders for EBNA1 and PfSEA-1A across study groups. A responder was defined as a child whose T cells produced significantly higher levels of cytokine after ex vivo antigen stimulation compared to their unstimulated, DMSO control cells using chi-square statistics. The frequency of responders was reported for each study group: Nandi, Kisumu and eBL children, and then compared across groups for each cytokine (i.e., IFN-γ, IL-10 or IL-17A), antigen (EBNA1 or PfSEA-1A) and type of T cell response (CD4+ and CD8+) by Fisher’s exact test; p-values < 0.05 were considered significant.
To provide a context for the observed differences in the T cell responses to EBNA1 we compared the frequency of responders within each group to a malaria antigen, PfSEA-1A. Nandi children, who resided in a low malaria exposure area, and thus putatively had acute malaria infections and demonstrably low or undetectable EBV loads, had the same frequency of CD4+ T cells (Figure 1A) expressing IFN-γ but significantly fewer IL-10 (p < 0.0001) and more IL-17A (p = 0.001) responders to EBNA1 compared to PfSEA-1A. In contrast, Kisumu children had similar frequencies of cytokine responders to the EBV and malaria antigens used in our assay, while children diagnosed with eBL had significantly more CD4+ T cell IL-17A responders to EBNA1 compared to PfSEA-1A (p = 0.0004). Conversely, the frequency of CD8+ T cell responders were more variable when comparing EBNA1 to PfSEA-1A within each group of children (Figure 1B). All Nandi children had IL-10 CD8+ T cell responses to PfSEA-1A, in contrast to only half of them responding to EBNA1 (p < 0.0001) even though they were all EBV seropositive, whereas more Nandi children had IFN-γ CD8+ T cell responses to EBNA1 compared to PfSEA-1A (p = 0.0006). Kisumu children had more IFN-γ CD8+ T cell responders to PfSEA-1A compared to EBNA1 (p = 0.01), but the eBL children had similarly low levels to both antigens. Kisumu children were also more likely to have IL-17A CD8+ T cell responses to PfSEA-1A compared to EBNA1 (p < 0.0001) in sharp contrast to eBL children who had more responders to EBNA1 and few to PfSEA-1A (p < 0.0001). The IL-10 CD8+ T cell responses to EBNA1 and PfSEA-1A also had an inverse detection with fewer Kisumu responders to PfSEA-1A compared to EBNA1 (p = 0.0009), yet more eBL responders to PfSEA-1A compared to EBNA1 (p = 0.0002). Taken together, if eBL children developed immune modifications to compensate for the loss of IFN-γ responses to EBNA1, IL-17A could play a role that warrants further exploration. In addition, eBL children seem to have developed more IL-17A expressing CD4+ and CD8+ T cells compared to Kisumu children who also resided in malaria holoendemic areas.
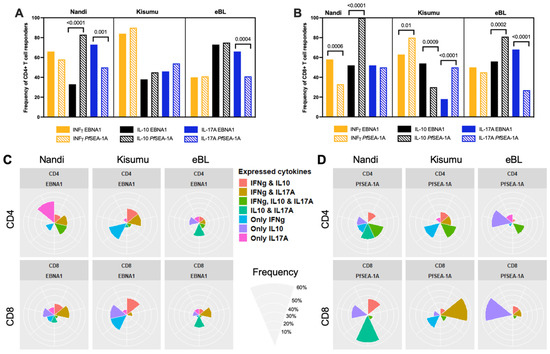
Figure 1.
Comparison of cytokine responses for Nandi, Kisumu and eBL children to EBNA1 and PfSEA-1A. Cytokine (IFN-γ, IL-10 or IL-17A) frequencies to EBNA1 and PfSEA-1A were compared for (A) CD4+ or (B) CD8+ T cells within each study group, χ2 test was applied and p values are indicated. Radar/flower plots represent the cytokine profile (frequency from 0 to 50% for Nandi and Kisumu groups and from 0 to 60% for eBL) within individuals from each group of children from CD4+ and/or CD8+ after (C) EBNA1 or (D) PfSEA-1A stimulation. Each petal represents a different combination of cytokine (IFN-γ, IL-10, IL-17A) expression found within individual children.
To determine which combinations of cytokine responses to EBNA1 were found within the same individuals and if they differed by malaria exposure or eBL diagnosis, results were summarized by flower plots (Figure 1C). The colored petals show the different combinations of cytokines being expressed and the size of the petal illustrates the frequency of finding those cytokines (alone or together) within individual children. Most Nandi children (33.3%) only had IL-17A expressing EBNA1-specific CD4+ T cells, yet 20% had both IFN-γ and IL-17A, and another 20% had both IL-10 and IL-17A expressing CD4+ T cells after EBNA1 stimulation. CD8+ T cell responses were more heterogeneous with most Nandi children (20%) expressing both IFN-γ and IL-17A to EBNA1. In contrast, 30% of Kisumu children had only IFN-γ expressing EBNA-1-specific CD4+ T cells, with 23.5% of children producing both IFN-γ and IL-17A, or IFN-γ and IL-10. However, their CD8+ T cell responses were equally prevalent (27.7%) as one of three phenotypes: IFN-γ, IL-10, or expressing both IFN-γ and IL-10. This might reflect a transition from pro-inflammatory to anti-inflammatory CD8+ T cell populations depending on the cumulative number of malaria infections experienced by each child. Of the relatively fewer eBL children who had EBNA1-specific CD4+ T cell responses, 27% expressed IL-10 and IL-17A and 20% expressed only IL-10, with lower frequencies of responders for other cytokine combinations. As for their CD8+ T cells, 25% of eBL children expressed either IL-10 and IL-17A, or IFN-γ and IL-17A to EBNA1, with some minor exceptions. In summary, it would appear that Kisumu children developed more IFN-γ and IL-10 CD4+ and CD8+ T cell responses to EBNA1 compared to Nandi and eBL children, suggesting their combined importance in limiting EBV-associated oncogenesis. In contrast, adult memory responses to EBNA1 included IFN-γ, IL-10 expressing cells, but also cells expressing IL-17A and those with a dual expression of IL-10 and IFN-γ (Figure S2).
Figure 1D shows the combinations of cytokine responses to PfSEA-1A found within the same individuals and summarized by the flower plots. CD4+ and CD8+ T cell responses to this malaria antigen were also found to be heterogeneous within individuals across our study populations. Most of the Nandi children generated IL-10 in combination with CD4+ and CD8+ T cells that expressed IL-17A and/or IFN-γ to PfSEA-1A, whereas Kisumu children relied on CD4+ T cells to express IFN-γ alone or in combination with IL-10 and/or IL-17A, and CD8+ T cell populations that expressed IFN-γ or IL-17A. These heterogeneous cytokine profiles were in sharp contrast to the eBL children who had predominantly CD4+ and CD8+ T cells expressing only IL-10 to PfSEA-1A. Together, these findings reinforce the premise that an immune suppressive environment against malaria developed for children diagnosed with eBL, and that their IL-10 production by T cells was elevated against both EBNA1 and PfSEA-1A.
3.2. Magnitude of Cytokine Responses to EBNA1 and PfSEA-1A
Even though there were significant differences in the frequency of IFN-γ expressing CD4+ T cell responders to EBNA1 across the study groups, we did not observe differences in the magnitude of EBNA1-specific IFN-γ expressing CD4+ or CD8+ T cells (Figure 2A) for those with responses. To determine if these differences were specific to EBNA1, we tested memory responses to other EBV latent antigens. No differences in IFN-γ responses were observed for EBNA2 and EBNA3A demonstrating that there was no immunodominant latent antigen T cell response within our study populations (Figure S3A,B, respectively).
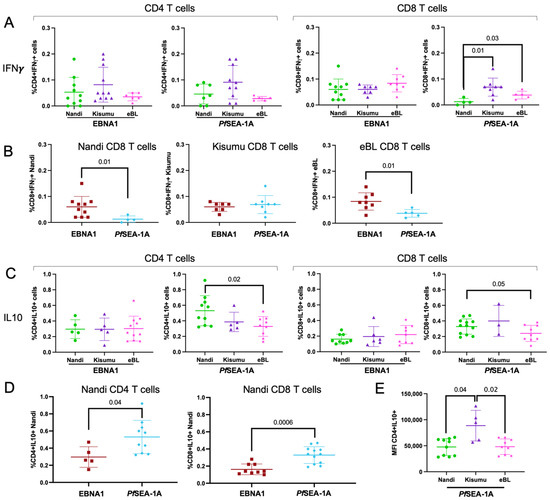
Figure 2.
Comparison of cytokine-expressing CD4+ and CD8+ T cell frequencies across study groups. (A) EBNA1 and PfSEA-1A responses were measured as a percentage of total CD4+ and CD8+ cells expressing IFN-γ and compared across the different groups of children: Nandi (green circle), Kisumu (purple triangle) and eBL (pink star). (B) IFN-γ+ CD8+ T cell responses to EBNA1 (red square) and PfSEA-1A (blue diamond) were compared within Nandi (left), Kisumu (middle), and eBL (right) children. (C) EBNA1 and PfSEA-1A responses were measured as the percentage of total CD4+ and CD8+ T cells expressing IL-10 and compared across the different groups of children: Nandi, Kisumu and eBL. (D) IL-10 response to EBNA1 and PfSEA-1A were compared for CD4+ (left) and CD8+ (right) T cells from Nandi children. (E) Comparison of IL-10+ expression intensity from CD4+ T cells (geometric mean fluorescent intensity, MFI) after PfSEA-1A stimulation across groups of children: Nandi, Kisumu and eBL. Mann–Whitney or Wilcoxon p-values are indicated as well as the mean with standard deviation (SD).
For PfSEA-1A, we observed significantly fewer IFN-γ expressing CD8+ T cells within Nandi children compared to the Kisumu and eBL children (p = 0.01 and p = 0.03, respectively, Figure 2A). This was expected as Kisumu and eBL children resided in high Pf-malaria transmission areas whereas Nandi children had fewer opportunities to develop memory T cell responses to a malaria antigen. However, all children in our study (Nandi, Kisumu and eBL) were EBV seropositive (Table S1) and should have developed T cell immunity to EBNA1. Therefore, it was not surprising to observe significantly higher frequencies of EBNA1-specific IFN-γ expressing CD8+ T cells compared to PfSEA-1A for Nandi children (p = 0.01, Figure 2B), whereas Kisumu children had similar frequencies of responses for both antigens. In contrast, eBL children had significantly higher EBNA1-specific IFN-γ-producing CD8+ T cells compared to PfSEA-1A (p = 0.01, Figure 2B) which suggests a loss of CD8+ T cells which could help clear malaria infections.
Consistent with the frequency of EBNA1-specific IL-10 responders, we did not observe any differences in the frequency of IL-10 expressing CD4+ and CD8+ T cells across study groups (Figure 2C). However, we found significantly more IL-10-producing CD4+ T cells to PfSEA-1A for Nandi compared to eBL children (p = 0.02, Figure 2C), with a similar trend for CD8+ T cells (p = 0.05). We also observed significantly higher frequencies of IL-10 expressing CD4+ and CD8+ T cells to PfSEA-1A compared to EBNA1 for Nandi children (p = 0.04 and p = 0.0006, respectively, Figure 2D). IL-10 responses to these antigens were not different for Kisumu or eBL children (Figure S4). Finally, even though Kisumu children did not have significantly higher frequencies of IL-10 expressing CD4+ T cells, they had significantly more cytokine production per T cell compared to Nandi and eBL children (p = 0.04 and p = 0.02, respectively, Figure 2E). Similar comparisons were made for IL-17A responses but no differences were observed (Supplemental Figure S5A,B). Interestingly, CD4+ and CD8+ T cell responses from eBL children had a lower IL-17A expression to PfSEA-1A (Figure S5C,D); however, the overall response rates were too infrequent to achieve significance.
These results demonstrate that the frequency of EBNA1-specific IFN-γ expressing CD4+ and CD8+ T cells did not differ between eBL and healthy children who developed these T cell responses, suggesting that the IFN-γ deficiency previously reported for eBL children [18] might not be due to the lack of cytotoxic T cells. These findings also suggest that Nandi children who experience acute malaria infections develop robust anti-inflammatory T cell responses to PfSEA-1A mediated by IL-10, in contrast to children who experience malaria as a chronic infection.
3.3. Mono- and Poly-Functional IFN-γ, IL-10 and IL-17A T Cell Responses to EBNA1 and PfSEA-1A
Polyfunctional T cells have been described for other chronic infections and shown to correlate with T cell efficiency and immune regulation [30,46]. However, we found EBNA1-specific CD4+ and CD8+ T cells to be mostly monofunctional, expressing IL-10 for our three groups of children (Figure 3A,B), thus supporting their role as antiviral, immunosuppressive mediators [47]. Interestingly, IL-17A expressing monofunctional CD4+ and CD8+ T cells were present in all three groups of children (Figure 3C,D) with a slightly higher trend for eBL compared to Kisumu children (p = 0.04, Figure 3C). We also observed a significantly higher frequency of double positive IFN-γ+IL-10+ EBNA1-specific CD4+ T cells for Kisumu compared to eBL children (p = 0.02, Figure 3C) identifying a T cell population engaged in immunity to EBV not found in children with eBL. In contrast, no significant differences were observed within EBNA1-specific CD8+ T cell profiles across our groups of children despite a trend of higher frequencies of EBNA1-specific CD8+ T cells expressing IL-17A from eBL compared to Kisumu children.
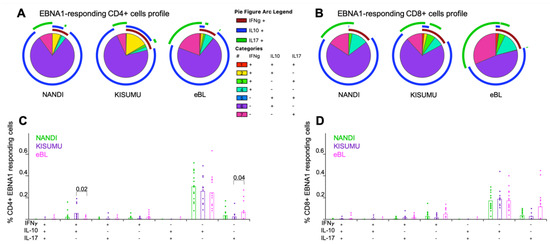
Figure 3.
Mono- and poly-functional T cell profiles to EBNA1. Functionality of cells after EBNA1 stimulation were compared across study groups. Each pie slice represents the type of cytokine expression from (A) CD4+ and (B) CD8+ T cells; arcs show co-cytokine expression. The frequencies of each type of cytokine-expressing T cells were compared across study groups (Nandi in green, Kisumu in purple and eBL in pink) after EBNA1 stimulation for (C) CD4+ and (D) CD8+ T cells. Bar plots show the mean for each group of children and each dot represents an individual. Wilcoxon Rank Sum Test p values < 0.05 are indicated.
Monofunctional PfSEA-1A-specific IL-10-producing CD4+ and CD8+ T cells were the predominant phenotype for Nandi children compared to eBL children (p = 0.01 for both, Figure 4A,B). Polyfunctional T cells were observed at higher frequencies for Kisumu children, with more double positive IFN-γ+IL-10+ PfSEA-1A-specific CD4+ T cells compared to eBL and Nandi children (Figure 4C, p = 0.003 and p = 0.02, respectively); and more IFN-γ+IL-17A+ PfSEA-1A-specific CD8+ T cells compared to eBL children (Figure 4D, p = 0.03). In contrast, eBL children had a higher frequency of IFN-γ+-PfSEA-1A-specific CD8+ T cells compared to Nandi children with low malaria exposure. Together, these data suggest that separate T cell populations serve distinct roles in fighting infection versus limiting immunopathology.
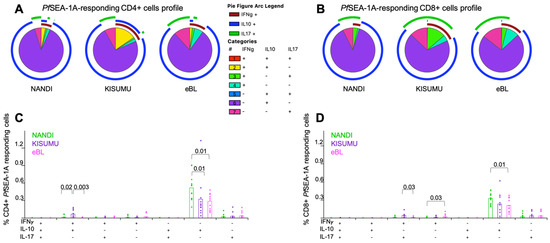
Figure 4.
Mono- and poly-functional T cell profiles to PfSEA-1A. Functionality of cells after PfSEA-1A stimulation were compared across study groups. Each pie slice represents the type of cytokine expression from (A) CD4+ and (B) CD8+ T cells; arcs show co-cytokine expression. The frequencies of each type of cytokine-expressing T cells were compared across study groups (Nandi in green, Kisumu in purple and eBL in pink) after PfSEA-1A stimulation for (C) CD4+ and (D) CD8+ T cells. Bar plots show the mean for each group of children and each dot represents an individual. Wilcoxon Rank Sum Test p values < 0.05 are indicated.
3.4. EBNA-1 and PfSEA-1A Specific T Cell Subsets
We next determined the effector memory phenotypes of cytokine-producing T cells characterized based on their CD45RA and CCR7 expression and compared subset proportions across our three groups of children and to African adults. EBNA1-specific IFN-γ-producing CD4+ T cells from both children and adults were predominantly TNAIVE-LIKE cells with smaller contributions from the other subsets (Figure 5A). Kisumu children displayed a significantly different distribution of EBNA1-specific IFN-γ CD4+ T cell subsets compared to Nandi, eBL and adults (p = 0.005, p = 0.0003 and p < 0.0001, respectively) with fewer TEMRA and TEM and no TCM. In contrast, EBNA1-specific, IL-10-producing CD4+ T cells (Figure 5A) had a larger proportion of TEMRA across our study groups, except for the eBL children who had significantly more TNAIVE-LIKE cells compared to Kisumu children and adults (p = 0.03). Additionally, EBNA1-specific, IFN-γ-producing CD8+ T cells (Figure 5B) were derived from both TNAIVE-LIKE and TEMRA cell subsets, and displayed significant differences in relative proportions between the Nandi children who had more TCM compared to the Kisumu children, eBL and adult groups, (p = 0.001, p = 0.001 and p < 0.0001, respectively). Interestingly, we observed that IL-10-producing CD8+ T cells (Figure 5B) were mostly TEMRA and TNAIVE-LIKE cell subsets, except for the eBL children who had significantly more TNAIVE-LIKE cells compared to the Nandi children and adults (p = 0.0005 and p = 0.001, respectively) suggesting an immature EBNA1-specific T cell signature for children with this EBV-associated cancer. Further study is needed to determine if this condition precedes oncogenesis or if it is an indication of recent T cell turnover.
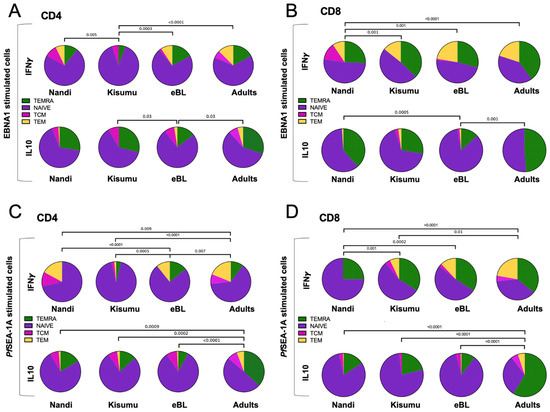
Figure 5.
Comparison of EBNA1- or PfSEA-1A- cytokine-expressing T cell subsets across groups of individuals. The mean proportion (pie slice) of IFN-γ or IL-10 expressing (A) CD4+ and (B) CD8+ after EBNA1 stimulation; or IFN-γ or IL-10 expressing (C) CD4+ and (D) CD8+ after PfSEA-1A stimulation were compared across our study groups (Nandi, Kisumu, eBL, and Adults). T cell subsets were defined by CD45RA and CCR7 expression: TEMRA (green, CD45RA+CCR7−), TNAIVE-LIKE (purple, CD45RA+CCR7+), TCM (pink, CD45RA−CCR7+) and TEM (yellow, CD45RA−CCR7−). χ2 test was applied and p values are indicated.
As a basis for comparison to persistent EBV latent antigen stimulation, we examined the T cell memory phenotypes to a malaria antigen, which tends to cause an acute infection for Nandi children but a repeated/chronic infection for Kisumu and eBL children. In general, the composition of CD4+ T cell subsets expressing IFN-γ and IL-10 to PfSEA-1A were significantly more mature for adults (TEMRA, TCM and TEM subsets) compared to Nandi, Kisumu and eBL children (Figure 5C). However, eBL children had more PfSEA-1A-specific IFN-γ CD4+ TEMRA and fewer TCM compared to Nandi children (p < 0.0001, Figure 5C) and more TEMRA and TEM compared to Kisumu children whose responses were dominated by the TNAIVE-LIKE cell subset (p = 0.0005). In contrast, PfSEA-1A-specific IFN-γ CD8+ T cells subset proportions were significantly different between malaria-exposed and non-exposed individuals. Nandi children had mostly TNAIVE-LIKE cells with a complete absence of TCM and TEM subsets compared to Kisumu, eBL and adults who resided in malaria holoendemic areas (Figure 5D). Interestingly, we observed no statistical differences for PfSEA-1A-specific IFN-γ CD8+ T subsets between eBL children and adults, whereas Kisumu children had more TNAIVE-LIKE and fewer TEM cell subsets compared to adults (p = 0.01, Figure 5D). This suggests that eBL children had a more “adult-like” T cell memory repertoire to malaria which could have resulted from a greater cumulative burden of malaria compared to their healthy counterparts. In contrast, differences for PfSEA-1A-specific IL-10-producing CD8+ T cell subsets were only apparent when comparing adults to children. IL-10 responses in children were dominated by TNAIVE-LIKE cells, whereas an adult response was governed by TEMRA cells (p < 0.0001, Figure 5D).
Overall, our findings suggest that the compositions of EBNA1 and PfSEA-1A-specific IFN-γ expressing T cell memory subsets were influenced by intensity of malaria exposure, whereas T cell subsets expressing IL-10 were dependent on an eBL diagnosis or age.
3.5. PD-1 Expression, as a Marker of Activation or Exhaustion
To determine if T cell exhaustion could account for the differences in the phenotypes and functions described above, we measured the PD-1 expression on CD4+ and CD8+ T cell subsets within our study groups. PD-1 is a classical T cell exhaustion marker but is also an indicator of the activation and functional adaptation after chronic antigen-engagement [48]. We found that EBNA1 stimulation significantly increased the percentage of PD-1 expression on CD4+ and CD8+ cells for Nandi children compared to their resting cells (p = 0.01 and p = 0.002, respectively, Figure 6A,B). Moreover, PD-1 expression, in response to EBNA1, was significantly higher compared to PfSEA-1A-stimulated cells (p = 0.005, Figure 6B), as expected for children with acute exposure to malaria. However, both EBNA1 and PfSEA-1A stimulation displayed significant increases in PD-1 expression for CD4+ T cells for Kisumu children compared to their resting cells, (p = 0.003 and p = 0.0005, respectively), and an equal level of induction was found between these two simulation conditions (Figure 6C). Interestingly, PfSEA-1A significantly induced PD-1+ expression in CD8+ T cells for Kisumu children compared to resting and EBNA1 stimulation levels (Figure 6D). For the eBL children, only PfSEA-1A stimulation increased the percentage of PD-1+CD4+ cells compared to the resting cells (p = 0.002, Figure 6E). However, PD-1 expression was the same as upon EBNA1 stimulation, suggesting a sustained expression for CD4+ T cells, a sign of exhaustion as opposed to the transient activation for children with eBL. The PD-1 expression pattern for CD8+ T cells was similar between eBL and Kisumu children with higher expressions after PfSEA-1A stimulation, compared to both the resting state and EBNA1 stimulation (p = 0.003 and p = 0.0005, respectively, Figure 6F). In addition, we found significantly higher overall PD-1 expression on CD4+ but not CD8+ T cells (Figure 7A,B) for eBL, compared to Nandi children after EBNA1 stimulation (p = 0.004). In a striking contrast to the cytokine-producing T cells, the exhausted CD4+ T cells were predominantly found in the TEM subset across study groups (including adults) with slightly more PD-1 expressed by TNAIVE-LIKE cells in Kisumu compared to eBL children and adults (p = 0.04 and p = 0.01, respectively, Figure 7C). However, PD-1 was highly expressed on CD8+ TEMRA, TEM and TNAIVE-LIKE cells among Nandi, Kisumu and eBL children which differed from adults, who had fewer PD-1 expressing TNAIVE-LIKE cells (p = 0.009, p < 0.0001 and p = 0.003, respectively, Figure 7C).
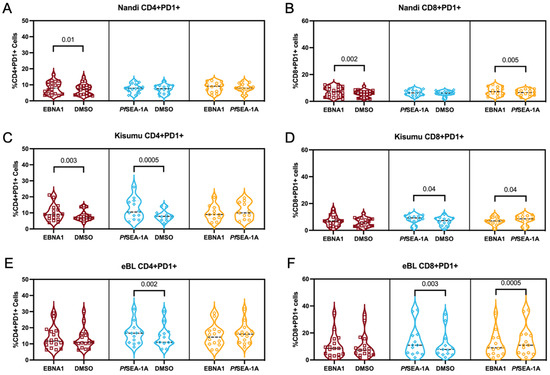
Figure 6.
PD-1 expression on stimulated vs. unstimulated CD4+ and CD8+ T cells. Percentage of CD4+PD-1+ cells from (A) Nandi, (C) Kisumu and (E) eBL children after EBNA1 and PfSEA-1A stimulation compared to unstimulated, DMSO control. Percentage of CD8+PD-1+ cells from (B) Nandi, (D) Kisumu and (F) eBL children after EBNA1 and PfSEA-1A stimulation compared to unstimulated, DMSO control. Wilcoxon two-tailed and matched-pairs test p values are indicated.
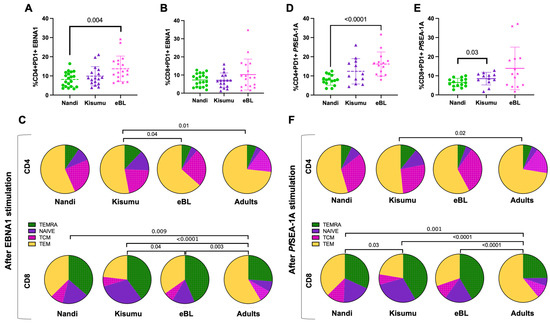
Figure 7.
Comparison of PD-1 expressing CD4+ and CD8+ T cell frequencies across study groups. Comparison of PD-1 expressing (A) CD4+ and (B) CD8+ T cells after EBNA1 stimulation between groups of children (Nandi green circles; Kisumu purple triangles and eBL pink stars) using Mann–Whitney test. (C) The mean proportion (pie slice) of PD-1-expressing CD4+ and CD8+ after EBNA1 stimulation was compared across study groups using χ2 test. Comparison of PD-1 expressing (D) CD4+ and (E) CD8+ T cells after PfSEA-1A stimulation between groups of children (Nandi green circles; Kisumu purple triangles and eBL pink starts) using Mann–Whitney test. (F) The mean proportion (pie slice) of PD-1-expressing CD4+ and CD8+ after PfSEA-1A stimulation was compared across study groups using χ2 test. T cell subsets were defined by CD45RA and CCR7 expressions: TEMRA (green, CD45RA+CCR7−), TNAIVE-LIKE (purple, CD45RA+CCR7+), TCM (pink, CD45RA−CCR7+) and TEM (yellow, CD45RA−CCR7−).
We observed similar patterns for PfSEA-1A stimulation with an overall increase in PD-1 expression in CD4+ T cells for eBL patients compared to Nandi children (p < 0.0001, Figure 7D), and a higher PD-1 expression in CD8+ T cells from Kisumu compared to Nandi children (p = 0.03, Figure 7E). This result reinforces the notion that PD-1 expression is context-dependent and might be a marker of activation rather than exhaustion. The CD4+ T cells expressing PD-1 after PfSEA-1A stimulation were also predominantly TEM and TCM cells (Figure 7F), but showed significant differences when comparing Kisumu children to adults (p = 0.02). More differences were observed for CD8+ T cells with adults expressing significantly more PD-1 for the TEM subset compared to children who had more PD-1 expression for TEMRA or TNAIVE-LIKE cells. Of note, PD-1 expressing CD8+ T cell subset proportions were also different between the Kisumu and Nandi children after PfSEA-1A stimulation (p = 0.03).
Finally, we found that the frequency of PD-1+CD4+ T cells in response to EBNA1 was higher for Kenyan adults compared to healthy Nandi and Kisumu children (p = 0.002 and p = 0.01, respectively, Supplemental Figure S6A). Interestingly, the percentage of PD-1+CD4+ T cells was not different between eBL children and adults, but differed from Nandi children (p = 0.004), which suggests that eBL children had exhausted CD4+ T cells due to more encounters with EBNA1. Our hypothesis that prolonged malaria exposure (or untreated chronic infections) as a risk factor for eBL is consistent with our finding that the frequency of exhausted CD4+ T cells to PfSEA1-1A were similar between eBL children and Kenyan adults, and were significantly higher compared to healthy Nandi children (p < 0.0001 and p = 0.0004, respectively, Figure S6A). Nandi and Kisumu children also had lower frequencies of PD-1+CD8+ T cells in response to EBNA1 compared to adults, (p = 0.01 and p = 0.02, respectively, Figure S6B), with eBL children being similar to adults for both EBNA1 and PfSEA1-A1. We found higher PD-1+CD8+ T cell frequencies in adults compared to both Nandi and Kisumu children (p < 0.0001 and p = 0.002, respectively, Figure S6B), which also differed based on their histories of malaria exposure (p = 0.03).
4. Discussion
The children diagnosed with eBL are deficient in T cell-mediated immune responses specific for EBNA1, the sole EBV antigen consistently expressed in this B cell tumor [18], and the only viral protein expressed during both lytic and latent cycles. Researchers postulate that the cumulative exposure to Pf malaria infections throughout early childhood leads to EBV-specific immune deficiencies that put these children at risk of eBL oncogenesis [3]. However, alternative cytokine profiles and the phenotypes of cytokine-producing cells have never been investigated in this context. Here, we characterized EBNA1-specific IFN-γ, IL-10 and IL-17A T cell responses in eBL children compared to healthy children residing in malaria holoendemic (Kisumu) and hypoendemic (Nandi) regions of Kenya to gain further insights into eBL immunopathology. We found that EBNA1-specific CD4+ T cells expressing IFN-γ were mostly TNAIVE-LIKE cells and were less frequent in children with eBL compared to healthy children, yet EBNA1-specific T cell cytokine profiles were highly heterogeneous within individuals and across the study groups. Kisumu children had both CD4+ and CD8+ EBNA1-specific T cells expressing either IFN-γ or IL-10 alone or in combination, compared to predominantly IL-17A expressing CD4+ T cells for Nandi children. Kisumu children were also more likely to have polyfunctional CD4+ T cells expressing IFN-γ and IL-10, whereas eBL children had more monofunctional IL-17A producing CD4+ T cells to EBNA1. The phenotype of cytokine expressing CD8+ T cell subsets was largely composed of the TNAIVE-LIKE and TEMRA subsets, with most differences in proportions associated with immune maturation. The studies of adolescents with infectious mononucleosis predominantly generated EBV-specific CD4+ TEM and TCM responses [49,50], consistent with our findings that the quality of EBV-specific T cell immunity is dependent on age. Interestingly, Nandi children had more CD8+ TCM cells compared to Kisumu and eBL children suggesting a difference in the EBNA1-specific memory T cell establishment relative to the acute versus chronic malaria infections.
In our seminal study, we reported a lower frequency of IFN-γ responders to EBNA1 within a group of eBL children compared to Kisumu and Nandi children, in addition to having fewer cytokine-producing cells, as measured by ELISPOT [18]. In this study, using a customized flow cytometry assay, we confirmed the lower frequency of responders within another group of children diagnosed with eBL; however, the frequency of IFN-γ-producing CD8+ T cells did not differ compared to healthy children. Other studies have demonstrated the importance of not only cytotoxic T cells but also natural killer (NK) cells in EBV control and the prevention of tumorigenesis [51,52]. We previously demonstrated a deficiency in IFN-γ production by NK cells for eBL compared to healthy children [42]. These NK cells were characterized as dysfunctional ‘chronic-infection induced’ cells in children exposed to holoendemic malaria as well as in those diagnosed with eBL [42]. Future studies are needed to explore malaria-induced mechanisms leading to NK cell dysfunction as a component of eBL immunopathology. A limitation of this study was not having enough cells to measure NKT cells, gamma-delta T cells as other non-classical T cell sources of IFN-γ, which were associated with malaria and eBL in other studies [53,54,55].
Overall, we observed the robust production of IL-10 from monofunctional CD4+ and CD8+ T cells (predominantly TNAIVE-LIKE and TEMRA subsets) for all groups of children in response to both EBNA1 and PfSEA-1A. IL-10 is an anti-inflammatory cytokine that down-regulates IFN-γ [56] and has been associated with immune suppression during severe malaria in children [57]. It is not surprising that these children developed an anti-inflammatory response after the prolonged exposure to this persistent EBV infection and that these cytokines may be generated by different T cell subsets compared to adults. IL-10 responses to EBV and malaria antigen were comparable for Kisumu and eBL children; however, Nandi children had significantly higher CD4+ and CD8+ T cell IL-10 responses to malaria (typically an acute infection in this study population) compared to EBNA1. Several types of CD4+ T cells are known to produce IL-10, such as regulatory T cells (Tregs) which can help reduce inflammation, and thereby reduce host tissue damage during chronic diseases. However, Tregs have been associated with poor outcomes for eBL patients [58], and here we observed the same trend with more EBNA1-specific CD4+ T cells (but not CD8+ T cells) producing IL-10 in eBL non-survivor compared to survivors (p = 0.04, Figure S7A,B). Morales et al. also observed a higher percentage of circulating IL10+CD4+ T cells in EBV-positive Hodgkin Lymphoma (HL) patients compared to HL patients without EBV and healthy controls [59]. However, they classified these IL10+CD4+ T cells as Tr1 (T regulatory type 1) using double staining for ITGA2 and ITGB2 [59]. Future studies are needed to assess a panel of IL-10-producing cell types [60] that may contribute to the immuno-regulatory environment in children infected with malaria and diagnosed with eBL.
Another mechanism used to down-regulate T cell function is PD-1. However, PD-1 can be considered as both a marker of exhaustion as well as activation depending on the type of infection [18,30]. Our study showed that phenotype of antigen-specific, cytokine-producing T cells were not the same subsets as those expressing PD-1 and that PD-1 expression was context-dependent. Children with eBL showed the most signs of T cell exhaustion; whereas, for healthy children, EBNA1 appeared to activate CD4+ T cells. Our results reinforce the theory that chronic Pf malaria infections and higher EBV loads could lead to persistent T cell activation and dysfunction. These qualitative differences and heterogeneity in T cell immunity to EBNA1 within individuals and across study groups provide more clues in the causal pathway of eBL. Studies are underway exploring this concept, including more markers and transcription factors that regulate T cell activation and exhaustion [61].
To determine the specificity of immune responses to EBNA1 within our study populations, we performed parallel investigations into the responses to a malaria vaccine candidate, PfSEA-1A. Antibodies against PfSEA-1A appear protective against reinfection and decrease parasite replication in vitro [38]. However, naturally acquired T cell responses to this antigen have not yet been characterized. We observed that T cells from Nandi children expressed IL-10, IFN-γ and IL-17A to EBNA1; whereas, they almost exclusively expressed IL-10 to PfSEA-1A. Interestingly, both double positive IFN-γ+IL-10+ and IFN-γ+IL-17A+ cells were involved in T cell responses to EBNA1 and PfSEA-1A for Kisumu children. In contrast, eBL children had more monofunctional EBNA1-specific T cell responses, whereas the variety of responses decreased for PfSEA-1A, impaired by an overall higher frequency of PD-1. Figure 8 illustrates the heterogeneity found for EBNA1 and PfSEA-1A cytokine responses from children with divergent malaria infection histories compared to children diagnosed with eBL. Deconvoluting human T cell adaptations, driven by the intensity of malaria exposure combined with high EBV loads, supports the prevailing theory of eBL pathogenesis that involves a multifaceted degradation of immune control over EBV-infected B cells.
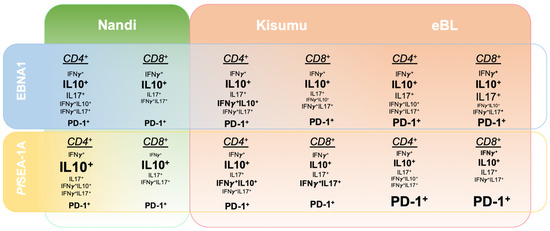
Figure 8.
Summary of relative cytokine expression patterns to EBNA1 compared to a malaria antigen, PfSEA-1A across three groups of children: Nandi, low malaria exposure; Kisumu, high malaria exposure; and eBL cancer patients, high malaria exposure. The summary is based on the 2D gating strategy from FlowJo and unbiased Boolean gating in SPICE.
There were several limitations to our study. Investigating human T cell immunity to most infections is challenging due to low frequencies (less than 2%) of cells captured in peripheral blood samples. However, the frequencies of EBV-specific T cells in our study were comparable to other study groups [20]. Our sample size was insufficient to test the association between all the different T cell responses we found and eBL survival. We also made assumptions about the histories of malaria exposure for our study populations, using study site as an epidemiologic surrogate. However, based on our previously published longitudinal studies, we are confident that cumulative malaria exposures and EBV loads were significantly different between Kisumu and Nandi children [8,40,62]. It will be informative to validate our results with another cohort of children from Kenya or from another sub-Saharan country where eBL occurs.
5. Conclusions
This study demonstrates that EBNA1-specific T cell responses are highly heterogeneous and are shaped by malaria and EBV-associated immune regulatory adaptations. IL-10-producing EBNA1-specific T cells that were enriched in children with eBL might be involved in preventing efficient immune control of this EBV-associated tumor. Understanding the devolution of immune responses to EBV during malaria co-infections will improve our ability to prevent or cure eBL.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/cancers13215375/s1, Table S1: Population characteristics, Figure S1: Flow cytometry gating strategy and T cell subsets, Figure S2: Adult T cell cytokine profile in response to EBNA1 and PfSEA-1A, Figure S3: Comparison of IFN-γ+ T cell responses between EBNA1, EBNA2 and EBNA3A stimulation for each group of children, Figure S4: No differences were observed in IL-10 expression between EBNA1 and PfSEA-1A for Kisumu or eBL children, Figure S5: IL-17 responses to EBNA1 and PfSEA-1A from CD4+ and CD8+ T cells within each group of children, Figure S6: Adults had significantly higher CD4+PD-1+ T cells compared to Nandi and Kisumu children but not compared to eBL children, Figure S7: EBNA1-specific IL-10+CD4+ and CD8+ T cell frequencies for eBL survivors compared to non-survivors.
Author Contributions
D.H.M., J.M.O. and A.M.M. conceived the study. J.F., J.A.O. and J.M.O. collected the blood samples. C.S.F., P.S.L. performed the flow experiments. C.S.F., J.D.K., L.J.B., C.M. and A.M.M. analyzed the data. C.S.F. and A.M.M. wrote the manuscript. Funding was acquired by A.M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Institutes of Health grants R01 CA189806 and R01 CA134051 from the National Cancer Institute (A.M.M.) and R01 AI127699 (J.D.K.).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board at the University of Massachusetts Chan Medical School, Worcester, MA, USA (H-00004587) and the Scientific and Ethics Review Unit (SERU 2844) at the Kenya Medical Research Institute, Nairobi, Kenya.
Informed Consent Statement
Informed consent was obtained from all subjects or parents of minor participants involved in the study.
Data Availability Statement
The data presented in this study are openly available upon request.
Acknowledgments
The authors thank the children and their families who participated in this study. We also thank Peter Oluoch for his R program expertise. This manuscript was published with the permission of the KEMRI. The graphical abstract was created with BioRender.com (accessed on 13 September 2021).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Parkin, D.M.; Bray, F.; Ferlay, J.; Jemal, A. Cancer in Africa 2012. Cancer Epidemiol. Biomarkers Prev. 2014, 23, 953–966. [Google Scholar] [CrossRef]
- Stefan, C.; Bray, F.; Ferlay, J.; Liu, B.; Maxwell Parkin, D. Cancer of Childhood in Sub-Saharan Africa. Ecancermedicalscience 2017, 11. [Google Scholar] [CrossRef]
- Moormann, A.M.; Bailey, J.A. Malaria—How This Parasitic Infection Aids and Abets EBV-Associated Burkitt Lymphomagenesis. Curr. Opin. Virol. 2016, 20, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Burkitt, D. A Tumour Safari in East and Central Africa. Br. J. Cancer 1962, 16, 379–386. [Google Scholar] [CrossRef]
- De-Thé, G. The Epidemiology of Burkitt’s Lymphoma: Evidence for a Causal Association with Epstein-Barr Virus. Epidemiol. Rev. 1979, 1, 32–54. [Google Scholar] [CrossRef]
- Moormann, A.M.; Chelimo, K.; Sumba, O.P.; Lutzke, M.L.; Ploutz-Snyder, R.; Newton, D.; Kazura, J.; Rochford, R. Exposure to Holoendemic Malaria Results in Elevated Epstein-Barr Virus Loads in Children. J. Infect. Dis. 2005, 191, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Piriou, E.; Asito, A.S.; Sumba, P.O.; Fiore, N.; Middeldorp, J.M.; Moormann, A.M.; Ploutz-Snyder, R.; Rochford, R. Early Age at Time of Primary Epstein-Barr Virus Infection Results in Poorly Controlled Viral Infection in Infants from Western Kenya: Clues to the Etiology of Endemic Burkitt Lymphoma. J. Infect. Dis. 2012, 205, 906–913. [Google Scholar] [CrossRef]
- Reynaldi, A.; Schlub, T.E.; Chelimo, K.; Sumba, P.O.; Piriou, E.; Ogolla, S.; Moormann, A.M.; Rochford, R.; Davenport, M.P. Impact of Plasmodium Falciparum Coinfection on Longitudinal Epstein-Barr Virus Kinetics in Kenyan Children. J. Infect. Dis. 2016, 213, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Snider, C.J.; Cole, S.R.; Chelimo, K.; Sumba, P.O.; Macdonald, P.D.M.; John, C.C.; Meshnick, S.R.; Moormann, A.M. Recurrent Plasmodium Falciparum Malaria Infections in Kenyan Children Diminish T-Cell Immunity to Epstein Barr Virus Lytic but Not Latent Antigens. PLoS ONE 2012, 7, e31753. [Google Scholar] [CrossRef]
- Moormann, A.M.; Chelimo, K.; Sumba, P.O.; Tisch, D.J.; Rochford, R.; Kazura, J.W. Exposure to Holoendemic Malaria Results in Suppression of Epstein-Barr Virus-Specific T Cell Immunosurveillance in Kenyan Children. J. Infect. Dis. 2007, 195, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Kaymaz, Y.; Oduor, C.I.; Yu, H.; Otieno, J.A.; Ong’echa, J.M.; Moormann, A.M.; Bailey, J.A. Comprehensive Transcriptome and Mutational Profiling of Endemic Burkitt Lymphoma Reveals EBV Type-Specific Differences. Mol. Cancer Res. 2017, 15, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Grande, B.M.; Gerhard, D.S.; Jiang, A.; Griner, N.B.; Abramson, J.S.; Alexander, T.B.; Allen, H.; Ayers, L.W.; Bethony, J.M.; Bhatia, K.; et al. Genome-Wide Discovery of Somatic Coding and Noncoding Mutations in Pediatric Endemic and Sporadic Burkitt Lymphoma. Blood 2019, 133, 1313–1324. [Google Scholar] [CrossRef] [PubMed]
- Panea, R.I.; Love, C.L.; Shingleton, J.R.; Reddy, A.; Bailey, J.A.; Moormann, A.M.; Otieno, J.A.; Ong’echa, J.M.; Oduor, C.I.; Schroeder, K.M.S.; et al. The Whole-Genome Landscape of Burkitt Lymphoma Subtypes. Blood 2019, 134, 1598–1607. [Google Scholar] [CrossRef]
- Roca-Feltrer, A.; Carneiro, I.; Armstrong, J.R. Estimates of the Burden of Malaria Morbidity in Africa in Children under the Age of 5 Years. Trop. Med. Int. Health 2008, 13, 771–783. [Google Scholar] [CrossRef]
- Chattopadhyay, P.K.; Chelimo, K.; Embury, P.B.; Mulama, D.H.; Sumba, P.O.; Gostick, E.; Ladell, K.; Brodie, T.M.; Vulule, J.; Roederer, M.; et al. Holoendemic Malaria Exposure Is Associated with Altered Epstein-Barr Virus-Specific CD8(+) T-Cell Differentiation. J. Virol. 2013, 87, 1779–1788. [Google Scholar] [CrossRef]
- Moss, D.J.; Burrows, S.R.; Castelino, D.J.; Kane, R.G.; Pope, J.H.; Rickinson, A.B.; Alpers, M.P.; Heywood, P.F. A Comparison of Epstein-Barr Virus-Specific T-Cell Immunity in Malaria-Endemic and -Nonendemic Regions of Papua New Guinea. Int. J. Cancer 1983, 31, 727–732. [Google Scholar] [CrossRef]
- Whittle, H.C.; Brown, J.; Marsh, K.; Greenwood, B.M.; Seidelin, P.; Tighe, H.; Wedderburn, L. T-Cell Control of Epstein-Barr Virus-Infected B Cells Is Lost during P. Falciparum Malaria. Nature 1984, 312, 449–450. [Google Scholar] [CrossRef]
- Moormann, A.M.; Heller, K.N.; Chelimo, K.; Embury, P.; Ploutz-Snyder, R.; Otieno, J.A.; Oduor, M.; Münz, C.; Rochford, R. Children with Endemic Burkitt Lymphoma Are Deficient in EBNA1-Specific IFN-Gamma T Cell Responses. Int. J. Cancer 2009, 124, 1721–1726. [Google Scholar] [CrossRef]
- Long, H.M.; Taylor, G.S.; Rickinson, A.B. Immune Defence against EBV and EBV-Associated Disease. Curr. Opin. Immunol. 2011, 23, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Hislop, A.D.; Taylor, G.S. T-Cell Responses to EBV. In Current Topics in Microbiology and Immunology; Springer: Heidelberg/Berlin, Germany, 2015; pp. 325–353. [Google Scholar]
- Shannon-Lowe, C.; Rickinson, A. The Global Landscape of EBV-Associated Tumors. Front. Oncol. 2019, 9, 713. [Google Scholar] [CrossRef]
- Kutok, J.L.; Wang, F. Spectrum of epstein-barr virus–associated diseases. Annu. Rev. Pathol. Mech. Dis. 2006, 1, 375–404. [Google Scholar] [CrossRef]
- Kang, M.-S.; Kieff, E. Epstein-Barr Virus Latent Genes. Exp. Mol. Med. 2015, 47, e131. [Google Scholar] [CrossRef]
- Paludan, C.; Schmid, D.; Landthaler, M.; Vockerodt, M.; Kube, D.; Tuschl, T.; Münz, C. Endogenous MHC Class II Processing of a Viral Nuclear Antigen after Autophagy. Science 2005, 307, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Paludan, C.; Bickham, K.; Nikiforow, S.; Tsang, M.L.; Goodman, K.; Hanekom, W.A.; Fonteneau, J.-F.; Stevanovic, S.; Munz, C. Epstein-Barr Nuclear Antigen 1-Specific CD4 Th1 Cells Kill Burkitt’s Lymphoma Cells. J. Immunol. 2002, 169, 1593–1603. [Google Scholar] [CrossRef]
- Khanna, R.; Burrows, S.R.; Steigerwald-Mullen, P.M.; Moss, D.J.; Kurilla, M.G.; Cooper, L. Targeting Epstein-Barr Virus Nuclear Antigen 1 (EBNA1) through the Class II Pathway Restores Immune Recognition by EBNA1-Specific Cytotoxic T Lymphocytes: Evidence for HLA-DM-Independent Processing. Int. Immunol. 1997, 9, 1537–1543. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lee, S.P.; Brooks, J.M.; Al-Jarrah, H.; Thomas, W.A.; Haigh, T.A.; Taylor, G.S.; Humme, S.; Schepers, A.; Hammerschmidt, W.; Yates, J.L.; et al. CD8 T Cell Recognition of Endogenously Expressed Epstein-Barr Virus Nuclear Antigen 1. J. Exp. Med. 2004, 199, 1409–1420. [Google Scholar] [CrossRef]
- Voo, K.S.; Fu, T.; Wang, H.Y.; Tellam, J.; Heslop, H.E.; Brenner, M.K.; Rooney, C.M.; Wang, R.-F. Evidence for the Presentation of Major Histocompatibility Complex Class I-Restricted Epstein-Barr Virus Nuclear Antigen 1 Peptides to CD8+ T Lymphocytes. J. Exp. Med. 2004, 199, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Tellam, J.; Connolly, G.; Green, K.J.; Miles, J.J.; Moss, D.J.; Burrows, S.R.; Khanna, R. Endogenous Presentation of CD8+ T Cell Epitopes from Epstein-Barr Virus-Encoded Nuclear Antigen 1. J. Exp. Med. 2004, 199, 1421–1431. [Google Scholar] [CrossRef]
- O’Garra, A.; Vieira, P. T(H)1 Cells Control Themselves by Producing Interleukin-10. Nat. Rev. Immunol. 2007, 7, 425–428. [Google Scholar] [CrossRef]
- Muranski, P.; Restifo, N.P. Essentials of Th17 Cell Commitment and Plasticity. Blood 2013, 121, 2402–2414. [Google Scholar] [CrossRef]
- Chelimo, K.; Embury, P.B.; Sumba, P.O.; Vulule, J.; Ofulla, A.V.; Long, C.; Kazura, J.W.; Moormann, A.M. Age-Related Differences in Naturally Acquired T Cell Memory to Plasmodium Falciparum Merozoite Surface Protein 1. PLoS ONE 2011, 6, e24852. [Google Scholar] [CrossRef]
- Sallusto, F.; Langenkamp, A.; Geginat, J.; Lanzavecchia, A. Functional Subsets of Memory T Cells Identified by CCR7 Expression. In Current Topics in Microbiology and Immunology; Springer: Heidelberg/Berlin, Germany, 2000; pp. 167–171. [Google Scholar]
- Salisch, N.C.; Kaufmann, D.E.; Awad, A.S.; Reeves, R.K.; Tighe, D.P.; Li, Y.; Piatak, M., Jr.; Lifson, J.D.; Evans, D.T.; Pereyra, F.; et al. Inhibitory TCR Coreceptor PD-1 Is a Sensitive Indicator of Low-Level Replication of SIV and HIV-1. J. Immunol. 2010, 184, 476–487. [Google Scholar] [CrossRef]
- Butler, N.S.; Moebius, J.; Pewe, L.L.; Traore, B.; Doumbo, O.K.; Tygrett, L.T.; Waldschmidt, T.J.; Crompton, P.D.; Harty, J.T. Therapeutic Blockade of PD-L1 and LAG-3 Rapidly Clears Established Blood-Stage Plasmodium Infection. Nat. Immunol. 2011, 13, 188–195. [Google Scholar] [CrossRef]
- Gutierrez, F.R.S.; Mariano, F.S.; Oliveira, C.J.F.; Pavanelli, W.R.; Guedes, P.M.M.; Silva, G.K.; Campanelli, A.P.; Milanezi, C.M.; Azuma, M.; Honjo, T.; et al. Regulation of Trypanosoma Cruzi-Induced Myocarditis by Programmed Death Cell Receptor 1. Infect. Immun. 2011, 79, 1873–1881. [Google Scholar] [CrossRef]
- Bhadra, R.; Gigley, J.P.; Weiss, L.M.; Khan, I.A. Control of Toxoplasma Reactivation by Rescue of Dysfunctional CD8+ T-Cell Response via PD-1-PDL-1 Blockade. Proc. Natl. Acad. Sci. USA 2011, 108, 9196–9201. [Google Scholar] [CrossRef]
- Raj, D.K.; Nixon, C.P.; Nixon, C.E.; Dvorin, J.D.; DiPetrillo, C.G.; Pond-Tor, S.; Wu, H.-W.; Jolly, G.; Pischel, L.; Lu, A.; et al. Antibodies to PfSEA-1 Block Parasite Egress from RBCs and Protect against Malaria Infection. Science 2014, 344, 871–877. [Google Scholar] [CrossRef]
- Kaymaz, Y.; Oduor, C.I.; Aydemir, O.; Luftig, M.A.; Otieno, J.A.; Ong’echa, J.M.; Bailey, J.A.; Moormann, A.M. Epstein-Barr Virus Genomes Reveal Population Structure and Type 1 Association with Endemic Burkitt Lymphoma. J. Virol. 2020, 94, e02007–e02019. [Google Scholar] [CrossRef] [PubMed]
- Rainey, J.J.; Mwanda, W.O.; Wairiumu, P.; Moormann, A.M.; Wilson, M.L.; Rochford, R. Spatial Distribution of Burkitt’s Lymphoma in Kenya and Association with Malaria Risk. Trop. Med. Int. Health 2007, 12, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Ernst, K.C.; Adoka, S.O.; Kowuor, D.O.; Wilson, M.L.; John, C.C. Malaria Hotspot Areas in a Highland Kenya Site Are Consistent in Epidemic and Non-Epidemic Years and Are Associated with Ecological Factors. Malar. J. 2006, 5, 78. [Google Scholar] [CrossRef] [PubMed]
- Forconi, C.S.; Cosgrove, C.P.; Saikumar-Lakshmi, P.; Nixon, C.E.; Foley, J.; Ong’echa, J.M.; Otieno, J.A.; Alter, G.; Münz, C.; Moormann, A.M. Poorly Cytotoxic Terminally Differentiated CD56negCD16pos NK Cells Accumulate in Kenyan Children with Burkitt Lymphomas. Blood Adv. 2018, 2, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Heller, K.N.; Upshaw, J.; Seyoum, B.; Zebroski, H.; Münz, C. Distinct Memory CD4+ T-Cell Subsets Mediate Immune Recognition of Epstein Barr Virus Nuclear Antigen 1 in Healthy Virus Carriers. Blood 2007, 109, 1138–1146. [Google Scholar] [CrossRef]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central Memory and Effector Memory T Cell Subsets: Function, Generation, and Maintenance. Annu. Rev. Immunol. 2004, 22, 745–763. [Google Scholar] [CrossRef]
- Roederer, M.; Nozzi, J.L.; Nason, M.C. SPICE: Exploration and Analysis of Post-Cytometric Complex Multivariate Datasets. Cytometry A 2011, 79, 167–174. [Google Scholar] [CrossRef]
- Seder, R.A.; Darrah, P.A.; Roederer, M. T-Cell Quality in Memory and Protection: Implications for Vaccine Design. Nat. Rev. Immunol. 2008, 8, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.B.; Brooks, D.G. The Role of IL-10 in Regulating Immunity to Persistent Viral Infections. Curr. Top. Microbiol. Immunol. 2011, 350, 39–65. [Google Scholar] [PubMed]
- Speiser, D.E.; Utzschneider, D.T.; Oberle, S.G.; Münz, C.; Romero, P.; Zehn, D. T Cell Differentiation in Chronic Infection and Cancer: Functional Adaptation or Exhaustion? Nat. Rev. Immunol. 2014, 14, 768–774. [Google Scholar] [CrossRef]
- Long, H.M.; Chagoury, O.L.; Leese, A.M.; Ryan, G.B.; James, E.; Morton, L.T.; Abbott, R.J.M.; Sabbah, S.; Kwok, W.; Rickinson, A.B. MHC II Tetramers Visualize Human CD4+ T Cell Responses to Epstein-Barr Virus Infection and Demonstrate Atypical Kinetics of the Nuclear Antigen EBNA1 Response. J. Exp. Med. 2013, 210, 933–949. [Google Scholar] [CrossRef]
- Meckiff, B.J.; Ladell, K.; McLaren, J.E.; Ryan, G.B.; Leese, A.M.; James, E.A.; Price, D.A.; Long, H.M. Primary EBV Infection Induces an Acute Wave of Activated Antigen-Specific Cytotoxic CD4+ T Cells. J. Immunol. 2019, 203, 1276–1287. [Google Scholar] [CrossRef] [PubMed]
- Münz, C. Cytotoxicity in Epstein Barr Virus Specific Immune Control. Curr. Opin. Virol. 2021, 46, 1–8. [Google Scholar] [CrossRef]
- Münz, C. Latency and Lytic Replication in Epstein-Barr Virus-Associated Oncogenesis. Nat. Rev. Microbiol. 2019, 17, 691–700. [Google Scholar] [CrossRef]
- Hviid, L.; Kurtzhals, J.A.; Adabayeri, V.; Loizon, S.; Kemp, K.; Goka, B.Q.; Lim, A.; Mercereau-Puijalon, O.; Akanmori, B.D.; Behr, C. Perturbation and Proinflammatory Type Activation of V Delta 1(+) Gamma Delta T Cells in African Children with Plasmodium Falciparum Malaria. Infect. Immun. 2001, 69, 3190–3196. [Google Scholar] [CrossRef] [PubMed]
- Hviid, L.; Kurtzhals, J.A.; Dodoo, D.; Rodrigues, O.; Rønn, A.; Commey, J.O.; Nkrumah, F.K.; Theander, T.G. The Gamma/delta T-Cell Response to Plasmodium Falciparum Malaria in a Population in Which Malaria Is Endemic. Infect. Immun. 1996, 64, 4359–4362. [Google Scholar] [CrossRef]
- Futagbi, G.; Welbeck, J.E.; Tetteh, J.K.A.; Hviid, L.; Akanmori, B.D. Selective Activation of TCR-Gammadelta+ Cells in Endemic Burkitt’s Lymphoma. Malar. J. 2007, 6, 69. [Google Scholar] [CrossRef]
- Price, A.M.; Luftig, M.A. To Be or Not IIb: A Multi-Step Process for Epstein-Barr Virus Latency Establishment and Consequences for B Cell Tumorigenesis. PLoS Pathog. 2015, 11, e1004656. [Google Scholar] [CrossRef]
- Artavanis-Tsakonas, K.; Tongren, J.E.; Riley, E.M. The War between the Malaria Parasite and the Immune System: Immunity, Immunoregulation and Immunopathology. Clin. Exp. Immunol. 2003, 133, 145–152. [Google Scholar] [CrossRef]
- Parsons, E.; Otieno, J.A.; Ong’echa, J.M.; Nixon, C.E.; Vulule, J.; Münz, C.; Stewart, V.A.; Moormann, A.M. Regulatory T Cells in Endemic Burkitt Lymphoma Patients Are Associated with Poor Outcomes: A Prospective, Longitudinal Study. PLoS ONE 2016, 11, e0167841. [Google Scholar] [CrossRef]
- Morales, O.; Mrizak, D.; François, V.; Mustapha, R.; Miroux, C.; Depil, S.; Decouvelaere, A.-V.; Lionne-Huyghe, P.; Auriault, C.; de Launoit, Y.; et al. Epstein-Barr Virus Infection Induces an Increase of T Regulatory Type 1 Cells in Hodgkin Lymphoma Patients. Br. J. Haematol. 2014, 166, 875–890. [Google Scholar] [CrossRef]
- Roncarolo, M.G.; Gregori, S.; Bacchetta, R.; Battaglia, M.; Gagliani, N. The Biology of T Regulatory Type 1 Cells and Their Therapeutic Application in Immune-Mediated Diseases. Immunity 2018, 49, 1004–1019. [Google Scholar] [CrossRef]
- McLane, L.M.; Abdel-Hakeem, M.S.; Wherry, E.J. CD8 T Cell Exhaustion During Chronic Viral Infection and Cancer. Annu. Rev. Immunol. 2019, 37, 457–495. [Google Scholar] [CrossRef]
- Buckle, G.; Maranda, L.; Skiles, J.; Ong’echa, J.M.; Foley, J.; Epstein, M.; Vik, T.A.; Schroeder, A.; Lemberger, J.; Rosmarin, A.; et al. Factors influencing survival among Kenyan children diagnosed with endemic Burkitt lymphoma between 2003 and 2011: A historical cohort study. Int. J. Cancer 2016, 139, 1231–1240. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).