
A Retrospective Cross-Sectional Cohort Trial Assessing the Prevalence of MTHFR Polymorphisms and the Influence of Diet on Platinum Resistance in Ovarian Cancer Patients


Abstract
Simple Summary
Abstract
1. Introduction
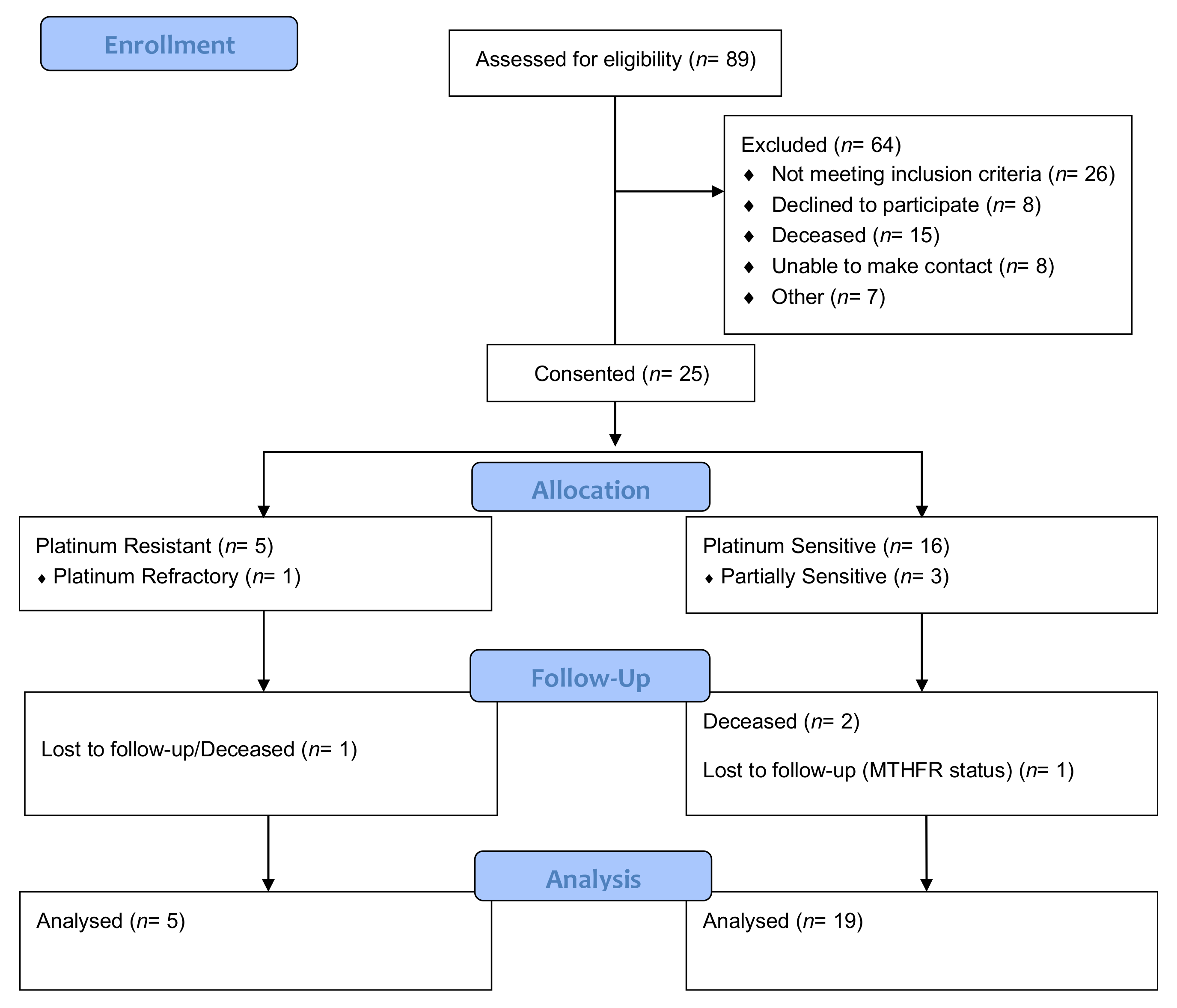
2. Materials and Methods
2.1. Data Collection
2.2. Data Analysis
3. Results
3.1. Participant Characteristics
3.2. Tumour and Treatment Characteristics
3.3. Pathology Results and Genetic Testing
3.4. Intergroup Analysis
3.5. Dietary Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vaughan, S.; Coward, J.I.; Bast, R.C.; Berchuck, A.; Berek, J.S.; Brenton, J.D.; Coukos, G.; Crum, C.C.; Drapkin, R.; Etemadmoghadam, D.; et al. Rethinking Ovarian Cancer: Recommendations for Improving Outcomes. Nat. Rev. Cancer 2011, 11, 719–725. [Google Scholar] [CrossRef] [PubMed]
- The World Ovarian Cancer Coalition. The World Ovarian Cancer Coalition Atlas Global Trends in Incidence, Mortality and Survival. Available online: https://worldovariancancercoalition.org/wp-content/uploads/2018/10/THE-WORLD-OVARIAN-CANCER-COALITION-ATLAS-2018.pdf (accessed on 23 June 2019).
- Australian Institute of Health and Welfare; Cancer Australia. Gynaecological Cancers in Australia: An Overview; Australian Institute of Health and Welfare: Canberra, Australia; Cancer Australia: Surry Hills, Australia, 2012; ISBN 9781742493480. [Google Scholar]
- Ledermann, J.A. First-line treatment of ovarian cancer: Questions and controversies to address. Ther. Adv. Med. Oncol. 2018, 10, 1758835918768232. [Google Scholar] [CrossRef]
- Polterauer, S.; Vergote, I.; Concin, N.; Braicu, I.; Chekerov, R.; Mahner, S.; Woelber, L.; Cadron, I.; Van Gorp, T.; Zeillinger, R.; et al. Prognostic value of residual tumor size in patients with epithelial ovarian cancer FIGO stages IIA-IV: Analysis of the OVCAD data. Int. J. Gynecol. Cancer 2012, 22, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Du Bois, A.; Quinn, M.; Thigpen, T.; Vermorken, J.; Avall-Lundqvist, E.; Bookman, M.; Bowtell, D.; Brady, M.; Casado, A.; Cervantes, A.; et al. 2004 consensus statements on the management of ovarian cancer: Final document of the 3rd International Gynecologic Cancer Intergroup Ovarian Cancer Consensus Conference (GCIG OCCC 2004). Ann. Oncol. 2005, 16, viii7–viii12. [Google Scholar] [CrossRef]
- Shen, D.-W.; Pouliot, L.M.; Hall, M.D.; Gottesman, M.M. Cisplatin Resistance: A Cellular Self-Defense Mechanism Resulting from Multiple Epigenetic and Genetic Changes. Pharmacol. Rev. 2012, 64, 706–721. [Google Scholar] [CrossRef]
- Martin, L.P.; Hamilton, T.C.; Schilder, R.J. Platinum Resistance: The Role of DNA Repair Pathways. Clin. Cancer Res. 2008, 14, 1291–1295. [Google Scholar] [CrossRef] [PubMed]
- Koukoura, O.; Spandidos, D.A.; Daponte, A.; Sifakis, S. DNA methylation profiles in ovarian cancer: Implication in diagnosis and therapy (Review). Mol. Med. Rep. 2014, 10, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Stuart, G.C.E.; Kitchener, H.; Bacon, M.; Dubois, A.; Friedlander, M.; Ledermann, J.; Marth, C.; Thigpen, T.; Trimble, E. 2010 Gynecologic Cancer InterGroup (GCIG) Consensus Statement on Clinical Trials in Ovarian Cancer Report From the Fourth Ovarian Cancer Consensus Conference. Int. J. Gynecol. Cancer 2011, 21, 750–755. [Google Scholar] [CrossRef]
- Wilson, M.K.; Pujade-Lauraine, E.; Aoki, D.; Mirza, M.R.; Lorusso, D.; Oza, A.M.; du Bois, A.; Vergote, I.; Reuss, A.; Bacon, M.; et al. 5th Ovarian Cancer Consensus Conference of the Gynecologic Cancer InterGroup: Recurrent Disease. Ann. Oncol. 2016, 28, 727–732. [Google Scholar] [CrossRef]
- Rizzo, A.; Napoli, A.; Roggiani, F.; Tomassetti, A.; Bagnoli, M.; Mezzanzanica, D. One-carbon metabolism: Biological players in epithelial ovarian cancer. Int. J. Mol. Sci. 2018, 19, 2092. [Google Scholar] [CrossRef] [PubMed]
- Bagnoli, M.; Granata, A.; Nicoletti, R.; Krishnamachary, B.; Bhujwalla, Z.M.; Canese, R.; Podo, F.; Canevari, S.; Iorio, E.; Mezzanzanica, D. Choline metabolism alteration: A focus on ovarian cancer. Front. Oncol. 2016, 6, 153. [Google Scholar] [CrossRef]
- Gonen, N.; Assaraf, Y.G. Drug Resistance Updates Antifolates in cancer therapy: Structure, activity and mechanisms of drug resistance. Drug Resist. Updates 2012, 15, 183–210. [Google Scholar] [CrossRef]
- Strickland, K.C.; Krupenko, N.I.; Krupenko, S.A. Molecular mechanisms underlying the potentially adverse effects of folate. Clin. Chem. Lab. Med. 2013, 51, 607–616. [Google Scholar] [CrossRef]
- Wang, Y.C.; Chen, Y.M.; Lin, Y.J.; Liu, S.P.; Chiang, E.P.I. GNMT expression increases hepatic folate contents and folate-dependent methionine synthase-mediated homocysteine remethylation. Mol. Med. 2011, 17, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Verschoor, M.L.; Singh, G. Ets-1 regulates intracellular glutathione levels: Key target for resistant ovarian cancer. Mol. Cancer 2013, 12, 138. [Google Scholar] [CrossRef]
- Si, M.; Lang, J. The roles of metallothioneins in carcinogenesis. J. Hematol. Oncol. 2018, 11, 107. [Google Scholar] [CrossRef]
- Feil, R.; Fraga, M.F. Epigenetics and the environment: Emerging patterns and implications. Nat. Rev. Genet. 2012, 13, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Ostrakhovitch, E.A.; Tabibzadeh, S. Homocysteine in Chronic Kidney Disease. Adv. Clin. Chem. 2015, 72, 77–106. [Google Scholar] [CrossRef]
- Schwahn, B.C.; Rozen, R. Methylenetetrahydrofolate Reductase Polymorphisms: Pharmacogenetic Effects. 2013. Available online: https://www.ncbi.nlm.nih.gov/books/NBK5968/ (accessed on 17 January 2019).
- Iverson Genetic Diagnostics Incorporated MTHFR Panel. Available online: http://www.iversongenetics.com/mthfr.html (accessed on 4 February 2019).
- Bogdanovic, O.; Santos-Pereira, J.M.; Mermoud, J.; Rotondo, J.C.; Tognon, M.; Martini, F.; Lanzillotti, C.; Mazziotta, C. Epigenetics of Male Infertility: The Role of DNA Methylation. Front. Cell Dev. Biol. 2021, 9, 1822. [Google Scholar] [CrossRef]
- Botezatu, A.; Socolov, D.; Iancu, I.V.; Huica, I.; Plesa, A.; Ungureanu, C.; Anton, G. Methylenetetrahydrofolate reductase (MTHFR) polymorphisms and promoter methylation in cervical oncogenic lesions and cancer. J. Cell. Mol. Med. 2013, 17, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Yin, F.; Wang, Q.; Zhang, W.; Li, L. Integration and bioinformatics analysis of DNA-methylated genes associated with drug resistance in ovarian cancer. Oncol. Lett. 2016, 12, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N. Epigenetic modulation of DNA methylation by nutrition and its mechanisms in animals. Anim. Nutr. 2015, 1, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, M.; Kagawa, Y. Genetic polymorphisms and folate status. Congenit. Anom. 2017, 57, 142–149. [Google Scholar] [CrossRef]
- Mckay, J.A.; Mathers, J.C. Diet induced epigenetic changes and their implications for health. Acta Physiol. 2011, 202, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Chiang, E.-P.; Wang, Y.-C.; Chiang, E.-P.; Wang, Y.-C.; Tang, F.-Y. Folate restriction and methylenetetrahydrofolate reductase 677T polymorphism decreases adoMet synthesis via folate-dependent remethylation in human-transformed lymphoblasts. Leukemia 2007, 21, 651–658. [Google Scholar] [CrossRef]
- Habel, N.; Hamidouche, Z.; Girault, I.; Patiñ O-Garcıá, A.; Lecanda, F.; Marie, P.J.; Fromigué, O. Zinc chelation: A metallothionein 2A’s mechanism of action involved in osteosarcoma cell death and chemotherapy resistance. Cit. Cell Death Dis. 2013, 4, e874. [Google Scholar] [CrossRef] [PubMed]
- Kelland, L.R. Preclinical Perspectives on Platinum Resistance. Drugs 2000, 59, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zaporozhan, V.; Tkachenko, O.; Rybin, A. Biomarkers of Platinum Resistance in Serous Ovarian Cancer. World Sci. 2018, 3, 17–18. [Google Scholar]
- Ostrakhovitch, E.A.; Olsson, P.-E.; Jiang, S.; Cherian, M.G. Interaction of metallothionein with tumor suppressor p53 protein. FEBS Lett. 2006, 580, 1235–1238. [Google Scholar] [CrossRef] [PubMed]
- Long, S.; Goldblatt, J. MTHFR genetic testing: Controversy and clinical implications. Aust. Fam. Physician 2016, 45, 237–240. [Google Scholar] [PubMed]
- Cancer Council Victoria Food Frequency Questionnaires—Cancer Council Victoria v3.2. Available online: https://www.cancervic.org.au/research/epidemiology/nutritional_assessment_services (accessed on 4 May 2018).
- FACIT.org FACT-O—Functional Assessment of Cancer Therapy—Ovarian Cancer. Available online: https://eprovide.mapi-trust.org/instruments/functional-assessment-of-cancer-therapy-ovarian-cancer (accessed on 30 April 2018).
- Gupta, D.; Braun, D.P.; Staren, E.D.; Markman, M. Longitudinal health-related quality of life assessment: Implications for prognosis in ovarian cancer. J. Ovarian Res. 2013, 6, 17. [Google Scholar] [CrossRef]
- Australian Government Department of Health Healthy Weight and You—Your Information Booklet. Available online: http://healthyweight.health.gov.au/wps/wcm/connect/healthyweight/9aa254c4-e7b0-4584-9c15-a185c5791a4b/FINAL+booklet+for+upload+%28D14-550905%29.PDF?MOD=AJPERES (accessed on 28 January 2019).
- Statistics—Ovarian Cancer Research Alliance. Available online: https://ocrahope.org/patients/about-ovarian-cancer/statistics/ (accessed on 2 February 2019).
- Gore, C.R.; Patvekar, M.M.; Kurade, S.J.; Kumar, H.; Pagaro, P.M. Malignant mixed mullerian tumor of the ovary. J. Obstet. Gynaecol. India 2014, 64, 62–64. [Google Scholar] [CrossRef][Green Version]
- Mackenzie, R.; Talhouk, A.; Eshragh, S.; Lau, S.; Cheung, D.; Chow, C.; Le, N.; Cook, L.S.; Wilkinson, N.; McDermott, J.; et al. Morphologic and Molecular Characteristics of Mixed Epithelial Ovarian Cancers. Am. J. Surg. Pathol. 2015, 39, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, E.L.; Abu-Rustum, N.R.; Sonoda, Y.; Aghajanian, C.; Barakat, R.R.; Chi, D.S. The effect of maximal surgical cytoreduction on sensitivity to platinum-taxane chemotherapy and subsequent survival in patients with advanced ovarian cancer. Gynecol. Oncol. 2008, 108, 276–281. [Google Scholar] [CrossRef]
- Palmirotta, R.; Silvestris, E.; D’Oronzo, S.; Cardascia, A.; Silvestris, F. Ovarian cancer: Novel molecular aspects for clinical assessment. Crit. Rev. Oncol. Hematol. 2017, 117, 12–29. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-W.; Friso, S. Epigenetics: A New Bridge between Nutrition and Health. Adv. Nutr. 2010, 1, 8–16. [Google Scholar] [CrossRef]
- Dixon, S.C.; Ibiebele, T.I.; Protani, M.M.; Beesley, J.; Defazio, A.; Crandon, A.J.; Gard, G.B.; Rome, R.M.; Webb, P.M.; Nagle, C.M. Dietary folate and related micronutrients, folate-metabolising genes, and ovarian cancer survival. Gynecol. Oncol. 2014, 132, 566–572. [Google Scholar] [CrossRef]
- Van Den Donk, M.; Pellis, L.; Crott, J.W.; Van Engeland, M.; Friederich, P.; Nagengast, F.M.; Van Bergeijk, J.D.; De Boer, S.Y.; Mason, J.B.; Kok, F.J.; et al. Folic Acid and Vitamin B-12 Smentation Does Not Favorably Influence Uracil Incorporation and Promoter Methylation in Rectal Mucosa DNA of Subjects with Previous Colorectal Adenomas. J. Nutr. 2007, 137, 2114–2120. [Google Scholar] [CrossRef]
- Markert, S.; Lassmann, S.; Gabriel, B.; Klar, M.; Werner, M.; Gitsch, G.; Kratz, F.; Hasenburg, A. Alpha-folate Receptor Expression in Epithelial Ovarian Carcinoma and Non-neoplastic Ovarian Tissue. Anticancer Res. 2008, 28, 3567–3572. [Google Scholar] [PubMed]
- Moore, K.N.; Martin, L.P.; O’Malley, D.M.; Matulonis, U.A.; Konner, J.A.; Vergote, I.; Ponte, J.F.; Birrer, M.J. A review of mirvetuximab soravtansine in the treatment of platinum-resistant ovarian cancer. Futur. Oncol. 2018, 14, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Méplan, C.; Richard, M.J.; Hainaut, P. Metalloregulation of the tumor suppressor protein p53: Zinc mediates the renaturation of p53 after exposure to metal chelators in vitro and in intact cells. Oncogene 2000, 19, 5227–5236. [Google Scholar] [CrossRef]
- Bell, D.; Berchuck, A.; Birrer, M.; Chien, J.; Cramer, D.W.; Dao, F.; Dhir, R.; Disaia, P.; Gabra, H.; Glenn, P.; et al. Integrated genomic analyses of ovarian carcinoma. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef]
- Vang, R.; Levine, D.A.; Soslow, R.A.; Zaloudek, C.; Shih, I.M.; Kurman, R.J. Molecular alterations of TP53 are a defining feature of ovarian high-grade serous carcinoma: A rereview of cases lacking tp53 mutations in the cancer genome atlas ovarian study. Int. J. Gynecol. Pathol. 2016, 35, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Chmelarova, M.; Krepinska, E.; Spacek, J.; Laco, J.; Beranek, M.; Palicka, V. Methylation in the p53 promoter in epithelial ovarian cancer. Clin. Transl. Oncol. 2013, 15, 160–163. [Google Scholar] [CrossRef]
- Feng, Q.; Deftereos, G.; Hawes, S.E.; Stern, J.E.; Willner, J.B.; Swisher, E.M.; Xi, L.; Drescher, C.; Urban, N.; Kiviat, N. DNA hypermethylation, Her-2/neu overexpression and p53 mutations in ovarian carcinoma. Gynecol. Oncol. 2008, 111, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Australian Dietary Guidelines | NHMRC. Available online: https://www.nhmrc.gov.au/about-us/publications/australian-dietary-guidelines#block-views-block-file-attachments-content-block-1 (accessed on 14 July 2020).
- De las Peñas, R.; Majem, M.; Perez-Altozano, J.; Virizuela, J.A.; Cancer, E.; Diz, P.; Donnay, O.; Hurtado, A.; Jimenez-Fonseca, P.; Ocon, M.J. SEOM clinical guidelines on nutrition in cancer patients (2018). Clin. Transl. Oncol. 2019, 21, 87–93. [Google Scholar] [CrossRef]
- Doyle, C.; Kushi, L.H.; Byers, T.; Courneya, K.S.; Demark-Wahnefried, W.; Grant, B.; McTiernan, A.; Rock, C.L.; Thompson, C.; Gansler, T.; et al. Nutrition and Physical Activity During and After Cancer Treatment: An American Cancer Society Guide for Informed Choices. CA. Cancer J. Clin. 2006, 56, 323–353. [Google Scholar] [CrossRef] [PubMed]
- Euser, A.M.; Zoccali, C.; Jager, K.J.; Dekker, F.W. Cohort Studies: Prospective versus Retrospective. Nephron Clin. Pract. 2009, 113, c214–c217. [Google Scholar] [CrossRef]
- Brattström, L.; Wilcken, D.E.L.; Öhrvik, J.; Brudin, L. Common Methylenetetrahydrofolate Reductase Gene Mutation Leads to Hyperhomocysteinemia but Not to Vascular Disease. Circulation 1998, 98, 2520–2526. [Google Scholar] [CrossRef]
- Vardavas, C.I.; Linardakis, M.K.; Hatzis, C.M.; Malliaraki, N.; Saris, W.H.; Kafatos, A.G. Tobacco Induced Diseases Smoking status in relation to serum folate and dietary vitamin intake. Tob. Induc. Dis. 2008, 4, 8. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genotype | Zygosity | % Normal Enzyme Activity |
|---|---|---|
| 1298A/677C | −/− (Normal) | 100% activity |
| A1298C | +/− (Heterozygous) | 83% activity |
| C677T | +/− (Heterozygous) | 66% activity |
| A1298C | +/+ (Homozygous) | 61% activity |
| C677T/A1298C | Compound heterozygous | 48% activity |
| C677T | +/+ (Homozygous) | 24% activity |
| Characteristics First-Line Therapy | Participants (n = ) |
|---|---|
| IP cisplatin 75 mg/m2 and paclitaxel 135 mg/m2 | 1 |
| Carboplatin AUC2 and paclitaxel 80 mg/m2 | 8 |
| Carboplatin AUC5, Caelyx® 30 mg/m2 | 1 |
| Carboplatin AUC5, Caelyx® 30 mg/m2 and Avastin® 15 mg/m2 | 1 |
| Carboplatin AUC5 and paclitaxel 175 mg/m2 | 2 |
| Carboplatin and ifosphamide | 1 |
| Carboplatin AUC5 and docetaxel 75 mg/m2 | 2 |
| Carboplatin AUC2 and paclitaxel 75 mg/m2 | 2 |
| Carboplatin AUC2 and paclitaxel 64 mg/m2 | 1 |
| Carboplatin and radiotherapy | 1 |
| Undisclosed carboplatin regime | 5 |
| Characteristics: Type of Cancer | Participants (n = ) |
|---|---|
| Ovarian | 14 |
| Peritoneal | 7 |
| Fallopian Tube | 3 |
| Stage of Cancer | Participants (n = ) |
| Stage 1 | 1 |
| Stage 1A | 1 |
| Stage 1C | 2 |
| Stage 2C | 1 |
| Stage 3 | 1 |
| Stage 3C | 8 |
| Stage 4 | 6 |
| Tumour Histotypes | Participants (n = ) |
| Serous | 18 |
| Endometriod | 3 |
| Clear-cell | 1 |
| Mixed clear-cell and endometrioid | 1 |
| Mixed serous and endometrioid | 1 |
| Carcinosarcoma (Müllerian) | 1 |
| Surgery | Participants (n = ) |
| Optimal debulking | 8 |
| Suboptimal debulking | 6 |
| Debulking status undisclosed | 11 |
| Platinum Response | Participants (n = ) |
| Sensitive | 16 |
| Partially sensitive | 3 |
| Resistant | 5 |
| Refractory | 1 |
| Tumour Grade | Participants (n = ) |
| High | 22 |
| Low | 1 |
| Undefined | 2 |
| MTHFR Allele. | p-Value | Coefficient | Stand. Error | 95% CI |
|---|---|---|---|---|
| Serum Folate | ||||
| Homozygous C677T | 0.199 | 0.27 | 0.21 | −0.14–0.7 |
| Homozygous A1298C | 0.53 | 0.05 | 0.08 | −0.11–0.21 |
| Heterozygous C677T | 0.57 | 0.04 | 0.08 | −0.11–0.21 |
| Heterozygous A1298C | ND | ND | ND | ND |
| Heterozygous C677T + A1298C | 0.76 | −0.02 | 0.07 | −0.17–0.12 |
| Serum Vitamin B12 | ||||
| Homozygous C677T | 0.46 | −0.005 | 0.003 | −0.1–0.001 |
| Homozygous A1298C | 0.40 | −0.001 | 0.001 | −0.003–0.001 |
| Heterozygous C677T | 0.14 | −0.002 | 0.001 | −0.006–0.0009 |
| Heterozygous A1298C | ND | ND | ND | ND |
| Heterozygous C677T + A1298C | 0.29 | −0.001 | 0.001 | −0.004–0.001 |
| Homocysteine | ||||
| Homozygous C677T | 0.068 | −0.02 | 0.11 | −0.42–0.01 |
| Homozygous A1298C | 0.12 | −0.09 | 0.06 | −0.22–0.2 |
| Heterozygous C677T | 0.12 | −0.09 | 0.06 | −0.21–0.02 |
| Heterozygous A1298C | ND | ND | ND | ND |
| Heterozygous C677T + A1298C | 0.344 | −0.044 | 0.04 | −0.13–0.04 |
| MTHFR Allele | Total no. of Mutations | Serum Folate | Serum Vitamin B12 | Active B12 | Homocysteine | Platinum Resistant | Partial Sensitivity | Platinum Sensitive |
|---|---|---|---|---|---|---|---|---|
| Homozygous C677T | 1 | P = 0.185 | 1 in range P = 0.890 | P = 0.055 | P = 0.940 | P = 0.610 | P = 0.706 | P = 0.444 |
| FE = 0.682 | FE = 1.000 | FE = 0.591 | FE = 1.000 | FE = 1.000 | FE = 1.000 | FE = 1.000 | ||
| Homozygous A1298C | 2 | P = 0.185 | 2 in range P = 0.783 | P = 0.590 | P = 0.661 | P = 0.461 | P = 0.586 | P = 0.269 |
| FE = 0.463 | FE = 1.000 | FE = 0.844 | FE = 0.983 | FE = 1.000 | FE = 0.520 | |||
| Heterozygous C677T | 3 | P = 0.500 | 3 in range P = 0.680 | P = 0.343 | P = 0.284 | P = 0.538 | P = 0.495 | P = 0.918 |
| FE = 0.829 | FE = 1.000 | FE = 0.642 | FE = 0.553 | FE = 0.504 | FE = 1.000 | FE = 1.000 | ||
| Heterozygous A1298C | 7 | P = 0.480 | 6 in range 1 over range P = 0.783 | P = 0.548 | P = 0.284 | P = 0.504 | P = 0.826 | P = 0.656 |
| FE = 0.872 | FE = 1.000 | FE = 1.000 | FE = 0.292 | FE = 0.597 | FE = 1.000 | FE = 0.673 | ||
| Heterozygous C677T + A1298C | 5 | P = 0.320 | 1 deficient 4 in range P = 0.117 | P = 0.202 | P = 0.285 | P = 1.000 | P = 0.538 | P = 0.211 |
| FE = 0.379 | FE = 0.326 | FE = 0.293 | FE = 0.388 | FE = 1.000 | FE = 0.504 | FE = 0.312 | ||
| No mutation | 4 | P = 0.185 | 2 in range 2 over range P = 0.062 | P = 0.565 | P = 0.284 | P = 0.275 | P = 0.420 | P = 0.102 |
| FE = 0.217 | FE = 0.135 | FE = 0.833 | FE = 0.460 | FE = 0.549 | FE = 1.000 | FE = 0.260 |
| MTHFR Allele | Total No. of Mutations | Dietary Folate Intake | Dietary Vitamin B12 | Dietary Vitamin B6 | Dietary Zinc | ||||
|---|---|---|---|---|---|---|---|---|---|
| Below RDI <400 mcg/d | Above RDI >400 mcg/d | Below RDI <2.4 mcg/d | Above RDI >2.4 mcg/d | Below RDI <1.5 mg/d | Above RDI >1.5 mg/d | Below RDI <8 mg/d | Above RDI >8 mg/d | ||
| Homozygous C677T | 1 | 0/6 | 1/19 | 1/14 | 0/11 | 1/22 | 0/3 | 0/8 | 1/17 |
| P = 0.566 Fisher’s Exact = 1.000 | P = 0.137 Fisher’s Exact = 0.320 | P = 0.763 Fisher’s Exact = 1.000 | P = 0.484 Fisher’s Exact = 1.000 | ||||||
| Homozygous A1298C | 2 | 1/6 | 1/19 | 0/14 | 2/11 | 0/22 | 0/3 | 1/8 | 1/17 |
| P = 0.369 Fisher’s exact = 0.430 | P = 0.312 Fisher’s exact = 1.000 | P = 0.664 Fisher’s exact = 1.000 | P = 0.569 Fisher’s exact = 1.000 | ||||||
| Heterozygous C677T | 3 | 2/6 | 1/19 | 3/14 | 0/11 | 3/22 | 0/3 | 3/8 | 0/17 |
| P = 0.065 Fisher’s Exact = 0.133 | P = 0.007 Fisher’s exact = 0.024 | P = 0.586 Fisher’s exact = 1.000 | P = 0.007 Fisher’s exact = 0.024 | ||||||
| Heterozygous A1298C | 7 | 1/6 | 6/19 | 1/14 | 6/11 | 5/22 | 2/2 | 2/8 | 5/17 |
| P = 0.478 Fisher’s exact = 0.637 | P = 0.236 Fisher’s exact = 0.362 | P = 0.018 Fisher’s exact = 0.070 | P = 0.819 Fisher’s exact = 1.000 | ||||||
| Heterozygous C677T + A1298C | 5 | ||||||||
| 0/6 | 5/19 | 1/14 | 4/11 | 5/22 | 0/2 | 1/8 | 4/17 | ||
| P = 0.160 Fisher’s Exact = 0.289 | P = 0.520 Fisher’s Exact = 1.000 | P = 0.461 Fisher’s exact = 1.000 | P = 0.520 Fisher’s exact = 1.000 | ||||||
| No mutation | 4 | 1/6 | 3/19 | 0/14 | 4/11 | 4/22 | 0/2 | 0/8 | 4/17 |
| P = 0.959 Fisher’s Exact = 1.000 | P = 0.134 Fisher’s Exact = 0.269 | P = 0.520 Fisher’s exact = 0.700 | P = 0.134 Fisher’s exact = 0.269 | ||||||
| MTHFR Allele | Total No. of Mutations | Platinum Response | Serum Folate (p = ) | Active B12 (p = ) | Homocysteine (p = ) | Dietary Folate Intake (p = ) | Dietary Vitamin B12 Intake (p = ) | Dietary Vitamin B6 Intake (p = ) | Dietary Zinc Intake (p = ) |
|---|---|---|---|---|---|---|---|---|---|
| Homozygous C677T | 1 | Sensitive | 0.149 | 0.4218 | 0.5844 | 0.5101 | 0.1424 | 0.5918 | 0.5101 |
| Partial | 0.1772 | 0.6514 | 0.7267 | 0.7142 | 0.1833 | 0.7959 | 0.6734 | ||
| Resistant | 0.1504 | 0.6137 | 0.6552 | 0.5918 | 0.2651 | 0.7551 | 0.5509 | ||
| Homozygous A1298C | 2 | Sensitive | 0.2013 | 0.4384 | 0.4284 | 0.2156 | 0.2096 | 0.3399 | 0.2156 |
| Partial | 0.337 | 0.8168 | 0.7264 | 0.4241 | 0.4441 | 0.6265 | 0.5148 | ||
| Resistant | 0.2233 | 0.6531 | 0.5635 | 0.4524 | 0.2496 | 0.5622 | 0.5151 | ||
| Heterozygous C677T | 3 | Sensitive | 0.9067 | 0.4997 | 0.6010 | 0.2161 | 0.0157 | 0.7643 | 0.0086 |
| Partial | 0.7221 | 0.2802 | 0.6079 | 0.0790 | 0.0030 | 0.4873 | 0.0030 | ||
| Resistant | 0.7308 | 0.4740 | 0.3925 | 0.1811 | 0.0068 | 0.5884 | 0.0189 | ||
| Heterozygous A1298C | 7 | Sensitive | 0.7106 | 0.4855 | 0.2326 | 0.6467 | 0.3560 | 0.0597 | 0.8259 |
| Partial | 0.8094 | 0.6194 | 0.1842 | 0.6571 | 0.1530 | 0.0547 | 0.8925 | ||
| Resistant | 0.5962 | 0.5254 | 0.1996 | 0.6278 | 0.4053 | 0.0610 | 0.7752 | ||
| Heterozygous C677T + A1298C | 5 | Sensitive | 0.2902 | 0.2632 | 0.1265 | 0.0627 | 0.2976 | 0.2652 | 0.2145 |
| Partial | 0.5873 | 0.4457 | 0.2161 | 0.0510 | 0.2657 | 0.5530 | 0.2657 | ||
| Resistant | 0.6793 | 0.5044 | 0.8448 | 0.2097 | 0.7991 | 0.6174 | 0.8006 | ||
| No mutation | 4 | Sensitive | 0.0478 | 0.1160 | 0.0886 | 0.1275 | 0.0350 | 0.1010 | 0.0514 |
| Partial | 0.2151 | 0.3066 | 0.5292 | 0.5314 | 0.1797 | 0.3739 | 0.1797 | ||
| Resistant | 0.240 | 0.1913 | 0.4013 | 0.3739 | 0.0514 | 0.2972 | 0.0731 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phillips-Chavez, C.; Coward, J.; Watson, M.; Schloss, J. A Retrospective Cross-Sectional Cohort Trial Assessing the Prevalence of MTHFR Polymorphisms and the Influence of Diet on Platinum Resistance in Ovarian Cancer Patients. Cancers 2021, 13, 5215. https://doi.org/10.3390/cancers13205215
Phillips-Chavez C, Coward J, Watson M, Schloss J. A Retrospective Cross-Sectional Cohort Trial Assessing the Prevalence of MTHFR Polymorphisms and the Influence of Diet on Platinum Resistance in Ovarian Cancer Patients. Cancers. 2021; 13(20):5215. https://doi.org/10.3390/cancers13205215
Chicago/Turabian StylePhillips-Chavez, Caitlin, Jermaine Coward, Michael Watson, and Janet Schloss. 2021. "A Retrospective Cross-Sectional Cohort Trial Assessing the Prevalence of MTHFR Polymorphisms and the Influence of Diet on Platinum Resistance in Ovarian Cancer Patients" Cancers 13, no. 20: 5215. https://doi.org/10.3390/cancers13205215
APA StylePhillips-Chavez, C., Coward, J., Watson, M., & Schloss, J. (2021). A Retrospective Cross-Sectional Cohort Trial Assessing the Prevalence of MTHFR Polymorphisms and the Influence of Diet on Platinum Resistance in Ovarian Cancer Patients. Cancers, 13(20), 5215. https://doi.org/10.3390/cancers13205215