
Therapeutic Potential of Innate Lymphoid Cells for Multiple Myeloma Therapy

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
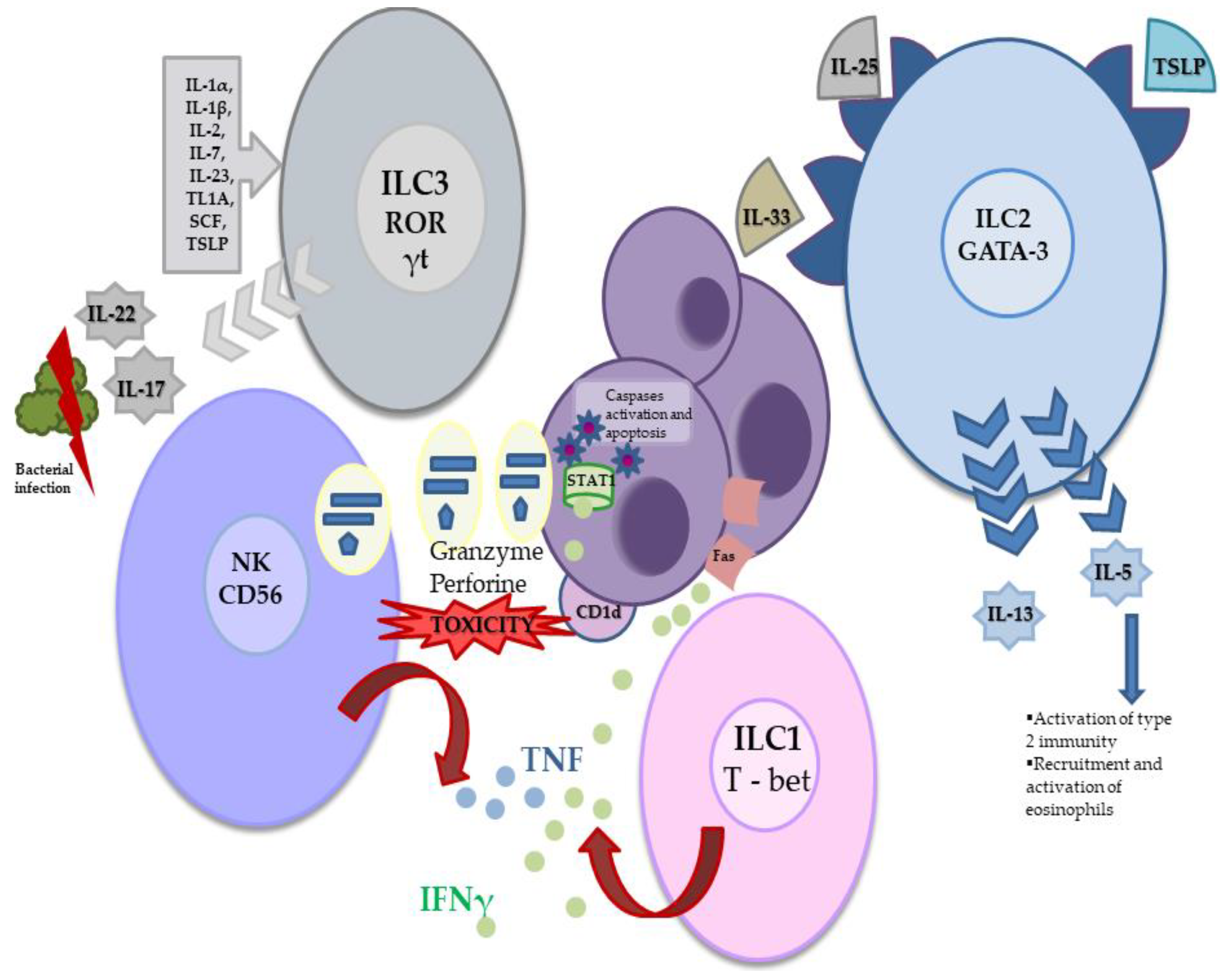
2. Innate Lymphoid Cells
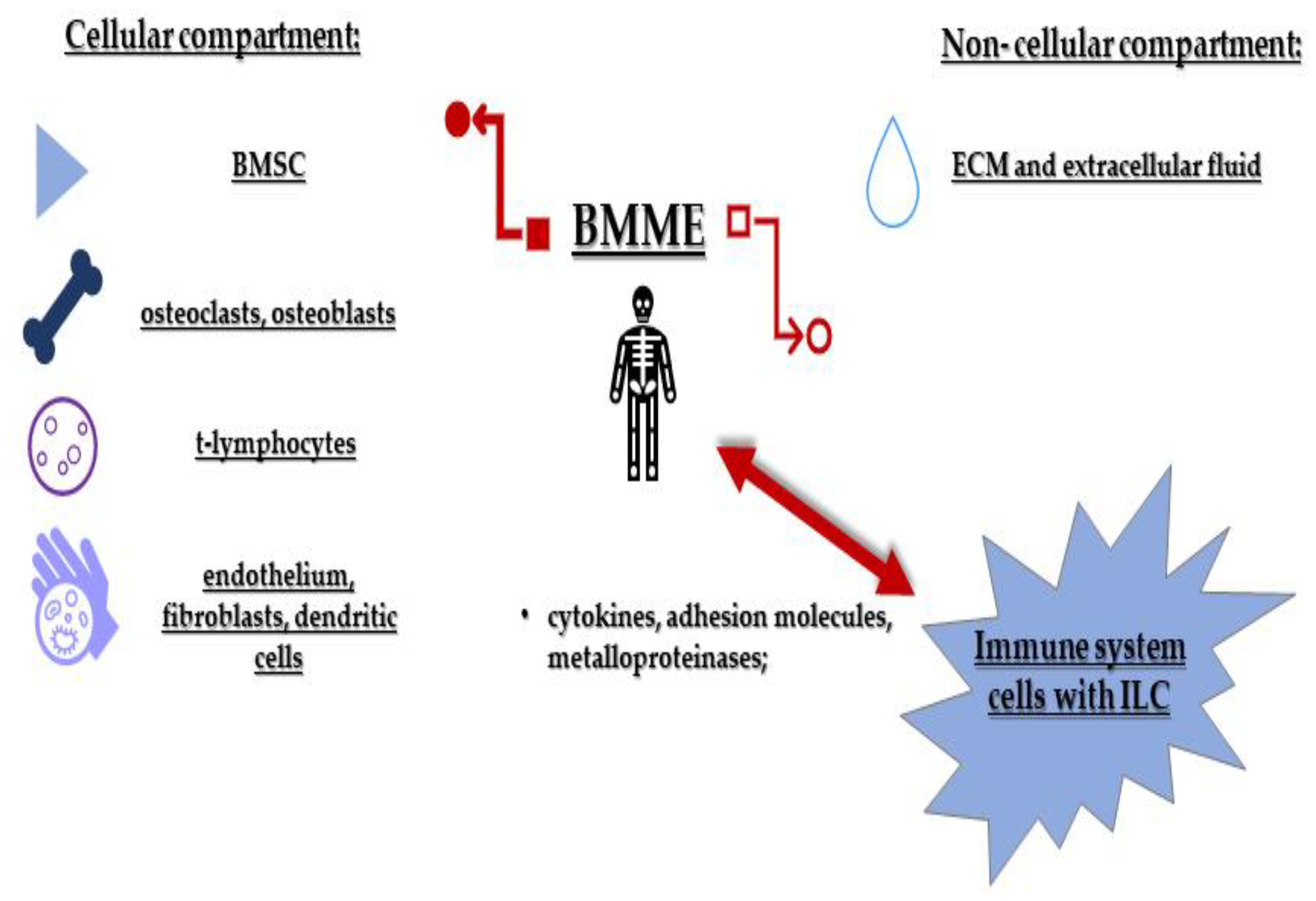
3. Innate Lymphoid Cells in the Multiple Myeloma Microenvironment
4. Innate Lymphoid Cells in the Prevention of Multiple Myeloma
4.1. The Role of NK Cells
4.2. The Role of ILC1s
5. Innate Lymphoid Cells in the Development of Multiple Myeloma
5.1. NK Cell Dysfunction
5.1.1. Humoral Mechanism
5.1.2. Signalling Mechanism
5.2. The Role of ILC1s
5.3. The Role of ILC2s
5.4. The Role of ILC3s
6. Targeting Innate Lymphoid Cells for Multiple Myeloma Immunotherapy
6.1. Non-NK ILC Cell Therapy for MM
ILCs in Graft-Versus-Host Disease
6.2. NK ILC Cell Therapy for MM
6.2.1. NK Cells during Daratumumab Therapy
6.2.2. CAR-NK Cell Therapy
Clinical Trials
6.3. Targeting ILCs through Receptors
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ludwig, H.; Durie, S.N.; Meckl, A.; Hinke, A.; Durie, B. Multiple Myeloma Incidence and Mortality Around the Globe; Interrelations Between Health Access and Quality, Economic Resources, and Patient Empowerment. Oncology 2020, 25, e1406–e1413. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, S.V. Multiple myeloma: 2020 update on diagnosis, risk-stratification and management. Am. J. Hematol. 2020, 95, 548–567. [Google Scholar] [CrossRef] [Green Version]
- Hájek, R.; Jarkovsky, J.; Maisnar, V.; Pour, L.; Špička, I.; Minařík, J.; Gregora, E.; Kessler, P.; Sýkora, M.; Fraňková, H.; et al. Real-world Outcomes of Multiple Myeloma: Retrospective Analysis of the Czech Registry of Monoclonal Gammopathies. Clin. Lymphoma Myeloma Leuk. 2018, 18, e219–e240. [Google Scholar] [CrossRef]
- Akizuki, K.; Matsuoka, H.; Toyama, T.; Kamiunten, A.; Sekine, M.; Shide, K.; Kameda, T.; Kawano, N.; Maeda, K.; Takeuchi, M.; et al. Real-World Data on Clinical Features, Outcomes, and Prognostic Factors in Multiple Myeloma from Miyazaki Prefecture, Japan. J. Clin. Med. 2020, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-C.; Lin, S.-F. Mechanisms of Drug Resistance in Relapse and Refractory Multiple Myeloma. BioMed Res. Int. 2015, 2015, 341430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadas, A.; Asimakopoulos, F. Mechanisms of Resistance in Multiple Myeloma. Sphingolipids Dis. 2017, 249, 251–288. [Google Scholar] [CrossRef]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate Lymphoid Cells: 10 Years On. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spits, H.; Cupedo, T. Innate Lymphoid Cells: Emerging Insights in Development, Lineage Relationships, and Function. Annu. Rev. Immunol. 2012, 30, 647–675. [Google Scholar] [CrossRef]
- Artis, D.; Spits, H. The biology of innate lymphoid cells. Nat. Cell Biol. 2015, 517, 293–301. [Google Scholar] [CrossRef]
- Eberl, G.; Di Santo, J.P.; Vivier, E. The brave new world of innate lymphoid cells. Nat. Immunol. 2014, 16, 1–5. [Google Scholar] [CrossRef]
- Fuchs, A.; Vermi, W.; Lee, J.S.; Lonardi, S.; Gilfillan, S.; Newberry, R.D.; Cella, M.; Colonna, M. Intraepithelial Type 1 Innate Lymphoid Cells Are a Unique Subset of IL-12- and IL-15-Responsive IFN-γ-Producing Cells. Immunity 2013, 38, 769–781. [Google Scholar] [CrossRef] [Green Version]
- Bernink, J.H.; Peters, C.P.; Munneke, M.; Velde, A.A.T.; Meijer, S.L.; Weijer, K.; Hreggvidsdottir, H.S.; Heinsbroek, S.E.; Legrand, N.; Buskens, C.J.; et al. Human type 1 innate lymphoid cells accumulate in inflamed mucosal tissues. Nat. Immunol. 2013, 14, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Hoyler, T.; Klose, C.S.; Souabni, A.; Turqueti-Neves, A.; Pfeifer, D.; Rawlins, E.L.; Voehringer, D.; Busslinger, M.; Diefenbach, A. The Transcription Factor GATA-3 Controls Cell Fate and Maintenance of Type 2 Innate Lymphoid Cells. Immunity 2012, 37, 634–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerbe, F.; Sidot, E.; Smyth, D.J.; Ohmoto, M.; Matsumoto, I.; Dardalhon, V.; Cesses, P.; Garnier, L.; Pouzolles, M.; Brulin, B.; et al. Intestinal epithelial tuft cells initiate type 2 mucosal immunity to helminth parasites. Nature 2016, 529, 226–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallrapp, A.; Riesenfeld, S.J.; Burkett, P.R.; Abdulnour, R.-E.E.; Nyman, J.; Dionne, D.; Hofree, M.; Cuoco, M.S.; Rodman, C.; Farouq, D.; et al. The neuropeptide NMU amplifies ILC2-driven allergic lung inflammation. Nature 2017, 549, 351–356. [Google Scholar] [CrossRef]
- Ibiza, S.; García-Cassani, B.; Ribeiro, H.; Carvalho, T.; Almeida, L.; Marques, R.; Misic, A.M.; Bartow-McKenney, C.; Larson, D.M.; Pavan, W.J.; et al. Glial-cell-derived neuroregulators control type 3 innate lymphoid cells and gut defence. Nature 2016, 535, 440–443. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Xia, P.; Chen, Y.; Qu, Y.; Xiong, Z.; Ye, B.; Du, Y.; Tian, Y.; Yin, Z.; Xu, Z.; et al. Regulatory Innate Lymphoid Cells Control Innate Intestinal Inflammation. Cell 2017, 171, 201–216.e18. [Google Scholar] [CrossRef] [Green Version]
- Klose, C.S.N.; Artis, D. Innate lymphoid cells control signaling circuits to regulate tissue-specific immunity. Cell Res. 2020, 30, 475–491. [Google Scholar] [CrossRef]
- Oliphant, C.J.; Hwang, Y.Y.; Walker, J.A.; Salimi, M.; Wong, S.H.; Brewer, J.; Englezakis, A.; Barlow, J.L.; Hams, E.; Scanlon, S.T.; et al. MHCII-Mediated Dialog between Group 2 Innate Lymphoid Cells and CD4+ T Cells Potentiates Type 2 Immunity and Promotes Parasitic Helminth Expulsion. Immunity 2014, 41, 283–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veiga-Fernandes, H.; Coles, M.C.; Foster, K.E.; Patel, A.; Williams, A.; Natarajan, D.; Barlow, A.J.; Pachnis, V.; Kioussis, D. Tyrosine kinase receptor RET is a key regulator of Peyer’s Patch organogenesis. Nat. Cell Biol. 2007, 446, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Eberl, G.; Colonna, M.; Di Santo, J.P.; McKenzie, A.N.J. Innate lymphoid cells: A new paradigm in immunology. Science 2015, 348, aaa6566. [Google Scholar] [CrossRef] [Green Version]
- García-Ortiz, A.; Rodríguez-García, Y.; Encinas, J.; Maroto-Martín, E.; Castellano, E.; Teixidó, J.; Martínez-López, J. The Role of Tumor Microenvironment in Multiple Myeloma Development and Progression. Cancers 2021, 13, 217. [Google Scholar] [CrossRef]
- De Raeve, H.R.; Vanderkerken, K. The role of the bone marrow microenvironment in multiple myeloma. Histol. Histopathol. 2005, 20, 1227–1250. [Google Scholar] [CrossRef]
- Bolkun, L.; Lemancewicz, D.; Jabłońska, E.; Kulczynska-Przybik, A.; Bolkun-Skornicka, U.; Kloczko, J.; Dzięcioł, J. BAFF and APRIL as TNF superfamily molecules and angiogenesis parallel progression of human multiple myeloma. Ann. Hematol. 2014, 93, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Ungefroren, H.; Sebens, S.; Seidl, D.; Lehnert, H.; Hass, R. Interaction of tumor cells with the microenvironment. Cell Commun. Signal. 2011, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Zamarron, B.; Chen, W. Dual Roles of Immune Cells and Their Factors in Cancer Development and Progression. Int. J. Biol. Sci. 2011, 7, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huang, P. Cancer-stromal interactions. Cancer Biol. Ther. 2011, 11, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narendra, B.L.; Reddy, K.E.; Shantikumar, S.; Ramakrishna, S. Immune system: A double-edged sword in cancer. Inflamm. Res. 2013, 62, 823–834. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, G.; Fan, X.; Dikiy, S.; Lee, S.Y.; Rudensky, A.Y. Tissue residency of innate lymphoid cells in lymphoid and nonlymphoid organs. Science 2015, 350, 981–985. [Google Scholar] [CrossRef] [Green Version]
- Pearson, C.; Thornton, E.; McKenzie, B.; Schaupp, A.-L.; Huskens, N.; Griseri, T.; West, N.; Tung, S.; Seddon, B.P.; Uhlig, H.H.; et al. ILC3 GM-CSF production and mobilisation orchestrate acute intestinal inflammation. eLife 2016, 5, e10066. [Google Scholar] [CrossRef]
- Lim, A.I.; Menegatti, S.; Bustamante, J.; Le Bourhis, L.; Allez, M.; Rogge, L.; Casanova, J.-L.; Yssel, H.; Di Santo, J.P. IL-12 drives functional plasticity of human group 2 innate lymphoid cells. J. Exp. Med. 2016, 213, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Lim, A.I.; Verrier, T.; Vosshenrich, C.; Di Santo, J.P. Developmental options and functional plasticity of innate lymphoid cells. Curr. Opin. Immunol. 2017, 44, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Bal, S.M.; Golebski, K.; Spits, H. Plasticity of innate lymphoid cell subsets. Nat. Rev. Immunol. 2020, 20, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Hinshaw, D.C.; Shevde, L.A. The tumor microenvironment innately modulates cancer progression. Cancer Res. 2019, 79, 4557–4566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Sanz, R.; González, M.; Orfão, A.; Moro, M.J.; Hernández, J.M.; Borrego, D.; Carnero, M.; Casanova, F.; Bárez, A.; Jiménez, R.; et al. Analysis of natural killer-associated antigens in peripheral blood and bone marrow of multiple myeloma patients and prognostic implications. Br. J. Haematol. 1996, 93, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Viel, S.; Charrier, E.; Marçais, A.; Rouzaire, P.; Bienvenu, J.; Karlin, L.; Salles, G.; Walzer, T. Monitoring NK cell activity in patients with hematological malignancies. OncoImmunology 2013, 2, e26011. [Google Scholar] [CrossRef] [PubMed]
- Guillerey, C.; Smyth, M.J. NK Cells and Cancer Immunoediting. Curr. Top. Microbiol. Immunol. 2016, 395, 115–145. [Google Scholar] [CrossRef] [PubMed]
- Exley, M.; Garcia, J.; Wilson, S.B.; Spada, F.; Gerdes, D.; Tahir, S.M.A.; Patton, K.T.; Blumberg¶, R.S.; Porcelli, S.; Chott, A.; et al. CD1d structure and regulation on human thymocytes, peripheral blood T cells, B cells and monocytes. Immunology 2000, 100, 37–47. [Google Scholar] [CrossRef]
- Zeng, Z.-H.; Castaño, A.R.; Segelke, B.W.; Stura, E.A.; Peterson, P.A.; Wilson, I.A. Crystal Structure of Mouse CD1: An MHC-Like Fold with a Large Hydrophobic Binding Groove. Science 1997, 277, 339–345. [Google Scholar] [CrossRef]
- Yu, C.; Milstein, C. A physical map linking the five CD1 human thymocyte differentiation antigen genes. EMBO J. 1989, 8, 3727–3732. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Rossjohn, J. New ways to turn on NKT cells. J. Exp. Med. 2011, 208, 1121–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanoudakis, E.; Hu, M.; Naresh, K.; Terpos, E.; Melo, V.; Reid, A.; Kotsianidis, I.; Abdalla, S.; Rahemtulla, A.; Karadimitris, A. Regulation of multiple myeloma survival and progression by CD1d. Blood 2009, 113, 2498–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seliger, B.; Ruiz-Cabello, F.; Garrido, F. Chapter 7 IFN Inducibility of Major Histocompatibility Antigens in Tumors. Adv. Cancer Res. 2008, 101, 249–276. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Chakravarty, S.D.; Ivashkiv, L.B. Regulation of interferon and Toll-like receptor signaling during macrophage activation by opposing feedforward and feedback inhibition mechanisms. Immunol. Rev. 2008, 226, 41–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, F.; Algarra, I. MHC antigens and tumor escape from immune surveillance. Adv. Cancer Res. 2001, 83, 117–158. [Google Scholar] [CrossRef]
- Kelker, H.C.; Le, J.; Rubin, B.Y.; Yip, Y.K.; Nagler, C.; Vilcek, J. Three molecular weight forms of natural human interferon-gamma revealed by immunoprecipitation with monoclonal antibody. J. Biol. Chem. 1984, 259, 4301–4304. [Google Scholar] [CrossRef]
- Takaoka, A.; Mitani, Y.; Suemori, H.; Sato, M.; Yokochi, T.; Noguchi, S.; Tanaka, N.; Taniguchi, T. Cross Talk Between Interferon-gamma and -alpha /beta Signaling Components in Caveolar Membrane Domains. Science 2000, 288, 2357–2360. [Google Scholar] [CrossRef]
- Xu, X.; Fu, X.Y.; Plate, J.; Chong, A.S. IFN-gamma induces cell growth inhibition by Fas-mediated apoptosis: Requirement of STAT1 protein for up-regulation of Fas and FasL expression. Cancer Res. 1998, 58, 2832–2837. [Google Scholar]
- Miller, C.H.; Maher, S.; Young, H.A. Clinical Use of Interferon-γ. Ann. N. Y. Acad. Sci. 2009, 1182, 69–79. [Google Scholar] [CrossRef]
- Palumbo, A.; Battaglio, S.; Napoli, P.; Omedè, P.; Fusaro, A.; Bruno, B.; Boccadoro, M.; Pileri, A. Recombinant interferon-gamma inhibits the in vitro proliferation of human myeloma cells. Br. J. Haematol. 1994, 86, 726–732. [Google Scholar] [CrossRef]
- Palumbo, A.; Bruno, B.; Boccadoro, M.; Pileri, A. Interferon-γ in Multiple Myeloma. Leuk. Lymphoma 1995, 18, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.; Morita, A.A.; Broto, G.E.; Takakura, É.; Da Silva, S.S.; Tomiotto-Pellissier, F.; Conchon-Costa, I.; Pavanelli, W.R.; Panis, C.; Barbosa, D.S. Interferon-gamma in mobilized stem cells: A possible prognostic marker in early post-transplant management in multiple myeloma. Cytokine 2018, 108, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Tsuyama, N.; Danjoh, I.; Otsuyama, K.-I.; Obata, M.; Tahara, H.; Ohta, T.; Ishikawa, H. IL-6-induced Bcl6 variant 2 supports IL-6-dependent myeloma cell proliferation and survival through STAT3. Biochem. Biophys. Res. Commun. 2005, 337, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Hideshima, T.; Mitsiades, C.; Ikeda, H.; Chauhan, D.; Raje, N.; Gorgun, G.; Hideshima, H.; Munshi, N.C.; Richardson, P.G.; Carrasco, D.R.; et al. A proto-oncogene BCL6 is up-regulated in the bone marrow microenvironment in multiple myeloma cells. Blood 2010, 115, 3772–3775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ujvari, D.; Nagy, N.; Madapura, H.S.; Kallas, T.; Kröhnke, M.C.; Stenke, L.; Klein, E.; Salamon, D. Interferon γ is a strong, STAT1-dependent direct inducer of BCL6 expression in multiple myeloma cells. Biochem. Biophys. Res. Commun. 2018, 498, 502–508. [Google Scholar] [CrossRef]
- Vesely, M.; Kershaw, M.; Schreiber, R.D.; Smyth, M. Natural Innate and Adaptive Immunity to Cancer. Annu. Rev. Immunol. 2011, 29, 235–271. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, J.; Benson, D.M. The role of natural killer cells in immunity against multiple myeloma. Leuk. Lymphoma 2012, 53, 1666–1676. [Google Scholar] [CrossRef]
- Caligiuri, M.A. Human natural killer cells. Blood 2008, 112, 461–469. [Google Scholar] [CrossRef]
- Matsuzaki, H.; Kagimoto, T.; Oda, T.; Kawano, F.; Takatsuki, K. Natural Killer Activity and Antibody-Dependent Cell-Mediated Cytotoxicity in Multiple Myeloma. Jpn. J. Clin. Oncol. 1985, 15, 611–617. [Google Scholar] [CrossRef]
- Komiyama, K.; Crago, S.; Itoh, K.; Moro, I.; Mestecky, J. Inhibition of natural killer cell activity by IgA. Cell. Immunol. 1986, 101, 143–155. [Google Scholar] [CrossRef]
- Gherman, M.; Manciulea, M.; Bancu, A.C. Regulation of human natural cytotoxicity by IgG—I. Characterization of the struc-tural site on monomeric IgG responsible for inhibiting natural killer cell activity. Int. Immunol. 1987, 24, 743–750. [Google Scholar]
- Frassanito, M.A.; Silvestris, F.; Cafforio, P.; Ma, F. IgG M-components in active myeloma patients induce a down-regulation of natural killer cell activity. Int. J. Clin. Lab. Res. 1997, 27, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Moţa, G.; Galatiuc, C.; Popescu, I.; Hirt, M.; Cialâcu, V.; Sulică, A. IgA monoclonal and polyclonal proteins as regulatory factors of the NK cytotoxic activity. Rom. J. Virol. 2001, 50, 17–31. [Google Scholar]
- Benson, D.M.; Yu, J.; Becknell, B.; Wei, M.; Freud, A.G.; Ferketich, A.K.; Trotta, R.; Perrotti, D.; Briesewitz, R.; Caligiuri, M.A. Stem cell factor and interleukin-2/15 combine to enhance MAPK-mediated proliferation of human natural killer cells. Blood 2009, 113, 2706–2714. [Google Scholar] [CrossRef] [Green Version]
- Becknell, B.; Caligiuri, M.A. Interleukin-2, Interleukin-15, and Their Roles in Human Natural Killer Cells. Adv. Immunol. 2005, 86, 209–239. [Google Scholar] [CrossRef]
- Kiessling, R.; Klein, E.; Wigzell, H. „Natural” killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype. Eur. J. Immunol. 1975, 5, 112–117. [Google Scholar] [CrossRef]
- Pratt, G.; Goodyear, O.; Moss, P. Immunodeficiency and immunotherapy in multiple myeloma. Br. J. Haematol. 2007, 138, 563–579. [Google Scholar] [CrossRef]
- Lu, Z.Y.; Bataille, R.; Poubelle, P.; Rapp, M.J.; Harousseau, J.L.; Klein, B. An interleukin 1 receptor antagonist blocks the IL-1-induced IL-6 paracrine production through a prostaglandin E2-related mechanism in multiple myeloma. Stem Cells 1995, 13, 28–34. [Google Scholar]
- Wu, J.; Gao, F.-X.; Wang, C.; Qin, M.; Han, F.; Xu, T.; Hu, Z.; Long, Y.; He, X.-M.; Deng, X.; et al. IL-6 and IL-8 secreted by tumour cells impair the function of NK cells via the STAT3 pathway in oesophageal squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 1–15. [Google Scholar] [CrossRef]
- Holt, D.; Ma, X.; Kundu, N.; Fulton, A. Prostaglandin E2 (PGE2) suppresses natural killer cell function primarily through the PGE2 receptor EP4. Cancer Immunol. Immunother. 2011, 60, 1577–1586. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Benson, D.M.; DeAngelis, S.; Bakan, C.E.; Li, P.-K.; Li, C.; Lin, J. A small molecule, LLL12 inhibits constitutive STAT3 and IL-6-induced STAT3 signaling and exhibits potent growth suppressive activity in human multiple myeloma cells. Int. J. Cancer 2011, 130, 1459–1469. [Google Scholar] [CrossRef] [Green Version]
- Tsuruma, T.; Yagihashi, A.; Hirata, K.; Torigoe, T.; Araya, J.; Watanabe, N.; Sato, N. Interleukin-10 Reduces Natural Killer (NK) Sensitivity of Tumor Cells by Downregulating NK Target Structure Expression. Cell. Immunol. 1999, 198, 103–110. [Google Scholar] [CrossRef]
- Martinet, L.; Jean, C.; Dietrich, G.; Fournié, J.-J.; Poupot, R. PGE2 inhibits natural killer and γδ T cell cytotoxicity triggered by NKR and TCR through a cAMP-mediated PKA type I-dependent signaling. Biochem. Pharmacol. 2010, 80, 838–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, H.; Nielsen, H.J.; Tvede, N.; Klarlund, K.; Mansa, B.; Moesgaard, F.; Drivsholm, A.; Nielsen, H.; Nielsen, H.J. Immune dysfunction in multiple myeloma. Reduced natural killer cell activity and increased levels of soluble interleukin-2 receptors. APMIS 1991, 99, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Mellor, A.L.; Munn, D.H. Tryptophan catabolism and T-cell tolerance: Immunosuppression by starvation? Immunol. Today 1999, 20, 469–473. [Google Scholar] [CrossRef]
- Della Chiesa, M.; Carlomagno, S.; Frumento, G.; Balsamo, M.; Cantoni, C.; Conte, R.; Moretta, L.; Moretta, A.; Vitale, M. The tryptophan catabolite l-kynurenine inhibits the surface expression of NKp46- and NKG2D-activating receptors and regulates NK-cell function. Blood 2006, 108, 4118–4125. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.R.; Carlson, L.M.; Koorella, C.; Rozanski, C.H.; Byrne, G.E.; Bergsagel, P.L.; Shaughnessy, J.P.; Boise, L.; Chanan-Khan, A.; Lee, K.P. CD28 Expressed on Malignant Plasma Cells Induces a Prosurvival and Immunosuppressive Microenvironment. J. Immunol. 2011, 187, 1243–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catlett-Falcone, R.; Landowski, T.H.; Oshiro, M.M.; Turkson, J.; Levitzki, A.; Savino, R.; Ciliberto, G.; Moscinski, L.; Fernández-Luna, J.L.; Nuñez, G.; et al. Constitutive Activation of Stat3 Signaling Confers Resistance to Apoptosis in Human U266 Myeloma Cells. Immunity 1999, 10, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Zwirner, N.W.; Fuertes, M.B.; Girart, M.V.; Domaica, C.I.; Rossi, L.E. Cytokine-driven regulation of NK cell functions in tumor immunity: Role of the MICA-NKG2D system. Cytokine Growth Factor Rev. 2007, 18, 159–170. [Google Scholar] [CrossRef]
- El-Sherbiny, Y.; Meade, J.L.; Holmes, T.D.; McGonagle, D.; Mackie, S.L.; Morgan, A.W.; Cook, G.; Feyler, S.; Richards, S.; Davies, F.; et al. The Requirement for DNAM-1, NKG2D, and NKp46 in the Natural Killer Cell-Mediated Killing of Myeloma Cells. Cancer Res. 2007, 67, 8444–8449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedel, R.; Thiery-Vuillemin, A.; Grandclement, C.; Balland, J.; Remy-Martin, J.-P.; Kantelip, B.; Pallandre, J.-R.; Pivot, X.; Ferrand, C.; Tiberghien, P.; et al. Novel Role for STAT3 in Transcriptional Regulation of NK Immune Cell Targeting Receptor MICA on Cancer Cells. Cancer Res. 2011, 71, 1615–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, E.; Neri, P.; Mesuraca, M.; Fulciniti, M.T.; Otsuki, T.; Pende, D.; Groh, V.; Spies, T.; Pollio, G.; Cosman, D.; et al. HLA class I, NKG2D, and natural cytotoxicity receptors regulate multiple myeloma cell recognition by natural killer cells. Blood 2005, 105, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinushi, M.; Vanneman, M.; Munshi, N.C.; Tai, Y.-T.; Prabhala, R.H.; Ritz, J.; Neuberg, D.; Anderson, K.C.; Carrasco, D.R.; Dranoff, G. MHC class I chain-related protein A antibodies and shedding are associated with the progression of multiple myeloma. Proc. Natl. Acad. Sci. USA 2008, 105, 1285–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Niu, G.; Kortylewski, M.; Burdelya, L.; Shain, K.; Zhang, S.; Bhattacharya, R.; Gabrilovich, D.; Heller, R.; Coppola, D.; et al. Regulation of the innate and adaptive immune responses by Stat-3 signaling in tumor cells. Nat. Med. 2004, 10, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Kortylewski, M.; Kujawski, M.; Wang, T.; Wei, S.; Zhang, S.; Pilon-Thomas, S.; Niu, G.; Kay, H.; Mulé, J.; Kerr, W.; et al. Inhibiting Stat3 signaling in the hematopoietic system elicits multicomponent antitumor immunity. Nat. Med. 2005, 11, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Perez-Andres, M.; Almeida, J.; Martín-Ayuso, M.; Moro, M.J.; Martín-Nuñez, G.; Galende, J.; Borrego, D.; Rodriguez, M.D.; Ortega, F.; Hernandez, J.; et al. Clonal plasma cells from monoclonal gammopathy of undetermined significance, multiple myeloma and plasma cell leukemia show different expression profiles of molecules involved in the interaction with the immunological bone marrow microenvironment. Leukemia 2005, 19, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Iwai, Y.; Ishida, M.; Tanaka, Y.; Okazaki, T.; Honjo, T.; Minato, N. Involvement of PD-L1 on tumor cells in the escape from host immune system and tumor immunotherapy by PD-L1 blockade. Proc. Natl. Acad. Sci. USA 2002, 99, 12293–12297. [Google Scholar] [CrossRef] [Green Version]
- Benson, D.M., Jr.; Bakan, C.E.; Mishra, A.; Hofmeister, C.; Efebera, Y.A.; Becknell, B.; Baiocchi, R.A.; Zhang, J.; Yu, J.; Smith, M.K.; et al. The PD-1/PD-L1 axis modulates the natural killer cell versus multiple myeloma effect: A therapeutic target for CT-011, a novel monoclonal anti–PD-1 antibody. Blood 2010, 116, 2286–2294. [Google Scholar] [CrossRef]
- Soriani, A.; Zingoni, A.; Cerboni, C.; Iannitto, M.L.; Ricciardi, M.R.; Di Gialleonardo, V.; Cippitelli, M.; Fionda, C.; Petrucci, M.T.; Guarini, A.; et al. ATM-ATR–dependent up-regulation of DNAM-1 and NKG2D ligands on multiple myeloma cells by therapeutic agents results in enhanced NK-cell susceptibility and is associated with a senescent phenotype. Blood 2009, 113, 3503–3511. [Google Scholar] [CrossRef] [Green Version]
- Von Lilienfeld-Toal, M.; Frank, S.; Leyendecker, C.; Feyler, S.; Jarmin, S.; Morgan, R.; Glasmacher, A.; Märten, A.; Schmidt-Wolf, I.G.H.; Brossart, P.; et al. Reduced immune effector cell NKG2D expression and increased levels of soluble NKG2D ligands in multiple myeloma may not be causally linked. Cancer Immunol. Immunother. 2010, 59, 829–839. [Google Scholar] [CrossRef]
- Fauriat, C.; Mallet, F.; Olive, D.; Costello, R. Impaired activating receptor expression pattern in natural killer cells from patients with multiple myeloma. Leukemia 2006, 20, 732–733. [Google Scholar] [CrossRef] [PubMed]
- Bernal, M.; Garrido, P.; Jiménez, P.; Carretero, R.; Almagro, M.; López, P.; Navarro, P.; Garrido, F.; Ruiz-Cabello, F. Changes in activatory and inhibitory natural killer (NK) receptors may induce progression to multiple myeloma: Implications for tumor evasion of T and NK cells. Hum. Immunol. 2009, 70, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Landowski, T.H.; Qu, N.; Buyuksal, I. Mutations in the Fas antigen in patients with multiple myeloma. Blood 1997, 90, 4266–4270. [Google Scholar] [CrossRef] [Green Version]
- Björklund, Å.K.; Forkel, M.; Picelli, S.; Konya, V.; Theorell, J.; Friberg, D.; Sandberg, R.; Mjösberg, J. The heterogeneity of human CD127+ innate lymphoid cells revealed by single-cell RNA sequencing. Nat. Immunol. 2016, 17, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Kini Bailur, J.; Mehta, S.; Zhang, L.; Neparidze, N.; Parker, T.L.; Bar, N.; Anderson, T.; Xu, M.; Dhodapkar, K.; Dhodapkar, M.V. Early Enrichment and Subsequent Dysfunction of Marrow ILC Subsets in Monoclonal Gammopathy: Target for IMiD Therapy. Blood 2017, 130 (Suppl. 1), 4370. [Google Scholar] [CrossRef]
- Wang, S.; Wu, P.; Chen, Y.; Chai, Y. Ambiguous roles and potential therapeutic strategies of innate lymphoid cells in different types of tumor (Review). Oncol. Lett. 2020, 20, 1513–1525. [Google Scholar] [CrossRef] [PubMed]
- Trabanelli, S.; Chevalier, M.F.; Martinez-Usatorre, A.; Gomez-Cadena, A.; Salomé, B.; Lecciso, M.; Salvestrini, V.; Verdeil, G.; Racle, J.; Papayannidis, C.; et al. Tumour-derived PGD2 and NKp30-B7H6 engagement drives an immunosuppressive ILC2-MDSC axis. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Yan, Y.; Su, Z.; Bie, Q.; Chen, X.; Barnie, P.A.; Guo, Q.; Wang, S.; Xu, H. Enhanced circulating ILC2s and MDSCs may contribute to ensure maintenance of Th2 predominant in patients with lung cancer. Mol. Med. Rep. 2017, 15, 4374–4381. [Google Scholar] [CrossRef] [Green Version]
- Guillerey, C.; Stannard, K.; Chen, J.; Krumeich, S.; Miles, K.; Nakamura, K.; Smith, J.; Yu, Y.; Ng, S.; Harjunpää, H.; et al. Systemic administration of IL-33 induces a population of circulating KLRG1 hi type 2 innate lymphoid cells and inhibits type 1 innate immunity against multiple myeloma. Immunol. Cell Biol. 2021, 99, 65–83. [Google Scholar] [CrossRef]
- Taylor, S.; Huang, Y.; Mallett, G.; Stathopoulou, C.; Felizardo, T.C.; Sun, M.-A.; Martin, E.L.; Zhu, N.; Woodward, E.L.; Elias, M.S.; et al. PD-1 regulates KLRG1+ group 2 innate lymphoid cells. J. Exp. Med. 2017, 214, 1663–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paiva, B.; Azpilikueta, A.; Puig, N.; Ocio, E.M.; Sharma, R.; Oyajobi, B.O.; Labiano, S.; San-Segundo, L.; Rodriguez, A.; Aires-Mejia, I.; et al. PD-L1/PD-1 presence in the tumor microenvironment and activity of PD-1 blockade in multiple myeloma. Leukemia 2015, 29, 2110–2113. [Google Scholar] [CrossRef]
- Geremia, A.; Arancibia-Cárcamo, C.V.; Fleming, M.P.; Rust, N.; Singh, B.; Mortensen, N.J.; Travis, S.P.; Powrie, F. IL-23–responsive innate lymphoid cells are increased in inflammatory bowel disease. J. Exp. Med. 2011, 208, 1127–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langowski, J.L.; Zhang, X.; Wu, L.; Mattson, J.D.; Chen, T.; Smith, K.; Basham, B.; McClanahan, T.; Kastelein, R.A.; Oft, M. IL-23 promotes tumour incidence and growth. Nat. Cell Biol. 2006, 442, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Wang, K.; Mucida, D.; Stewart, C.A.; Schnabl, B.; Jauch, D.; Taniguchi, K.; Yu, G.Y.; Osterreicher, C.H.; Hung, K.E.; et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumour growth. Nature 2012, 491, 254–258. [Google Scholar] [CrossRef] [Green Version]
- Tsirakis, G.; Pappa, C.A.; Kolovou, A.; Kokonozaki, M.; Neonakis, I.; Alexandrakis, M.G. Clinical significance of interleukin-22 in multiple myeloma. Hematology 2014, 20, 143–147. [Google Scholar] [CrossRef]
- Prabhala, R.H.; Pelluru, D.; Fulciniti, M.; Prabhala, H.K.; Nanjappa, P.; Song, W.; Pai, C.; Amin, S.; Tai, Y.-T.; Richardson, P.G.; et al. Elevated IL-17 produced by Th17 cells promotes myeloma cell growth and inhibits immune function in multiple myeloma. Blood 2010, 115, 5385–5392. [Google Scholar] [CrossRef]
- Gu, J.; Huang, X.; Zhang, Y.; Bao, C.; Zhou, Z.; Jin, J. Cytokine profiles in patients with newly diagnosed multiple myeloma: Survival is associated with IL-6 and IL-17A levels. Cytokine 2021, 138, 155358. [Google Scholar] [CrossRef]
- Munshi, N.C.; Anderson, J.L.D.; Shah, N.; Madduri, D.; Berdeja, J.; Lonial, S.; Raje, N.; Lin, Y.; Siegel, D.; Oriol, A.; et al. Idecabtagene Vicleucel in Relapsed and Refractory Multiple Myeloma. N. Engl. J. Med. 2021, 384, 705–716. [Google Scholar] [CrossRef]
- Marofi, F.; Saleh, M.M.; Rahman, H.S.; Suksatan, W.; Al-Gazally, M.E.; Abdelbasset, W.K.; Thangavelu, L.; Yumashev, A.V.; Hassanzadeh, A.; Yazdanifar, M.; et al. CAR-engineered NK cells; a promising therapeutic option for treatment of hematological malignancies. Stem Cell Res. Ther. 2021, 12, 1–18. [Google Scholar] [CrossRef]
- Seillet, C.; Brossay, L.; Vivier, E. Natural killers or ILC1s? That is the question. Curr. Opin. Immunol. 2021, 68, 48–53. [Google Scholar] [CrossRef]
- Michieletto, M.F.; Henao-Mejia, J. Ontogeny and heterogeneity of innate lymphoid cells and the noncoding genome. Immunol. Rev. 2021, 300, 152–166. [Google Scholar] [CrossRef] [PubMed]
- Cobb, L.M.; Verneris, M.R. Therapeutic manipulation of innate lymphoid cells. JCI Insight 2021, 6, 146006. [Google Scholar] [CrossRef] [PubMed]
- Di Censo, C.; Marotel, M.; Mattiola, I.; Müller, L.; Scarno, G.; Pietropaolo, G.; Peruzzi, G.; Laffranchi, M.; Mazej, J.; Hasim, M.S.; et al. Granzyme A and CD160 expression delineates ILC1 with graded functions in the mouse liver. Eur. J. Immunol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, G.; Ganter, S.; Bärenwaldt, A.; Finke, D. NKp46 Calibrates Tumoricidal Potential of Type 1 Innate Lymphocytes by Regulating TRAIL Expression. J. Immunol. 2018, 200, 3762–3768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazitt, Y. TRAIL is a potent inducer of apoptosis in myeloma cells derived from multiple myeloma patients and is not cytotoxic to hematopoietic stem cells. Leukemia 1999, 13, 1817–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krabbendam, L.; Heesters, B.A.; Kradolfer, C.M.; Spits, H.; Bernink, J.H. Identification of human cytotoxic ILC3s. Eur. J. Immunol. 2021, 51, 811–823. [Google Scholar] [CrossRef]
- Kim, J.; Kim, W.; Moon, U.J.; Kim, H.J.; Choi, H.-J.; Sin, J.-I.; Park, N.H.; Cho, H.R.; Kwon, B. Intratumorally Establishing Type 2 Innate Lymphoid Cells Blocks Tumor Growth. J. Immunol. 2016, 196, 2410–2423. [Google Scholar] [CrossRef] [Green Version]
- Rosiñol, L.; Jiménez, R.; Rovira, M.; Martínez, C.; Fernández-Avilés, F.; Marín, P.; Suárez-Lledó, M.; Gutiérrez-García, G.; De Larrea, C.F.; Carreras, E.; et al. Allogeneic hematopoietic SCT in multiple myeloma: Long-term results from a single institution. Bone Marrow Transplant. 2015, 50, 658–662. [Google Scholar] [CrossRef] [Green Version]
- Greil, C.; Engelhardt, M.; Ihorst, G.; Schoeller, K.; Bertz, H.; Marks, R.; Zeiser, R.; Duyster, J.; Einsele, H.; Finke, J.; et al. Allogeneic transplantation of multiple myeloma patients may allow long-term survival in carefully selected patients with acceptable toxicity and preserved quality of life. Haematologica 2018, 104, 370–379. [Google Scholar] [CrossRef] [Green Version]
- Depil, S.; Duchateau, P.; Grupp, S.A.; Mufti, G.; Poirot, L. ‘Off-the-shelf’ allogeneic CAR T cells: Development and challenges. Nat. Rev. Drug Discov. 2020, 19, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Munneke, J.M.; Björklund, A.T.; Mjösberg, J.; Garming-Legert, K.; Bernink, J.H.; Blom, B.; Huisman, C.; Van Oers, M.H.J.; Spits, H.; Malmberg, K.-J.; et al. Activated innate lymphoid cells are associated with a reduced susceptibility to graft-versus-host disease. Blood 2014, 124, 812–821. [Google Scholar] [CrossRef] [PubMed]
- Hanash, A.M.; Dudakov, J.; Hua, G.; O’Connor, M.H.; Young, L.F.; Singer, N.V.; West, M.L.; Jenq, R.R.; Holland, A.M.; Kappel, L.W.; et al. Interleukin-22 Protects Intestinal Stem Cells from Immune-Mediated Tissue Damage and Regulates Sensitivity to Graft versus Host Disease. Immunity 2012, 37, 339–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudakov, J.A.; Hanash, A.M.; Jenq, R.R.; Young, L.F.; Ghosh, A.; Singer, N.V.; West, M.L.; Smith, O.M.; Holland, A.M.; Tsai, J.J.; et al. Interleukin-22 Drives Endogenous Thymic Regeneration in Mice. Science 2012, 336, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Dudakov, J.A.; Mertelsmann, A.M.; O’Connor, M.H.; Jenq, R.R.; Velardi, E.; Young, L.F.; Smith, O.M.; Boyd, R.L.; Brink, M.R.M.V.D.; Hanash, A.M. Loss of thymic innate lymphoid cells leads to impaired thymopoiesis in experimental graft-versus-host disease. Blood 2017, 130, 933–942. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Zhu, X.; Wu, J.; He, L.; Lu, T.; Wang, Y.; Liu, B.; Ye, B.; Sun, L.; Fan, D.; et al. IL-13 secreted by ILC2s promotes the self-renewal of intestinal stem cells through circular RNA circPan3. Nat. Immunol. 2019, 20, 183–194. [Google Scholar] [CrossRef]
- Bruce, D.W.; Stefanski, H.E.; Vincent, B.G.; Dant, T.A.; Reisdorf, S.; Bommiasamy, H.; Serody, D.A.; Wilson, J.E.; McKinnon, K.P.; Shlomchik, W.D.; et al. Type 2 innate lymphoid cells treat and prevent acute gastrointestinal graft-versus-host disease. J. Clin. Investig. 2017, 127, 1813–1825. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Y.; Hughes, T.; Zhang, J.; Caligiuri, M.A.; Benson, D.M.; Yu, J. Fratricide of NK Cells in Daratumumab Therapy for Multiple Myeloma Overcome by Ex Vivo–Expanded Autologous NK Cells. Clin. Cancer Res. 2018, 24, 4006–4017. [Google Scholar] [CrossRef] [Green Version]
- Casneuf, T.; Xu, X.S.; Adams, H.C., 3rd; Axel, A.E.; Chiu, C.; Khan, I.; Ahmadi, T.; Yan, X.; Lonial, S.; Plesner, T.; et al. Effects of daratumumab on natural killer cells and impact on clinical outcomes in relapsed or refractory multiple myeloma. Blood Adv. 2017, 1, 2105–2114. [Google Scholar] [CrossRef]
- Lejeune, M.; Duray, E.; Peipp, M.; Clémenceau, B.; Baron, F.; Beguin, Y.; Caers, J. Balancing the CD38 Expression on Effector and Target Cells in Daratumumab-Mediated NK Cell ADCC against Multiple Myeloma. Cancers 2021, 13, 3072. [Google Scholar] [CrossRef] [PubMed]
- Malavasi, F.; Faini, A.C.; Morandi, F.; Castella, B.; Incarnato, D.; Oliviero, S.; Horenstein, A.L.; Massaia, M.; van de Donk, N.W.C.J.; Richardson, P.G. Molecular dynamics of targeting CD38 in multiple myeloma. Br. J. Haematol. 2021, 193, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Viola, D.; Dona, A.; Caserta, E.; Troadec, E.; Besi, F.; McDonald, T.; Ghoda, L.; Gunes, E.G.; Sanchez, J.F.; Khalife, J.; et al. Daratumumab induces mechanisms of immune activation through CD38+ NK cell targeting. Leukemia 2021, 35, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Motais, B.; Charvátová, S.; Walek, Z.; Hrdinka, M.; Smolarczyk, R.; Cichoń, T.; Czapla, J.; Giebel, S.; Šimíček, M.; Jelínek, T.; et al. Selection, Expansion, and Unique Pretreatment of Allogeneic Human Natural Killer Cells with Anti-CD38 Monoclonal Antibody for Efficient Multiple Myeloma Treatment. Cells 2021, 10, 967. [Google Scholar] [CrossRef] [PubMed]
- Kararoudi, M.N.; Nagai, Y.; Elmas, E.; Pereira, M.D.S.F.; Ali, S.A.; Imus, P.H.; Wethington, D.; Borrello, I.M.; Lee, D.A.; Ghiaur, G. CD38 deletion of human primary NK cells eliminates daratumumab-induced fratricide and boosts their effector activity. Blood 2020, 136, 2416–2427. [Google Scholar] [CrossRef]
- Sarkar, S.; Chauhan, S.K.S.; Daly, J.; Natoni, A.; Fairfield, H.; Henderson, R.; Nolan, E.; Swan, D.; Hu, J.; Reagan, M.R.; et al. The CD38low natural killer cell line KHYG1 transiently expressing CD16F158V in combination with daratumumab targets multiple myeloma cells with minimal effector NK cell fratricide. Cancer Immunol. Immunother. 2020, 69, 421–434. [Google Scholar] [CrossRef]
- Bigley, A.B.; Spade, S.; Agha, N.H.; Biswas, S.; Tang, S.; Malik, M.H.; Dai, L.; Masoumi, S.; Patiño-Escobar, B.; Hale, M.; et al. FcεRIγ-negative NK cells persist in vivo and enhance efficacy of therapeutic monoclonal antibodies in multiple myeloma. Blood Adv. 2021, 5, 3021–3031. [Google Scholar] [CrossRef]
- Shah, U.A.; Mailankody, S. CAR T and CAR NK cells in multiple myeloma: Expanding the targets. Best Pract. Res. Clin. Haematol. 2020, 33, 101141. [Google Scholar] [CrossRef]
- Holthof, L.C.; Stikvoort, A.; van der Horst, H.J.; Gelderloos, A.T.; Poels, R.; Li, F.; Groen, R.W.J.; Zweegman, S.; van de Donk, N.W.C.J.; O’Dwyer, M.; et al. Bone Marrow Mesenchymal Stromal Cell-mediated Resistance in Multiple Myeloma Against NK Cells can be Overcome by Introduction of CD38-CAR or TRAIL-variant. HemaSphere 2021, 5, e561. [Google Scholar] [CrossRef]
- Goodridge, J.P.; Bjordahl, R.; Mahmood, S.; Reiser, J.; Gaidarova, M.S.; Blum, M.R.; Cichocki, F.; Chu, H.-Y.; Bonello, G.; Lee, T.; et al. FT576: Multi-Specific Off-the-Shelf CAR-NK Cell Therapy Engineered for Enhanced Persistence, Avoidance of Self-Fratricide and Optimized Mab Combination Therapy to Prevent Antigenic Escape and Elicit a Deep and Durable Response in Multiple Myeloma. Blood 2020, 136, 4–5. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, W.; Shang, P.; Zhang, H.; Fu, W.; Ye, F.; Zeng, T.; Huang, H.; Zhang, X.; Sun, W.; et al. Transfection of chimeric anti-CD138 gene enhances natural killer cell activation and killing of multiple myeloma cells. Mol. Oncol. 2014, 8, 297–310. [Google Scholar] [CrossRef]
- Grosicki, S.; Bednarczyk, M.; Barchnicka, A.; Grosicka, O. Elotuzumab in the treatment of relapsed and refractory multiple myeloma. Future Oncol. 2021, 17, 1581–1591. [Google Scholar] [CrossRef]
- Chu, J.; Deng, Y.; Benson, D.M.; He, S.; Hughes, T.P.; Zhang, J.; Peng, Y.; Mao, H.; Yi, L.; Ghoshal, K.; et al. CS1-specific chimeric antigen receptor (CAR)-engineered natural killer cells enhance in vitro and in vivo antitumor activity against human multiple myeloma. Leukemia 2014, 28, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Leivas, A.; Valeri, A.; Córdoba, L.; García-Ortiz, A.; Ortiz, A.; Sánchez-Vega, L.; Graña-Castro, O.; Fernández, L.; Carreño-Tarragona, G.; Pérez, M.; et al. NKG2D-CAR-transduced natural killer cells efficiently target multiple myeloma. Blood Cancer J. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Zhu, H.; Blum, R.H.; Bjordahl, R.; Gaidarova, S.; Rogers, P.; Lee, T.T.; Abujarour, R.; Bonello, G.B.; Wu, J.; Tsai, P.-F.; et al. Pluripotent stem cell–derived NK cells with high-affinity noncleavable CD16a mediate improved antitumor activity. Blood 2020, 135, 399–410. [Google Scholar] [CrossRef]
- Williams, B.A.; Law, A.D.; Routy, B.; Denhollander, N.; Gupta, V.; Wang, X.-H.; Chaboureau, A.; Viswanathan, S.; Keating, A. A phase I trial of NK-92 cells for refractory hematological malignancies relapsing after autologous hematopoietic cell transplantation shows safety and evidence of efficacy. Oncotarget 2017, 8, 89256–89268. [Google Scholar] [CrossRef] [Green Version]
- Ducimetière, L.; Lucchiari, G.; Litscher, G.; Nater, M.; Heeb, L.; Nuñez, N.G.; Wyss, L.; Burri, D.; Vermeer, M.; Gschwend, J.; et al. Conventional NK cells and tissue-resident ILC1s join forces to control liver metastasis. Proc. Natl. Acad. Sci. USA 2021, 118, 2026271118. [Google Scholar] [CrossRef]
- Jacquelot, N.; Belz, G.T.; Seillet, C. Neuroimmune Interactions and Rhythmic Regulation of Innate Lymphoid Cells. Front. Neurosci. 2021, 15, 657081. [Google Scholar] [CrossRef]
- Oh, S.; Chun, S.; Hwang, S.; Kim, J.; Cho, Y.; Lee, J.; Kwack, K.; Choi, S.-W. Vitamin D and Exercise Are Major Determinants of Natural Killer Cell Activity, Which Is Age- and Gender-Specific. Front. Immunol. 2021, 12, 594356. [Google Scholar] [CrossRef] [PubMed]
- Poznanski, S.M.; Ashkar, A.A. What Defines NK Cell Functional Fate: Phenotype or Metabolism? Front. Immunol. 2019, 10, 1414. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.; Finlay, D.K. Optimising NK cell metabolism to increase the efficacy of cancer immunotherapy. Stem Cell Res. Ther. 2021, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, C.; Keam, S.P.; Yeang, H.X.A.; Neeson, M.; Richardson, K.; Hsu, A.K.; Canfield, R.; Bezman, N.; Robbins, M.; Quach, H.; et al. Myeloma natural killer cells are exhausted and have impaired regulation of activation. Haematologica 2021, 106, 2522–2526. [Google Scholar] [CrossRef] [PubMed]
- Pazina, T.; Macfarlane, A.W.; Bernabei, L.; Dulaimi, E.; Kotcher, R.; Yam, C.; Bezman, N.A.; Robbins, M.D.; Ross, E.A.; Campbell, K.S.; et al. Alterations of NK Cell Phenotype in the Disease Course of Multiple Myeloma. Cancers 2021, 13, 226. [Google Scholar] [CrossRef] [PubMed]
- Carlsten, M.; Korde, N.; Kotecha, R.; Reger, R.; Bor, S.; Kazandjian, D.; Landgren, O.; Childs, R.W. Checkpoint Inhibition of KIR2D with the Monoclonal Antibody IPH2101 Induces Contraction and Hyporesponsiveness of NK Cells in Patients with Myeloma. Clin. Cancer Res. 2016, 22, 5211–5222. [Google Scholar] [CrossRef] [Green Version]
- Benson, D.M.; Bakan, C.E.; Zhang, S.; Collins, S.M.; Liang, J.; Srivastava, S.; Hofmeister, C.; Efebera, Y.; Andre, P.; Romagne, F.; et al. IPH2101, a novel anti-inhibitory KIR antibody, and lenalidomide combine to enhance the natural killer cell versus multiple myeloma effect. Blood 2011, 118, 6387–6391. [Google Scholar] [CrossRef] [Green Version]
- Benson, D.M., Jr.; Hofmeister, C.C.; Padmanabhan, S.; Suvannasankha, A.; Jagannath, S.; Abonour, R.; Bakan, C.; Andre, P.; Efebera, Y.; Tiollier, J.; et al. A phase 1 trial of the anti-KIR antibody IPH2101 in patients with relapsed/refractory multiple myeloma. Blood 2012, 120, 4324–4333. [Google Scholar] [CrossRef]
- Alfarra, H.; Weir, J.; Grieve, S.; Reiman, T. Targeting NK Cell Inhibitory Receptors for Precision Multiple Myeloma Immunotherapy. Front. Immunol. 2020, 11, 575609. [Google Scholar] [CrossRef]
- Guia, S.; Fenis, A.; Vivier, E.; Narni-Mancinelli, E. Activating and inhibitory receptors expressed on innate lymphoid cells. Semin. Immunopathol. 2018, 40, 331–341. [Google Scholar] [CrossRef]
- Mariotti, F.R.; Quatrini, L.; Munari, E.; Vacca, P.; Moretta, L. Innate Lymphoid Cells: Expression of PD-1 and Other Checkpoints in Normal and Pathological Conditions. Front. Immunol. 2019, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zheng, N.; Luo, Q.; Jiang, L.; He, B.; Yuan, X.; Shen, L. Probiotics Lactobacillus reuteri Abrogates Immune Checkpoint Blockade-Associated Colitis by Inhibiting Group 3 Innate Lymphoid Cells. Front. Immunol. 2019, 10, 1235. [Google Scholar] [CrossRef]
- Pesce, S.; Trabanelli, S.; Di Vito, C.; Greppi, M.; Obino, V.; Guolo, F.; Minetto, P.; Bozzo, M.; Calvi, M.; Zaghi, E.; et al. Cancer Immunotherapy by Blocking Immune Checkpoints on Innate Lymphocytes. Cancers 2020, 12, 3504. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| NCT Number | Phase | Participants | Status | Location | Drug |
|---|---|---|---|---|---|
| NCT04634435 | 1/2 | 25 | Active and not yet recruiting | USA | Autologous cytokine-induced memory-like NK cells + KP12347: CD38 targeting antibody recruiting molecule + IL-2 |
| NCT04558931 | 2 | 60 | Active and not yet recruiting | Sweden | Activated autologous NK cells + Isatuximab |
| NCT03940833 | 1/2 | 20 | Recruiting | China | BCMA CAR-NK 92 cells |
| NCT04614636 | 1 | 105 | Recruiting | USA | FT538: CD38KO 158V CD16 + Daratumumab/Elotuzumab |
| NCT02727803 | 2 | 100 | Recruiting | USA | NK-92 after UCBT + Anti-thymocyte globulin |
| NCT04309084 | 1 | 29 | Recruiting | USA | CYNK-001: Placental CD56+/CD3- NK cells |
| NCT01729091 | 2 | 72 | Recruiting | USA | UCB NK cells after Elotuzumab/Lenalidomide/Melphalan before UCBT |
| NCT03019666 | 1 | 24 | Recruiting | USA | NAM-NK: Nicotinamide expanded haploidentical or mismatched related donor NK cells + IL-2 + Elotuzumab after lymphodepletion |
| NCT02890758 | 1 | 14 | Active, not recruiting | USA | Donor NK cells from healthy unmatched individuals + ALT-803: IL-15 superagonist |
| NCT01040026 | 1/2 | 10 | Active, not recruiting | Switzerland | Haploidentical NK cells after Melphalan + ASCT |
| NCT01619761 | 1 | 13 | Active, not recruiting | USA | UCB NK cells after Chemo + Lenalidomide +/− RT before UCBT |
| NCT04558853 | 1 | 12 | Active, not recruiting | Sweden | Autologous ex vivo expanded NK cells |
| NCT00720785 | 1 | 35 | Completed April 2021 | USA | Autologous expanded NK cells + Bortezomib |
| NCT02955550 | 1 | 15 | Completed June 2019 | USA | UCB NK + IL-2 after ASCT |
| NCT01313897 | 2 | 10 | Completed October 2016 | USA | Autologous expanded NK cells + Bortezomib |
| NCT02481934 | 1 | 5 | Completed October 2016 | Spain | NKAEs: Autologous ex vivo activated and expanded NK cell + Lenalidomide or Bortezomi |
| NCT01884688 | 2 | 3 | Completed October 2016 | USA | Autologous expanded NK cells + IL-2 |
| NCT00823524 | 1/2 | 47 | Completed February 2013 | South Korea | Donor NK cells after haploidentical familial donor BMT |
| NCT00660166 | 1 | 13 | Completed June 2012 | USA | Mismatched related donor NK cells after ASCT |
| NCT00990717 | 1 | 12 | Completed July 2012 | USA | NK 92 cells for RRMM of patients treated with ASCT |
| NCT00089453 | 1 | 10 | Completed May 2010 | USA | KIR-mismatched haploidentical donor NK cells + Bortezomib + Interleukin before ASCT |
| NCT00569283 | 1 | 18 | Completed December 2008 | South Korea | Donor NK cells after haploidentical familial donor BMT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szudy-Szczyrek, A.; Ahern, S.; Kozioł, M.; Majowicz, D.; Szczyrek, M.; Krawczyk, J.; Hus, M. Therapeutic Potential of Innate Lymphoid Cells for Multiple Myeloma Therapy. Cancers 2021, 13, 4806. https://doi.org/10.3390/cancers13194806
Szudy-Szczyrek A, Ahern S, Kozioł M, Majowicz D, Szczyrek M, Krawczyk J, Hus M. Therapeutic Potential of Innate Lymphoid Cells for Multiple Myeloma Therapy. Cancers. 2021; 13(19):4806. https://doi.org/10.3390/cancers13194806
Chicago/Turabian StyleSzudy-Szczyrek, Aneta, Sean Ahern, Magdalena Kozioł, Daria Majowicz, Michał Szczyrek, Janusz Krawczyk, and Marek Hus. 2021. "Therapeutic Potential of Innate Lymphoid Cells for Multiple Myeloma Therapy" Cancers 13, no. 19: 4806. https://doi.org/10.3390/cancers13194806
APA StyleSzudy-Szczyrek, A., Ahern, S., Kozioł, M., Majowicz, D., Szczyrek, M., Krawczyk, J., & Hus, M. (2021). Therapeutic Potential of Innate Lymphoid Cells for Multiple Myeloma Therapy. Cancers, 13(19), 4806. https://doi.org/10.3390/cancers13194806