Simple Summary
Pancreatic cancer is the seventh leading cause of cancer related death worldwide. In the United States, pancreatic cancer remains the fourth leading cause of cancer related death. The lack of early diagnosis and effective therapy contributes to the high mortality of pancreatic cancer. Therefore, there is an urgent need to find novel and effective biomarkers for the diagnosis and treatment of pancreatic cancer. Long noncoding RNA, circular RNAs and piwi-interacting RNA are non-coding RNAs and could become new biomarkers for the diagnosis, prognosis, and treatment of pancreatic cancer. We summarize the new findings on the roles of these non-coding RNAs in pancreatic cancer diagnosis, prognosis and targeted therapy.
Abstract
Pancreatic cancer is an aggressive malignance with high mortality. The lack of early diagnosis and effective therapy contributes to the high mortality of this deadly disease. For a long time being, the alterations in coding RNAs have been considered as major targets for diagnosis and treatment of pancreatic cancer. However, with the advances in high-throughput next generation of sequencing more alterations in non-coding RNAs (ncRNAs) have been discovered in different cancers. Further mechanistic studies have demonstrated that ncRNAs such as long noncoding RNAs (lncRNA), circular RNAs (circRNA) and piwi-interacting RNA (piRNA) play vital roles in the regulation of tumorigenesis, tumor progression and prognosis. In recent years, increasing studies have focused on the roles of ncRNAs in the development and progression of pancreatic cancer. Novel findings have demonstrated that lncRNA, circRNA, and piRNA are critically involved in the regulation of gene expression and cellular signal transduction in pancreatic cancer. In this review, we summarize the current knowledge of roles of lncRNA, circRNA, and piRNA in the diagnosis and prognosis of pancreatic cancer, and molecular mechanisms underlying the regulation of these ncRNAs and related signaling in pancreatic cancer therapy. The information provided here will help to find new strategies for better treatment of pancreatic cancer.
1. Introduction
Non-coding RNAs (ncRNAs) are short or long single strands of linear or circular RNA molecules, which have no ability to be translated into proteins [1,2,3,4]. The most abundant ncRNAs are ribosome RNA (rRNA) and transfer RNA (tRNA), which are critically involved in the process of translation and serve as house-keeping gene. Other ncRNAs can be categorized into long non-coding RNA (lncRNA) and small non-coding RNA according to their length (Figure 1). Small non-coding RNAs are usually shorter than 200 nt (nucleotides) and include microRNA (miRNA), small interfering RNA (siRNA), Piwi-interacting RNA (piRNA), small nucleolar RNA (snoRNA), small nuclear RNA (snRNA) and guide RNA (gRNA). Mechanistic studies have shown that these small ncRNAs play important roles in the regulation of transcription, post-transcription, and translation [2,3,4]. lncRNAs are RNA molecules which have lengths exceeding 200 nt and are not translated into proteins. One of the important functions of lncRNAs is epigenetic, transcriptional, and the post-transcriptional regulation of gene expression [5]. In addition to these long and small linear ncRNAs, there is another type of ncRNA molecule which forms a covalently closed loop, and it is named circular RNA (circRNA). The size of circRNA can range from under 100 nucleotide to over 4 kilobases. Similar as miRNAs and lncRNAs, circRNAs have recently received much attention in the field of cancer research because of their abilities to regulate gene expression [6,7,8].
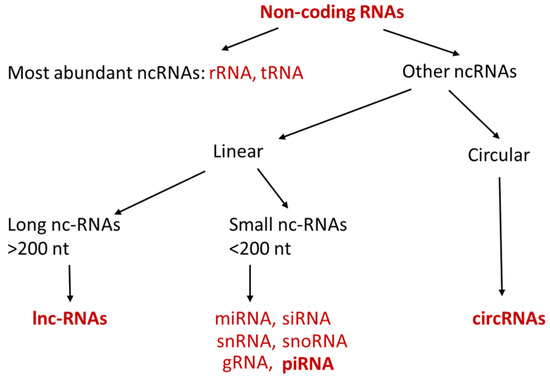
Figure 1.
Types of ncRNAs. ncRNAs can be categorized into linear (long and small) and circular ncRNAs. ncRNAs in bold are discussed in the below sections.
2. Biogenesis and Functions of lncRNA, circRNA and piRNA
2.1. lncRNA
Among the ncRNAs discovered, miRNAs are the most intensively studied and the roles of miRNAs in physiological and pathological regulations are extensively reported [6,7,9]. However, lncRNAs are also widely investigated because aberrant expression levels of lncRNAs are frequently found in various types of cancers [10,11,12]. H19 RNA was the first reported lncRNA in mammalian genome [13], followed by other lncRNA such as X-inactive-specific transcript [14]. In an earlier international collaborative study on mouse cDNAs, it was found that 11,665 cDNA are new non-coding messages and these ncRNAs represent a major component of the transcriptome [15]. From then on, lncRNAs were recognized and more lncRNAs were found through high-throughput genome analysis. Most lncRNAs are transcribed by RNA polymerase II, polyadenylated at 3′ end, spliced or un-spliced and capped at 5′ end [12,16] (Figure 2). The sequences of lncRNAs are either sense or antisense compared to protein-coding genes. The locations of lncRNA sequences are within introns or intergenic regions of genes. lncRNAs have diverse cellular functions including transcriptional regulation, epigenetic gene regulation, chromatin remodeling, cis and trans gene expression, etc. [5,12,17,18]. Because of the diverse functions of lncRNAs, the aberrant levels of lncRNAs in pathological status including cancers and inflammations have received much attention in the field of cancer research [2,5,19].
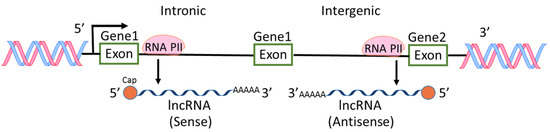
Figure 2.
Biogenesis of lncRNAs. lncRNAs are transcribed by RNA polymerase II (RNA PII) intronically and intergenically. The sequences of lncRNAs are either sense or antisense compared to protein-coding genes.
2.2. circRNA
circRNAs are a sub-group of ncRNAs and play significant roles in the regulation of transcription and post-transcription [20,21]. The first report on circRNA was published 40 years ago [22]. In the report, viroids were recognized as covalently closed circular RNA molecules. circRNAs are derived from pre-mRNAs; however, circRNAs lack the ability as mRNA to translate to protein. circRNAs can be generated from exons, introns, intergenic regions, 3′-UTR or 5′-UTR, but most circRNAs are derived from exons through spliceosomal and other machineries [20,21,22,23,24] (Figure 3). The investigations on circRNA production showed that RNA splicing such as exon skipping and back-splicing is important for circRNA creation [20,21,22,23,24]. The fragments of RNA created by RNA splicing form a closed circular nucleotide structure connected by covalent bond (Figure 3). Because of the circular structure, circRNAs are more stable than linear mRNAs. With the development of RNA-sequencing technology, new circRNAs have been discovered rapidly. The mechanisms of circRNAs regulating cellular signaling could include interaction with RNA binding proteins (RBPs), sponge with miRNAs, and competing with endogenous RNAs or RNA splicing, leading to posttranscriptional regulation of gene expression and protein translation [20,21,23]. In addition, most circRNAs are evolutionarily conserved and stable [25]. Any alternations in the expression level of specific types of circRNAs could reflect a pathological state. Therefore, circRNAs could be used as biomarkers for cancer diagnosis and treatment, and are considered a hot topic in cancer research [7,8,26,27].
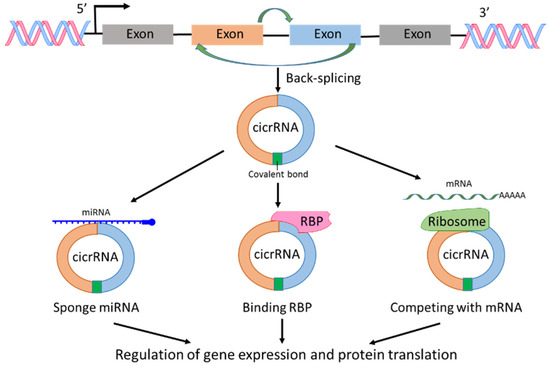
Figure 3.
Biogenesis and functions of circRNAs. circRNA is produced by back-splicing and the fragments of RNA form a closed circular nucleotide structure connected by covalent bond. circRNAs interact with RNA binding protein (RBP), sponge with miRNAs and compete with mRNAs, leading to regulation of gene expression and protein translation.
2.3. piRNAs
piRNAs are a class of short (about 21 to 35 nucleotides) single-stranded RNAs. They interact with PIWI proteins to silence genetic elements such as transposons in the maintenance of genome stability and integrity [28,29,30,31]. Four PIWI proteins exist in humans: PIWIL1 (also known as HIWI), PIWIL2 (also known as HILI), PIWIL3 (also known as HIWI3), and PIWIL4 (as known as HIWI2) [32]. PIWI proteins play critical roles in the control of cell proliferation, apoptosis, cell movement, and genomic stability [32]. In 2006, piRNAs were first identified and named as “PIWI-interacting RNAs” in a mouse study on PIWI proteins and genome stability [33]. From then on, more piRNAs were identified based on their association with PIWI proteins and their biogenesis [34,35]. Two steps exist in the process of piRNA biogenesis [28]. First, a single-strand primary piRNA is transcripted by RNA polymerase II (Pol II) from a specific genomic location known as piRNA clusters (Figure 4). Through binding to a RNA-binding protein MAEL, the primary piRNA is transported into cytoplasm where the primary piRNA undergoes 5′ and 3′-end modification with MOV10L1 and Tdrkh [28,36,37], leading to increased stability and affinity to PIWI protein [38]. Then, primary piRNAs go through a ping-pong cycle to guide PIWI protein mediated cleavage of transcripts and create secondary piRNAs (Figure 4). The most investigated function of piRNA is its suppression of transposable elements (TEs) through association with PIWI proteins [28,39]. However, piRNAs could also regulate endogenous gene expression in stem cells during development [40,41]. piRNA regulates mRNA mainly through RNA degradation. piRNAs combined with MIWI (one of mouse PIWI proteins) could target mRNAs with imperfect base pairing and induce mRNA degradation through MIWI-dependent cleavage [42,43]. Although piRNAs play crucial roles in germ cells and adult stem cells, growing evidences have shown that piRNA and PIWI proteins are involved in tumorigenesis [44,45,46,47]. Recently, more investigations are focused on the roles of piRNAs in cancer development and progression [45,48,49,50,51]. Because piRNAs regulate gene expression and genome stability in stem cells, any deregulation of piRNAs could cause the development of cancer stem cells and promote cancer progression.
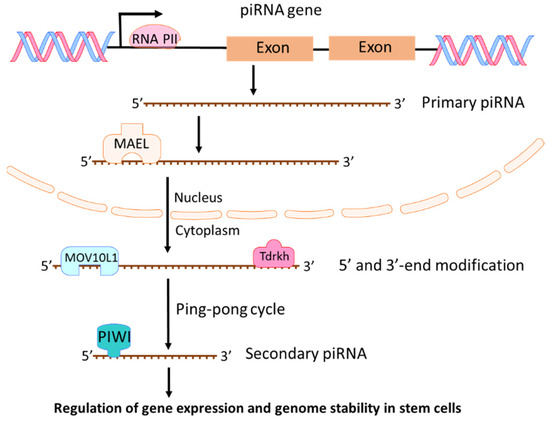
Figure 4.
Biogenesis and functions of piRNAs. Primary piRNA is transcripted by RNA polymerase II (Pol II) from piRNA genes. By binding to MAEL, the primary piRNA is transported into cytoplasm where the primary piRNA undergoes 5′ and 3′-end modification and a ping-pong cycle to create secondary piRNAs. piRNAs can regulate gene expression and genome stability in stem cells.
3. Major lncRNAs in Pancreatic Cancer
Because of the diverse biofunctions of lncRNAs, the roles of lncRNAs in pancreatic cancers can be oncogenic or tumor suppressive. Among the growing number of lncRNAs found in pancreatic cancers, in this section, we summarized some important lncRNAs which play critical roles in pancreatic cancer (Table 1).

Table 1.
Major lncRNAs in pancreatic cancer.
3.1. HOTAIR
lncRNA HOTAIR (HOX transcript antisense RNA) is an oncogene in pancreatic cancer [52]. It was found that a higher level of HOTAIR had strong associations with susceptibility of pancreatic cancer [52]. A high expression of HOTAIR was also associated with proliferation and the metastasis of pancreatic cancers [55]. HOTAIR could transcriptionally regulate the expression of hundreds of genes in both PRC2-dependent and PRC2-independent manner in pancreatic cancer cells [53]. HOTAIR could also couple with EZH2 to silence tumor suppressor miR-34a in pancreatic cancer cells, thereby inducing cancer cell proliferation [54], suggesting the regulatory role of HOTAIR in EZH2/EMT pathway. Moreover, studies showed that HOTAIR enhanced pancreatic cancer resistance to gemcitabine and TNF-related apoptosis-inducing ligand, causing chemoresistance and progression of pancreatic cancer [56,57]. In addition, HOTAIR could also regulate the sensitivity of radiotherapy. Knockdown of HOTAIR enhanced the radiosensitivity of pancreatic cancer [95]. Furthermore, HOTAIR expression was associated with more aggressive pancreatic cancer, suggesting its prognostic role in pancreatic cancer [53].
3.2. MALAT1
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT-1) is highly conserved and ubiquitously expressed lncRNA [96,97]. MALAT-1 was firstly discovered in non-small cell lung cancer [96,97]. However, high expression of MALAT-1 has been found in various cancer tissues including pancreatic cancer [63,98,99]; therefore, MALAT-1 is considered as an oncogenic lncRNA. In pancreatic cancer, increased expression of MALAT-1 promoted cell growth, migration, and invasion [59]. MALAT-1 also transcriptionally regulated Sox-2 expression and enhanced stem cell-like phenotypes in pancreatic cancer cells, suggesting its role in pancreatic stemness and tumorigenesis [60]. A study also showed that MALAT-1 promoted aggressive pancreatic cancer proliferation and metastasis via stimulation of autophagy [61]. MALAT-1 could also exert its oncogenic effects through mediation of EZH2, miR-216a, miR-217, miR-200c, or Hippo-YAP signaling in pancreatic cancers [58,62,64,65,66]. Therefore, MALAT1 regulates miRNAs, mRNAs, and proteins in stemness and autophagy pathways in pancreatic cancer. Importantly, overexpression of MALAT-1 has been correlated with advanced tumor stages, metastasis, and poor survival in pancreatic cancers [63,100].
3.3. GAS5
lncRNA growth arrest-specific 5 (GAS5) is a tumor suppressor found in various cancers including pancreatic cancer [101,102]. In pancreatic cancer, downregulation of GAS5 increased cell proliferation by regulating CDK6 transcriptionally, suggesting the tumor suppressive role of GAS5 [89]. GAS5 also reversed EMT, inhibited metastasis and increased the sensitivity of pancreatic cancer stem cells to gemcitabine through targeting miR-221/SOCS3 signaling [88]. Thus, GAS5 has a role in the regulation of cell cycle and EMT pathways. Studies also showed that GAS5 suppressed pancreatic cancer metastasis and chemoresistance through epigenetic regulation of miR-32 or miR-181c [86,87]. All findings demonstrate that GAS5 is a tumor suppressive lncRNA.
3.4. MEG3
MEG3 is another tumor suppressive lncRNA in cancers including pancreatic cancer [103]. The expression levels of MEG3 in both pancreatic cancer tissues and cells were found to be much lower than that in normal tissues and cells [92]. Knockdown of MEG3 promoted pancreatic cancer cell proliferation, migration, and invasion [92] while overexpression of MEG3 suppressed pancreatic neuroendocrine tumor cell viability, invasion, and migration [94]. The mechanisms underlying the tumor suppressive effects of MEG3 could be mediated through epigenetic regulation of c-Met. MEG3 was found to bind to unique genomic regions in and around c-Met gene and inhibit c-Met expression, leading to tumor suppression [91,93]. In addition, MEG3 could also exert its anti-cancer effects on pancreatic cancer by regulation of the PI3K/AKT signaling pathway [90]. Therefore, MEG3 affects multiple cellular signaling in pancreatic cancer.
3.5. H19
H19 is an oncogenic lncRNA which antagonizes tumor suppressive let-7 [104]. In pancreatic cancer, the expression of H19 was significantly increased and the overexpression of H19 was correlated with histological grade and invasion of pancreatic cancer [68]. Knockdown of H19 in pancreatic cancer cells inhibited cell proliferation and tumor growth with G0/G1 arrest and downregulation of E2F-1 transcription factor [68]. Knockdown of H19 also inhibited metastasis of pancreatic cancer [72]. It was found that H19 promoted pancreatic cancer cell invasion and migration by upregulation of HMGA2-mediated EMT through antagonizing let-7 [67], demonstrating the critical role of H19/let-7/HMGA2/EMT signaling axis in pancreatic cancer progression. In addition to the association of H19 with let-7, H19 could also correlate with miR-675 or miR194 to modulate EMT, cell proliferation, migration, and metastasis of pancreatic cancer [69,70,71], suggesting its role in the regulation of the EMT pathway.
3.6. PVT1
PVT1 is an oncogenic lncRNA [105]. In pancreatic cancer, higher expression of PVT1 was found in cancer tissues, and the high expression of PVT1 was positively correlated with poor survival of patients [73,76]. PVT1 promoted cell proliferation and migration through transcriptional and epigenetic regulation of p21 and miR-448 [78,80]. PVT1 also promoted autophagy and cell growth by regulating miR-20a-5p and ULK1 signaling, leading to the development of pancreatic cancer [74]. More importantly, PVT1 was identified as a regulator of gemcitabine sensitivity [77,79,81]. Functional inactivation of PVT1 resulted in the enhanced sensitivity to gemcitabine in pancreatic cancers [79]. In addition, PVT1 also promoted pancreatic cancer development through regulation of miR519, HIF-1, YKT6, RAB7, and VAMP3 [75,76], suggesting its diverse oncogenic effects with multiple signaling regulation.
3.7. HOTTIP
HOXA transcript at the distal tip (HOTTIP) is another oncogenic lncRNA [106]. HOTTIP is located at the 5′ end of HOXA cluster [106]. The expression of HOTTIP is increased in pancreatic cancer [84]. It was found that ectopic HOTTIP expression promoted growth and invasiveness in pancreatic ductal adenocarcinoma [84] and that HOTTIP modulated pancreatic cancer stem cell properties by regulating HOXA9 epigenetically [82]. More importantly, HOTTIP could also regulate drug resistance through regulation of miR-137 and HOXA13 [83,85], suggesting its role in HOXA pathway.
4. circRNAs in Pancreatic Cancer
In recent years, studies on circRNAs in cancer have become a hot topic in cancer research, especially after advanced next-generation sequencing technology was developed [7,21,107]. However, compared to other types of cancers, research on circRNAs in pancreatic cancer is still in its early stages. By conducting expression profiling of circRNAs using pancreatic cancer tissues and further mechanistic studies, it was found that some circRNAs are oncogenic while others are tumor suppressive in pancreatic cancers [108,109] (Table 2).
Table 2.
Major circRNAs in pancreatic cancer.
4.1. circRNA Expression Profiling in Pancreatic Cancer
In 2015, a group of investigators reported for the first time the microarray analysis of circRNA profiles in six pairs of pancreatic ductal adenocarcinoma and adjacent normal tissues [108]. They further analyzed the circRNA expression profile and identified a class of circRNAs that was responsible for tumorigenesis of pancreatic cancer. The top five up-regulated circRNAs in pancreatic cancers were hsa_circ_0072088, hsa_circ_0030235, hsa_circ_0001946, hsa_circ_0060055 and hsa_circ_0005397 whereas the top five down-regulated circRNAs were hsa_circ_0013587, hsa_circ_0075410, hsa_circ_0008768, hsa_circ_0080712, and hsa_circ_0000257 [122]. GO enrichment and pathway analysis showed that the most significantly altered circRNAs in pancreatic cancer were related to small GTPase regulator activity, Ras GTPase binding, RNA binding, and VEGF signaling [122], which have been known to be significantly altered in pancreatic cancer [123,124,125]. These findings suggest that these circRNAs could be molecular targets for the treatment of pancreatic cancer.
In 2016, another group of investigators conducted a microarray analysis of circRNA profiles in 20 pairs of pancreatic ductal adenocarcinoma and adjacent normal tissues and deposited the data in GEO database (GEO79634) [109]. The top 10 differently expressed circRNAs were identified in this study. Among them, five circRNAs (hsa_circ_102051, hsa_circ_102619, hsa_circ_104270, hsa_circ_102049 and hsa_circ_104227) were up-regulated while five circRNAs (hsa_circ_000167, hsa_circ_103809, hsa_circ_104700, hsa_circ_001846 and hsa_circ_102728) were down-regulated. They also found that a specific set of circRNAs (hsa_circRNA_100435, hsa_circRNA_103076, hsa_circRNA_103309, hsa_circRNA_000780, hsa_circRNA_101252, hsa_circRNA_102374, hsa_circRNA_104433, hsa_circRNA_104882, hsa_circRNA_101717, hsa_circRNA_104084, hsa_circRNA_100646 and hsa_circRNA_102213) could bind to miR-15a or miR-505 to modulate gene expression in pancreatic cancer [109]. miR-15a and miR-506 have been found to inhibit cell proliferation, EMT, and chemoresistance in pancreatic cancer [126,127].
Because circRNAs can function as miRNA sponges, the above mentioned circRNA profiling data from two sets of pancreatic tissues were further analyzed with miRNA profiling data. In the study, circRNA and miRNA interactions were predicted between differentially expressed circRNAs and miRNAs using computerized analysis [128]. A total of 51 interactions between circRNAs and miRNAs were found. Further analysis showed that mitogen-activated protein kinase, PI3K/AKT, and WNT/β-catenin signaling pathways were associated with the development of pancreatic cancer [128].
In addition to the microarray technique used for circRNA profiling, high-throughput circRNA-sequencing has also been utilized for circRNA expression profiling [129,130]. We have conducted a circRNA-sequencing experiment to obtain the circRNA expression profiles before and after thymoquinone treatment in pancreatic cancer cells. By circRNA-sequencing, we were able to identify circRNAs which were altered by thymoquinone, a natural product with potential anti-cancer activity. Gene enrichment analysis showed that the altered circRNAs were associated with binding of proteins, metal ions, nucleotide, ATP, and RNAs, and correlated with Wnt and Hedgehog signaling pathways (Figure 5), demonstrating the inhibitory effects of thymoquinone on pancreatic cancer. Another group of investigators also utilized circRNA-sequencing to find novel mechanisms of a potential anti-cancer drug nigericin in the treatment of pancreatic cancer [131]. From the profiling data, they found that nigericin could regulate focal adhesion, TNF, MAPK, PI3K-Akt, and HIF-1 signaling pathway, pyrimidine metabolism and purine metabolism through modulation of circRNA-miRNA-mRNA network [131].
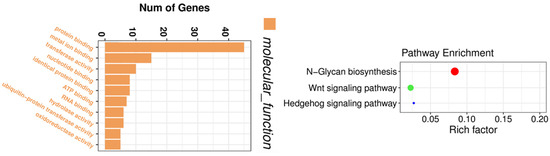
Figure 5.
Gene enrichment analysis on circRNAs altered by thymoquinone. The altered circRNAs were associated with the binding of proteins, metal ions, nucleotide, ATP, and RNAs, and correlated with Wnt and Hedgehog signaling pathways.
4.2. Oncogenic circRNAs
4.2.1. ciRS-7
ciRS-7 is an oncogenic circRNA which acts as a sponge of miR-7 [132]. A recent study found that ciRS-7 expression was significantly higher in pancreatic cancer tissues compared to adjacent pancreatic tissues [110]. Moreover, ciRS-7 expression was significantly increased in the area of invasion and metastasis of pancreatic cancer. A mechanistic study showed that knockdown of ciRS-7 inhibited the proliferation and invasion of pancreatic cancer cells with up-regulation of miR-7 through releasing sponge and down-regulation of EGFR and STAT3 pathways by miR-7 [110].
4.2.2. CircEIF6
Oncogenic circRNA circEIF6 (hsa_circ_0060055) was found to be aberrantly up-regulated in pancreatic tumor tissues and cells [111]. Silence of circEIF6 significantly induced apoptosis and inhibited pancreatic cancer cell proliferation, migration, and invasion [111]. A mechanistic study demonstrated that circEIF6 bond to miR-557 led to the up-regulation of SLC7A11 and activation of PI3K/AKT signaling in pancreatic cancer cells [111]. An animal study showed that knockdown of circEIF6 significantly suppressed pancreatic tumor xenograft growth in vivo [111]. These findings suggest that circEIF6 exerts its oncogenic effects through miR-557/SLC7A11/PI3K/AKT signaling transduction.
4.2.3. CircFOXK2
circFOXK2 is an oncogenic circRNA in pancreatic cancer [112]. circFOXK2 was significantly upregulated in pancreatic cancer cells and tissues, leading to increased cell proliferation, migration, and invasion [112]. circFOXK2 acted as a sponge for miR-942, causing the altered expression of downstream target ANK1, GDNF, and PAX6, which could play an oncogenic effect in pancreatic cancer [133]. circFOXK2 also bonded to RNA-binding protein YBX1 and hnRNPK, enhancing the expression of oncogenes NUF2 and PDXK. By interactions with miR-942 and YBX1/hnRNPK, circFOXK2 promoted the progression of pancreatic cancers [112].
4.2.4. circRNA_100782 and circ_001653
A study found that circRNA_100782 was significantly upregulated in pancreatic cancers [117]. Inhibition of circRNA_100782 suppressed cell proliferation and colony formation through downregulation of IL6R and STAT3, which are oncogenes in pancreatic cancer [134]. Luciferase assay showed that miR-124 was a direct target of circRNA_100782. An animal study showed that knockdown of circRNA_100782 inhibited pancreatic cancer xenografts in nude mice [117]. Therefore, it is believed that circRNA_100782 exerts its oncogenic effects through sponging miR-124 to activate IL6/STAT3 signaling [117]. Similarly, circ_001653 was upregulated in pancreatic cancers [135]. Inhibition of hsa_circ_001653 by siRNA suppressed pancreatic cancer cell proliferation and invasion through sponging miR-377, leading to the downregulation of oncogenic HOXC6 expression [135].
4.2.5. hsa_circ_0071036 and hsa_circ_0007534
Recent study showed that hsa_circ_0071036 plays important roles as oncogenic circRNA in tumorigenesis and progression of pancreatic cancers [118]. A mechanistic study showed that hsa_circ_0071036 acted as an efficient sponge for tumor suppressive miR-489 in pancreatic cancer. An in vivo animal study demonstrated that knockdown of hsa_circ_0071036 significantly inhibited pancreatic cancer growth in mice [118]. Similarly, hsa_circ_0007534 was also found to be up-regulated in pancreatic cancer tissues and cells [119]. hsa_circ_0007534 could inhibit pancreatic cancer cell apoptotic death through modulation of Bcl-2/caspase-3 by sponging tumor suppressive miR-625 and miR-892b [119].
4.2.6. circBFAR
circBFAR (hsa_circ_0009065) was found to be upregulated in pancreatic cancer tissues [113]. Moreover, the high expression of circBFAR was correlated with high TNM stage and poor prognosis of patients with pancreatic cancer. Mechanistic study showed that circBFAR promoted expression of MET through sponging tumor suppressive miR-34b-5p, leading to the activation of MET/PI3K/Akt signaling. Importantly, knockdown of circBFAR significantly suppressed pancreatic cancer cell proliferation and motility in vitro and inhibited pancreatic tumor growth and metastasis in mouse model, demonstrating its oncogenic function of circBFAR [113].
4.2.7. circ-ASH2L
circ-ASH2L was highly up-regulated in pancreatic cancer cells and tissues [114]. The high expression of circ-ASHL was positively correlated with lymphatic invasion and TNM stage. A mechanistic study showed that circ-ASH2L induced cell proliferation, tumor invasion, and angiogenesis by sponging miR-34a, which is a tumor suppressive miRNA that suppresses Notch 1. Therefore, by sponging miR-34a, circ-ASH2L enhanced the expression of Notch1, one of the oncogenic signals, leading to tumor progression and poor survival of patients with pancreatic cancer [114].
4.2.8. circRHOT1
circRHOT1 (hsa_circ_0005397) is highly expressed in pancreatic cancer and is mainly located in the cytoplasm of pancreatic cancer cells [115,116]. Down-regulation of circRHOT1 suppressed pancreatic cell proliferation, invasion, and migration. The effects of circRHOT1 could be mediated through sponging miR-26b, miR-125a, miR-330, and miR-382, which could be tumor suppressive miRNAs in pancreatic cancer [116]. A similar study also showed that circRHOT1 served as a sponge and bond to miR-125a-3p, upregulating oncogenic E2F3. Knockdown of circRHOT1 significantly suppressed pancreatic cancer cells through the regulation miR-125a-3p/E2F3 axis [115]. These findings suggest the oncogenic effects of circRHOT1.
4.3. Tumor Suppressive circRNAs
4.3.1. circNFIB1
circNFIB1 (hsa_circ_0086375) is a tumor suppressive circRNA in pancreatic cancer [120]. circNFIB1 is downregulated in pancreatic cancer tissues and negatively associated with lymph node metastasis [120]. A study found that knockdown of circNFIB1 enhanced lymph node metastasis of pancreatic cancer both in vitro and in vivo [120]. A mechanistic study showed that circNFIB1 acted as an anti-cancer sponge of oncogenic miR-486-5p. By binding to miR-486-5p, circNFIB1 induced expression of PIK3R1 and, in turn, inhibited the expression of VEGF-C through inactivating PI3K/Akt signaling [120]. An animal study confirmed that circNFIB1 regulated miR-486-5p/PIK3R1/VEGF-C signal axis, leading to the inhibition of lymph node metastasis in pancreatic cancer [120].
4.3.2. hsa_circ_001587
hsa_circ_001587 is another tumor suppressive circRNA in pancreatic cancer [121]. The expression of hsa_circ_001587 was significantly down-regulated in pancreatic cancer tissues and cells [121]. Overexpression of hsa_circRNA_001587 significantly decreased abilities of pancreatic cancer cell proliferation, migration, invasion, angiogenesis and tumorigenesis through inhibition of oncogenic MMP-2, MMP-9, MCM2, and VEGF expression in pancreatic cancer [121]. Mechanistic studies showed that hsa_circRNA_001587 bond to oncogenic miR-223 and, in turn, up-regulated SLC4A4 [121]. Therefore, the tumor suppressive effects of hsa_circ_001587 is mediated through the regulation of miR-223/SLC4A4/MMP signaling pathway.
4.3.3. hsa_circ_0001649
Similarly to other tumor suppressive circRNA, hsa_circ_0001649 was found to be down-regulated in pancreatic cancer tissues and cells [136]. Moreover, the down-regulated hsa_circ_0001649 was associated with the clinical tumor stage of the patients with pancreatic cancer [136]. Study found that overexpression of hsa_circ_0001649 induced apoptotic cell death and inhibited cell proliferation and colony formation in pancreatic cancer cells [136], suggesting the tumor suppressive effects of hsa_circ_0001649 in pancreatic cancer. However, the mechanism underlying hsa_circ_0001649 as a tumor suppressive remains unclear.
5. piRNA in Pancreatic Cancer
In recent years, more studies on the roles of piRNAs and PIWI proteins in the development and progression of breast, lung, and gastric cancers have been reported [47,48,50,137,138]. Because piRNAs regulate gene expression and genome stability in stem cells [40,41], any deregulation of piRNAs could cause the development of cancer stem cells and promote cancer progression. A recent study showed that piRNA-651 promoted proliferation and migration of breast cancer cells through epigenetic regulation of PTEN promoter inactivation [48]. It was also found that piRNA-211106 could serve as tumor suppressor to inhibit the progression of lung cancer and enhance the chemotherapy sensitivity of cancer cells [49]. In gastric cancer, piRNA-823 was found to be significantly down-regulated and induction of piRNA-823 inhibited cancer cell growth in vitro and in vivo [139]. These results suggest the importance of piRNAs in cancer. However, the reported research on the roles of piRNA and PIWI proteins in pancreatic cancer is limited.
5.1. Genotyping and piRNA Profiling in Pancreatic Cancer
In a recent study on the genetic mechanism of the co-effects of single nucleotide polymorphisms (SNP) and piRNA in cancers, a group of investigators conducted expression quantitative trait locus (eQTL) analysis using data including 10,997 samples across 33 cancer types from The Cancer Genome Atlas (TCGA) [140]. They identified millions of SNP-piRNA pairs in tumor and normal tissues. They also developed a database, piRNA-eQTL, containing eQTL results with differential expression and survival analyses. From the database, it was found that the expression level of piR-317 in pancreatic cancer tissues was lower compared to normal tissue, suggesting its tumor suppressive effect of piR-317 [140]. The expression level of piR-1945036 was significantly up-regulated in pancreatic cancer tissues. The patients with low piR-1945036 had longer survival than the patients with higher piR-1945036 expression [140], suggesting the oncogenic effect of piR-1945036.
5.2. Exosome piRNAs in Pancreatic Cancer
Serum exosomes from liquid biopsy are a good source of samples that could be tested for investigation of cell communications. Exosomes in blood circulation contain proteins and RNAs including piRNAs, which could reflect the healthy or pathological conditions of patients [141,142]. In a study on the analysis of exosomes, the exosomes from the serum of healthy people, intraductal papillary mucosal neoplasms, and pancreatic cancer were isolated and total RNAs from exosomes were extracted [141]. The total RNAs were further analyzed by next generation sequencing. Sequencing analysis showed that piRNAs including hsa-piR-52959, hsa-piR-53108, hsa-piR-30690, hsa-piR-54479, and hsa-piR-56621 were up-regulated in patients with pancreatic cancer compared to healthy people [141], suggesting the oncogenic effects of these piRNA. Whereas, several piRNAs such as hsa-piR-54888, hsa-piR-42185, hsa-piR-46410, hsa-piR-58897, and hsa-piR-43043 were down-regulated in patients with pancreatic cancer [141], suggesting that these piRNAs could have tumor suppressive functions. In addition, exosome hsa-piR-001311 and hsa-piR-016658 were found to have abundant level differences between pancreatic cancer and controls [143]. However, the roles of these exosome piRNAs need further investigations.
5.3. piR-017061
Next-generation sequencing (NGS) has also been used in the analysis of pancreatic cancer tissues. In a study using NGS, coding RNAs, lncRNAs, and small non-coding RNAs including piRNAs in tissue samples from six pancreatic cancers and five pancreatic controls were analyzed [144]. It was found that piR-017061, a piRNA located within the HBII-296A snoRNA, was significantly downregulated in pancreatic cancer tissues [144]. This result suggests that piR-017061 could be a tumor suppressive piRNA in pancreas. A recent report by another group confirmed the tumor suppressive effects of piR-017061 [44]. They found that the expression of piR-017061 was significantly downregulated in pancreatic cancer tissues and cells. Functional analysis showed that piR-017061 suppressed the growth of pancreatic cancer cell in vitro and in vivo [44]. A mechanistic study identified that EFNA5 mRNA was a target of piR-017061 and that piR-017061 directly bond to EFNA5 mRNA mediated by PIWIL1, leading to the degradation of EFNA5 mRNA. Loss of EFNA5, which had an oncogenic effect, inhibited the development and progression of pancreatic cancer [44]. Therefore, targeting PIWI/piRNA-mediated EFNA5 gene regulation could be a new strategy for the treatment of pancreatic cancer.
6. The Role of Non-Coding RNAs in the Diagnosis of Pancreatic Cancer
Several lncRNAs have their tissue-specific and cancer type-specific distribution, suggesting that they can be used as diagnostic biomarkers [145,146,147,148]. Among them, PCA3 (Prostate Cancer Antigen 3) is the most well-recognized lncRNA which can be used for the diagnosis of prostate cancer [148] and is approved by FDA. For diagnosis of pancreatic cancer, no such FDA approved lncRNA-based diagnosis has been reported so far. However, researches on the development of ncRNA testing as diagnosis or supplementary diagnostic tools for pancreatic cancer have received much attention and revealed some novel markers for pancreatic cancer [149,150,151,152].
6.1. SNHG15
SNHG15 is an lncRNA which plays critical roles in the development and progression of various cancers through regulation of Akt/mTOR signaling [153,154]. It has been reported as a prognostic marker in colorectal and liver cancers [155,156]. A study was focused on the investigation of SNHG15 as potential diagnostic markers of pancreatic cancer. It was found that the level of SNHG15 was significantly higher in both plasma and cancer tissues from patients with pancreatic cancer [157]. Receiver operator characteristic (ROC) analysis, which is usually used for determining diagnosis ability, showed that plasma SNHG15 could be a biomarker for distinguishing patients with pancreatic cancer from healthy people [157]. Moreover, a high level of SNHG15 was also significantly correlated with lymph node metastasis, higher tumor stage, and shorter overall survival [157], suggesting that SNHG15 is a promising biomarker for the diagnosis and prognosis of pancreatic cancer.
6.2. ABHD11-AS1
Circulating lncRNA ABHD11-AS1 has also been identified as a biomarker for the early detection of pancreatic cancer [151]. In a study on lncRNA, 11 pancreatic cancer related lncRNAs were investigated to reveal their roles as diagnostic markers [151]. The levels of these lncRNAs in plasma samples from 114 patients with pancreatic cancer, 97 patients with chronic pancreatitis, and 46 healthy controls were measured and ROC analysis was conducted. It was found that among 11 lncRNAs, ABHD11-AS1 had the best diagnostic performance for the diagnosis of pancreatic cancer [151]. Moreover, ROC analysis suggested that ABHD11-AS1 could be a better biomarker than CEA, CA19-9, and CA125 for early detection of pancreatic cancer. Furthermore, it was found that the combination of ABHD11-AS1 and CA19-9 was more efficient for the diagnosis of pancreatic cancer [151], suggesting the importance of the new diagnostic marker ABHD11-AS1.
6.3. HULC
HULC is a lncRNA that promotes cancer cell EMT, invasion, and metastasis [158,159]. The roles of HULC as a biomarker in the diagnosis of pancreatic cancer have been investigated. The study found that extracellular vesicle encapsulated HULC in blood could be a potential biomarker for the diagnosis of pancreatic cancer [150]. HULC level was significantly higher in the patients with pancreatic cancer compared to healthy individuals [150]. Further analysis showed that HULC in blood extracellular vesicle had good predictive performance for diagnosis of pancreatic cancer. In addition, another separate study using blood samples from 60 patients with pancreatic cancer and 60 patients with benign pancreatic diseases showed that high expression of HULC in serum could be used to distinguish patients with pancreatic cancer from patients with benign pancreatic diseases [149]. Interestingly, HULC could serve as a serum biomarker for the diagnosis and prognosis of gastric cancer [160]. These findings suggest that lncRNA HULC could be a novel biomarker for the diagnosis and prognosis of pancreatic and gastric cancers.
6.4. UFC1
The lncRNA UFC1 has been known as an oncogenic lncRNA regulating EZH2 signaling in cancer [161]. The role of serum UFC1 in the diagnosis of pancreatic cancer was investigated. Compared to healthy subjects, the level of serum UFC1 in pancreatic cancer patients was significantly higher [162]. ROC analysis showed that serum UFC1 levels could effectively distinguish patients with pancreatic cancer from healthy subjects. Moreover, the serum UFC1 level was correlated with lymph node metastasis, distant metastasis, and clinical stage [162]. The patients with high level of UFC1 had shorter progression-free and overall survival than those with a low level of UFC1 [162]. These results clearly suggest that serum lncRNA UFC1 could be a novel biomarker for the diagnosis and prognosis of pancreatic cancer.
6.5. circ_001569
circ_001569 is an oncogenic circRNA regulating PI3K/Akt and Wnt signaling [163]. High expression of circ_001569 have been found in colorectal and liver cancers [164,165,166]. To find the diagnostic and prognostic values of circ_001569 in pancreatic cancer, the expression levels of circ_001569 in plasma samples from 71 patients with pancreatic cancer and 71 healthy subjects were measured [167]. It was found that the level of plasma circ_001569 was significantly increased in patients with pancreatic cancer [167]. ROC analysis showed that plasma circ_001569 could efficiently distinguish patients with pancreatic cancer from healthy subjects. In addition, the high level of circ_001569 was positively associated with lymph node metastasis, tumor stage, and invasion [167]. The pancreatic cancer patients with a high level of circ_001569 had a poor prognosis. These observations suggest that circ_001569 in plasma could be a promising biomarker for the diagnosis and prognosis of pancreatic cancers.
6.6. circ-LDLRAD3
circ-LDLRAD3 is another circRNA which could be used for the diagnosis of pancreatic cancer. In a study on circ-LDLRAD3, the expression levels of circ-LDLRAD3 in pancreatic cancer and adjacent non-tumorous tissues, plasma samples from patients with pancreatic cancer and plasma samples from healthy subjects were measured [168]. It was found that the level of circ-LDLRAD3 was significantly increased in pancreatic cancer tissues and plasma compared to normal tissues or plasma [168]. ROC analysis showed that circ-LDLRAD3 alone could efficiently diagnose pancreatic cancer [168]. A combination of circ-LDLRAD3 with CA19-9 had a much higher diagnostic sensitivity and specificity for pancreatic cancer. In addition, the high level of circ-LDLRAD3 was also significantly correlated with invasion and metastasis pancreatic cancer [168]. These results demonstrate that the level of circ-LDLRAD3 could be used as diagnostic and prognostic markers for pancreatic cancer.
7. The Role of Non-Coding RNAs in the Therapy of Pancreatic Cancer
In the development of cancer therapy targeting ncRNAs, strategies for the delivery of anti-ncRNA molecules include nanoparticle-packed siRNA delivery, LNA GapmeRs delivery and exosome-based delivery [169,170,171]. Nanoparticle-packed siRNA has been utilized to inhibit lncRNAs in clinical trial [169]. LNA GapmeRs delivery system has been successfully used to inhibit lncRNA MALAT1 [170]. Studies have also confirmed that exosome can deliver exogenous RNAs including siRNAs and other ncRNAs [171,172,173,174]. Therefore, exosome-based delivery of siRNAs and other ncRNAs could be another novel strategy for the treatment of cancers.
ncRNAs also play important roles in drug resistance and the prognosis of pancreatic cancer [175,176,177,178]. In the treatment of pancreatic cancer, ncRNAs could influence the efficacy of cancer therapeutics including chemotherapy, radiotherapy, immunotherapy, and other targeted therapy by regulation of cancer cell sensitivity to therapies. In addition, some ncRNAs could be used for predicting the prognosis of pancreatic cancer patients after treatment.
7.1. ncRNAs Regulated Drug Sensitivity in Pancreatic Cancer
Gemcitabine resistance is a major chemoresistance in the treatment of pancreatic cancer. A study was conducted to find the difference of lncRNA profiles between gemcitabine sensitive and resistant pancreatic cancer cells using genomic analysis [178]. By comparing lncRNA profiles between gemcitabine sensitive and resistant cells, it was found that 4983 of 13,310 detected lncRNAs demonstrated more than 2-fold changes, suggesting that lncRNA regulation is significantly involved in drug resistance in pancreatic cancer [178]. PVT1 is a lncRNA and significantly overexpressed in pancreatic cancer [79]. PVT1 has been identified as a regulator of gemcitabine sensitivity. Study showed that functional inactivation of PVT1 resulted in enhanced gemcitabine sensitivity whereas overexpression of PVT1 enhanced resistance to gemcitabine in human pancreatic cancer [79]. Another lncRNA HOTTIP is also significantly upregulated in human pancreatic cancer [83]. High expression of HOTTIP in pancreatic cancer induced gemcitabine resistance while inhibition of HOTTIP potentiated anticancer activity of gemcitabine in vitro and in vivo [83]. In addition, linc-ROR is also a lncRNA which is up-regulated in pancreatic cancer [179]. Linc-ROR was also involved in the regulation of gemcitabine sensitivity in pancreatic cancer [180]. Down-regulation of linc-ROR significantly sensitized pancreatic cancer cells to gemcitabine whereas overexpression of linc-ROR significantly reduced the sensitivity of pancreatic cancer cells to gemcitabine [180]. lncRNA MALAT-1 is also associated with chemoresistance of pancreatic cancer [60]. A study showed that an elevated level of MALAT-1 decreased sensitivity to gemcitabine in pancreatic cancer cells [60], suggesting that down-regulation of MALAT-1 could increase the sensitivity of pancreatic cancer cells to gemcitabine.
In addition to lncRNAs, circRNAs also regulate the sensitivity of pancreatic cancer to gemcitabine [176,181]. A study showed that silencing two circRNAs (chr14:101402109-101464448+ and chr4:52729603-52780244+) in gemcitabine resistant pancreatic cancer cells restored sensitivity to gemcitabine, suggesting the role of these circRNAs in the regulation of gemcitabine resistance [176]. By profiling circRNAs in gemcitabine resistant and sensitive pancreatic cancer cells, it was found that circ_101672, circ_004077, and circ_003251 were the top three upregulated circRNAs while circ_101543, circ_102747, and circ_000926 were the top three down-regulated circRNAs in gemcitabine resistant pancreatic cancers [182], suggesting that regulation of these circRNAs could restore the sensitivity of pancreatic cancer cells to gemcitabine. CircHIPK3 is another circRNA up-regulated in gemcitabine resistant pancreatic cancer cells and tumor tissues [181]. Knockdown of circHIPK3 in PANC-1-GEM and SW-1990-GEM gemcitabine resistant cells could significantly inhibit cell proliferation, invasion, migration, and sensitize pancreatic cancer cells to gemcitabine [181].
Since ncRNAs play important roles in the chemoresistance of pancreatic cancer, therapies targeting ncRNAs could reduce chemoresistance in pancreatic cancer. Therefore, therapeutic strategy targeting ncRNAs could be designed as to block chemoresistance related ncRNAs by siRNAs. Because the short nucleotides such as siRNAs are easy to be degraded in vivo, novel efficient delivery systems are needed for successful delivery of short nucleotides into target cells [183]. Recently, nanoparticle delivery systems have been used for delivery of ncRNAs to pancreatic cancer cells and tumor microenvironments for the treatment of pancreatic cancer [184,185].
7.2. ncRNAs for the Prognosis of Pancreatic Cancer
Recent studies have demonstrated that certain lncRNAs could be used for the prognosis of pancreatic cancer [53,186,187]. A study on 80 patients with pancreatic cancer showed that high expression of lncRNA AFAP1-AS1 and UCA1, and low expression of ENSG00000218510 could predict poor overall survival and tumor progression in pancreatic cancer [186]. Similar results from another group also demonstrated that AFAP1-AS1 overexpression was associated with lymph node metastasis, perineural invasion, and poor survival [187]. The study also confirmed that knockdown of AFAP1-AS1 led to the inhibition of pancreatic cancer cell proliferation, migration, and invasion, demonstrating the value of AFAP1-AS1 as a prognosis marker [187]. HOTAIR is another lncRNA which is highly expressed in pancreatic cancer [55]. HOTAIR could be another prognostic markers for pancreatic cancer. A study showed that patients with low HOTAIR expression had significantly increased overall survival compared to patients with high HOTAIR expression [53]. As mention early, MALAT1 is another oncogenic lncRNA highly expressed in pancreatic cancer [63]. Clinical studies demonstrated that MALAT1 was correlated with clinical progression and unfavorable prognosis in pancreatic cancer and that patients with higher expression of MALAT1 had a poorer disease free survival compared to patients with low expression of MALAT1 [63,100]. In a study on lncRNA profiling analysis with clinical data, it was found that the expression level of lncRNA BC008363 was significantly lower in pancreatic cancer tissues compared to normal pancreatic tissues [175]. More importantly, patients with high expression level of BC008363 had significantly better survival than those with low expression level of BC00836 [175], suggesting its value as a prognostic marker. In addition, oncogenic lncRNA PVT1 was found to be an independent prognostic factor for poor overall survival in patients with pancreatic cancer [73]. Pancreatic cancer patients with high expression of PVT1 had shorter overall survival times compared to those with low expression of PVT1 [73].
In addition to lncRNAs, circRNAs could also function as prognostic markers. circ-ADAM9 is an oncogenic circRNA [188]. In a study on 58 patients with pancreatic cancer, high expression of circ-ADAM9 was found in pancreatic cancer tissues with advanced clinical stage and lymph node metastasis [188]. More importantly, it was found that pancreatic cancer patients with high expression of circ-ADAM9 had poorer survival rate than those with low expression of circ-ADAM9 [188]. Another circRNA circ-LDLRAD3 was shown to be overexpressed in pancreatic cancer tissues and cells [168,189]. Further analysis showed that high expression of circ-LDLRAD3 was a marker indicating a poor prognosis in patients with pancreatic cancer [189]. CircRNA circ_0030235 was also significantly increased in pancreatic cancer tissues and cells compared to normal pancreatic tissues and cells [177]. Clinical study found that overexpression of circ_0030235 in tumor samples was associated with higher tumor stage and lymph node invasion [177]. Kaplan-Meier analysis showed that high expression of circ_0030235 significantly predicted poor prognosis in pancreatic cancer [177]. circRNA hsa_circ_0001649 could also serve as an independent prognostic biomarker of pancreatic cancer [136]. Fisher’s exact test found that patients with low expression of hsa_circ_0001649 had an advanced tumor stage and lower differentiation grade [136]. Kaplan-Meier analysis showed that the patients with high expression of hsa_circ_0001649 had better 5-year overall survival [136], suggesting the prognostic value of hsa_circ_0001649. In addition, testing circRNA in plasma samples to predict the prognosis of patients with pancreatic cancer is a more convenient way in clinic. Studies found that high level of circ-IARS or circ-PDE8A in plasma exosomes secreted by pancreatic cancer indicated a poor overall survival of patients with pancreatic cancer [190,191]. These findings on ncRNAs could help to design targeted therapeutic strategy more optimally to better treat pancreatic cancers.
8. Conclusions
Pancreatic cancer is the seventh leading cause of cancer related deaths worldwide. In the United States [192], pancreatic cancer remains the fourth leading cause of cancer related death with an estimated 60,430 new cases and 48,220 deaths in 2021 in the United States [193]. It is expected that pancreatic cancer will become the second leading cause of cancer-related death by 2030 [194]. Both incidence and mortality rates for pancreatic cancer have increased since 2000 in US [193]. There are no early screening biomarkers for pancreatic cancer. Currently, CA19-9 is a biomarker for the diagnosis and prognosis of pancreatic cancer. However, CA19-9 has limitation in the early diagnosis of this disease because of its false positivity and nonspecific characteristics [195]. Therefore, there is an urgent need to find novel and effective biomarkers for the diagnosis of pancreatic cancer. Moreover, so far there are no specific or effective drugs to overcome chemoresistance for the treatment of pancreatic cancer. Thus, finding novel therapeutic and prognostic biomarkers is critically important for the improvement of cancer therapy for pancreatic cancer.
The recent studies described above indicate that ncRNAs including lncRNAs, circRNAs, and piRNAs play critical roles in the regulation of the development and progression of pancreatic cancer. Therefore, these ncRNAs could be novel biomarkers for the diagnosis and treatment of pancreatic cancer. For the diagnosis of pancreatic cancer, ncRNAs in extracellular vesicles such as SNHG15, ABHD11-AS1, HULC, UFC1, circ_001569, and circ-LDLRAD3 described above could serve as diagnostic biomarkers combined with CA19-9 in liquid biopsy for better diagnosis of pancreatic cancer. This non-invasive biopsy is more convenient in clinic. For the treatment of pancreatic cancer, currently available studies have shown that several ncRNAs have effects on the regulation of drug sensitivity in pancreatic cancer. These ncRNAs include PVT1, HOTTIP, linc-ROR, MALAT-1, circHIPK3, circ_101672, circ_004077, circ_003251, circ_101543, circ_102747, and circ_000926. Therefore, regulations of these ncRNAs could increase pancreatic cancer cell sensitivity to conventional therapies to achieve better treatment outcome. In addition, several ncRNAs such as AFAP1-AS1, UCA1, HOTAIR, PVT1, MALAT-1, circ-ADAM9, BC008363, circ-LDLRAD3, circ-IARS, circ-PDE8A, circ_0030235, and circ_0001649 have been associated with prognosis of pancreatic cancer. Detection of these ncRNAs could predict the treatment outcome in pancreatic cancer. Targeting ncRNAs by siRNA is main strategy for targeted therapy [172,173]. However, easy degradation and difficult delivery of siRNA are challenges for ncRNA targeted therapy. Nevertheless, using nanoparticle-packed siRNA, LNA GapmeRs, and exosome-based delivery to protect siRNA and target oncogenic ncRNAs is on its way and will be tested in pre-clinical and clinical studies. It is important to note that more molecular studies and in vivo pre-clinical studies are needed before these strategies targeting ncRNAs could be used in clinical practice.
Author Contributions
Conceptualization: A.S.A. and R.M.M.; Writing—original draft preparation: Y.L.; Review and editing: Y.L., M.N.A.H., P.A.P., A.S.A. and R.M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partly funded by the National Institutes of Health (R37CA215427 and R01CA240607), SKY Foundation and Perri Family Foundation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Toden, S.; Zumwalt, T.J.; Goel, A. Non-coding RNAs and potential therapeutic targeting in cancer. Biochim. Biophys Acta Rev. Cancer 2021, 1875, 188491. [Google Scholar] [CrossRef]
- Eddy, S.R. Non-coding RNA genes and the modern RNA world. Nat. Rev. Genet. 2001, 2, 919–929. [Google Scholar] [CrossRef]
- Slack, F.J.; Chinnaiyan, A.M. The role of non-coding RNAs in oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Cheng, G. Circulating miRNAs: Roles in cancer diagnosis, prognosis and therapy. Adv. Drug Deliv. Rev. 2015, 81, 75–93. [Google Scholar] [CrossRef]
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in human diseases and potential use as biomarkers. Cell Death Dis. 2021, 12, 468. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Huang, Q.Y.; Liu, G.F.; Qian, X.L.; Tang, L.B.; Huang, Q.Y.; Xiong, L.X. Long non-coding RNA: Dual Effects on breast cancer metastasis and clinical applications. Cancers 2019, 11, 1802. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Meng, X.M.; Huang, C.; Wu, B.M.; Zhang, L.; Lv, X.W.; Li, J. Long noncoding RNAs: Novel insights into hepatocelluar carcinoma. Cancer Lett. 2014, 344, 20–27. [Google Scholar] [CrossRef]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Brannan, C.I.; Dees, E.C.; Ingram, R.S.; Tilghman, S.M. The product of the H19 gene may function as an RNA. Mol. Cell Biol. 1990, 10, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.J.; Ballabio, A.; Rupert, J.L.; Lafreniere, R.G.; Grompe, M.; Tonlorenzi, R.; Willard, H.F. A gene from the region of the human X inactivation centre is expressed exclusively from the inactive X chromosome. Nature 1991, 349, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Furuno, M.; Kasukawa, T.; Adachi, J.; Bono, H.; Kondo, S.; Nikaido, I.; Osato, N.; Saito, R.; Suzuki, H.; et al. Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs. Nature 2002, 420, 563–573. [Google Scholar]
- Kung, J.T.; Colognori, D.; Lee, J.T. Long noncoding RNAs: Past, present, and future. Genetics 2013, 193, 651–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, R.W.; Wang, Y.; Chen, L.L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.J.; Dang, H.X.; Lim, D.A.; Feng, F.Y.; Maher, C.A. Long noncoding RNAs in cancer metastasis. Nat. Rev. Cancer 2021, 21, 446–460. [Google Scholar] [CrossRef]
- Chen, L.L. The biogenesis and emerging roles of circular RNAs. Nat. Rev. Mol. Cell Biol. 2016, 17, 205–211. [Google Scholar] [CrossRef]
- Hentze, M.W.; Preiss, T. Circular RNAs: Splicing’s enigma variations. EMBO J. 2013, 32, 923–925. [Google Scholar] [CrossRef] [PubMed]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [Green Version]
- Vicens, Q.; Westhof, E. Biogenesis of Circular RNAs. Cell 2014, 159, 13–14. [Google Scholar] [CrossRef] [Green Version]
- Cocquerelle, C.; Mascrez, B.; Hétuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. FASEB J. 1993, 7, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.L.; Bao, Y.; Yee, M.C.; Barrett, S.P.; Hogan, G.J.; Olsen, M.N.; Dinneny, J.R.; Brown, P.O.; Salzman, J. Circular RNA is expressed across the eukaryotic tree of life. PLoS ONE 2014, 9, e90859. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Jiang, J.; Shi, H.; Qian, H.; Zhang, X.; Xu, W. CircRNA: A rising star in gastric cancer. Cell Mol. Life Sci. 2020, 77, 1661–1680. [Google Scholar] [CrossRef]
- Su, M.; Xiao, Y.; Ma, J.; Tang, Y.; Tian, B.; Zhang, Y.; Li, X.; Wu, Z.; Yang, D.; Zhou, Y.; et al. Circular RNAs in cancer: Emerging functions in hallmarks, stemness, resistance and roles as potential biomarkers. Mol. Cancer 2019, 18, 90. [Google Scholar] [CrossRef]
- Ernst, C.; Odom, D.T.; Kutter, C. The emergence of piRNAs against transposon invasion to preserve mammalian genome integrity. Nat. Commun. 2017, 8, 1411. [Google Scholar] [CrossRef] [Green Version]
- Lin, H. piRNAs in the germ line. Science 2007, 316, 397. [Google Scholar] [CrossRef]
- Teixeira, F.K.; Okuniewska, M.; Malone, C.D.; Coux, R.X.; Rio, D.C.; Lehmann, R. piRNA-mediated regulation of transposon alternative splicing in the soma and germ line. Nature 2017, 552, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Grivna, S.T.; Beyret, E.; Wang, Z.; Lin, H. A novel class of small RNAs in mouse spermatogenic cells. Genes Dev. 2006, 20, 1709–1714. [Google Scholar] [CrossRef] [Green Version]
- Ross, R.J.; Weiner, M.M.; Lin, H. PIWI proteins and PIWI-interacting RNAs in the soma. Nature 2014, 505, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Aravin, A.; Gaidatzis, D.; Pfeffer, S.; Lagos-Quintana, M.; Landgraf, P.; Iovino, N.; Morris, P.; Brownstein, M.J.; Kuramochi-Miyagawa, S.; Nakano, T.; et al. A novel class of small RNAs bind to MILI protein in mouse testes. Nature 2006, 442, 203–207. [Google Scholar] [CrossRef]
- Guo, B.; Li, D.; Du, L.; Zhu, X. piRNAs: Biogenesis and their potential roles in cancer. Cancer Metastasis Rev. 2020, 39, 567–575. [Google Scholar] [CrossRef]
- Wu, W.; Lu, B.F.; Jiang, R.Q.; Chen, S. The function and regulation mechanism of piRNAs in human cancers. Histol. Histopathol. 2021. [Google Scholar] [CrossRef]
- Ipsaro, J.J.; Haase, A.D.; Knott, S.R.; Joshua-Tor, L.; Hannon, G.J. The structural biochemistry of Zucchini implicates it as a nuclease in piRNA biogenesis. Nature 2012, 491, 279–283. [Google Scholar] [CrossRef] [Green Version]
- Saxe, J.P.; Chen, M.; Zhao, H.; Lin, H. Tdrkh is essential for spermatogenesis and participates in primary piRNA biogenesis in the germline. EMBO J. 2013, 32, 1869–1885. [Google Scholar] [CrossRef]
- Lim, S.L.; Qu, Z.P.; Kortschak, R.D.; Lawrence, D.M.; Geoghegan, J.; Hempfling, A.L.; Bergmann, M.; Goodnow, C.C.; Ormandy, C.J.; Wong, L.; et al. HENMT1 and piRNA Stability are required for adult male germ cell transposon repression and to define the spermatogenic program in the mouse. PLoS Genet. 2015, 11, e1005620. [Google Scholar]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O’Carroll, D.; Zamore, P.D. PIWI-interacting RNAs: Small RNAs with big functions. Nat. Rev. Genet. 2019, 20, 89–108. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Tu, S.; Stubna, M.; Wu, W.S.; Huang, W.C.; Weng, Z.; Lee, H.C. The piRNA targeting rules and the resistance to piRNA silencing in endogenous genes. Science 2018, 359, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Rios, P.; Simonelig, M. piRNAs and PIWI proteins: Regulators of gene expression in development and stem cells. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Kang, J.Y.; Gou, L.T.; Wang, J.; Xue, Y.; Skogerboe, G.; Dai, P.; Huang, D.W.; Chen, R.; Fu, X.D.; et al. MIWI and piRNA-mediated cleavage of messenger RNAs in mouse testes. Cell Res. 2015, 25, 193–207. [Google Scholar] [CrossRef]
- Goh, W.S.; Falciatori, I.; Tam, O.H.; Burgess, R.; Meikar, O.; Kotaja, N.; Hammell, M.; Hannon, G.J. piRNA-directed cleavage of meiotic transcripts regulates spermatogenesis. Genes Dev. 2015, 29, 1032–1044. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Xing, S.; Shen, B.Y.; Chen, H.T.; Sun, B.; Wang, Z.T.; Wang, J.W.; Lu, X.X. PIWIL1 interacting RNA piR-017061 inhibits pancreatic cancer growth via regulating EFNA5. Hum. Cell 2021, 34, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Fan, G.; Song, S.; Jiang, Y.; Qian, C.; Zhang, W.; Su, Q.; Xue, X.; Zhuang, W.; Li, B. piRNA-30473 contributes to tumorigenesis and poor prognosis by regulating m6A RNA methylation in DLBCL. Blood 2021, 137, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zheng, J.; Lin, D. PIWI-interacting RNAs in human cancer. Semin Cancer Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, N.; Zhang, F.; Jin, S.; Dong, Y.; Dong, X.; Chen, Y.; Kong, X.; Tong, Y.; Mi, Q.; et al. PIWI-interacting RNAs are aberrantly expressed and may serve as novel biomarkers for diagnosis of lung adenocarcinoma. Thorac. Cancer 2021. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, J.; Sun, L.; Li, M.; He, X.; Jiang, J.; Zhou, Q. Piwi-interacting RNA-651 promotes cell proliferation and migration and inhibits apoptosis in breast cancer by facilitating DNMT1-mediated PTEN promoter methylation. Cell Cycle 2021. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dong, Y.; He, X.; Gong, A.; Gao, J.; Hao, X.; Wang, S.; Fan, Y.; Wang, Z.; Li, M.; et al. piR-hsa-211106 inhibits the progression of lung adenocarcinoma through pyruvate carboxylase and enhances chemotherapy sensitivity. Front. Oncol. 2021, 11, 651915. [Google Scholar] [CrossRef]
- Ding, X.; Li, Y.; Lü, J.; Zhao, Q.; Guo, Y.; Lu, Z.; Ma, W.; Liu, P.; Pestell, R.G.; Liang, C.; et al. piRNA-823 is involved in cancer stem cell regulation through altering dna methylation in association with luminal breast cancer. Front. Cell Dev. Biol. 2021, 9, 641052. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Meng, X.; Li, D.; Han, X. piR-001773 and piR-017184 promote prostate cancer progression by interacting with PCDH9. Cell Signal. 2020, 76, 109780. [Google Scholar] [CrossRef]
- Jiang, D.; Xu, L.; Ni, J.; Zhang, J.; Cai, M.; Shen, L. Functional polymorphisms in LncRNA HOTAIR contribute to susceptibility of pancreatic cancer. Cancer Cell Int. 2019, 19, 47. [Google Scholar] [CrossRef]
- Kim, K.; Jutooru, I.; Chadalapaka, G.; Johnson, G.; Frank, J.; Burghardt, R.; Kim, S.; Safe, S. HOTAIR is a negative prognostic factor and exhibits pro-oncogenic activity in pancreatic cancer. Oncogene 2013, 32, 1616–1625. [Google Scholar] [CrossRef] [Green Version]
- Li, C.H.; Xiao, Z.; Tong, J.H.; To, K.F.; Fang, X.; Cheng, A.S.; Chen, Y. EZH2 coupled with HOTAIR to silence MicroRNA-34a by the induction of heterochromatin formation in human pancreatic ductal adenocarcinoma. Int. J. Cancer 2017, 140, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; He, Y.; Ye, X.; Song, J.; Wang, Q.; Li, Y.; Xie, X. High expression of long noncoding RNA HOTAIRM1 is associated with the proliferation and migration in pancreatic ductal adenocarcinoma. Pathol. Oncol. Res. 2019, 25, 1567–1577. [Google Scholar] [CrossRef]
- Wang, L.; Dong, P.; Wang, W.; Huang, M.; Tian, B. Gemcitabine treatment causes resistance and malignancy of pancreatic cancer stem-like cells via induction of lncRNA HOTAIR. Exp. Med. 2017, 14, 4773–4780. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.Z.; Xu, F.; Zhou, T.; Zhao, X.; McDonald, J.M.; Chen, Y. The long non-coding RNA HOTAIR enhances pancreatic cancer resistance to TNF-related apoptosis-inducing ligand. J. Biol. Chem. 2017, 292, 10390–10397. [Google Scholar] [CrossRef] [Green Version]
- Han, T.; Jiao, F.; Hu, H.; Yuan, C.; Wang, L.; Jin, Z.L.; Song, W.F.; Wang, L.W. EZH2 promotes cell migration and invasion but not alters cell proliferation by suppressing E-cadherin, partly through association with MALAT-1 in pancreatic cancer. Oncotarget 2016, 7, 11194–11207. [Google Scholar] [CrossRef] [Green Version]
- Jiao, F.; Hu, H.; Yuan, C.; Wang, L.; Jiang, W.; Jin, Z.; Guo, Z.; Wang, L. Elevated expression level of long noncoding RNA MALAT-1 facilitates cell growth, migration and invasion in pancreatic cancer. Oncol. Rep. 2014, 32, 2485–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, F.; Hu, H.; Han, T.; Yuan, C.; Wang, L.; Jin, Z.; Guo, Z.; Wang, L. Long noncoding RNA MALAT-1 enhances stem cell-like phenotypes in pancreatic cancer cells. Int. J. Mol. Sci. 2015, 16, 6677–6693. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Chen, H.; Gao, Y.; Wang, Y.W.; Zhang, G.Q.; Pan, S.H.; Ji, L.; Kong, R.; Wang, G.; Jia, Y.H.; et al. Long noncoding RNA MALAT1 promotes aggressive pancreatic cancer proliferation and metastasis via the stimulation of autophagy. Mol. Cancer 2016, 15, 2232–2243. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Yang, H.; Zhang, J.; Peng, X.; Lu, Z.; Tong, W.; Chen, J. The lncRNA MALAT1 acts as a competing endogenous RNA to regulate KRAS expression by sponging miR-217 in pancreatic ductal adenocarcinoma. Sci. Rep. 2017, 7, 5186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, E.J.; Yang, R.; Fu, X.B.; Liu, Y.F. Overexpression of long non-coding RNA MALAT1 is correlated with clinical progression and unfavorable prognosis in pancreatic cancer. Tumour Biol. 2015, 36, 2403–2407. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, X.; Shi, M.; Wen, C.; Shen, B. MiR-216a decreases MALAT1 expression, induces G2/M arrest and apoptosis in pancreatic cancer cells. Biochem. Biophys. Res. Commun. 2017, 483, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shan, T.; Ding, W.; Hua, Z.; Shen, Y.; Lu, Z.; Chen, B.; Dai, T. Study on mechanism about long noncoding RNA MALAT1 affecting pancreatic cancer by regulating hippo-YAP signaling. J. Cell Physiol. 2018, 233, 5805–5814. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, M.; Yuan, C.; Han, T.; Cui, J.; Jiao, F.; Wang, L. A novel feedback loop between high MALAT-1 and low miR-200c-3p promotes cell migration and invasion in pancreatic ductal adenocarcinoma and is predictive of poor prognosis. BMC Cancer 2018, 18, 1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Nong, K.; Zhu, H.; Wang, W.; Huang, X.; Yuan, Z.; Ai, K. H19 promotes pancreatic cancer metastasis by derepressing let-7’s suppression on its target HMGA2-mediated EMT. Tumour Biol. 2014, 35, 9163–9169. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Tian, X.; Wang, F.; Zhang, Z.; Du, C.; Xie, X.; Kornmann, M.; Yang, Y. The long noncoding RNA H19 promotes cell proliferation via E2F-1 in pancreatic ductal adenocarcinoma. Cancer Biol. 2016, 17, 1051–1061. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Tian, X.; Guo, H.; Zhang, Z.; Du, C.; Wang, F.; Xie, X.; Gao, H.; Zhuang, Y.; Kornmann, M.; et al. Long noncoding RNA H19 derived miR-675 regulates cell proliferation by down-regulating E2F-1 in human pancreatic ductal adenocarcinoma. J. Cancer 2018, 9, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhu, Q.; Yang, W.; Shan, Y.; Yu, Z.; Zhang, Q.; Wu, H. LncRNA H19/miR-194/PFTK1 axis modulates the cell proliferation and migration of pancreatic cancer. J. Cell Biochem. 2019, 120, 3874–3886. [Google Scholar] [CrossRef]
- Wang, F.; Rong, L.; Zhang, Z.; Li, M.; Ma, L.; Ma, Y.; Xie, X.; Tian, X.; Yang, Y. LncRNA H19-derived miR-675-3p promotes epithelial-mesenchymal transition and stemness in human pancreatic cancer cells by targeting the STAT3 pathway. J. Cancer 2020, 11, 4771–4782. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, H.; Matsuda, Y.; Yamamoto, M.; Michishita, M.; Takahashi, K.; Sasaki, N.; Ishikawa, N.; Aida, J.; Takubo, K.; Arai, T.; et al. Reduced expression of the H19 long non-coding RNA inhibits pancreatic cancer metastasis. Lab. Investig. 2018, 98, 814–824. [Google Scholar] [CrossRef]
- Huang, C.; Yu, W.; Wang, Q.; Cui, H.; Wang, Y.; Zhang, L.; Han, F.; Huang, T. Increased expression of the lncRNA PVT1 is associated with poor prognosis in pancreatic cancer patients. Minerva Med. 2015, 106, 143–149. [Google Scholar]
- Huang, F.; Chen, W.; Peng, J.; Li, Y.; Zhuang, Y.; Zhu, Z.; Shao, C.; Yang, W.; Yao, H.; Zhang, S. LncRNA PVT1 triggers cyto-protective autophagy and promotes pancreatic ductal adenocarcinoma development via the miR-20a-5p/ULK1 Axis. Mol. Cancer 2018, 17, 98. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Wang, P.; Dong, W.; Liu, H.; Sun, J.; Zhao, L. LncRNA PVT1 promotes exosome secretion through YKT6, RAB7, and VAMP3 in pancreatic cancer. Aging 2020, 12, 10427–10440. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, P.; Yin, T.; Zhang, F.; Wang, W. Upregulation of LncRNA PVT1 facilitates pancreatic ductal adenocarcinoma cell progression and glycolysis by regulating MiR-519d-3p and HIF-1A. J. Cancer 2020, 11, 2572–2579. [Google Scholar] [CrossRef]
- Wang, C.J.; Shi, S.B.; Tian, J.; Xu, J.; Niu, Z.X. lncRNA MALAT1, HOTTIP and PVT1 as predictors for predicting the efficacy of GEM based chemotherapy in first-line treatment of pancreatic cancer patients. Oncotarget 2017, 8, 95108–95115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.Q.; Jiang, Y.; Zhu, F.; Sun, D.L.; He, X.Z. Long noncoding RNA PVT1 promotes EMT and cell proliferation and migration through downregulating p21 in pancreatic cancer cells. Technol. Cancer Res. Treat. 2017, 16, 819–827. [Google Scholar] [CrossRef] [Green Version]
- You, L.; Chang, D.; Du, H.Z.; Zhao, Y.P. Genome-wide screen identifies PVT1 as a regulator of gemcitabine sensitivity in human pancreatic cancer cells. Biochem. Biophys. Res. Commun. 2011, 407, 1–6. [Google Scholar] [CrossRef]
- Zhao, L.; Kong, H.; Sun, H.; Chen, Z.; Chen, B.; Zhou, M. LncRNA-PVT1 promotes pancreatic cancer cells proliferation and migration through acting as a molecular sponge to regulate miR-448. J. Cell Physiol. 2018, 233, 4044–4055. [Google Scholar] [CrossRef]
- Zhou, C.; Yi, C.; Yi, Y.; Qin, W.; Yan, Y.; Dong, X.; Zhang, X.; Huang, Y.; Zhang, R.; Wei, J.; et al. LncRNA PVT1 promotes gemcitabine resistance of pancreatic cancer via activating Wnt/β-catenin and autophagy pathway through modulating the miR-619-5p/Pygo2 and miR-619-5p/ATG14 axes. Mol. Cancer 2020, 19, 118. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Chen, C.; Zhou, Q.; Wang, Y.; Zhao, Y.; Zhao, X.; Li, W.; Zheng, S.; Ye, H.; Wang, L.; et al. LncRNA HOTTIP modulates cancer stem cell properties in human pancreatic cancer by regulating HOXA9. Cancer Lett. 2017, 410, 68–81. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, X.; Zhou, Y.; Liu, Y.; Zhou, Q.; Ye, H.; Wang, Y.; Zeng, J.; Song, Y.; Gao, W.; et al. The long non-coding RNA HOTTIP promotes progression and gemcitabine resistance by regulating HOXA13 in pancreatic cancer. J. Transl. Med. 2015, 13, 84. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.H.; Li, C.H.; He, Q.; Chan, S.L.; Tong, J.H.; To, K.F.; Lin, L.Z.; Chen, Y. Ectopic HOTTIP expression induces noncanonical transactivation pathways to promote growth and invasiveness in pancreatic ductal adenocarcinoma. Cancer Lett. 2020, 477, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Zhang, Q.; Dong, Z.; Hu, J.; Ma, Z. LncRNA HOTTIP Participates in cisplatin resistance of tumor cells by regulating miR-137 expression in pancreatic cancer. Onco Targets Ther. 2020, 13, 2689–2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.Q.; Wang, J.F.; Chen, D.H.; Ma, X.S.; Wu, Y.; Tang, Z.; Dang, X.W. Long non-coding RNA GAS5 suppresses pancreatic cancer metastasis through modulating miR-32-5p/PTEN axis. Cell Biosci. 2017, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.Q.; Wang, J.F.; Chen, D.H.; Ma, X.S.; Yang, W.; Zhe, T.; Dang, X.W. Long non-coding RNA GAS5 antagonizes the chemoresistance of pancreatic cancer cells through down-regulation of miR-181c-5p. Biomed. Pharm. 2018, 97, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wu, S.; Ma, J.; Yan, S.; Xiao, Z.; Wan, L.; Zhang, F.; Shang, M.; Mao, A. lncRNA GAS5 reverses EMT and tumor stem cell-mediated gemcitabine resistance and metastasis by targeting miR-221/SOCS3 in pancreatic cancer. Mol. Nucleic Acids 2018, 13, 472–482. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Fang, Y.; Wang, Z.; Xie, J.; Zhan, Q.; Deng, X.; Chen, H.; Jin, J.; Peng, C.; Li, H.; et al. Downregulation of gas5 increases pancreatic cancer cell proliferation by regulating CDK6. Cell Tissue Res. 2013, 354, 891–896. [Google Scholar] [CrossRef]
- Gu, L.; Zhang, J.; Shi, M.; Zhan, Q.; Shen, B.; Peng, C. lncRNA MEG3 had anti-cancer effects to suppress pancreatic cancer activity. Biomed. Pharm. 2017, 89, 1269–1276. [Google Scholar] [CrossRef]
- Iyer, S.; Modali, S.D.; Agarwal, S.K. Long noncoding RNA MEG3 is an epigenetic determinant of oncogenic signaling in functional pancreatic neuroendocrine tumor cells. Mol. Cell Biol. 2017, 37, e00278-17. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Wang, F.; Du, C.; Zhang, Z.; Guo, H.; Xie, X.; Gao, H.; Zhuang, Y.; Kornmann, M.; Gao, H.; et al. Long non-coding RNA MEG3 functions as a tumour suppressor and has prognostic predictive value in human pancreatic cancer. Oncol. Rep. 2018, 39, 1132–1140. [Google Scholar] [CrossRef]
- Modali, S.D.; Parekh, V.I.; Kebebew, E.; Agarwal, S.K. Epigenetic regulation of the lncRNA MEG3 and its target c-MET in pancreatic neuroendocrine tumors. Mol. Endocrinol. 2015, 29, 224–237. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Y.; Feng, H.M. MEG3 suppresses human pancreatic neuroendocrine tumor cells growth and metastasis by down-regulation of Mir-183. Cell Physiol. Biochem. 2017, 44, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Li, Z.; Zheng, S.; Chen, H.; Zhao, X.; Gao, W.; Bi, Z.; You, K.; Wang, Y.; Li, W.; et al. The long non-coding RNA HOTAIR affects the radiosensitivity of pancreatic ductal adenocarcinoma by regulating the expression of Wnt inhibitory factor 1. Tumour Biol. 2016, 37, 3957–3967. [Google Scholar] [CrossRef]
- Ji, P.; Diederichs, S.; Wang, W.; Böing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [Green Version]
- Muller-Tidow, C.; Diederichs, S.; Thomas, M.; Serve, H. Genome-wide screening for prognosis-predicting genes in early-stage non-small-cell lung cancer. Lung Cancer 2004, 45 (Suppl. 2), S145–S150. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Wang, Y.; Li, H.; Chen, L.; Liu, Q. Regulatory networks of LncRNA MALAT-1 in cancer. Cancer Manag. Res. 2020, 12, 10181–10198. [Google Scholar] [CrossRef] [PubMed]
- Pei, C.; Gong, X.; Zhang, Y. LncRNA MALAT-1 promotes growth and metastasis of epithelial ovarian cancer via sponging microrna-22. Am. J. Transl. Res. 2020, 12, 6977–6987. [Google Scholar] [PubMed]
- Liu, J.H.; Chen, G.; Dang, Y.W.; Li, C.J.; Luo, D.Z. Expression and prognostic significance of lncRNA MALAT1 in pancreatic cancer tissues. Asian Pac. J. Cancer Prev. 2014, 15, 2971–2977. [Google Scholar] [CrossRef] [PubMed]
- Filippova, E.A.; Fridman, M.V.; Burdennyy, A.M.; Loginov, V.I.; Pronina, I.V.; Lukina, S.S.; Dmitriev, A.A.; Braga, E.A. Long noncoding RNA GAS5 in breast cancer: Epigenetic mechanisms and biological functions. Int. J. Mol. Sci. 2021, 22, 6810. [Google Scholar] [CrossRef]
- Pickard, M.R.; Williams, G.T. Molecular and cellular mechanisms of action of tumour suppressor GAS5 LncRNA. Genes 2015, 6, 484–499. [Google Scholar] [CrossRef] [Green Version]
- Ghafouri-Fard, S.; Taheri, M. Maternally expressed gene 3 (MEG3): A tumor suppressor long non coding RNA. Biomed. Pharm. 2019, 118, 109129. [Google Scholar] [CrossRef]
- Kallen, A.N.; Zhou, X.B.; Xu, J.; Qiao, C.; Ma, J.; Yan, L.; Lu, L.; Liu, C.; Yi, J.S.; Zhang, H.; et al. The imprinted H19 lncRNA antagonizes let-7 microRNAs. Mol. Cell 2013, 52, 101–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onagoruwa, O.T.; Pal, G.; Ochu, C.; Ogunwobi, O.O. Oncogenic role of PVT1 and therapeutic implications. Front. Oncol. 2020, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Ghafouri-Fard, S.; Dashti, S.; Taheri, M. The HOTTIP (HOXA transcript at the distal tip) lncRNA: Review of oncogenic roles in human. Biomed. Pharm. 2020, 127, 110158. [Google Scholar] [CrossRef]
- Shang, Q.; Yang, Z.; Jia, R.; Ge, S. The novel roles of circRNAs in human cancer. Mol. Cancer 2019, 18, 6. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Song, W.; Yang, X.; Wang, J.; Zhang, R.; Zhang, Z.; Zhang, H.; Li, H. Microarray expression profile of circular RNAs in human pancreatic ductal adenocarcinoma. Genom Data 2015, 5, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Xu, X.; Ouyang, Y.; Wang, Y.; Yang, J.; Yin, L.; Ge, J.; Wang, H. Microarray expression profile analysis of circular RNAs in pancreatic cancer. Mol. Med. Rep. 2018, 17, 7661–7671. [Google Scholar] [CrossRef]
- Liu, L.; Liu, F.B.; Huang, M.; Xie, K.; Xie, Q.S.; Liu, C.H.; Shen, M.J.; Huang, Q. Circular RNA ciRS-7 promotes the proliferation and metastasis of pancreatic cancer by regulating miR-7-mediated EGFR/STAT3 signaling pathway. Hepatobiliary Pancreat. Dis. Int. 2019, 18, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, M.; Lu, H.; Peng, T. Up-regulation of circEIF6 contributes to pancreatic cancer development through targeting miR-557/SLC7A11/PI3K/AKT signaling. Cancer Manag. Res. 2021, 13, 247–258. [Google Scholar] [CrossRef]
- Wong, C.H.; Lou, U.K.; Li, Y.; Chan, S.L.; Tong, J.H.; To, K.F.; Chen, Y. CircFOXK2 promotes growth and metastasis of pancreatic ductal adenocarcinoma by complexing with rna-binding proteins and sponging MiR-942. Cancer Res. 2020, 80, 2138–2149. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Zhou, Q.; Su, D.; Luo, Y.; Fu, Z.; Huang, L.; Li, Z.; Jiang, D.; Kong, Y.; Li, Z.; et al. Circular RNA circBFAR promotes the progression of pancreatic ductal adenocarcinoma via the miR-34b-5p/MET/Akt axis. Mol. Cancer 2020, 19, 83. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Z.; Zhang, M.; Wang, B.; Ye, J.; Zhang, Y.; Tang, D.; Ma, D.; Jin, W.; Li, X.; et al. Circ-ASH2L promotes tumor progression by sponging miR-34a to regulate Notch1 in pancreatic ductal adenocarcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 466. [Google Scholar] [CrossRef] [Green Version]
- Ling, S.; He, Y.; Li, X.; Hu, M.; Ma, Y.; Li, Y.; Lu, Z.; Shen, S.; Kong, B.; Zou, X.; et al. CircRHOT1 mediated cell proliferation, apoptosis and invasion of pancreatic cancer cells by sponging miR-125a-3p. J. Cell Mol. Med. 2020, 24, 9881–9889. [Google Scholar] [CrossRef]
- Qu, S.; Hao, X.; Song, W.; Niu, K.; Yang, X.; Zhang, X.; Shang, R.; Wang, Q.; Li, H.; Liu, Z. Circular RNA circRHOT1 is upregulated and promotes cell proliferation and invasion in pancreatic cancer. Epigenomics 2019, 11, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Shi, Y.; Zhang, Y.; Sun, J. CircRNA_100782 regulates pancreatic carcinoma proliferation through the IL6-STAT3 pathway. Onco Targets Ther. 2017, 10, 5783–5794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Fang, Y.; Chen, P.; Xu, Y.; Zhou, W.; Rong, Y.; Li, J.A.; Chen, W.; Lou, W. Upregulated circRNA hsa_circ_0071036 promotes tumourigenesis of pancreatic cancer by sponging miR-489 and predicts unfavorable characteristics and prognosis. Cell Cycle 2021, 4, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Rong, W.; Bai, L.; Cui, H.; Zhang, S.; Li, Y.; Chen, D.; Meng, X. Upregulated circular RNA circ_0007534 indicates an unfavorable prognosis in pancreatic ductal adenocarcinoma and regulates cell proliferation, apoptosis, and invasion by sponging miR-625 and miR-892b. J. Cell Biochem. 2019, 120, 3780–3789. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Li, Y.; Luo, Y.; Zhu, J.; Zheng, H.; Gao, B.; Guo, X.; Li, Z.; Chen, R.; Chen, C. circNFIB1 inhibits lymphangiogenesis and lymphatic metastasis via the miR-486-5p/PIK3R1/VEGF-C axis in pancreatic cancer. Mol. Cancer 2020, 19, 82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tan, P.; Zhuang, Y.; Du, L. hsa_circRNA_001587 upregulates SLC4A4 expression to inhibit migration, invasion, and angiogenesis of pancreatic cancer cells via binding to microRNA-223. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G703–G717. [Google Scholar] [CrossRef]
- Li, H.; Hao, X.; Wang, H.; Liu, Z.; He, Y.; Pu, M.; Zhang, H.; Yu, H.; Duan, J.; Qu, S. Circular RNA expression profile of pancreatic ductal adenocarcinoma revealed by microarray. Cell Physiol. Biochem. 2016, 40, 1334–1344. [Google Scholar] [CrossRef]
- Qian, Y.; Gong, Y.; Fan, Z.; Luo, G.; Huang, Q.; Deng, S.; Cheng, H.; Jin, K.; Ni, Q.; Yu, X.; et al. Molecular alterations and targeted therapy in pancreatic ductal adenocarcinoma. J. Hematol. Oncol. 2020, 13, 130. [Google Scholar] [CrossRef]
- Buscail, L.; Bournet, B.; Cordelier, P. Role of oncogenic KRAS in the diagnosis, prognosis and treatment of pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Van Mackelenbergh, M.G.; Stroes, C.I.; Spijker, R.; van Eijck, C.H.J.; Wilmink, J.W.; Bijlsma, M.F.; van Laarhoven, H.W.M. Clinical trials targeting the stroma in pancreatic cancer: A systematic review and meta-analysis. Cancers 2019, 11, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathi, M.; Ghafouri-Fard, S.; Abak, A.; Taheri, M. Emerging roles of miRNAs in the development of pancreatic cancer. Biomed. Pharm. 2021, 141, 111914. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ju, J.; Ni, B.; Wang, H. The emerging role of miR-506 in cancer. Oncotarget 2016, 7, 62778–62788. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, J.Y.; Zhou, S.Y.; Yang, S.J.; Zhong, S.L. Circular RNA expression in pancreatic ductal adenocarcinoma. Oncol. Lett. 2019, 18, 2923–2930. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Ye, T.; Liu, H.; Lv, P.; Duan, C.; Wu, X.; Jiang, K.; Lu, H.; Xia, D.; Peng, E.; et al. Expression profiles, biological functions and clinical significance of circRNAs in bladder cancer. Mol. Cancer 2021, 20, 4. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, F. Computational strategies for exploring circular RNAs. Trends Genet. 2018, 34, 389–400. [Google Scholar] [CrossRef]
- Xu, Z.; Shen, J.; Hua, S.; Wan, D.; Chen, Q.; Han, Y.; Ren, R.; Liu, F.; Du, Z.; Guo, X.; et al. High-throughput sequencing of circRNAs reveals novel insights into mechanisms of nigericin in pancreatic cancer. BMC Genom. 2019, 20, 716. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.B.; Kjems, J.; Damgaard, C.K. Circular RNA and miR-7 in cancer. Cancer Res. 2013, 73, 5609–5612. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Li, X.; Xu, Q.; Lv, S.; Li, J.; Ma, Q. Role of glial cell line-derived neurotrophic factor in perineural invasion of pancreatic cancer. Biochim. Biophys. Acta 2012, 1826, 112–120. [Google Scholar] [CrossRef]
- Van, D.G.; Griffin, M.D.W.; Putoczki, T.L. Emerging roles for the IL-6 family of cytokines in pancreatic cancer. Clin. Sci. 2020, 134, 2091–2115. [Google Scholar]
- Shi, H.; Li, H.; Zhen, T.; Dong, Y.; Pei, X.; Zhang, X. hsa_circ_001653 implicates in the development of pancreatic ductal adenocarcinoma by regulating microrna-377-mediated HOXC6 Axis. Mol. Nucleic Acids 2020, 20, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, T.; Yan, L.; Qu, L. A novel prognostic biomarker for pancreatic ductal adenocarcinoma: Hsa_circ_0001649. Gene 2018, 675, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Xie, H.; Zhang, L.; Zhao, Q.; Lü, J.; Yu, Z. Piwi-interacting RNAs: A new class of regulator in human breast cancer. Front. Oncol. 2021, 11, 695077. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, C.G.; Azevedo Dos Santos, P.J.; Vidal, A.F.; Santos, S.; Ribeiro-Dos-Santos, Ã. piRNAs in gastric cancer: A new approach towards translational research. Int. J. Mol. Sci. 2020, 21, 2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Deng, H.; Xiao, B.; Zhou, H.; Zhou, F.; Shen, Z.; Guo, J. piR-823, a novel non-coding small RNA, demonstrates in vitro and in vivo tumor suppressive activity in human gastric cancer cells. Cancer Lett. 2012, 315, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.; Du, M.; Jiang, X.; Wu, Y.; Ben, S.; Zheng, R.; Chu, H.; Li, S.; Zhang, Z.; Wang, M. Systematic evaluation of the effects of genetic variants on PIWI-interacting RNA expression across 33 cancer types. Nucleic Acids Res. 2021, 49, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Kimchi, E.T.; Manjunath, Y.; Gajagowni, S.; Stuckel, A.J.; Kaifi, J.T. RNA cargos in extracellular vesicles derived from blood serum in pancreas associated conditions. Sci. Rep. 2020, 10, 2800. [Google Scholar] [CrossRef] [Green Version]
- Yee, N.S.; Zhang, S.; He, H.Z.; Zheng, S.Y. Extracellular vesicles as potential biomarkers for early detection and diagnosis of pancreatic cancer. Biomedicines 2020, 8, 581. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Huang, X.; Woodcock, M.; Du, M.; Dittmar, R.; Wang, Y.; Tsai, S.; Kohli, M.; Boardman, L.; Patel, T.; et al. Plasma extracellular RNA profiles in healthy and cancer patients. Sci. Rep. 2016, 6, 19413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, S.; Raulefs, S.; Bruns, P.; Afonso-Grunz, F.; Plotner, A.; Thermann, R.; Jäger, C.; Schlitter, A.M.; Kong, B.; Regel, I.; et al. Next-generation sequencing reveals novel differentially regulated mRNAs, lncRNAs, miRNAs, sdRNAs and a piRNA in pancreatic cancer. Mol. Cancer 2015, 14, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukowati, C.H.C.; Cabral, L.K.D.; Tiribelli, C.; Pascut, D. Circulating long and circular noncoding RNA as non-invasive diagnostic tools of hepatocellular carcinoma. Biomedicines 2021, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Zhang, S.; Yao, Z.; Liu, Y.; Lin, K.; Zhao, Z.; Zhu, Y. The diagnostic and prognostic value of exosome-derived long non-coding RNAs in cancer patients: A meta-analysis. Clin. Exp. Med. 2020, 20, 339–348. [Google Scholar] [CrossRef]
- Galamb, O.; Barták, B.K.; Kalmár, A.; Nagy, Z.B.; Szigeti, K.A.; Tulassay, Z.; Igaz, P.; Molnár, B. Diagnostic and prognostic potential of tissue and circulating long non-coding RNAs in colorectal tumors. World J. Gastroenterol. 2019, 25, 5026–5048. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.J.; Ying, X.L.; Jiang, J.H.; Xu, Y.H. Prostate cancer antigen 3 as a biomarker in the urine for prostate cancer diagnosis: A meta-analysis. J. Cancer Res. 2014, 10, C218–C221. [Google Scholar] [CrossRef]
- Ou, Z.L.; Luo, Z.; Lu, Y.B. Long non-coding RNA HULC as a diagnostic and prognostic marker of pancreatic cancer. World J. Gastroenterol. 2019, 25, 6728–6742. [Google Scholar] [CrossRef]
- Takahashi, K.; Ota, Y.; Kogure, T.; Suzuki, Y.; Iwamoto, H.; Yamakita, K.; Kitano, Y.; Fujii, S.; Haneda, M.; Patel, T.; et al. Circulating extracellular vesicle-encapsulated HULC is a potential biomarker for human pancreatic cancer. Cancer Sci. 2020, 111, 98–111. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Feng, W.; Liu, W.; Kong, X.; Li, L.; He, J.; Wang, D.; Zhang, M.; Zhou, G.; Xu, W.; et al. Circulating lncRNA ABHD11-AS1 serves as a biomarker for early pancreatic cancer diagnosis. J. Cancer 2019, 10, 3746–3756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortoglou, M.; Tabin, Z.K.; Arisan, E.D.; Kocher, H.M.; Uysal-Onganer, P. Non-coding RNAs in pancreatic ductal adenocarcinoma: New approaches for better diagnosis and therapy. Transl. Oncol. 2021, 14, 101090. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Jiang, J.; Qiao, Z. The lncRNA SNHG15/miR-18a-5p axis promotes cell proliferation in ovarian cancer through activating Akt/mTOR signaling pathway. J. Cell Biochem. 2020, 121, 4699–4710. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Feng, Y.; Wang, J.; Liang, Y.; Zou, W. Long non-coding RNA SNHG15 in various cancers: A meta and bioinformatic analysis. BMC Cancer 2020, 20, 1156. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Lin, H.; Kang, L.; Huang, P.; Huang, J.; Cai, J.; Xian, Z.; Zhu, P.; Huang, M.; Wang, L.; et al. Aberrant expression of long noncoding RNA SNHG15 correlates with liver metastasis and poor survival in colorectal cancer. J. Cell Physiol. 2019, 234, 7032–7039. [Google Scholar] [CrossRef]
- Zhang, J.H.; Wei, H.W.; Yang, H.G. Long noncoding RNA SNHG15, a potential prognostic biomarker for hepatocellular carcinoma. Eur. Rev. Med. Pharm. Sci. 2016, 20, 1720–1724. [Google Scholar]
- Guo, X.B.; Yin, H.S.; Wang, J.Y. Evaluating the diagnostic and prognostic value of long non-coding RNA SNHG15 in pancreatic ductal adenocarcinoma. Eur. Rev. Med. Pharm. Sci. 2018, 22, 5892–5898. [Google Scholar]
- Li, S.P.; Xu, H.X.; Yu, Y.; He, J.D.; Wang, Z.; Xu, Y.J.; Wang, C.Y.; Zhang, H.M.; Zhang, R.X.; Zhang, J.J.; et al. LncRNA HULC enhances epithelial-mesenchymal transition to promote tumorigenesis and metastasis of hepatocellular carcinoma via the miR-200a-3p/ZEB1 signaling pathway. Oncotarget 2016, 7, 42431–42446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, T.; Wu, J.; Hu, Y.; Zhang, M.; He, J. Long non-coding RNA HULC stimulates the epithelial-mesenchymal transition process and vasculogenic mimicry in human glioblastoma. Cancer Med. 2021, 10, 5270–5282. [Google Scholar] [CrossRef]
- Jin, C.; Shi, W.; Wang, F.; Shen, X.; Qi, J.; Cong, H.; Yuan, J.; Shi, L.; Zhu, B.; Luo, X.; et al. Long non-coding RNA HULC as a novel serum biomarker for diagnosis and prognosis prediction of gastric cancer. Oncotarget 2016, 7, 51763–51772. [Google Scholar] [CrossRef] [Green Version]
- Zang, X.; Gu, J.; Zhang, J.; Shi, H.; Hou, S.; Xu, X.; Chen, Y.; Zhang, Y.; Mao, F.; Qian, H.; et al. Exosome-transmitted lncRNA UFC1 promotes non-small-cell lung cancer progression by EZH2-mediated epigenetic silencing of PTEN expression. Cell Death Dis. 2020, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Sun, Q.Q.; Liu, T.X.; Lu, K.; Zhang, N.; Zhu, Y.; Chen, M. Serum lncRNA-UFC1 as a potential biomarker for diagnosis and prognosis of pancreatic cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 4125–4129. [Google Scholar]
- Xu, J.H.; Wang, Y.; Xu, D. Hsa_circ_001569 is an unfavorable prognostic factor and promotes cell proliferation and metastasis by modulating PI3K-AKT pathway in breast cancer. Cancer Biomark. 2019, 25, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xue, L.; Song, C.; Liu, F.; Jiang, T.; Yang, X. Overexpression of circular RNA circ_001569 indicates poor prognosis in hepatocellular carcinoma and promotes cell growth and metastasis by sponging miR-411-5p and miR-432-5p. Biochem. Biophys. Res. Commun. 2018, 503, 2659–2665. [Google Scholar] [CrossRef]
- Xie, H.; Ren, X.; Xin, S.; Lan, X.; Lu, G.; Lin, Y.; Yang, S.; Zeng, Z.; Liao, W.; Ding, Y.Q.; et al. Emerging roles of circRNA_001569 targeting miR-145 in the proliferation and invasion of colorectal cancer. Oncotarget 2016, 7, 26680–26691. [Google Scholar] [CrossRef] [PubMed]
- Mai, S.; Zhang, Z.; Mi, W. Upregulation of circ_PVT1 and circ_001569 indicate unfavorable prognosis in colorectal cancer. Ann. Clin. Lab. Sci. 2021, 51, 55–60. [Google Scholar] [PubMed]
- Shen, X.; Chen, Y.; Li, J.; Huang, H.; Liu, C.; Zhou, N. Identification of Circ_001569 as a potential biomarker in the diagnosis and prognosis of pancreatic cancer. Technol. Cancer Res. Treat. 2021, 20, 1533033820983302. [Google Scholar] [CrossRef]
- Yang, F.; Liu, D.Y.; Guo, J.T.; Ge, N.; Zhu, P.; Liu, X.; Wang, S.; Wang, G.X.; Sun, S.Y. Circular RNA circ-LDLRAD3 as a biomarker in diagnosis of pancreatic cancer. World J. Gastroenterol. 2017, 23, 8345–8354. [Google Scholar] [CrossRef]
- Davis, M.E.; Zuckerman, J.E.; Choi, C.H.; Seligson, D.; Tolcher, A.; Alabi, C.A.; Yen, Y.; Heidel, J.D.; Ribas, A. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 2010, 464, 1067–1070. [Google Scholar] [CrossRef]
- Michalik, K.M.; You, X.; Manavski, Y.; Doddaballapur, A.; Zörnig, M.; Braun, T.; John, D.; Ponomareva, Y.; Chen, W.; Uchida, S.; et al. Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth. Circ. Res. 2014, 114, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Kandimalla, R.; Wallen, M.; Tyagi, N.; Wilcher, S.; Yan, J.; Schultz, D.J.; Spencer, W.; et al. Exosome-mediated delivery of RNA and DNA for gene therapy. Cancer Lett. 2021, 505, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Orefice, N.S. Development of new strategies using extracellular vesicles loaded with exogenous nucleic acid. Pharmaceutics 2020, 12, 705. [Google Scholar] [CrossRef]
- Maheshwari, R.; Tekade, M.; Gondaliya, P.; Kalia, K.; D’Emanuele, A.; Tekade, R.K. Recent advances in exosome-based nanovehicles as RNA interference therapeutic carriers. Nanomedicine 2017, 12, 2653–2675. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zhang, G.; Zhang, L.; Hu, Y.; Zhang, K.; Sun, X.; Zhao, C.; Li, H.; Li, Y.M.; Zhao, J. Mesenchymal stem cells deliver exogenous miR-21 via exosomes to inhibit nucleus pulposus cell apoptosis and reduce intervertebral disc degeneration. J. Cell Mol. Med. 2018, 22, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, D.; Hua, R.; Zhang, J.; Liu, W.; Huo, Y.; Cheng, Y.; Hong, J.; Sun, Y. Long non-coding RNAs expressed in pancreatic ductal adenocarcinoma and lncRNA BC008363 an independent prognostic factor in PDAC. Pancreatology 2014, 14, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Shao, F.; Huang, M.; Meng, F.; Huang, Q. Circular RNA Signature predicts gemcitabine resistance of pancreatic ductal adenocarcinoma. Front. Pharm. 2018, 9, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Yao, Y.; Gao, P.; Cui, Y. Upregulated circular RNA circ_0030235 predicts unfavorable prognosis in pancreatic ductal adenocarcinoma and facilitates cell progression by sponging miR-1253 and miR-1294. Biochem. Biophys. Res. Commun. 2019, 509, 138–142. [Google Scholar] [CrossRef]
- Zhou, M.; Ye, Z.; Gu, Y.; Tian, B.; Wu, B.; Li, J. Genomic analysis of drug resistant pancreatic cancer cell line by combining long non-coding RNA and mRNA expression profling. Int. J. Clin. Exp. Pathol. 2015, 8, 38–52. [Google Scholar]
- Zhan, H.X.; Wang, Y.; Li, C.; Xu, J.W.; Zhou, B.; Zhu, J.K.; Han, H.F.; Wang, L.; Wang, Y.S.; Hu, S.Y. LincRNA-ROR promotes invasion, metastasis and tumor growth in pancreatic cancer through activating ZEB1 pathway. Cancer Lett. 2016, 374, 261–271. [Google Scholar] [CrossRef]
- Li, C.; Zhao, Z.; Zhou, Z.; Liu, R. Linc-ROR confers gemcitabine resistance to pancreatic cancer cells via inducing autophagy and modulating the miR-124/PTBP1/PKM2 axis. Cancer Chemother. Pharm. 2016, 78, 1199–1207. [Google Scholar] [CrossRef]
- Liu, Y.; Xia, L.; Dong, L.; Wang, J.; Xiao, Q.; Yu, X.; Zhu, H. CircHIPK3 promotes gemcitabine (GEM) Resistance in pancreatic cancer cells by sponging miR-330-5p and targets RASSF1. Cancer Manag. Res. 2020, 12, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Yu, Y.; Ding, F. Microarray analysis of circular RNA expression profiles associated with gemcitabine resistance in pancreatic cancer cells. Oncol. Rep. 2018, 40, 395–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—Challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Cheng, C.J.; Bahal, R.; Babar, I.A.; Pincus, Z.; Barrera, F.; Liu, C.; Svoronos, A.; Braddock, D.T.; Glazer, P.M.; Engelman, D.M.; et al. MicroRNA silencing for cancer therapy targeted to the tumour microenvironment. Nature 2015, 518, 107–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Ansarullah; Kumar, D.; Jaggi, M.; Chauhan, S.C. Targeting microRNAs in pancreatic cancer: Microplayers in the big game. Cancer Res. 2013, 73, 6541–6547. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.L.; Liu, D.J.; Yan, T.T.; Yang, J.Y.; Yang, M.W.; Li, J.; Huo, Y.M.; Liu, W.; Zhang, J.F.; Hong, J.; et al. Analysis of long non-coding RNA expression profiles in pancreatic ductal adenocarcinoma. Sci. Rep. 2016, 6, 33535. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Chen, J.; Zhou, Y.; Fu, Z.; Zhou, Q.; Wang, Y.; Gao, W.; Zheng, S.; Zhao, X.; Chen, T.; et al. High expression of AFAP1-AS1 is associated with poor survival and short-term recurrence in pancreatic ductal adenocarcinoma. J. Transl. Med. 2015, 13, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, C.; Ye, H.; Wang, W.; Sun, M.; Zhang, J.; Zhao, Z.; Jiang, G. Circular RNA ADAM9 facilitates the malignant behaviours of pancreatic cancer by sponging miR-217 and upregulating PRSS3 expression. Artif. Cells Nanomed. 2019, 47, 3920–3928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Zhang, C.; Chen, Y.; Gao, S. Downregulation of circular RNA circ-LDLRAD3 suppresses pancreatic cancer progression through miR-137-3p/PTN axis. Life Sci. 2019, 239, 116871. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.; Jiang, P.; Peng, M.; Zhang, X.; Chen, K.; Liu, H.; Bi, H.; Liu, X.; Li, X. Circular RNA IARS (circ-IARS) secreted by pancreatic cancer cells and located within exosomes regulates endothelial monolayer permeability to promote tumor metastasis. J. Exp. Clin. Cancer Res. 2018, 37, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Yanfang, W.; Li, J.; Jiang, P.; Peng, T.; Chen, K.; Zhao, X.; Zhang, Y.; Zhen, P.; Zhu, J.; et al. Tumor-released exosomal circular RNA PDE8A promotes invasive growth via the miR-338/MACC1/MET pathway in pancreatic cancer. Cancer Lett. 2018, 432, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 3, 209–249. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Xu, R.; Wang, C.; Qiu, J.; Ren, B.; You, L. Early screening and diagnosis strategies of pancreatic cancer: A comprehensive review. Cancer Commun. 2021. [Google Scholar] [CrossRef] [PubMed]
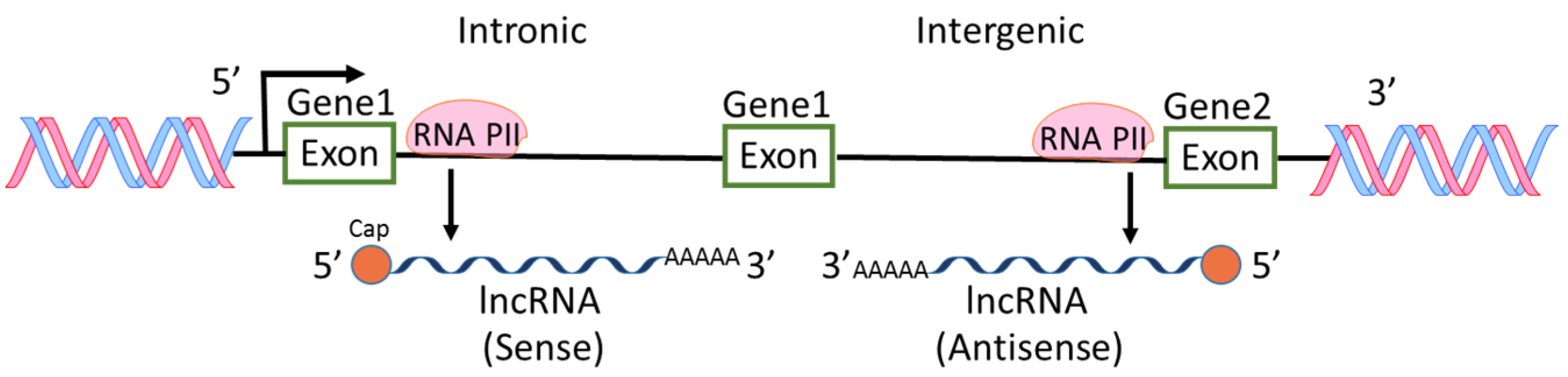
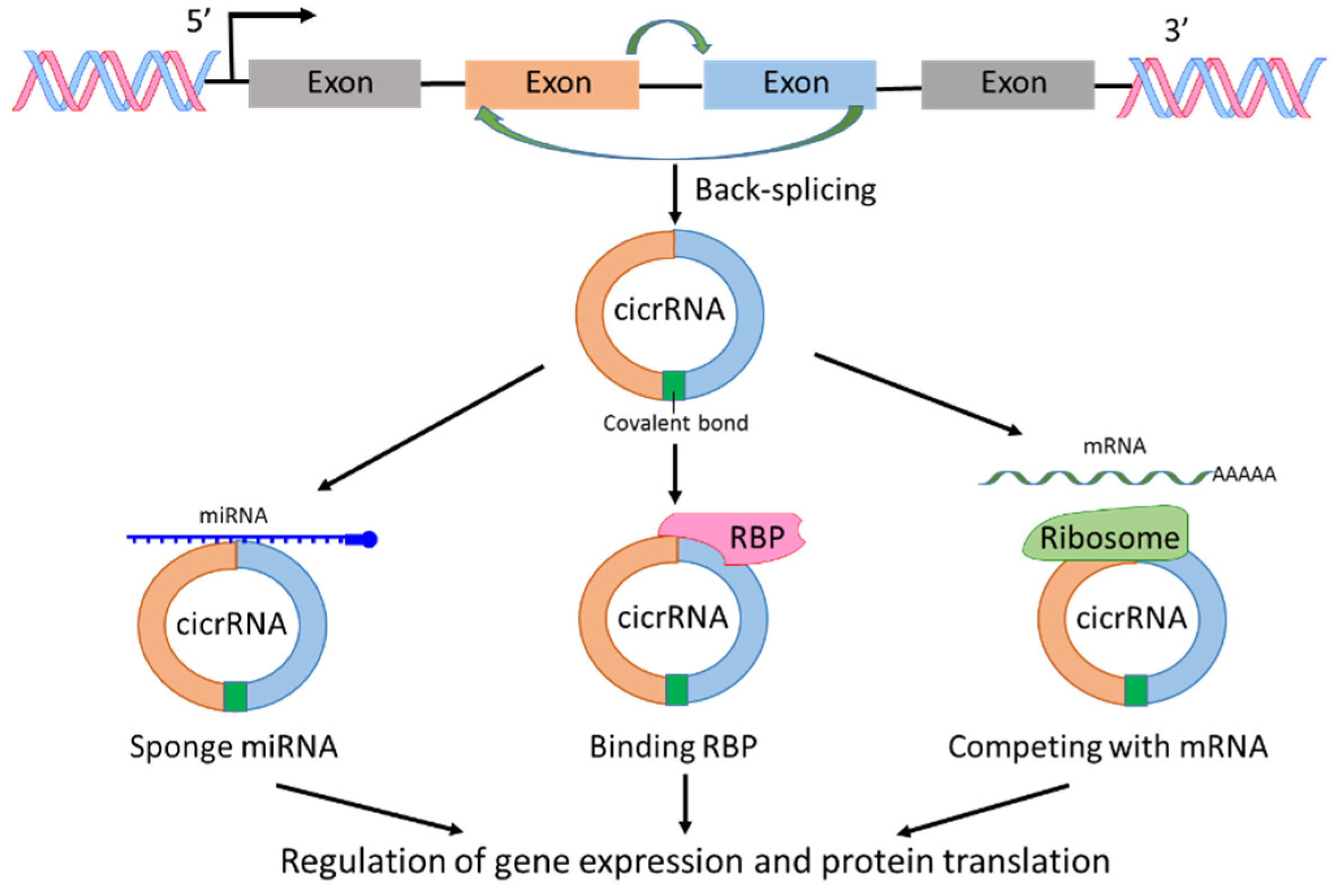
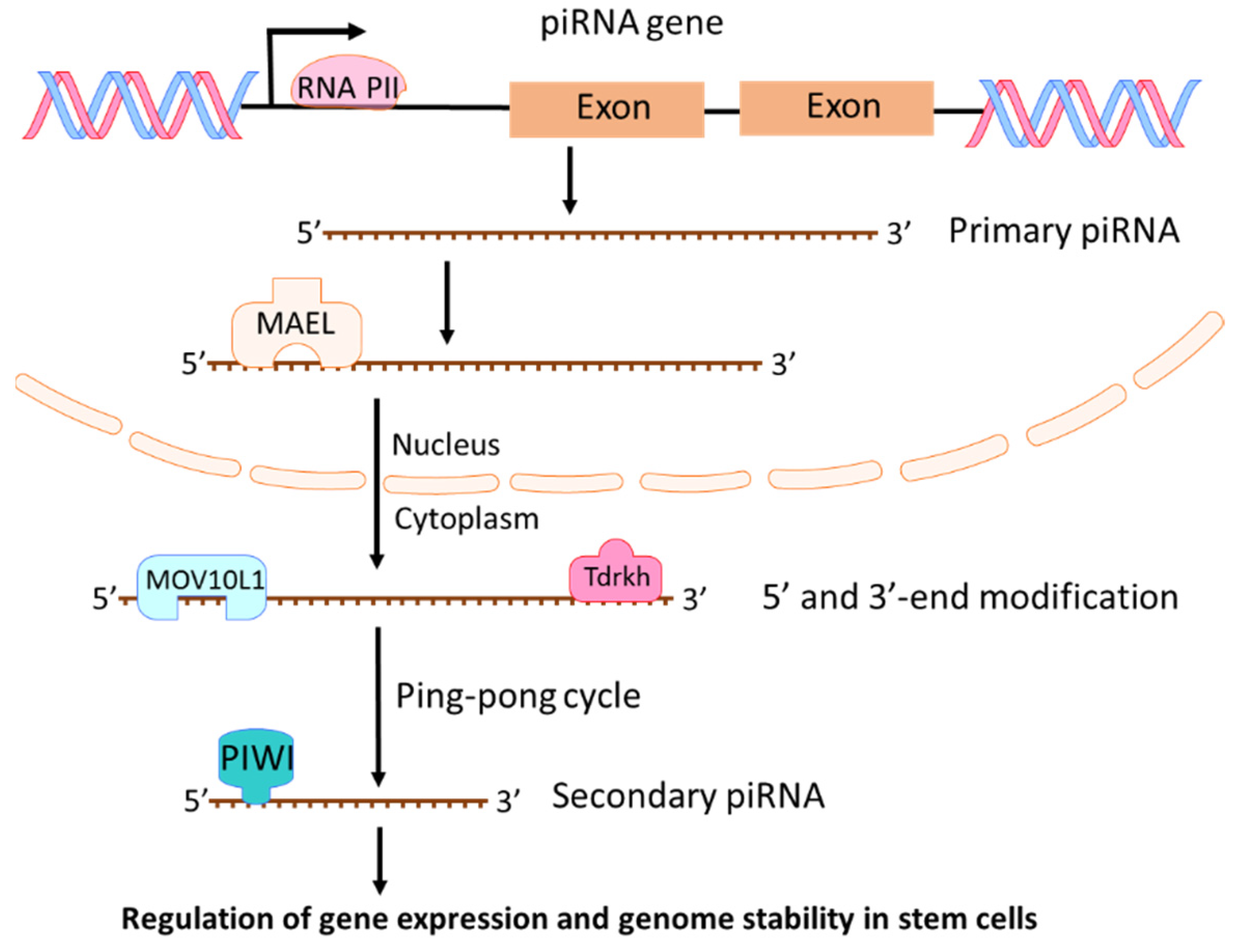
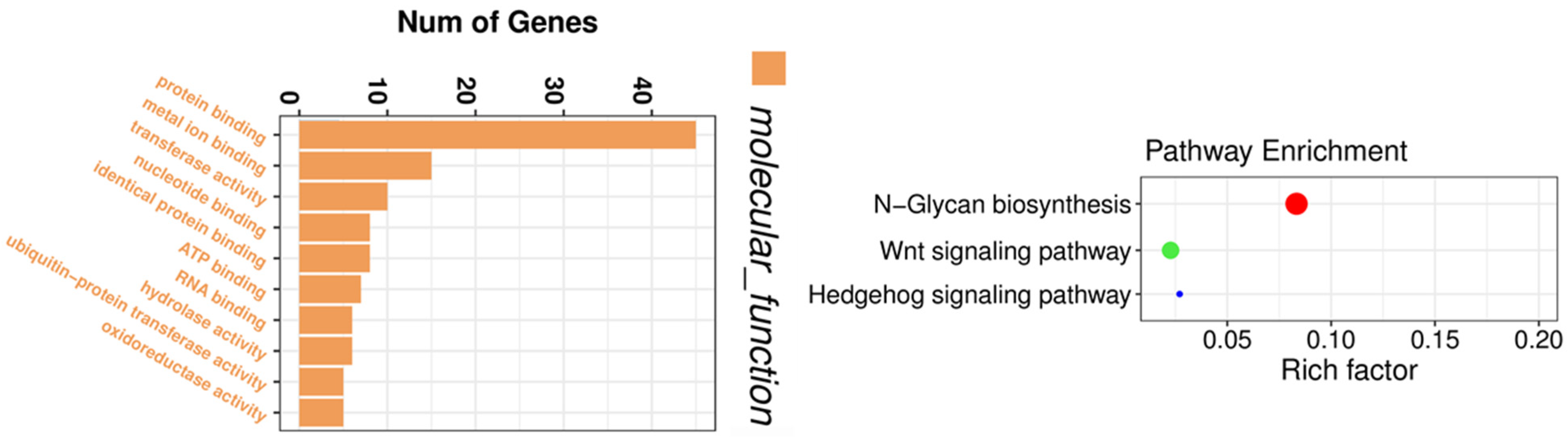
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).