
CCL20/TNF/VEGFA Cytokine Secretory Phenotype of Tumor-Associated Macrophages Is a Negative Prognostic Factor in Cutaneous Melanoma

 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Cohorts and Selection Criteria
2.2. Multicolor Fluorescence Confocal Microscopy
2.3. Cell Isolation from Human Melanomas
2.4. Monocyte Isolation and Cell Culture
2.5. In Vitro Measurements
2.6. Statistical Analyses
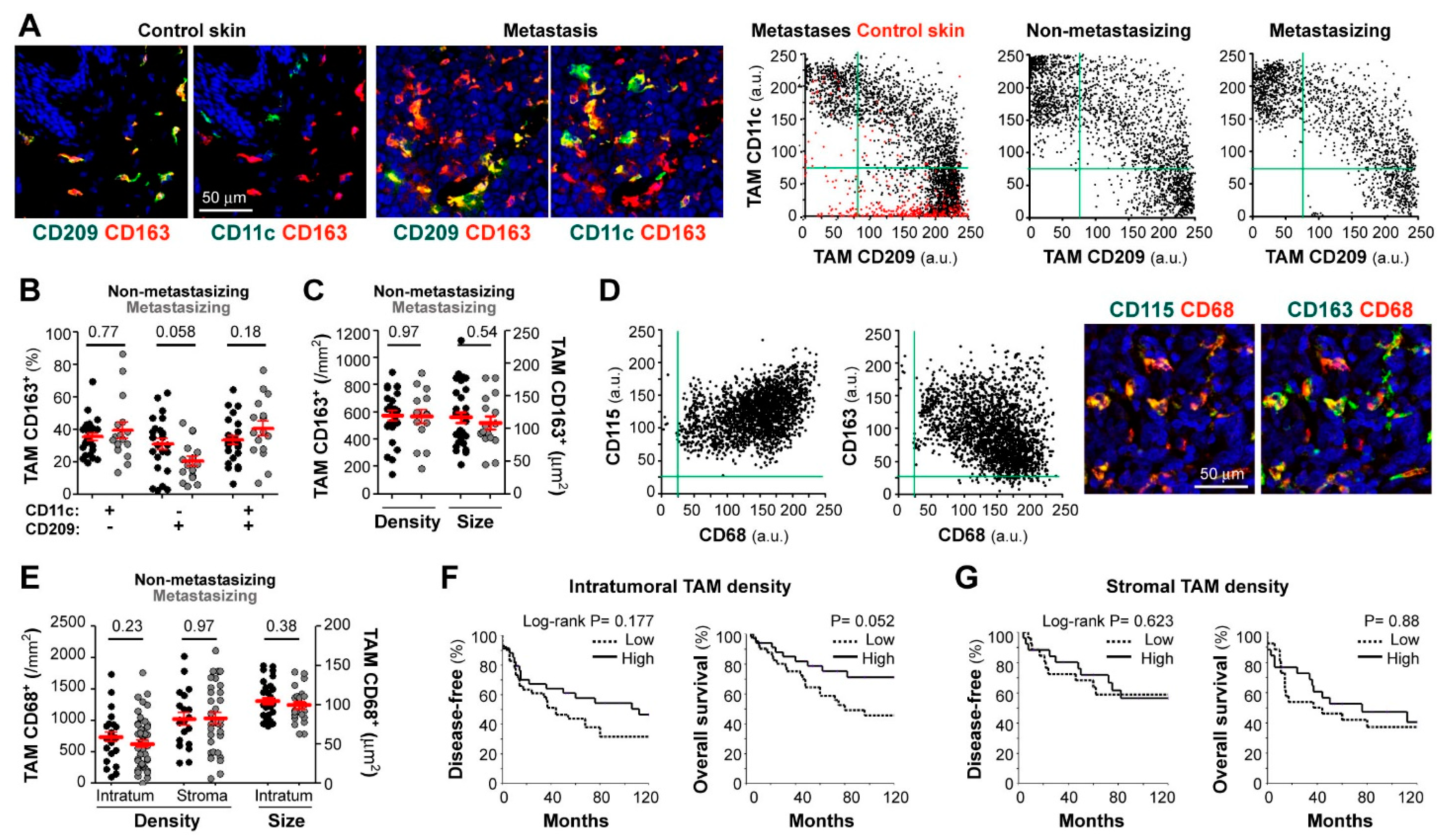
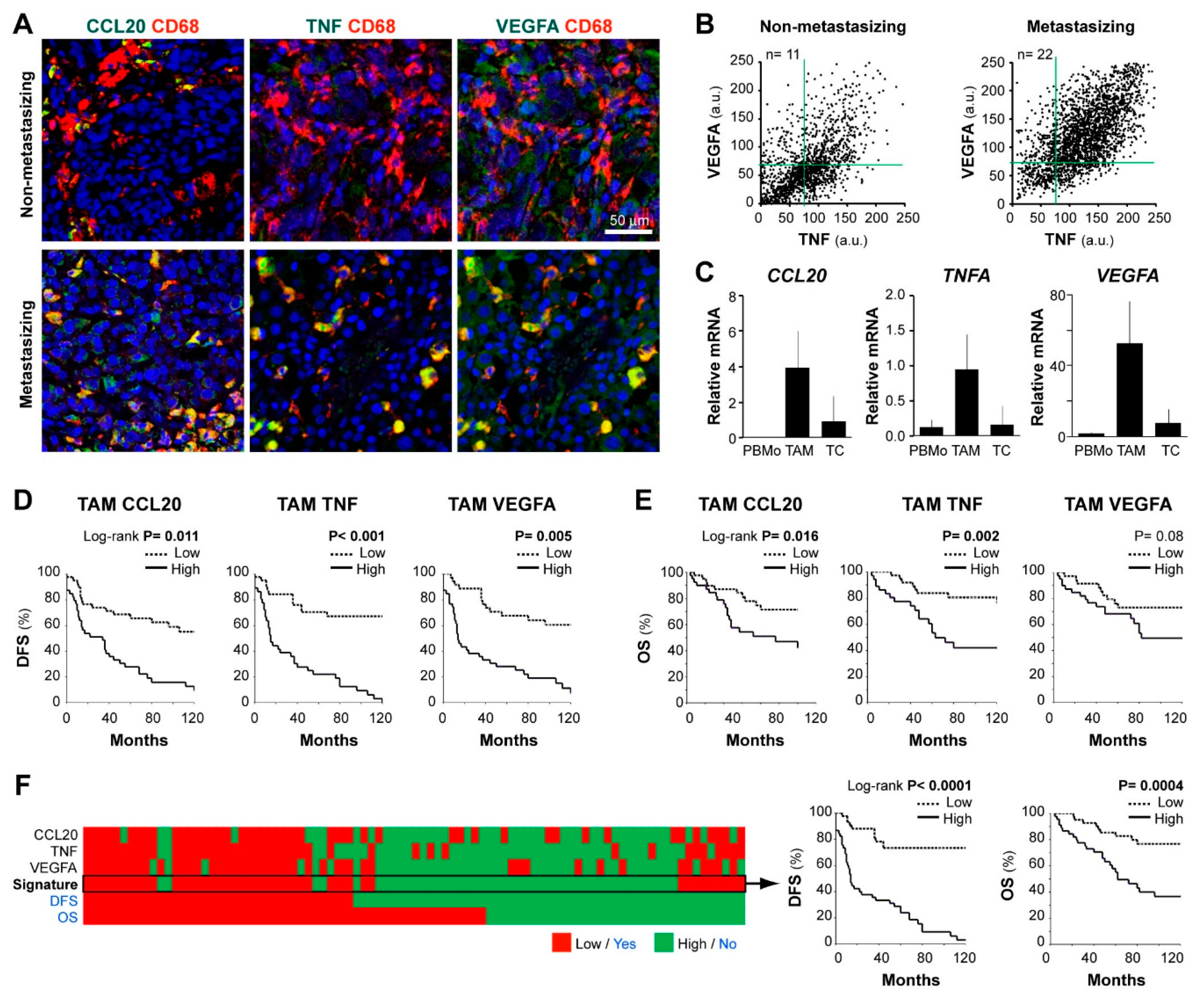
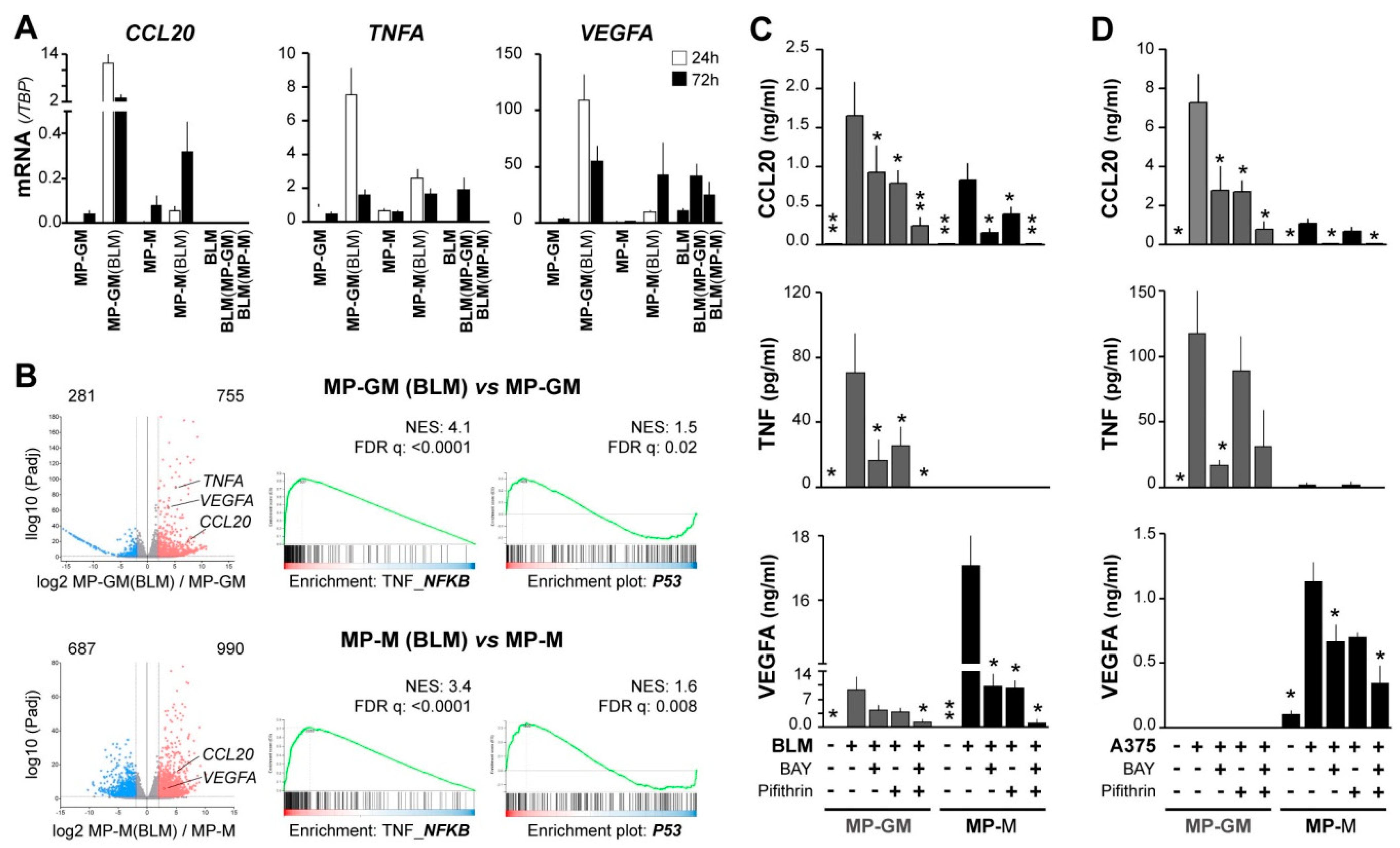
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Curti, B.D.; Faries, M.B. Recent Advances in the Treatment of Melanoma. N. Engl. J. Med. 2021, 384, 2229–2240. [Google Scholar] [CrossRef] [PubMed]
- Balch, C.M.; Gershenwald, J.E.; Soong, S.J.; Thompson, J.F.; Atkins, M.B.; Byrd, D.R.; Buzaid, A.C.; Cochran, A.J.; Coit, D.G.; Ding, S.; et al. Final version of 2009 AJCC melanoma staging and classification. J. Clin. Oncol. 2009, 27, 6199–6206. [Google Scholar] [CrossRef] [Green Version]
- Schuchter, L.M. Adjuvant Melanoma Therapy—Head-Spinning Progress. N. Engl. J. Med. 2017, 377, 1888–1890. [Google Scholar] [CrossRef]
- Miller, R.; Walker, S.; Shui, I.; Brandtmuller, A.; Cadwell, K.; Scherrer, E. Epidemiology and survival outcomes in stages II and III cutaneous melanoma: A systematic review. Melanoma Manag. 2020, 7, MMT39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquelot, N.; Pitt, J.M.; Enot, D.P.; Roberti, M.P.; Duong, C.P.M.; Rusakiewicz, S.; Eggermont, A.M.; Zitvogel, L. Immune biomarkers for prognosis and prediction of responses to immune checkpoint blockade in cutaneous melanoma. Oncoimmunology 2017, 6, e1299303. [Google Scholar] [CrossRef]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevrier, S.; Levine, J.H.; Zanotelli, V.R.T.; Silina, K.; Schulz, D.; Bacac, M.; Ries, C.H.; Ailles, L.; Jewett, M.A.S.; Moch, H.; et al. An Immune Atlas of Clear Cell Renal Cell Carcinoma. Cell 2017, 169, 736–749.e718. [Google Scholar] [CrossRef] [Green Version]
- Lavin, Y.; Kobayashi, S.; Leader, A.; Amir, E.D.; Elefant, N.; Bigenwald, C.; Remark, R.; Sweeney, R.; Becker, C.D.; Levine, J.H.; et al. Innate Immune Landscape in Early Lung Adenocarcinoma by Paired Single-Cell Analyses. Cell 2017, 169, 750–765.e717. [Google Scholar] [CrossRef] [Green Version]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Yang, M.; McKay, D.; Pollard, J.W.; Lewis, C.E. Diverse Functions of Macrophages in Different Tumor Microenvironments. Cancer Res. 2018, 78, 5492–5503. [Google Scholar] [CrossRef] [Green Version]
- Barros, M.H.; Hauck, F.; Dreyer, J.H.; Kempkes, B.; Niedobitek, G. Macrophage polarisation: An immunohistochemical approach for identifying M1 and M2 macrophages. PLoS ONE 2013, 8, e80908. [Google Scholar] [CrossRef] [Green Version]
- Ladanyi, A. Prognostic and predictive significance of immune cells infiltrating cutaneous melanoma. Pigment Cell Melanoma Res. 2015, 28, 490–500. [Google Scholar] [CrossRef]
- Jensen, T.O.; Schmidt, H.; Moller, H.J.; Hoyer, M.; Maniecki, M.B.; Sjoegren, P.; Christensen, I.J.; Steiniche, T. Macrophage markers in serum and tumor have prognostic impact in American Joint Committee on Cancer stage I/II melanoma. J. Clin. Oncol. 2009, 27, 3330–3337. [Google Scholar] [CrossRef]
- Storr, S.J.; Safuan, S.; Mitra, A.; Elliott, F.; Walker, C.; Vasko, M.J.; Ho, B.; Cook, M.; Mohammed, R.A.; Patel, P.M.; et al. Objective assessment of blood and lymphatic vessel invasion and association with macrophage infiltration in cutaneous melanoma. Mod. Pathol. 2012, 25, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Tremble, L.F.; McCabe, M.; Walker, S.P.; McCarthy, S.; Tynan, R.F.; Beecher, S.; Werner, R.; Clover, A.J.P.; Power, X.D.G.; Forde, P.F.; et al. Differential association of CD68(+) and CD163(+) macrophages with macrophage enzymes, whole tumour gene expression and overall survival in advanced melanoma. Br. J. Cancer 2020, 123, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Samaniego, R.; Gutierrez-Gonzalez, A.; Gutierrez-Seijo, A.; Sanchez-Gregorio, S.; Garcia-Gimenez, J.; Mercader, E.; Marquez-Rodas, I.; Aviles, J.A.; Relloso, M.; Sanchez-Mateos, P. CCL20 Expression by Tumor-Associated Macrophages Predicts Progression of Human Primary Cutaneous Melanoma. Cancer Immunol. Res. 2018, 6, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Baade, P.; Meng, X.; Youlden, D.; Aitken, J.; Youl, P. Time trends and latitudinal differences in melanoma thickness distribution in Australia, 1990–2006. Int. J. Cancer 2012, 130, 170–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samaniego, R.; Estecha, A.; Relloso, M.; Longo, N.; Escat, J.L.; Longo-Imedio, I.; Aviles, J.A.; del Pozo, M.A.; Puig-Kroger, A.; Sanchez-Mateos, P. Mesenchymal contribution to recruitment, infiltration, and positioning of leukocytes in human melanoma tissues. J. Investig. Dermatol. 2013, 133, 2255–2264. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Gonzalez, A.; Martinez-Moreno, M.; Samaniego, R.; Arellano-Sanchez, N.; Salinas-Munoz, L.; Relloso, M.; Valeri, A.; Martinez-Lopez, J.; Corbi, A.L.; Hidalgo, A.; et al. Evaluation of the potential therapeutic benefits of macrophage reprogramming in multiple myeloma. Blood 2016, 128, 2241–2252. [Google Scholar] [CrossRef] [Green Version]
- Estecha, A.; Sanchez-Martin, L.; Puig-Kroger, A.; Bartolome, R.A.; Teixido, J.; Samaniego, R.; Sanchez-Mateos, P. Moesin orchestrates cortical polarity of melanoma tumour cells to initiate 3D invasion. J. Cell Sci. 2009, 122, 3492–3501. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberzon, A.; Birger, C.; Thorvaldsdottir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Donadon, M.; Torzilli, G.; Cortese, N.; Soldani, C.; Di Tommaso, L.; Franceschini, B.; Carriero, R.; Barbagallo, M.; Rigamonti, A.; Anselmo, A.; et al. Macrophage morphology correlates with single-cell diversity and prognosis in colorectal liver metastasis. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef] [PubMed]
- Gartrell, R.D.; Marks, D.K.; Hart, T.D.; Li, G.; Davari, D.R.; Wu, A.; Blake, Z.; Lu, Y.; Askin, K.N.; Monod, A.; et al. Quantitative Analysis of Immune Infiltrates in Primary Melanoma. Cancer Immunol. Res. 2018, 6, 481–493. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGovern, N.; Schlitzer, A.; Gunawan, M.; Jardine, L.; Shin, A.; Poyner, E.; Green, K.; Dickinson, R.; Wang, X.N.; Low, D.; et al. Human dermal CD14(+) cells are a transient population of monocyte-derived macrophages. Immunity 2014, 41, 465–477. [Google Scholar] [CrossRef] [Green Version]
- Wunderlich, C.M.; Ackermann, P.J.; Ostermann, A.L.; Adams-Quack, P.; Vogt, M.C.; Tran, M.L.; Nikolajev, A.; Waisman, A.; Garbers, C.; Theurich, S.; et al. Obesity exacerbates colitis-associated cancer via IL-6-regulated macrophage polarisation and CCL-20/CCR-6-mediated lymphocyte recruitment. Nat. Commun. 2018, 9, 1646. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.P.; Sanchez-Laorden, B.; O’Brien, K.; Brunton, H.; Ferguson, J.; Young, H.; Dhomen, N.; Flaherty, K.T.; Frederick, D.T.; Cooper, Z.A.; et al. The immune microenvironment confers resistance to MAPK pathway inhibitors through macrophage-derived TNFalpha. Cancer Discov. 2014, 4, 1214–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta, C.; Rimoldi, M.; Raes, G.; Brys, L.; Ghezzi, P.; Di Liberto, D.; Dieli, F.; Ghisletti, S.; Natoli, G.; De Baetselier, P.; et al. Tolerance and M2 (alternative) macrophage polarization are related processes orchestrated by p50 nuclear factor kappaB. Proc. Natl. Acad. Sci. USA 2009, 106, 14978–14983. [Google Scholar] [CrossRef] [Green Version]
- Hagemann, T.; Lawrence, T.; McNeish, I.; Charles, K.A.; Kulbe, H.; Thompson, R.G.; Robinson, S.C.; Balkwill, F.R. “Re-educating” tumor-associated macrophages by targeting NF-kappaB. J. Exp. Med. 2008, 205, 1261–1268. [Google Scholar] [CrossRef]
- Lowe, J.M.; Menendez, D.; Bushel, P.R.; Shatz, M.; Kirk, E.L.; Troester, M.A.; Garantziotis, S.; Fessler, M.B.; Resnick, M.A. p53 and NF-kappaB coregulate proinflammatory gene responses in human macrophages. Cancer Res. 2014, 74, 2182–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Univariate | Disease-Free Survival | Overall Survival | ||||
|---|---|---|---|---|---|---|
| Gender (F vs. M) | 1.602 | 0.91–2.81 | 0.100 | 1.168 | 0.55–2.47 | 0.685 |
| Age (years) | 1.003 | 0.99–1.02 | 0.704 | 1.007 | 0.98–1.03 | 0.574 |
| Location (H/L vs. T) | 1.452 | 0.74–2.85 | 0.278 | 0.895 | 0.40–1.99 | 0.785 |
| Subtype (nod vs. others) | 0.831 | 0.46–1.52 | 0.547 | 1.305 | 0.60–2.82 | 0.498 |
| Ulceration (yes vs. no) | 1.354 | 0.76–2.41 | 0.301 | 2.582 | 1.15–5.82 | 0.022 |
| Breslow (mm) | 1.130 | 1.07–1.20 | <0.001 | 1.124 | 1.04–1.21 | 0.002 |
| Stage (II vs. III–IV) | 1.865 | 1.19–2.92 | 0.006 | 1.793 | 0.74–4.33 | 0.194 |
| TAM-CCL20 (MFI, each 10 a.u.) | 1.195 | 1.10–1.29 | <0.001 | 1.164 | 1.05–1.29 | 0.003 |
| TAM-TNF (MFI, each 10 a.u.) | 1.295 | 1.18–1.42 | <0.001 | 1.250 | 1.11–1.41 | 0.000 |
| TAM-VEGFA (MFI, each 10 a.u.) | 1.185 | 1.11–1.26 | <0.001 | 1.051 | 0.96–1.15 | 0.264 |
| TC-VEGFA (MFI, each 10 a.u.) | 1.050 | 0.98–1.13 | 0.189 | 1.026 | 0.92–1.14 | 0.629 |
| Multivariate | HR | 95% CI | p | HR | 95% CI | p |
| Gender (F vs. M) | 1.019 | 0.56–1.87 | 0.951 | 0.612 | 0.25–1.47 | 0.274 |
| Age (years) | 0.999 | 0.98–1.02 | 0.861 | 1.010 | 0.99–1.03 | 0.387 |
| Breslow (mm) | 1.164 | 1.09–1.25 | <0.001 | 1.173 | 1.06–1.30 | 0.002 |
| Stage (II vs. III–IV) | 0.716 | 0.33–1.53 | 0.390 | 0.596 | 0.17–2.10 | 0.421 |
| TAM-CCL20 (MFI, each 10 a.u.) | 1.244 | 1.12–1.38 | <0.001 | 1.260 | 1.08–1.47 | 0.004 |
| Gender (F vs. M) | 1.245 | 0.71–2.20 | 0.449 | 0.666 | 0.27–1.64 | 0.376 |
| Age (years) | 0.996 | 0.98–1.01 | 0.692 | 1.005 | 0.98–1.03 | 0.715 |
| Breslow (mm) | 1.115 | 1.05–1.18 | <0.001 | 1.123 | 1.03–1.22 | 0.008 |
| Stage (II vs. III–IV) | 1.401 | 0.90–2.19 | 0.138 | 1.013 | 0.35–2.92 | 0.982 |
| TAM-TNF (MFI, each 10 a.u.) | 1.304 | 1.19–1.43 | <0.001 | 1.255 | 1.09–1.44 | 0.001 |
| Gender (F vs. M) | 1.103 | 0.61–2.00 | 0.747 | 0.811 | 0.34–1.91 | 0.632 |
| Age (years) | 1.005 | 0.99–1.02 | 0.577 | 1.008 | 0.98–1.03 | 0.550 |
| Breslow (mm) | 1.138 | 1.06–1.22 | <0.001 | 1.124 | 1.03–1.23 | 0.008 |
| Stage (II vs. III–IV) | 1.122 | 0.63–1.99 | 0.694 | 1.230 | 0.46–3.32 | 0.683 |
| TAM-VEGFA (MFI, each 10 a.u.) | 1.188 | 1.11–1.27 | <0.001 | 1.040 | 0.95–1.14 | 0.404 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Seijo, A.; García-Martínez, E.; Barrio-Alonso, C.; Pareja-Malagón, M.; Acosta-Ocampo, A.; Fernández-Santos, M.E.; Puig-Kröger, A.; Parra-Blanco, V.; Mercader, E.; Márquez-Rodas, I.; et al. CCL20/TNF/VEGFA Cytokine Secretory Phenotype of Tumor-Associated Macrophages Is a Negative Prognostic Factor in Cutaneous Melanoma. Cancers 2021, 13, 3943. https://doi.org/10.3390/cancers13163943
Gutiérrez-Seijo A, García-Martínez E, Barrio-Alonso C, Pareja-Malagón M, Acosta-Ocampo A, Fernández-Santos ME, Puig-Kröger A, Parra-Blanco V, Mercader E, Márquez-Rodas I, et al. CCL20/TNF/VEGFA Cytokine Secretory Phenotype of Tumor-Associated Macrophages Is a Negative Prognostic Factor in Cutaneous Melanoma. Cancers. 2021; 13(16):3943. https://doi.org/10.3390/cancers13163943
Chicago/Turabian StyleGutiérrez-Seijo, Alba, Elena García-Martínez, Celia Barrio-Alonso, Miriam Pareja-Malagón, Alejandra Acosta-Ocampo, María Eugenia Fernández-Santos, Amaya Puig-Kröger, Verónica Parra-Blanco, Enrique Mercader, Iván Márquez-Rodas, and et al. 2021. "CCL20/TNF/VEGFA Cytokine Secretory Phenotype of Tumor-Associated Macrophages Is a Negative Prognostic Factor in Cutaneous Melanoma" Cancers 13, no. 16: 3943. https://doi.org/10.3390/cancers13163943
APA StyleGutiérrez-Seijo, A., García-Martínez, E., Barrio-Alonso, C., Pareja-Malagón, M., Acosta-Ocampo, A., Fernández-Santos, M. E., Puig-Kröger, A., Parra-Blanco, V., Mercader, E., Márquez-Rodas, I., Avilés-Izquierdo, J. A., Samaniego, R., & Sánchez-Mateos, P. (2021). CCL20/TNF/VEGFA Cytokine Secretory Phenotype of Tumor-Associated Macrophages Is a Negative Prognostic Factor in Cutaneous Melanoma. Cancers, 13(16), 3943. https://doi.org/10.3390/cancers13163943