
Germline ERBB2/HER2 Coding Variants Are Associated with Increased Risk of Myeloproliferative Neoplasms


 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Recruitment
2.2. Whole Exome Sequencing and Variant Analysis
2.3. Variant Confirmation
2.4. Targeted Sequencing
2.5. Clinical Sequencing Analysis
2.6. Statistical Analysis
3. Results
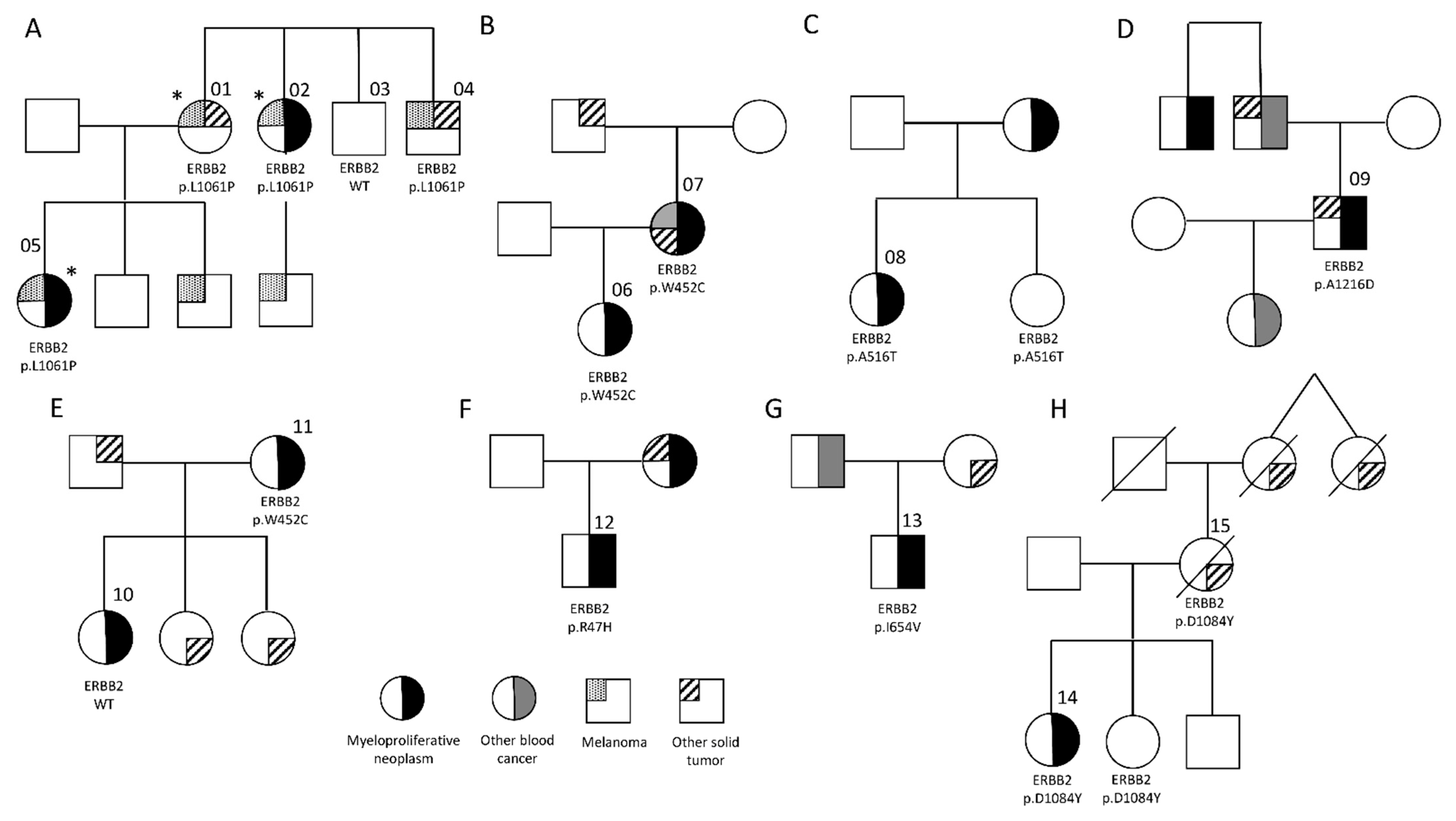
3.1. Germline ERBB2 Variants in Familial MPN
3.2. Frequency of ERBB2 Variants in Hematologic Malignancies
3.3. ERBB2 Variants Overrepresented in MPN
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sud, A.; Chattopadhyay, S.; Thomsen, H.; Sundquist, K.; Sundquist, J.; Houlston, R.S.; Hemminki, K. Familial risks of acute myeloid leukemia, myelodysplastic syndromes, and myeloproliferative neoplasms. Blood 2018, 132, 973–976. [Google Scholar] [CrossRef]
- Landgren, O.; Goldin, L.R.; Kristinsson, S.Y.; Helgadottir, E.A.; Samuelsson, J.; Björkholm, M. Increased risks of polycythemia vera, essential thrombocythemia, and myelofibrosis among 24 577 first-degree relatives of 11 039 patients with myeloproliferative neoplasms in Sweden. Blood 2008, 112, 2199–2204. [Google Scholar] [CrossRef]
- Rumi, E.; Passamonti, F.; Della Porta, M.G.; Elena, C.; Arcaini, L.; Vanelli, L.; Del Curto, C.; Pietra, D.; Boveri, E.; Pascutto, C.; et al. Familial Chronic Myeloproliferative Disorders: Clinical Phenotype and Evidence of Disease Anticipation. J. Clin. Oncol. 2007, 25, 5630–5635. [Google Scholar] [CrossRef] [PubMed]
- Harutyunyan, A.S.; Giambruno, R.; Krendl, C.; Stukalov, A.; Klampfl, T.; Berg, T.; Chen, R.; Feenstra, J.D.M.; Jäger, R.; Gisslinger, B.; et al. Germline RBBP6 mutations in familial myeloproliferative neoplasms. Blood 2016, 127, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Babushok, D.V.; Stanley, N.L.; Morrissette, J.J.D.; Lieberman, D.B.; Olson, T.S.; Chou, S.T.; Hexner, E.O. Germline duplication of ATG2B and GSKIP genes is not required for the familial myeloid malignancy syndrome associated with the duplication of chromosome 14q32. Leukemia 2018, 32, 2720–2723. [Google Scholar] [CrossRef] [PubMed]
- Saliba, J.; Saint-Martin, C.; Di Stefano, A.; Lenglet, G.; Marty, C.; Keren, B.; Pasquier, F.; Della Valle, V.; Secardin, L.; Leroy, G.; et al. Germline duplication of ATG2B and GSKIP predisposes to familial myeloid malignancies. Nat. Genet. 2015, 47, 1131–1140. [Google Scholar] [CrossRef]
- Quesada, A.E.; Routbort, M.J.; Dinardo, C.D.; Bueso-Ramos, C.E.; Kanagal-Shamanna, R.; Khoury, J.D.; Thakral, B.; Zuo, Z.; Yin, C.C.; Loghavi, S.; et al. DDX41 mutations in myeloid neoplasms are associated with male gender, TP53 mutations and high-risk disease. Am. J. Hematol. 2019, 94, 757–766. [Google Scholar] [CrossRef]
- Jones, A.V.; Chase, A.; Silver, R.T.; Oscier, D.; Zoi, K.; Wang, Y.L.; Cario, H.; Pahl, H.L.; Collins, A.; Reiter, A.; et al. JAK2 haplotype is a major risk factor for the development of myeloproliferative neoplasms. Nat. Genet. 2009, 41, 446–449. [Google Scholar] [CrossRef]
- Kilpivaara, O.; Mukherjee, S.; Schram, A.; Wadleigh, M.; Mullally, A.; Ebert, B.L.; Bass, A.; Marubayashi, S.; Heguy, A.; Garcia-Manero, G.; et al. A germline JAK2 SNP is associated with predisposition to the development of JAK2V617F-positive myeloproliferative neoplasms. Nat. Genet. 2009, 41, 455–459. [Google Scholar] [CrossRef]
- Olcaydu, D.; Harutyunyan, A.; Jäger, R.; Berg, T.; Gisslinger, B.; Pabinger, I.; Gisslinger, H.; Kralovics, R. A common JAK2 haplotype confers susceptibility to myeloproliferative neoplasms. Nat. Genet. 2009, 41, 450–454. [Google Scholar] [CrossRef]
- Olcaydu, D.; Rumi, E.; Harutyunyan, A.; Passamonti, F.; Pietra, D.; Pascutto, C.; Berg, T.; Jäger, R.; Hammond, E.; Cazzola, M.; et al. The role of the JAK2 GGCC haplotype and the TET2 gene in familial myeloproliferative neoplasms. Haematologica 2010, 96, 367–374. [Google Scholar] [CrossRef]
- Hinds, D.A.; Barnholt, K.E.; Mesa, R.A.; Kiefer, A.K.; Do, C.B.; Eriksson, N.; Mountain, J.L.; Francke, U.; Tung, J.Y.; Nguyen, H.M.; et al. Germ line variants predispose to both JAK2 V617F clonal hematopoiesis and myeloproliferative neoplasms. Blood 2016, 128, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Oddsson, A.; Kristinsson, S.Y.; Helgason, H.; Gudbjartsson, D.F.; Masson, G.; Sigurdsson, A.; Jonasdottir, A.; Steingrimsdottir, H.; Vidarsson, B.; Reykdal, S.; et al. The germline sequence variant rs2736100_C in TERT associates with myeloproliferative neoplasms. Leukemia 2014, 28, 1371–1374. [Google Scholar] [CrossRef] [PubMed]
- Tapper, W.; Jones, A.V.; Kralovics, R.; Harutyunyan, A.; Zoi, K.; Leung, W.; Godfrey, A.L.; Guglielmelli, P.; Callaway, A.; Ward, D.; et al. Genetic variation at MECOM, TERT, JAK2 and HBS1L-MYB predisposes to myeloproliferative neoplasms. Nat. Commun. 2015, 6, 6691. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yin, Z.; Wu, W.; Li, X.; Zhou, B. Genetic variants in TERT–CLPTM1L genetic region associated with several types of cancer: A meta-analysis. Gene 2013, 526, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Krahling, T.; Balassa, K.; Kiss, K.P.; Bors, A.; Batai, A.; Halm, G.; Egyed, M.; Fekete, S.; Remenyi, P.; Masszi, T.; et al. Co-occurrence of Myeloproliferative Neoplasms and Solid Tumors Is Attributed to a Synergism Between Cytoreductive Therapy and the Common TERT Polymorphism rs2736100. Cancer Epidemiol. Biomark. Prev. 2016, 25, 98–104. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bao, E.L.; Nandakumar, S.K.; Liao, X.; Bick, A.G.; Karjalainen, J.; Tabaka, M.; Gan, O.I.; Havulinna, A.S.; Kiiskinen, T.T.J.; Lareau, C.A.; et al. Inherited Myeloproliferative Neoplasm Risk Affects Haematopoietic Stem Cells. Nature 2020, 586, 769–775. [Google Scholar] [CrossRef]
- Moasser, M.M.; Krop, I.E. The Evolving Landscape of HER2 Targeting in Breast Cancer. JAMA Oncol. 2015, 1, 1154–1161. [Google Scholar] [CrossRef]
- Cousin, S.; Khalifa, E.; Crombe, A.; Laizet, Y.; Lucchesi, C.; Toulmonde, M.; Le Moulec, S.; Auzanneau, C.; Soubeyran, I.; Italiano, A. Targeting ERBB2 mutations in solid tumors: Biological and clinical implications. J. Hematol. Oncol. 2018, 11, 86. [Google Scholar] [CrossRef]
- Li, R.; Sobreira, N.; Witmer, P.D.; Pratz, K.W.; Braunstein, E.M. Two novel germline DDX41 mutations in a family with inherited myelodysplasia/acute myeloid leukemia. Haematologica 2016, 101, e228–e231. [Google Scholar] [CrossRef]
- Sobreira, N.; Schiettecatte, F.; Boehm, C.; Valle, D.; Hamosh, A. New Tools for Mendelian Disease Gene Identification: PhenoDB Variant Analysis Module; and GeneMatcher, a Web-Based Tool for Linking Investigators with an Interest in the Same Gene. Hum. Mutat. 2015, 36, 425–431. [Google Scholar] [CrossRef] [PubMed]
- 1000 Genomes Project Consortium; Abecasis, G.R.; Auton, A.; Brooks, L.D.; DePristo, M.A.; Durbin, R.M.; Handsaker, R.E.; Kang, H.M.; Marth, G.T.; McVean, G.A. An Integrated Map of Genetic Variation from 1,092 Human Genomes. Nature 2012, 491, 56–65. [Google Scholar] [PubMed]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. Variation across 141,456 human exomes and genomes reveals the spectrum of loss-of-function intolerance across human protein-coding genes. bioRxiv 2019, 531210. [Google Scholar] [CrossRef]
- Karantanos, T.; Chaturvedi, S.; Braunstein, E.M.; Spivak, J.; Resar, L.; Karanika, S.; Williams, D.M.; Rogers, O.; Gocke, C.D.; Moliterno, A.R. Sex determines the presentation and outcomes in MPN and is related to sex-specific differences in the mutational burden. Blood Adv. 2020, 4, 2567–2576. [Google Scholar] [CrossRef]
- Zheng, G.; Chen, P.; Pallavajjalla, A.; Haley, L.; Gondek, L.; DeZern, A.; Ling, H.; De Marchi, F.; Lin, M.; Gocke, C. The diagnostic utility of targeted gene panel sequencing in discriminating etiologies of cytopenia. Am. J. Hematol. 2019, 94, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Frank, B.; Hemminki, K.; Wirtenberger, M.; Bermejo, J.; Bugert, P.; Klaes, R.; Schmutzler, R.; Wappenschmidt, B.; Bartram, C.; Burwinkel, B. The rare ERBB2 variant Ile654Val is associated with an increased familial breast cancer risk. Carcinogenesis 2005, 26, 643–647. [Google Scholar] [CrossRef][Green Version]
- Appenzeller, S.; Gesierich, A.; Thiem, A.; Hufnagel, A.; Jessen, C.; Kneitz, H.; Regensburger, M.; Schmidt, C.; Zirkenbach, V.; Bischler, T.; et al. The identification of patient-specific mutations reveals dual pathway activation in most patients with melanoma and activated receptor tyrosine kinases in BRAF/NRAS wild-type melanomas. Cancer 2018, 125, 586–600. [Google Scholar] [CrossRef]
- Yamamoto, H.; Higasa, K.; Sakaguchi, M.; Shien, K.; Soh, J.; Ichimura, K.; Furukawa, M.; Hashida, S.; Tsukuda, K.; Takigawa, N.; et al. Novel Germline Mutation in the Transmembrane Domain of HER2 in Familial Lung Adenocarcinomas. J. Natl. Cancer Inst. 2014, 106, djt338. [Google Scholar] [CrossRef]
- Pahuja, K.B.; Nguyen, T.T.; Jaiswal, B.S.; Prabhash, K.; Thaker, T.M.; Senger, K.; Chaudhuri, S.; Kljavin, N.M.; Antony, A.; Phalke, S.; et al. Actionable Activating Oncogenic ERBB2/HER2 Transmembrane and Juxtamembrane Domain Mutations. Cancer Cell 2018, 34, 792–806.e5. [Google Scholar] [CrossRef]
- Braunstein, E.M.; Li, R.; Sobreira, N.; Marosy, B.; Hetrick, K.; Doheny, K.; Gocke, C.D.; Valle, D.; Brodsky, R.A.; Cheng, L. A germline ERBB3 variant is a candidate for predisposition to erythroid MDS/erythroleukemia. Leukemia 2016, 30, 2242–2245. [Google Scholar] [CrossRef]
- Grossman, R.L.; Heath, A.P.; Ferretti, V.; Varmus, H.E.; Lowy, D.R.; Kibbe, W.A.; Staudt, L.M. Toward a Shared Vision for Cancer Genomic Data. N. Engl. J. Med. 2016, 375, 1109–1112. [Google Scholar] [CrossRef]
- Adzhubei, I.; Jordan, D.; Sunyaev, S.R. Predicting Functional Effect of Human Missense Mutations Using PolyPhen-2. Curr. Protoc. Hum. Genet. 2013, 76, 7.20.1–7.20.41. [Google Scholar] [CrossRef] [PubMed]
- Furutani, E.; Shimamura, A. Germline Genetic Predisposition to Hematologic Malignancy. J. Clin. Oncol. 2017, 35, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Foulkes, W.D. Inherited Susceptibility to Common Cancers. N. Engl. J. Med. 2008, 359, 2143–2153. [Google Scholar] [CrossRef]
- Pharoah, P.D.; Antoniou, A.C.; Bobrow, M.; Zimmern, R.L.; Easton, D.F.; Ponder, B.A. Polygenic susceptibility to breast cancer and implications for prevention. Nat. Genet. 2002, 31, 33–36. [Google Scholar] [CrossRef]
- Bodmer, W.; Bonilla, C. Common and rare variants in multifactorial susceptibility to common diseases. Nat. Genet. 2008, 40, 695–701. [Google Scholar] [CrossRef]
- Fletcher, O.; Houlston, R. Architecture of inherited susceptibility to common cancer. Nat. Rev. Cancer 2010, 10, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Arteaga, C.L.; Engelman, J.A. ERBB Receptors: From Oncogene Discovery to Basic Science to Mechanism-Based Cancer Therapeutics. Cancer Cell 2014, 25, 282–303. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.K.; Keck, J.M.; Eide, C.A.; Bottomly, D.; Traer, E.; Tyner, J.W.; McWeeney, S.K.; Tognon, C.E.; Druker, B.J. ERBB2/HER2 mutations are transforming and therapeutically targetable in leukemia. Leukemia 2020, 34, 2798–2804. [Google Scholar] [CrossRef] [PubMed]
- Network, Cancer Genome Atlas Research. Genomic and Epigenomic Landscapes of Adult De Novo Acute Myeloid Leukemia. N. Engl. J. Med. 2013, 368, 2059–2074. [Google Scholar] [CrossRef]
- Zabransky, D.J.; Yankaskas, C.; Cochran, R.L.; Wong, H.Y.; Croessmann, S.; Chu, D.; Kavuri, S.M.; Brewer, M.R.; Rosen, D.M.; Dalton, W.; et al. HER2 missense mutations have distinct effects on oncogenic signaling and migration. Proc. Natl. Acad. Sci. USA 2015, 112, E6205–E6214. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.S.; Gay, L.M.; Wang, K.; Ali, S.M.; Chumsri, S.; Elvin, J.A.; Bose, R.; Vergilio, J.-A.; Suh, J.; Yelensky, R.; et al. NonamplificationERBB2genomic alterations in 5605 cases of recurrent and metastatic breast cancer: An emerging opportunity for anti-HER2 targeted therapies. Cancer 2016, 122, 2654–2662. [Google Scholar] [CrossRef] [PubMed]
- Aloraifi, F.; McDevitt, T.; Martiniano, R.; McGreevy, J.; McLaughlin, R.; Egan, C.M.; Cody, N.; Meany, M.; Kenny, E.; Green, A.J.; et al. Detection of Novel Germline Mutations for Breast Cancer in Non-BRCA1/2 Families. FEBS J. 2015, 282, 3424–3437. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Walsh, M.F.; Wu, G.; Edmonson, M.N.; Gruber, T.A.; Easton, J.; Hedges, D.; Aman, P.; Zhou, X.; Yergeau, D.A.; et al. Germline Mutations in Predisposition Genes in Pediatric Cancer. N. Engl. J. Med. 2015, 373, 2336–2346. [Google Scholar] [CrossRef]
- Macauda, A.; Giaccherini, M.; Sainz, J.; Gemignani, F.; Sgherza, N.; Sánchez-Maldonado, J.M.; Gora-Tybor, J.; Martinez-Lopez, J.; Carreño-Tarragona, G.; Jerez, A.; et al. Do myeloproliferative neoplasms and multiple myeloma share the same genetic susceptibility loci? Int. J. Cancer 2021, 148, 1616–1624. [Google Scholar] [CrossRef]
- Bick, A.G.; NHLBI Trans-Omics for Precision Medicine Consortium; Weinstock, J.S.; Nandakumar, S.K.; Fulco, C.P.; Bao, E.L.; Zekavat, S.M.; Szeto, M.D.; Liao, X.; Leventhal, M.J.; et al. Inherited causes of clonal haematopoiesis in 97,691 whole genomes. Nat. Cell Biol. 2020, 586, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.M.; Çolak, Y.; Ellervik, C.; Hasselbalch, H.C.; Bojesen, S.E.; Nordestgaard, B.G. Loss-of-function polymorphism in IL6R reduces risk of JAK2V617F somatic mutation and myeloproliferative neoplasm: A Mendelian randomization study. EClinicalMedicine 2020, 21, 100280. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Diagnosis | N Cases | ERBB2 MAF < 0.01 | ERBB2 MAF < 0.005 |
| MPN | 236 | 21 (8.9%) | 11 (4.7%) |
| CMML | 92 | 6 (6.5%) | 2 (2.2%) |
| MDS/AML | 771 | 30 (3.9%) | 9 (1.2%) |
| AA | 61 | 3 (4.9%) | 2 (3.3%) |
| CML | 27 | 1 (3.7%) | 0 |
| Lymphoid | 161 | 6 (3.7%) | 1 (0.06) |
| Control (Non-Blood Cancer) | 256 | 7 (2.7%) | 3 (1.2%) |
| All Blood Cancer | 1348 | 67 (5.0%) | 26 (1.9%) |
| Non-MPN Blood Cancer | 1112 | 46 (4.1%) | 14 (1.3%) |
| Validation Control Cohort | 1587 | 83 (5.2%) | 47 (3.0%) |
| Comparison | Odds ratio (95% confidence interval), p value | ||
| MPN vs. Control | 3.5 (1.4 to 8.3), p = 0.0053 | 4.1 (1.1 to 13.5), p = 0.0313 | |
| MPN vs. MDS/AML | 2.4 (1.4 to 4.0), p = 0.0028 | 4.1 (1.7 to 10.1) p = 0.0018 | |
| MPN vs. Non-MPN Blood Cancer | 2.3 (1.3 to 3.9) p = 0.0028 | 3.8 (1.7 to 8.6) p = 0.0010 | |
| MPN vs. Validation Control | 1.7 (1.0 to 2.8) p = 0.0400 | 1.6 (0.8 to 3.1) p = 0.1698 | |
| MPN Cohort (n = 292) | Non-MPN Cancer Cohort (n = 1112) | Validation Control Cohort (n = 1587) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Variant | Observed (%) | Expected # | O/E Ratio | Observed (%) | Expected # | O/E Ratio | Observed (%) | Expected # | O/E Ratio |
| I654V | 6 (2.05) | 2.81 | 2.13 | 17 (1.53) | 10.72 | 1.59 | 17 (1.07) | 15.29 | 1.11 |
| A386D | 2 (0.68) | 2.12 | 0.94 | 8 (0.72) | 8.09 | 0.99 | 13 (0.82) | 11.55 | 1.13 |
| E930D | 2 (0.68) | 0.30 | 6.67 | 2 (0.18) | 1.14 | 1.75 | 1 (0.06) | 1.63 | 0.61 |
| P489L | 1 (0.34) | 0.42 | 2.36 | 3 (0.27) | 1.61 | 1.86 | 2 (0.13) | 2.30 | 0.87 |
| R1161Q | 0 | 0.44 | NA | 3 (0.27) | 1.69 | 1.78 | 2 (0.13) | 2.41 | 0.83 |
| R143Q | 0 | 0.28 | NA | 2 (0.18) | 1.10 | 1.89 | 0 | 1.51 | NA |
| I961T | 0 | 0.004 | NA | 2 (0.18) | 0.02 | 113 | 0 | 0.03 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braunstein, E.M.; Chen, H.; Juarez, F.; Yang, F.; Tao, L.; Makhlin, I.; Williams, D.M.; Chaturvedi, S.; Pallavajjala, A.; Karantanos, T.; et al. Germline ERBB2/HER2 Coding Variants Are Associated with Increased Risk of Myeloproliferative Neoplasms. Cancers 2021, 13, 3246. https://doi.org/10.3390/cancers13133246
Braunstein EM, Chen H, Juarez F, Yang F, Tao L, Makhlin I, Williams DM, Chaturvedi S, Pallavajjala A, Karantanos T, et al. Germline ERBB2/HER2 Coding Variants Are Associated with Increased Risk of Myeloproliferative Neoplasms. Cancers. 2021; 13(13):3246. https://doi.org/10.3390/cancers13133246
Chicago/Turabian StyleBraunstein, Evan M., Hang Chen, Felicia Juarez, Fanghan Yang, Lindsay Tao, Igor Makhlin, Donna M. Williams, Shruti Chaturvedi, Aparna Pallavajjala, Theodoros Karantanos, and et al. 2021. "Germline ERBB2/HER2 Coding Variants Are Associated with Increased Risk of Myeloproliferative Neoplasms" Cancers 13, no. 13: 3246. https://doi.org/10.3390/cancers13133246
APA StyleBraunstein, E. M., Chen, H., Juarez, F., Yang, F., Tao, L., Makhlin, I., Williams, D. M., Chaturvedi, S., Pallavajjala, A., Karantanos, T., Martin, R., Wohler, E., Sobreira, N., Gocke, C. D., & Moliterno, A. R. (2021). Germline ERBB2/HER2 Coding Variants Are Associated with Increased Risk of Myeloproliferative Neoplasms. Cancers, 13(13), 3246. https://doi.org/10.3390/cancers13133246