Friend or Foe: S100 Proteins in Cancer

 ,
,
Abstract
1. The S100 Family
1.1. The S100 Family Members
1.1.1. Structure
1.1.2. Expression and Regulation
1.1.3. Functions
1.2. Clinical Relevance of S100 Proteins
2. S100 Proteins in Cancer
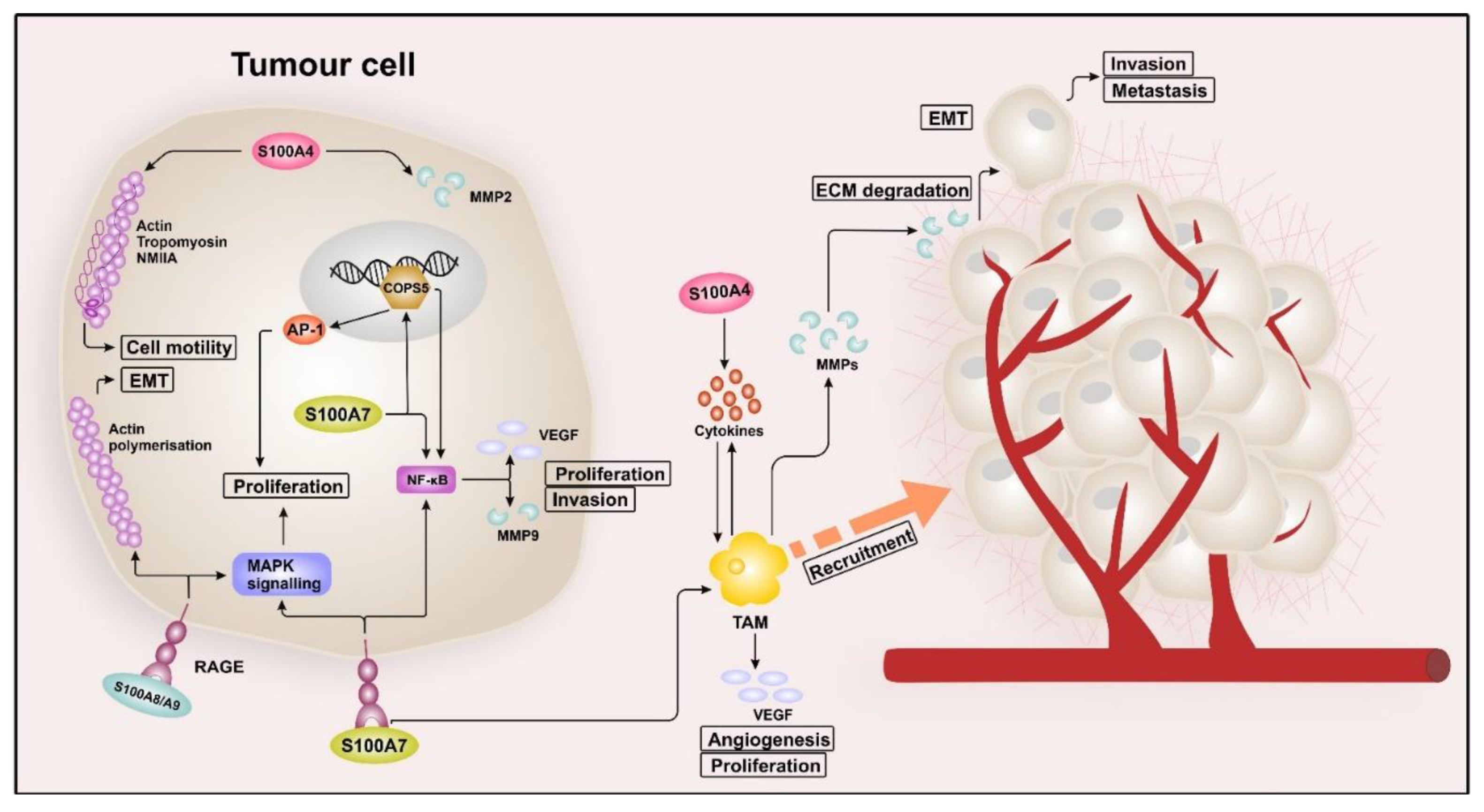
2.1. S100 Proteins in Breast Cancer
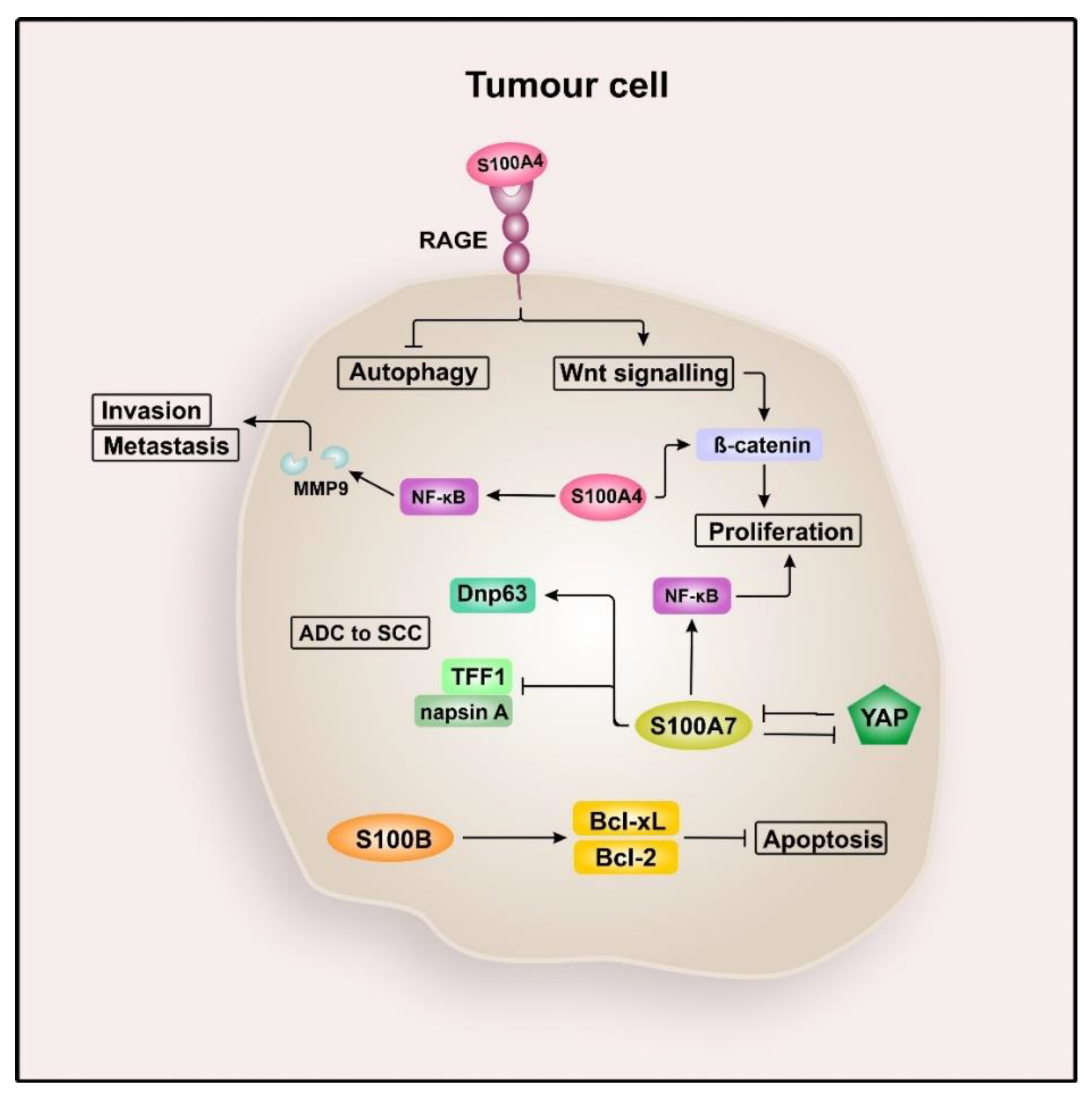
2.2. S100 Proteins in Lung Cancer
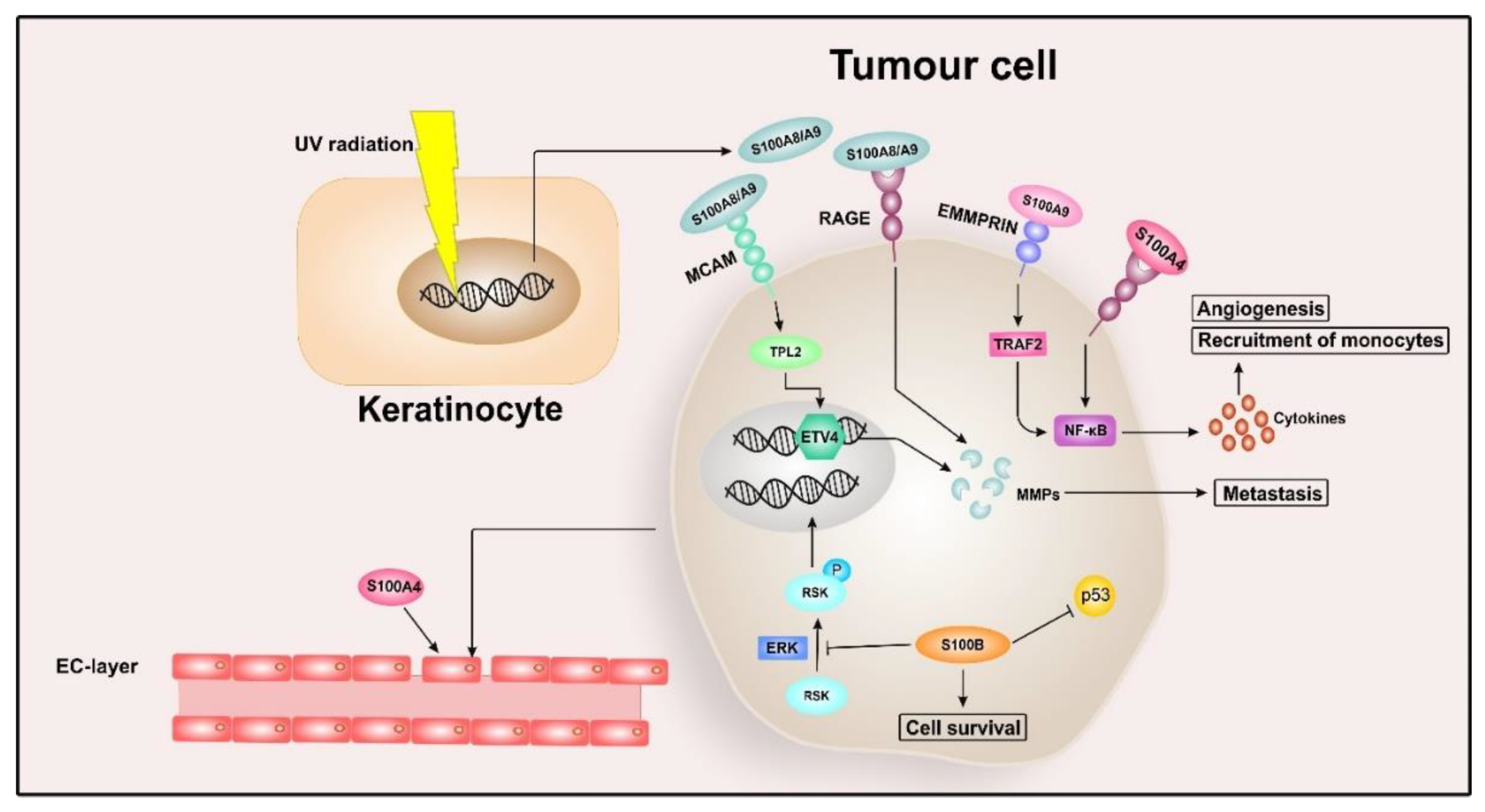
2.3. S100 Proteins in Malignant Melanoma
2.4. S100 Proteins in Ovarian Cancer
2.5. S100 Proteins in Colorectal Cancer
2.6. S100 Proteins in Pancreatic Cancer
2.7. S100 Proteins in Other Cancer Types
3. Targeting S100 Proteins for Cancer Therapy
3.1. Small Molecule Inhibitors
3.2. Neutralising Antibodies
3.3. Micro RNA (miRNA) Mimics
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| ADC | adenocarcinoma |
| AMP | antimicrobial peptide |
| ANT | adenine nucleotide translocator |
| ATP | adenosine triphosphate |
| Bcl-2 | namely B-cell lymphoma 2 |
| Bcl-xL | B-cell lymphoma extra-large |
| Ca2+ | calcium ion |
| Cacy/SIP | calcyclin-binding protein/Siah-1-interacting protein |
| CCL2 | chemokine (C-C motif) ligand 2 |
| COP9 | constitutive photomorphogenesis 9 |
| COPS5 | COP9 constitutive photomorphogenic homolog subunit 5 |
| COX2 | cyclooxygenase-2 |
| CP | calprotectin |
| CRC | colorectal carcinoma |
| CT | computerised tomography |
| Cu2+ | copper ion |
| CXCL | chemokine (C-X-C motif) ligand |
| DAMPs | danger-associated molecular patterns DAMPs |
| ECG | electrocardiogram |
| ECM | extracellular matrix |
| EDC | epidermal differentiation complex |
| EGF | epidermal growth factor |
| EMMPRIN | extracellular matrix metalloprotease inducer |
| EMT | epithelial-mesenchymal transition |
| ERK | extracellular signal-regulated kinase |
| ERα | in estrogen receptor α |
| FABP | fatty acid-binding proteins |
| FAK | focal adhesion kinase |
| FDA | Food and Drug Administration |
| Fe2+ | iron ion |
| HCC | hepatocellular carcinoma |
| HIF-1 | hypoxia-inducible factor 1 |
| IBD | inflammatory bowel disease |
| ICI | intracranial injuries |
| IL-1β | interleukin-1β |
| IFN-γ | interferon gamma |
| Jab1 | c-Jun activation domain-binding protein-1 |
| MAPK | mitogen-activated protein kinase |
| MAP3K8 | mitogen-activated protein kinase kinase kinase 8 |
| MCAM | melanoma cell adhesion molecule |
| MDR1 | multidrug resistance gene 1 |
| Mef2 | myocyte enhancer factor-2 |
| MRP1 | MDR-associated protein 1 |
| miRNA | micro RNA |
| MMP | metalloproteinase |
| Mn2+ | manganese ion |
| mTOR | mechanistic target of rapamycin |
| NCX1 | Na+/Ca2+ exchanger |
| Ndr | nuclear dbf2-related |
| NF-kB | nuclear factor ’kappa-light-chain-enhancer’ of activated B-cells |
| NMIBC | non-muscle-invasive bladder cancer |
| NMIIA | non-muscle myosin heavy chain IIA |
| NOXA | phorbol-12-myristate-13-acetate-induced protein 1 |
| NSCLC | non-small cell lung cancer |
| OS | overall survival |
| OCSC | ovarian cancer stem cells |
| PAD3 | phytoalexin deficient 3 |
| PDGF | platelet-derived growth factor |
| PI3K | phosphoinositide 3-kinase |
| PMCA1b | plasma membrane Ca2+ transport ATPase |
| PUMA | p53 upregulated modulator of apoptosis |
| RA | rheumatoid arthritis |
| RAGE | receptor for advanced glycation end products |
| ROS | reactive oxygen species |
| RSK | ribosomal S6 kinase |
| RyR2 | ryanodine receptor 2 |
| SCC | squamous-cell carcinoma |
| SCLC | small cell lung cancer |
| SERCA2a | sarcoplasmic reticulum ATPase |
| Shh | sonic hedgehog |
| SNC | Scandinavian Neurotrauma Committee |
| STEMI | ST-segment elevation myocardial infarction |
| TAMs | tumour-associated macrophages |
| TBA | traumatic brain injury |
| TCF | T-cell factor |
| TLR4 | Toll-like receptor 4 |
| TME | tumour microenvironment |
| TNFα | tumour necrosis factor α |
| tPA | tissue plasminogen activator |
| TPL2 | tumour progression locus 2 |
| TRAF | TNF receptor-associated factor 2 |
| TRAIL | tumour necrosis factor-related apoptosis-inducing ligand |
| TRPV6 | transient receptor potential vanilloid 6 |
| VEGF | vascular endothelial growth factor |
| YAP | yes-associated protein |
| Zn2+ | zinc ion |
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Family Member | Expression | Regulation | Targets/Interaction | Function | References |
|---|---|---|---|---|---|
| S100A1 | Cardiomyocytes, skeletal muscle cells, endothelial cells, neurological cells | Transcription factor binding sites for Nkx 2.5, Mef2 and CEF | SERCA2a/PLB, RyR2, F1-ATPase, ANT, Titin | Modulates contractile performance in cardiomyocytes: interaction with SERCA2a/PLB and RyR2 improves systolic Ca2+ release and diastolic Ca2+ uptake; binding to the PEVK subdomain of titin results in improved sarcomeric compliance Mitochondrial metabolism: plays a role in cardiac energy homeostasis by interacting with F1-ATPase and ANT | [36,37,256] |
| S100A2 (S100L; CaN19) | Epithelial tissue | Transcriptional activation by p53 EGF-induced transcription in keratinocytes | p53, RAGE | Tumour suppressor: calcium-dependent activation of p53 | [51,78] |
| S100A3 (S100E) | Hair cuticular cells | PAD3 induces the formation of a homotetramer via citrullination | RARα | Maintenance of the hair texture: Ca2+-dependent epithelial barrier formation and cuticular maturation | [22,257,258] |
| S100A4 (metastasin; CAPL; calvasculin) | Fibroblasts, immune cells, cancer cells | Transcription is promoted by β-catenin and growth factors | actin, NMIIA, tropomyosin, p53, p37, MMP2, liprin-β1, RAGE, TLR4, EGFR, IL-10 receptor, annexin II | Cell migration: interaction with proteins of the cytoskeleton and induces secretion of matrix metalloproteinases (MMPs) Cell growth and proliferation: activates NF-κB and MAP kinases | [45,52,89] |
| S100A5 | olfactory bulb, brainstem, spinal trigeminal tract | - | RAGE, NCXI | - | [34,259] |
| S100A6 (Calcylin; Cacy; CABP; PRA) | Fibroblasts, epithelial cells | Extracellular by PDGF, EGF, retonic acid, estrogen, gastrin Under stress conditions: ischemia, irradiation, oxidative stress Intracellular: NF-κB activates S100A6-promotor, while p53 indirectly suppresses transcription | caldesmon, calponin, tropomyosin, kinesin light chain, RAGE | Control of cell cycle progression: involved in ubiquitination of β-catenin Regulation of microfilament dynamics: interaction with proteins of the cytoskeleton Neuronal apoptosis: binding to RAGE leads to ROS-dependent activation of JNK, caspase-3, and caspase-7 As extracellular factor: involved in the release of lactogen II, insulin, and histamine | [27,260] |
| S100A7 (Psoriasin) | Keratinocytes | Secretion induced by disruptive stimuli (e.g., UVB, irradiation, retonic acid, infection with bacteria) Upregulated by pro-inflammatory cytokines (e.g., IL-1α, TNFα, IL-17, IL-22) | RAGE, FABP, TGM | Role as AMP: lyses bacteria (e.g., E. coli) Inhibition of epidermal differentiation: RAGE-dependent activation of NF-κB-signaling results in upregulation of IL-6 Inflammation: selective chemotactic for CD4+ lymphocytes and neutrophils | [28,42] |
| S100A8 (Calgranulin A) | Neutrophils, osteoclasts, hypertrophic chondrocytes, myeloid dendric cells | Induced by LPS, IL-10, TLR4, glucocorticoids | Telomerase | Inflammation: chemotactic for neutrophils, protects from oxidation by scavenging oxidants, acts as NO shuttle, thereby activating mast cells Regulation of differentiation: binds and inhibits telomerase, therefore promoting differentiation in keratinocytes | [43,261] |
| S100A9 (Calgranulin B) | Neutrophil granulocytes, monocytes | Upregulated by glucocorticoids, cytokines, and growth factors | RAGE, TLR4 | Inflammation: Zn2+-dependent interaction with RAGE and TLR4 leads to upregulation of pro-inflammatory cytokines (e.g., TNFα) via NF-κB signalling; acts as a chemoattractant and is involved in differentiation of myeloid cells | [21,40,43] |
| S100A8/S100A9 (Calprotectin) | Neutrophils, monocytes, macrophages, inducible expression in keratinocytes and epithelial cells (e.g., under inflammatory conditions) | TNFα, IL-1β, HIF-1 stimulate expression Secretion is Ca2+-dependent | RAGE, TLR4 | Modulation of the cytoskeleton: interacts with several proteins of the cytoskeleton (e.g., keratin and F-actin) and promotes polymerisation of microtubules Protection against pathogens: S100A8/S100A9 expressing epithelial cells are more resistant to bacterial infection Inflammation: RAGE- and TLR4-mediated upregulation of pro-inflammatory cytokines (e.g., IL-6 and IL-8) and adhesion proteins (e.g., ICAM-1) | [29] |
| S100A10 (p11) | Endothelial cells, macrophages, fibroblasts, epithelial cells | The expression can be induced by: Sp1, IFN-γ, glucocorticoids, TGFβ, EGF, IL-1β, thrombin, oncogenes (e.g., PML-RARα, KRas) | Annexin II, 5-HT1B receptor, TRPV5, TRPV6, TASK-1 | Role as plasminogen receptor: regulates plasmin production by forming a complex with annexin II, which leads to fibrinolysis, activation of MMPs, ECM degradation, stimulation of the JAK1/TYK2 signalling pathway, and recruitment of macrophages Trafficking of plasma membrane proteins: S100A10 interacts with the 5-HT1B receptor, TRPV5, TRPV6, and TASK-1 and participates in their trafficking | [13,35] |
| S100A11 (S100C; calgizzarin) | Chondrocytes, keratinocytes, fibroblasts, luteal cells | TGFβ and high extracellular Ca2+ concentrations stimulate expression and activity | RAGE, annexin I, nucleolin | Cell growth regulation: intracellular S100A11 inhibits cell growth by binding to nucleolin, which allows expression of p21, leading to inhibition of DNA synthesis; extracellular S100A11 promotes cell proliferation by RAGE-mediated transcription of EGF Interaction with RAGE: RAGE activates p38 MAPK kinase, which increases type X collagen | [49,50] |
| S100A12 (Calgranulin C; EN-RAGE) | Neutrophil granulocytes, monocytes, macrophages, early-stage differentiating epithelial and dendric cells | Upregulated by TNFα and LPS Secretion in neutrophils involves ROS and K+ exchange | RAGE, TLR4, CacyBP/SIP | Wnt signalling: binds to CacyBP/SIP and is part of the ubiquitinylation complex Inflammation: binding to TLR4 leads to activation and migration of monocytes and release of IL-1β, IL-6, and IL-8, RAGE-mediated NF-κB and MAPK signalling induces secretion of TNFα and IL-1β Inhibition of pathogens: antifungal and antibacterial activity via Zn2+ sequestration | [41,262,263,264] |
| S100A13 | Leydig cells of testis, follicle cells of thyroid, smooth muscle, endothelial and epithelial cells | Induced by stress stimuli | RAGE, Syt1, SPHK1 | Non-canonical secretion pathway: forms a copper-dependent multiprotein complex with Syt1 and SPHK and is thereby involved in the non-classical stress-dependent release of FGF-1, IL-1α, and prothymosin-α | [265,266,267] |
| S100A14 | High expression in epithelial tissue, lower expression in mesenchymal tissue | Regulated by p53 | RAGE, p53 | Interaction with RAGE: binding in lower concentrations activates MAPK and NF-κB signalling, leading to cell proliferation; binding in high concentrations activates ROS production, resulting in apoptosis | [46,53] |
| S100A15 (S100A7A; koebnerisin) | Dendric cells, endothelial cells, vascular smooth muscle cells, peripheral nerves, keratinocytes | Upregulated by E. coli through TLR4 Transcription can be induced by IFN-γ, IL-1β, TNFα, and Th1 Co-expression with S100A7 in keratinocytes | GPCR | Epidermal cell maturation: upregulated in epidermal differentiation Initial immune response: functions as an antibacterial agent by reducing the survival of E. coli and other strains Inflammation: acts as a chemoattractant for leucocytes via GPCR | [23,268] |
| S100A16 | Astrocytes, adipocytes | Ca2+ influences nuclear import/export | p53 | Overexpression in preadipocytes resulted in increased proliferation and reduction in insulin-stimulated glucose uptake and Akt phosphorylation | [269,270] |
| S100B | Astrocytes, oligodendrocytes, Schwann cells, ependymal cells, melanocytes, adipocytes, chondrocytes | Secretion is regulated by IL-1β, extracellular Ca2+ and K+, inhibitors of gap junctions, antioxidants, lipopolysaccharide, and apomorphine p53 upregulates expression | Extracellular: RAGE, FGFR1 Intracellular: Ndr kinase, Src, Rac1, IQGAP1, p53 | Cell migration: regulates F-actin-based cytoskeleton via Src, IQGAP1, and Rac1 Cell division: interaction with Ndr kinase leads to the promotion of PI3K/Akt signalling Role in tumour suppression: inhibits p53 activity and reduces p53 Proliferation: at higher concentrations, S100B blocks RAGE and stimulates FGFR1, leading to Ras/MEK/Erk-mediated proliferation; at low concentrations, S100B additionally stimulates RAGE-mediated p38 MAPK signalling, resulting in activation of the mitogenic program | [47,65,271] |
| 100G (CaBP-9k) | Epithelial cells | Vitamin D-dependent in the intestine | - | Cellular Ca2+ homeostasis: regulates intracellular Ca2+ levels and prevents toxic concentrations | [33] |
| S100P | Epithelial cells, leucocytes, | Promoter has binding sites for SMAD, STAT/CREB, and SP/KLF During embryonic implantation: highly expressed in the trophoblastic layer of the embryo, and in the endometrium of the uterine wall | RAGE, IQGAP1, enzrin, NMIIA | Cell proliferation and survival: activates RAGE-mediated NF-κB signalling; interaction with IQGAP1 induces MAPK signalling cascade Cell migration: promotes interaction with F-actin (via enzrin) and reduces focal adhesion sites (via NMIIA) | [48,272] |
| S100Z | Leucocytes | - | - | - | [8] |
References
- Moore, B.W. A soluble protein characteristic of the nervous system. Biochem. Biophys. Res. Commun. 1965, 19, 739–744. [Google Scholar] [CrossRef]
- Isobe, T.; Okuyama, T. The amino-acid sequence of S-100 protein (PAP I-b Protein) and Its relation to the calcium-binding proteins. Eur. J. Biochem. 1978, 89, 379–388. [Google Scholar] [CrossRef]
- Isobe, T.; Okuyama, T. The Amino-Acid Sequence Of The α subunit in bovine brain S-100a protein. Eur. J. Biochem. 1981, 116, 79–86. [Google Scholar] [CrossRef]
- Kozlyuk, N.; Monteith, A.J.; Garcia, V.; Damo, S.M.; Skaar, E.P.; Chazin, W.J. S100 proteins in the innate immune response to pathogens. Methods Mol. Biol. 2019, 1929, 275–290. [Google Scholar] [CrossRef]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef] [PubMed]
- Gilston, B.A.; Skaar, E.P.; Chazin, W.J. Binding of transition metals to S100 proteins. Sci. China Life Sci. 2016, 59, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Kiss, B.; Ecsédi, P.; Simon, M.; Nyitray, L. Isolation and characterization of S100 protein-protein complexes. Methods Mol. Biol. 2019, 1929, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Calderone, V.; Fragai, M.; Luchinat, C. Reviewing the crystal structure of S100Z and other members of the S100 family: Implications in calcium-regulated quaternary structure. Methods Mol. Biol. 2019, 1929, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Donato, R. S100: A multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int. J. Biochem. Cell Biol. 2001, 33, 637–668. [Google Scholar] [CrossRef]
- Spratt, D.E.; Barber, K.R.; Marlatt, N.M.; Ngo, V.; Macklin, J.A.; Xiao, Y.; Konermann, L.; Duennwald, M.L.; Shaw, G.S. A subset of calcium-binding S100 proteins show preferential heterodimerization. FEBS 2019, 286, 1859–1876. [Google Scholar] [CrossRef]
- Melville, Z.; Aligholizadeh, E.; McKnight, L.E.; Weber, D.J.; Pozharski, E.; Weber, D.J. X-ray crystal structure of human calcium-bound S100A1. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2017, 73, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Bunick, C.G.; Chazin, W.J. Target selectivity in EF-hand calcium binding proteins. Biochim. Biophys. Acta 2004, 1742, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Saiki, Y.; Horii, A. Multiple functions of S100A10, an important cancer promoter. Pathol. Int. 2019, 69, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, D.; Kessel, C.; Foell, D. S100 proteins in autoinflammation. In Textbook of Autoinflammation; Hashkes, P.J., Laxer, R.M., Simon, A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; Volume 19, pp. 149–163. [Google Scholar]
- Nakashige, T.G.; Zhang, B.; Krebs, C.; Nolan, E.M. Human calprotectin is an iron-sequestering host-defense protein. Nat. Chem. Biol. 2015, 11, 765–771. [Google Scholar] [CrossRef]
- Corbin, B.D.; Seeley, E.H.; Raab, A.; Feldmann, J.; Miller, M.R.; Torres, V.J.; Anderson, K.L.; Dattilo, B.M.; Dunman, P.M.; Gerads, R.; et al. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 2008, 319, 962–965. [Google Scholar] [CrossRef]
- Damoa, S.M.; Kehl-Fieb, T.E.; Sugitania, N.; Holta, M.E.; Rathia, S.; Murphya, W.J.; Zhangb, Y.; Betzc, C.; Hencha, L.; Fritzc, G.; et al. Molecular basis for manganese sequestration by calprotectin and roles in the innate immune response to invading bacterial pathogens. Proc. Natl. Acad. Sci. USA 2013, 110, 3841–3846. [Google Scholar] [CrossRef]
- Marenholz, I.; Heizmann, C.W.; Fritz, G. S100 proteins in mouse and man: From evolution to function and pathology (including an update of the nomenclature). Biochem. Biophys. Res. Commun. 2004, 322, 1111–1122. [Google Scholar] [CrossRef]
- Völkers, M.; Rohde, D.; Goodman, C.; Most, P. S100A1: A regulator of striated muscle sarcoplasmic reticulum Ca2+ handling, sarcomeric, and mitochondrial function. J. Biomed. Biotechnol. 2010, 2010, 178614. [Google Scholar] [CrossRef]
- Leśniak, W. Epigenetic regulation of S100 protein expression. Clin. Epigenet. 2011, 2, 77–83. [Google Scholar] [CrossRef]
- Källberg, E.; Vogl, T.; Liberg, D.; Olsson, A.; Bjo, P.; Wikstro, P.; Bergh, A.; Roth, J.; Ivars, F.; Leanderson, T. S100A9 interaction with tlr4 promotes tumor growth. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Ite, K.; Yonezawa, K.; Kitanishi, K.; Shimizu, N.; Unno, M. Optimal mutant model of human S100A3 protein citrullinated at arg51 by peptidylarginine deiminase type III and its solution structural properties. ACS Omega 2020, 5, 4032–4042. [Google Scholar] [CrossRef] [PubMed]
- Wolf, R.; Ruzicka, T.; Yuspa, S.H. Novel S100A7 (psoriasin)/S100A15 (koebnerisin) subfamily: Highly homologous but distinct in regulation and function. Amino Acids 2011, 41, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Kypriotou, M.; Huber, M.; Hohl, D. The human epidermal differentiation complex: Cornified envelope precursors, S100 proteins and the ’fused genes’ family. Exp. Dermatol. 2012, 21, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Choe, N.; Kwon, D.-H.; Shin, S.; Kim, Y.S.; Kim, Y.-K.; Kim, J.; Ahn, Y.; Eom, G.H.; Kook, H. The microRNA miR-124 inhibits vascular smooth muscle cell proliferation by targeting S100 calcium-binding protein A4 (S100A4). FEBS Lett. 2017, 591, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Sun, J.; Chen, X.; Du, R. MiR-135b-dependent down-regulation of S100B promotes neural stem cell differentiation in a hypoxia/ischemia-induced cerebral palsy rat model. Am. J. Physiol. Cell Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Sorci, G.; Giambanco, I. S100A6 protein: Functional roles. Cell. Mol. Life Sci. CMLS 2017, 74, 2749–2760. [Google Scholar] [CrossRef]
- Son, E.D.; Kim, H.-J.; Kim, K.H.; Bin, B.H.; Bae, I.-H.; Lim, K.-M.; Yu, S.J.; Cho, E.-G.; Lee, T.R. S100A7 (psoriasin) inhibits human epidermal differentiation by enhanced IL-6 secretion through IκB/NF-κB signalling. Exp. Dermatol. 2016, 25, 636–641. [Google Scholar] [CrossRef]
- Pruenster, M.; Vogl, T.; Roth, J.; Sperandio, M. S100A8/A9: From basic science to clinical application. Pharmacol. Ther. 2016, 167, 120–131. [Google Scholar] [CrossRef]
- Bertheloot, D.; Latz, E. HMGB1, IL-1α, IL-33 and S100 proteins: Dual-function alarmins. Cell. Mol. Immunol. 2017, 14, 43–64. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Bagur, R.; Hajnóczky, G. Intracellular Ca2+ sensing: Its role in calcium homeostasis and signaling. Mol. Cell 2017, 66, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.-J.; Jeung, E.-B. Biological significance of calbindin-D9k within duodenal epithelium. Int. J. Mol. Sci. 2013, 14, 23330–23340. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, L.C.; Anderson, J.A.; Morrison, A.J.; Wong, C.E.; Harms, M.J. Conservation of specificity in two low-specificity proteins. Biochemistry 2018, 57, 684–695. [Google Scholar] [CrossRef] [PubMed]
- Madureira, P.A.; O’Connell, P.A.; Surette, A.P.; Miller, V.A.; Waisman, D.M. The biochemistry and regulation of S100A10: A multifunctional plasminogen receptor involved in oncogenesis. J. Biomed. Biotechnol. 2012, 2012, 353687. [Google Scholar] [CrossRef]
- Ritterhoff, J.; Most, P. Targeting S100A1 in heart failure. Gene Ther. 2012, 19, 613–621. [Google Scholar] [CrossRef]
- Rohde, D.; Ritterhoff, J.; Voelkers, M.; Katus, H.A.; Parker, T.G.; Most, P. S100A1: A multifaceted therapeutic target in cardiovascular disease. J. Cardiovasc. Transl. Res. 2010, 3, 525–537. [Google Scholar] [CrossRef]
- Kessel, C.; Holzinger, D.; Foell, D. Phagocyte-derived S100 proteins in autoinflammation: Putative role in pathogenesis and usefulness as biomarkers. Clin. Immunol. 2013, 147, 229–241. [Google Scholar] [CrossRef]
- Timmermans, K.; Kox, M.; Scheffer, G.J.; Pickkers, P. Danger in the intensive care unit: Damps in critically ill patients. Shock 2016, 45, 108–116. [Google Scholar] [CrossRef]
- Markowitz, J.; Carson, W.E. Review of S100A9 biology and its role in cancer. Biochim. Biophys. Acta 2013, 1835, 100–109. [Google Scholar] [CrossRef]
- Carvalho, A.; Lu, J.; Francis, J.D.; Moore, R.E.; Haley, K.P.; Doster, R.S.; Townsend, S.D.; Johnson, J.G.; Damo, S.M.; Gaddy, J.A. S100A12 in digestive diseases and health: A scoping review. Gastroenterol. Res. Pract. 2020, 2020, 2868373. [Google Scholar] [CrossRef]
- Krop, I.; März, A.; Carlsson, H.; Li, X.; Bloushtain-Qimron, N.; Hu, M.; Gelman, R.; Sabel, M.S.; Schnitt, S.; Ramaswamy, S.; et al. A putative role for psoriasin in breast tumor progression. Cancer Res. 2005, 65, 11326–11334. [Google Scholar] [CrossRef]
- Goyette, J.; Geczy, C.L. Inflammation-associated S100 proteins: New mechanisms that regulate function. Amino Acids 2011, 41, 821–842. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, R.; Kastrisianaki, E.; Giambanco, I.; Donato, R. S100B protein stimulates microglia migration via RAGE-dependent up-regulation of chemokine expression and release. J. Biol. Chem. 2011, 286, 7214–7226. [Google Scholar] [CrossRef] [PubMed]
- Dahlmann, M.; Kobelt, D.; Walther, W.; Mudduluru, G.; Stein, U. S100A4 in cancer metastasis: Wnt signaling-driven interventions for metastasis restriction. Cancers 2016, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.e.; Chen, H.; Luo, A.; Ding, F.; Liu, Z. S100A14 stimulates cell proliferation and induces cell apoptosis at different concentrations via receptor for advanced glycation end products (RAGE). PLoS ONE 2011, 6, e19375. [Google Scholar] [CrossRef]
- Riuzzi, F.; Sorci, G.; Beccafico, S.; Donato, R. S100B engages RAGE or bFGF/FGFR1 in myoblasts depending on its own concentration and myoblast density. Implications for muscle regeneration. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Prica, F.; Radon, T.; Cheng, Y.; Crnogorac-Jurcevic, T. The life and works of S100P–from conception to cancer. Am. J. Cancer Res. 2016, 6, 562–576. [Google Scholar]
- Sakaguchi, M.; Huh, N.-H. S100A11, a dual growth regulator of epidermal keratinocytes. Amino Acids 2011, 41, 797–807. [Google Scholar] [CrossRef]
- He, H.; Li, J.; Weng, S.; Li, M.; Yu, Y. S100A11: Diverse function and pathology corresponding to different target proteins. Cell Biochem. Biophys. 2009, 55, 117–126. [Google Scholar] [CrossRef]
- Pan, S.-C.; Li, C.-Y.; Kuo, C.-Y.; Kuo, Y.-Z.; Fang, W.-Y.; Huang, Y.-H.; Hsieh, T.-C.; Kao, H.-Y.; Kuo, Y.; Kang, Y.-R.; et al. The p53-S100A2 positive feedback loop negatively regulates epithelialization in cutaneous wound healing. Sci. Rep. 2018, 8, 5458. [Google Scholar] [CrossRef]
- Boye, K.; Maelandsmo, G.M. S100A4 and metastasis: A small actor playing many roles. Am. J. Pathol. 2010, 176, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Basnet, S.; Sharma, S.; Costea, D.E.; Sapkota, D. Expression profile and functional role of S100A14 in human cancer. Oncotarget 2019, 10, 2996–3012. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Yang, Q.; Wilder, P.T.; Carrier, F.; Weber, D.J. The calcium-binding protein S100B down-regulates p53 and apoptosis in malignant melanoma. J. Biol. Chem. 2010, 285, 27487–27498. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Austermann, J.; Spiekermann, C.; Roth, J. S100 proteins in rheumatic diseases. Nat. Rev. Rheumatol. 2018, 14, 528–541. [Google Scholar] [CrossRef] [PubMed]
- Perera, C.; McNeil, H.P.; Geczy, C.L. S100 calgranulins in inflammatory arthritis. Immunol. Cell Biol. 2010, 88, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Friedman, B.; Cronstein, B. Methotrexate mechanism in treatment of rheumatoid arthritis. Jt. Bone Spine 2019, 86, 301–307. [Google Scholar] [CrossRef]
- Boschetti, G.; Garnero, P.; Moussata, D.; Cuerq, C.; Préaudat, C.; Duclaux-Loras, R.; Mialon, A.; Drai, J.; Flourié, B.; Nancey, S. Accuracies of serum and fecal S100 proteins (calprotectin and calgranulin C) to predict the response to TNF antagonists in patients with Crohn’s disease. Inflamm. Bowel Dis. 2015, 21, 331–336. [Google Scholar] [CrossRef]
- Foell, D.; Frosch, M.; Sorg, C.; Roth, J. Phagocyte-specific calcium-binding S100 proteins as clinical laboratory markers of inflammation. Clin. Chim. Acta 2004, 344, 37–51. [Google Scholar] [CrossRef]
- Heizmann, C.W. S100 proteins: Diagnostic and prognostic biomarkers in laboratory medicine. Biochim. Biophys. Acta 2019, 1866, 1197–1206. [Google Scholar] [CrossRef]
- Undén, J.; Ingebrigtsen, T.; Romner, B. Scandinavian guidelines for initial management of minimal, mild and moderate head injuries in adults: An evidence and consensus-based update. BMC Med. 2013, 11. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.C.; Harmon, C.; McCann, M.; Gunyan, H.; Bazarian, J.J. S100B outperforms clinical decision rules for the identification of intracranial injury on head CT scan after mild traumatic brain injury. Brain Inj. 2020, 34, 407–414. [Google Scholar] [CrossRef]
- Vedin, T.; Svensson, S.; Edelhamre, M.; Karlsson, M.; Bergenheim, M.; Larsson, P.-A. Management of mild traumatic brain injury-trauma energy level and medical history as possible predictors for intracranial hemorrhage. Eur. J. Trauma Emerg. Surg. 2019, 45, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Michetti, F.; D’Ambrosi, N.; Toesca, A.; Puglisi, M.A.; Serrano, A.; Marchese, E.; Corvino, V.; Geloso, M.C. The S100B story: From biomarker to active factor in neural injury. J. Neurochem. 2019, 148, 168–187. [Google Scholar] [CrossRef] [PubMed]
- Cristóvão, J.S.; Gomes, C.M. S100 proteins in alzheimer’s disease. Front. Neurosci. 2019, 13, 463. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Liu, B.; Guo, R.; Luo, J.; Li, H.; Li, Z.; Xu, W. Elevated plasma S100A1 level is a risk factor for ST-segment elevation myocardial infarction and associated with post-infarction cardiac function. Int. J. Med. Sci. 2019, 16, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Heizmann, C.W. Ca2+-Binding proteins of the EF-hand superfamily: Diagnostic and prognostic biomarkers and novel therapeutic targets. Methods Mol. Biol. 2019, 1929, 157–186. [Google Scholar] [CrossRef] [PubMed]
- Rohde, D.; Brinks, H.; Ritterhoff, J.; Qui, G.; Ren, S.; Most, P. S100A1 gene therapy for heart failure: A novel strategy on the verge of clinical trials. J. Mol. Cell. Cardiol. 2011, 50, 777–784. [Google Scholar] [CrossRef]
- Gaynor, R.; Irie, R.; Morton, D.; Herschman, H.R. S100 protein is present in cultured human malignant melanomas. Nature 1980, 286, 400–401. [Google Scholar] [CrossRef]
- Gaynor, R.; Herschman, H.R.; Irie, R.; Jones, P.; Morton, D.; Cochran, A. S100 protein: A marker for human malignant melanomas? Lancet 1981, 1, 869–871. [Google Scholar] [CrossRef]
- Takashi, M.; Haimoto, H.; Murase, T.; Mitsuya, H.; Kato, K. An immunochemical and immunohistochemical study of S100 protein in renal cell carcinoma. Cancer 1988, 61, 889–895. [Google Scholar] [CrossRef]
- Gillett, C.E.; Bobrow, L.G.; Millis, R.R. S100 protein in human mammary tissue--immunoreactivity in breast carcinoma, including Paget’s disease of the nipple, and value as a marker of myoepithelial cells. J. Pathol. 1990, 160, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Kuźnicki, J.; Filipek, A.; Hunziker, P.E.; Huber, S.; Heizmann, C.W. Calcium-binding protein from mouse Ehrlich ascites-tumour cells is homologous to human calcyclin. Biochem. J. 1989, 263, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Budka, H. Glial fibrillary acidic protein and S-100 protein in human hepatic encephalopathy: Immunocytochemical demonstration of dissociation of two glia-associated proteins. Acta Neuropathol. 1986, 70, 17–21. [Google Scholar] [CrossRef]
- Bresnick, A.R.; Weber, D.J.; Zimmer, D.B. S100 proteins in cancer. Nat. Rev. Cancer 2015, 15, 96–109. [Google Scholar] [CrossRef]
- Hua, X.; Zhang, H.; Jia, J.; Chen, S.; Sun, Y.; Zhu, X. Roles of S100 family members in drug resistance in tumors: Status and prospects. Biomed. Pharmacother. 2020, 127, 110156. [Google Scholar] [CrossRef]
- Wolf, S.; Haase-Kohn, C.; Pietzsch, J. S100A2 in cancerogenesis: A friend or a foe? Amino Acids 2011, 41, 849–861. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Yates, L.R.; Knappskog, S.; Wedge, D.; Farmery, J.H.R.; Gonzalez, S.; Martincorena, I.; Alexandrov, L.B.; van Loo, P.; Haugland, H.K.; Lilleng, P.K.; et al. Genomic evolution of breast cancer metastasis and relapse. Cancer Cell 2017, 32, 169–184.e167. [Google Scholar] [CrossRef]
- Li, F.; Men, X.; Zhang, W. S100 protein in breast tumor. Indian J. Cancer 2014, 51 (Suppl. S3), 67–71. [Google Scholar] [CrossRef]
- Cancemi, P.; Bivona, S.; Pucci-Minafra, I.; Buttacavoli, M.; Di Cara, G.; Albanese, N.N.; Feo, S. A multiomics analysis of S100 protein family in breast cancer. Oncotarget 2018, 9, 29064–29081. [Google Scholar] [CrossRef] [PubMed]
- Yen, M.C.; Huang, Y.C.; Kan, J.Y.; Kuo, P.L.; Hou, M.F.; Hsu, Y.L. S100B expression in breast cancer as a predictive marker for cancer metastasis. Int. J. Oncol. 2018, 52, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, Z.; Liu, W.; Lei, R.; Shan, J.; Li, L.; Wang, X. Distinct prognostic values of S100 mRNA expression in breast cancer. Sci. Rep. 2017, 7, 39786. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.M.; Li, J.; Kang, A.D.; Huang, S.Q.; Liu, W.B.; Zhang, Y.; Liu, Z.H.; Zeng, L. Protein S100-A8: A potential metastasis-associated protein for breast cancer determined via iTRAQ quantitative proteomic and clinicopathological analysis. Oncol. Lett. 2018, 15, 5285–5293. [Google Scholar] [CrossRef]
- Charmsaz, S.; Hughes, É.; Bane, F.T.; Tibbitts, P.; McIlroy, M.; Byrne, C.; Cocchiglia, S.; McBryan, J.; Hennessy, B.T.; Dwyer, R.M.; et al. S100β as a serum marker in endocrine resistant breast cancer. BMC Med. 2017, 15, 79. [Google Scholar] [CrossRef]
- Darlix, A.; Lamy, P.-J.; Lopez-Crapez, E.; Braccini, A.L.; Firmin, N.; Romieu, G.; Thezenas, S.; Jacot, W. Serum HER2 extra-cellular domain, S100ß and CA 15-3 levels are independent prognostic factors in metastatic breast cancer patients. BMC Cancer 2016, 16, 428. [Google Scholar] [CrossRef]
- McIlroy, M.; McCartan, D.; Early, S.; Gaora, P.Ó.; Pennington, S.; Hill, A.D.K.; Young, L.S. Interaction of developmental transcription factor HOX11 with steroid receptor coactivator SRC-1 mediates resistance to endocrine therapy in breast cancer. Cancer Res. 2010. [Google Scholar] [CrossRef]
- Fei, F.; Qu, J.; Zhang, M.; Li, Y.; Zhang, S. S100A4 in cancer progression and metastasis: A systematic review. Oncotarget 2017, 8, 73219–73239. [Google Scholar] [CrossRef]
- Jenkinson, S.R.; Barraclough, R.; West, C.R.; Rudland, P.S. S100A4 regulates cell motility and invasion in an in vitro model for breast cancer metastasis. Br. J. Cancer 2004, 90, 253–262. [Google Scholar] [CrossRef]
- Barraclough, R.; Rudland, P.S. The S-100-related calcium-binding protein, p9Ka, and metastasis in rodent and human mammary cells. Eur. J. Cancer 1994, 30a, 1570–1576. [Google Scholar] [CrossRef]
- Li, Z.H.; Dulyaninova, N.G.; House, R.P.; Almo, S.C.; Bresnick, A.R. S100A4 regulates macrophage chemotaxis. Mol. Biol. Cell 2010, 21, 2598–2610. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.S.; Wang, M.Y.; Wu, S.N.; Su, J.L.; Hong, C.C.; Chuang, S.E.; Chen, M.W.; Hua, K.T.; Wu, Y.L.; Cha, S.T.; et al. CTGF enhances the motility of breast cancer cells via an integrin-alphavbeta3-ERK1/2-dependent S100A4-upregulated pathway. J. Cell Sci. 2007, 120, 2053–2065. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, S.; Fernig, D.G.; Martin-Fernandez, M.; Rudland, P.S.; Barraclough, R. Mutually antagonistic actions of S100A4 and S100A1 on normal and metastatic phenotypes. Oncogene 2005, 24, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, M.; Zhou, Y.; Wang, F.; Li, X.; Wang, L.; Fan, Q. S100A4 participates in epithelial-mesenchymal transition in breast cancer via targeting MMP2. Tumour Biol. 2016, 37, 2925–2932. [Google Scholar] [CrossRef] [PubMed]
- Cabezón, T.; Celis, J.E.; Skibshøj, I.; Klingelhöfer, J.; Grigorian, M.; Gromov, P.; Rank, F.; Myklebust, J.H.; Maelandsmo, G.M.; Lukanidin, E.; et al. Expression of S100A4 by a variety of cell types present in the tumor microenvironment of human breast cancer. Int. J. Cancer 2007, 121, 1433–1444. [Google Scholar] [CrossRef]
- Prasmickaite, L.; Tenstad, E.M.; Pettersen, S.; Jabeen, S.; Egeland, E.V.; Nord, S.; Pandya, A.; Haugen, M.H.; Kristensen, V.N.; Børresen-Dale, A.-L.; et al. Basal-like breast cancer engages tumor-supportive macrophages via secreted factors induced by extracellular S100A4. Mol. Oncol. 2018, 12, 1540–1558. [Google Scholar] [CrossRef]
- Wang, N.; Ma, Q.; Wang, Y.; Ma, G.; Zhai, H. CacyBP/SIP expression is involved in the clinical progression of breast cancer. World J. Surg. 2010, 34, 2545–2552. [Google Scholar] [CrossRef]
- Topolska-Woś, A.M.; Chazin, W.J.; Filipek, A. CacyBP/SIP—Structure and variety of functions. Biochim. Biophys. Acta 2016, 1860, 79–85. [Google Scholar] [CrossRef]
- Leong, S.; Christopherson, R.I.; Baxter, R.C. Profiling of Apoptotic changes in human breast cancer cells using seldi-tof mass spectrometry. Cell. Physiol. Biochem. 2007, 20, 579–590. [Google Scholar] [CrossRef]
- Nasser, M.W.; Wani, N.A.; Ahirwar, D.K.; Powell, C.A.; Ravi, J.; Elbaz, M.; Zhao, H.; Padilla, L.; Zhang, X.; Shilo, K.; et al. RAGE mediates S100A7-induced breast cancer growth and metastasis by modulating the tumor microenvironment. Cancer Res. 2015, 75, 974–985. [Google Scholar] [CrossRef]
- Liu, H.; Wang, L.; Wang, X.; Cao, Z.; Yang, Q.; Zhang, K. S100A7 enhances invasion of human breast cancer MDA-MB-468 cells through activation of nuclear factor-κB signaling. World J. Surg. Oncol. 2013, 11. [Google Scholar] [CrossRef] [PubMed]
- Nasser, M.W.; Qamri, Z.; Deol, Y.S.; Ravi, J.; Powell, C.A.; Trikha, P.; Schwendener, R.A.; Bai, X.-F.; Shilo, K.; Zou, X.; et al. S100A7 enhances mammary tumorigenesis through upregulation of inflammatory pathways. Cancer Res. 2012, 72, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Emberley, E.D.; Murphy, L.C.; Watson, P.H. S100A7 and the progression of breast cancer. Breast Cancer Res. 2004, 6, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Li, H.; Zhang, B.; Liu, Y.; Lu, G.; Lu, S.; Sun, L.; Qi, Y.; Li, X.; Chen, W. RAGE-binding S100A8/A9 promotes the migration and invasion of human breast cancer cells through actin polymerization and epithelial-mesenchymal transition. Breast Cancer Res. Treat. 2013, 142, 297–309. [Google Scholar] [CrossRef]
- Cassim, S.; Chepulis, L.; Keenan, R.; Kidd, J.; Firth, M.; Lawrenson, R. Patient and carer perceived barriers to early presentation and diagnosis of lung cancer: A systematic review. BMC Cancer 2019, 19, 25. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Ju, D.-T.; Chang, C.-F.; Muralidhar Reddy, P.; Velmurugan, B.K. A review on the effects of current chemotherapy drugs and natural agents in treating non-small cell lung cancer. Biomedicine 2017, 7, 23. [Google Scholar] [CrossRef]
- Khanmohammadi, A.; Aghaie, A.; Vahedi, E.; Qazvini, A.; Ghanei, M.; Afkhami, A.; Hajian, A.; Bagheri, H. Electrochemical biosensors for the detection of lung cancer biomarkers: A review. Talanta 2020, 206, 120251. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, J.; Tang, Y.-L.; Huang, L.; Zhou, C.-Y.; Xu, J.-X. Prognostic roles of mRNA expression of S100 in non-small-cell lung cancer. BioMed Res. Int. 2018, 2018, 9815806. [Google Scholar] [CrossRef]
- Wang, T.; Huo, X.; Chong, Z.; Khan, H.; Liu, R. A review of S100 protein family in lung cancer. Clin. Chim. Acta 2018, 476, 54–59. [Google Scholar] [CrossRef]
- Feng, G.; Xu, X.-C.; Youssef, E.M.; Lotan, R. Diminished expression of S100A2, a putative tumor suppressor, at early stage of human lung carcinogenesis. Cancer Res. 2001, 61, 7999–8004. [Google Scholar]
- Heighway, J.; Knapp, T.; Boyce, L.; Brennand, S.; Field, J.K.; Betticher, D.C.; Ratschiller, D.; Gugger, M.; Donovan, M.; Lasek, A.; et al. Expression profiling of primary non-small cell lung cancer for target identification. Oncogene 2002, 21, 7749–7763. [Google Scholar] [CrossRef] [PubMed]
- Bulk, E.; Sargin, B.; Krug, U.; Hascher, A.; Jun, Y.; Knop, M.; Kerkhoff, C.; Gerke, V.; Liersch, R.; Mesters, R.M.; et al. S100A2 induces metastasis in non-small cell lung cancer. Clin. Cancer Res. 2009, 15, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Hountis, P.; Matthaios, D.; Froudarakis, M.; Bouros, D.; Kakolyris, S. S100A2 protein and non-small cell lung cancer. The dual role concept. Tumour Biol. 2014, 35, 7327–7333. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Siddique, H.R.; Saleem, M. S100A4 calcium-binding protein is key player in tumor progression and metastasis: Preclinical and clinical evidence. Cancer Metastasis Rev. 2012, 31, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Sato, D.; Saiki, Y.; Sunamura, M.; Fukushige, S.; Horii, A. S100A4 is frequently overexpressed in lung cancer cells and promotes cell growth and cell motility. Biochem. Biophys. Res. Commun. 2014, 447, 459–464. [Google Scholar] [CrossRef]
- Stewart, R.L.; Carpenter, B.L.; West, D.S.; Knifley, T.; Liu, L.; Wang, C.; Weiss, H.L.; Gal, T.S.; Durbin, E.B.; Arnold, S.M.; et al. S100A4 drives non-small cell lung cancer invasion, associates with poor prognosis, and is effectively targeted by the FDAapproved anti-helminthic agent niclosamide. Oncotarget 2016, 7, 34630–34642. [Google Scholar] [CrossRef]
- Hou, S.; Tian, T.; Qi, D.; Sun, K.; Yuan, Q.; Wang, Z.; Qin, Z.; Wu, Z.; Chen, Z.; Zhang, J. S100A4 promotes lung tumor development through β-catenin pathway-mediated autophagy inhibition. Cell Death Dis. 2018, 9, 277. [Google Scholar] [CrossRef]
- Liu, G.; Wu, Q.; Liu, G.; Song, X.; Zhang, J. Knockdown of S100A7 reduces lung squamous cell carcinoma cell growth in vitro and in vivo. Int. J. Clin. Exp. Pathol. 2014, 7, 8279–8289. [Google Scholar]
- Wang, R.; Li, Y.; Hu, E.; Kong, F.; Wang, J.; Liu, J.; Shao, Q.; Hao, Y.; He, D.; Xiao, X. S100A7 promotes lung adenocarcinoma to squamous carcinoma transdifferentiation, and its expression is differentially regulated by the Hippo-YAP pathway in lung cancer cells. Oncotarget 2017, 8, 24804–24814. [Google Scholar] [CrossRef]
- Choi, H.; Puvenna, V.; Brennan, C.; Mahmoud, S.; Wang, X.-F.; Phillips, M.; Janigro, D.; Mazzone, P. S100B and S100B autoantibody as biomarkers for early detection of brain metastases in lung cancer. Transl. Lung Cancer Res. 2016, 5, 413–419. [Google Scholar] [CrossRef]
- Pang, X.; Min, J.; Liu, L.; Liu, Y.; Ma, N.; Zhang, H. S100B protein as a possible participant in the brain metastasis of NSCLC. Med. Oncol. 2012, 29, 2626–2632. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Jia, Q.; Liu, L.; Zhao, X.; Tan, A.; Ma, N.; Zhang, H. S100B promotes the proliferation, migration and invasion of specific brain metastatic lung adenocarcinoma cell line. Cell Biochem. Funct. 2011, 29, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Domingues, B.; Lopes, J.M.; Soares, P.; Pópulo, H. Melanoma treatment in review. ImmunoTargets Ther. 2018, 7, 35–49. [Google Scholar] [CrossRef]
- Deeks, E.D. Pembrolizumab: A review in advanced melanoma. Drugs 2016, 76, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.-F.; Pan, F.-Q.; Li, D. Expression and clinical significance of S100 family genes in patients with melanoma. Melanoma Res. 2019, 29, 23–29. [Google Scholar] [CrossRef]
- Herwig, N.; Belter, B.; Wolf, S.; Haase-Kohn, C.; Pietzsch, J. Interaction of extracellular S100A4 with RAGE prompts prometastatic activation of A375 melanoma cells. J. Cell. Mol. Med. 2016, 20, 825–835. [Google Scholar] [CrossRef]
- Herwig, N.; Belter, B.; Pietzsch, J. Extracellular S100A4 affects endothelial cell integrity and stimulates transmigration of A375 melanoma cells. Biochem. Biophys. Res. Commun. 2016, 477, 963–969. [Google Scholar] [CrossRef]
- Haase-Kohn, C.; Wolf, S.; Lenk, J.; Pietzsch, J. Copper-mediated cross-linking of S100A4, but not of S100A2, results in proinflammatory effects in melanoma cells. Biochem. Biophys. Res. Commun. 2011, 413, 494–498. [Google Scholar] [CrossRef]
- Bettum, I.J.; Vasiliauskaite, K.; Nygaard, V.; Clancy, T.; Pettersen, S.J.; Tenstad, E.; Mælandsmo, G.M.; Prasmickaite, L. Metastasis-associated protein S100A4 induces a network of inflammatory cytokines that activate stromal cells to acquire pro-tumorigenic properties. Cancer Lett. 2014, 344, 28–39. [Google Scholar] [CrossRef]
- Gebhardt, C.; Sevko, A.; Jiang, H.; Lichtenberger, R.; Reith, M.; Tarnanidis, K.; Holland-Letz, T.; Umansky, L.; Beckhove, P.; Sucker, A.; et al. Myeloid cells and related chronic inflammatory factors as novel predictive markers in melanoma treatment with ipilimumab. Clin. Cancer Res. 2015, 21, 5453–5459. [Google Scholar] [CrossRef]
- Shirley, S.H.; Maltzan, K.V.; Robbins, P.O.; Kusewitt, D.F. Melanocyte and melanoma cell activation by calprotectin. J. Ski. Cancer 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Lee, Y.-C.; Zhang, Z.; Chandra, G.; Su, S.-B.; Mukherjee, A.B. Lack of an endogenous anti-inflammatory protein in mice enhances colonization of B16F10 melanoma cells in the lungs. J. Biol. Chem. 2010, 285, 10822–10831. [Google Scholar] [CrossRef] [PubMed]
- Ruma, I.M.W.; Putranto, E.W.; Kondo, E.; Murata, H.; Watanabe, M.; Huang, P.; Kinoshita, R.; Futami, J.; Inoue, Y.; Yamauchi, A.; et al. MCAM, as a novel receptor for S100A8/A9, mediates progression of malignant melanoma through prominent activation of NF-κB and ROS formation upon ligand binding. Clin. Exp. Metastasis 2016, 33, 609–627. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sumardika, I.W.; Tomonobu, N.; Winarsa Ruma, I.M.; Kinoshita, R.; Kondo, E.; Inoue, Y.; Sato, H.; Yamauchi, A.; Murata, H.; et al. Melanoma cell adhesion molecule is the driving force behind the dissemination of melanoma upon S100A8/A9 binding in the original skin lesion. Cancer Lett. 2019, 452, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Hibino, T.; Sakaguchi, M.; Miyamoto, S.; Yamamoto, M.; Motoyama, A.; Hosoi, J.; Shimokata, T.; Ito, T.; Tsuboi, R.; Huh, N.-H. S100A9 is a novel ligand of EMMPRIN that promotes melanoma metastasis. Cancer Res. 2013, 73, 172–183. [Google Scholar] [CrossRef]
- Tomonobu, N.; Kinoshita, R.; Sakaguchi, M. S100 soil sensor receptors and molecular targeting therapy against them in cancer metastasis. Transl. Oncol. 2020, 13, 100753. [Google Scholar] [CrossRef]
- Harpio, R.; Einarsson, R. S100 proteins as cancer biomarkers with focus on S100B in malignant melanoma. Clin. Biochem. 2004, 37, 512–518. [Google Scholar] [CrossRef]
- Frauchiger, A.L.; Dummer, R.; Mangana, J. Serum S100B levels in melanoma. Methods Mol. Biol. 2019, 1929, 691–700. [Google Scholar] [CrossRef]
- Kruijff, S.; Hoekstra, H.J. The current status of S-100B as a biomarker in melanoma. Eur. J. Surg. Oncol. 2012, 38, 281–285. [Google Scholar] [CrossRef]
- Hartman, K.G.; Vitolo, M.I.; Pierce, A.D.; Fox, J.M.; Shapiro, P.; Martin, S.S.; Wilder, P.T.; Weber, D.J. Complex formation between S100B protein and the p90 ribosomal S6 kinase (RSK) in malignant melanoma is calcium-dependent and inhibits extracellular signal-regulated kinase (ERK)-mediated phosphorylation of RSK. J. Biol. Chem. 2014, 289, 12886–12895. [Google Scholar] [CrossRef]
- Lin, J.; Yang, Q.; Yan, Z.; Markowitz, J.; Wilder, P.T.; Carrier, F.; Weber, D.J. Inhibiting S100B restores p53 levels in primary malignant melanoma cancer cells. J. Biol. Chem. 2004, 279, 34071–34077. [Google Scholar] [CrossRef] [PubMed]
- Webster, M.R.; Fane, M.E.; Alicea, G.M.; Basu, S.; Kossenkov, A.V.; Marino, G.E.; Douglass, S.M.; Kaur, A.; Ecker, B.L.; Gnanapradeepan, K.; et al. Paradoxical role for wild-type p53 in driving therapy resistance in melanoma. Mol. Cell 2020, 77, 633–644.e635. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Li, L.-D.; Li, J.; Lu, X. Prognostic values of S100 family members in ovarian cancer patients. BMC Cancer 2018, 18, 1256. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Zhu, L.; Yang, L.; Cui, Y.; Zhan, Y. Prognostic values of S100 family mRNA expression in ovarian cancer. Cancer Biomark. 2019, 25, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Izycka, N.; Sterzynska, K.; Januchowski, R.; Nowak-Markwitz, E. Semaphorin 3A (SEMA3A), protocadherin 9 (PCdh9), and S100 calcium binding protein A3 (S100A3) as potential biomarkers of carcinogenesis and chemoresistance of different neoplasms, including ovarian cancer–review of literature. Ginekol. Pol. 2019, 90, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Gillet, J.-P.; Calcagno, A.M.; Varma, S.; Davidson, B.; Bunkholt Elstrand, M.; Ganapathi, R.; Kamat, A.A.; Sood, A.K.; Ambudkar, S.V.; Seiden, M.V.; et al. Multidrug resistance-linked gene signature predicts overall survival of patients with primary ovarian serous carcinoma. Clin. Cancer Res. 2012, 18, 3197–3206. [Google Scholar] [CrossRef]
- Yang, T.; Cheng, J.; You, J.; Yan, B.; Liu, H.; Li, F. S100B promotes chemoresistance in ovarian cancer stem cells by regulating p53. Oncol. Rep. 2018, 40, 1574–1582. [Google Scholar] [CrossRef]
- Yang, T.; Cheng, J.; Yang, Y.; Qi, W.; Zhao, Y.; Long, H.; Xie, R.; Zhu, B. S100B mediates stemness of ovarian cancer stem-like cells through inhibiting p53. Stem Cells 2017, 35, 325–336. [Google Scholar] [CrossRef]
- Lokman, N.A.; Pyragius, C.E.; Ruszkiewicz, A.; Oehler, M.K.; Ricciardelli, C. Annexin A2 and S100A10 are independent predictors of serous ovarian cancer outcome. Transl. Res. 2016, 171, 83–95. [Google Scholar] [CrossRef]
- Lokman, N.A.; Ho, R.; Gunasegaran, K.; Bonner, W.M.; Oehler, M.K.; Ricciardelli, C. Anti-tumour effects of all-trans retinoid acid on serous ovarian cancer. J. Exp. Clin. Cancer Res. 2019, 38, 10. [Google Scholar] [CrossRef]
- Noye, T.M.; Lokman, N.A.; Oehler, M.K.; Ricciardelli, C. S100A10 and cancer hallmarks: Structure, functions, and its emerging role in ovarian cancer. Int. J. Mol. Sci. 2018, 19, 4122. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ma, Y.; Fei, F.; Zheng, M.; Li, Z.; Zhao, Q.; Du, J.; Liu, K.; Lu, R.; Zhang, S. Critical role and its underlying molecular events of the plasminogen receptor, S100A10 in malignant tumor and non-tumor diseases. J. Cancer 2020, 11, 826–836. [Google Scholar] [CrossRef] [PubMed]
- Moravkova, P.; Kohoutova, D.; Rejchrt, S.; Cyrany, J.; Bures, J. Role of S100 proteins in colorectal carcinogenesis. Gastroenterol. Res. Pract. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Hemandas, A.K.; Salto-Tellez, M.; Maricar, S.H.; Leong, A.F.P.K.; Leow, C.K. Metastasis-associated protein S100A4--a potential prognostic marker for colorectal cancer. J. Surg. Oncol. 2006, 93, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-G.; Kim, C.-J.; Nam, S.-W.; Yoon, S.-H.; Lee, S.-H.; Yoo, N.-J.; Lee, J.-Y.; Park, W.-S. Overexpression of S100A4 is closely associated with progression of colorectal cancer. World J. Gastroenterol. 2005, 11, 4852–4856. [Google Scholar] [CrossRef] [PubMed]
- Moris, D.; Spartalis, E.; Angelou, A.; Margonis, G.-A.; Papalambros, A.; Petrou, A.; Athanasiou, A.; Schizas, D.; Dimitroulis, D.; Felekouras, E. The value of calprotectin S100A8/A9 complex as a biomarker in colorectal cancer: A systematic review. JBUON 2016, 21, 859–866. [Google Scholar]
- Hwang, C.-C.; Chai, H.-T.; Chen, H.-W.; Tsai, H.-L.; Lu, C.-Y.; Yu, F.-J.; Huang, M.-Y.; Wang, J.-Y. S100B protein expressions as an independent predictor of early relapse in UICC stages II and III colon cancer patients after curative resection. Ann. Surg. Oncol. 2011, 18, 139–145. [Google Scholar] [CrossRef]
- Boye, K.; Nesland, J.M.; Sandstad, B.; Mælandsmo, G.M.; Flatmark, K. Nuclear S100A4 is a novel prognostic marker in colorectal cancer. Eur. J. Cancer 2010, 46, 2919–2925. [Google Scholar] [CrossRef]
- Dahlmann, M.; Okhrimenko, A.; Marcinkowski, P.; Osterland, M.; Herrmann, P.; Smith, J.; Heizmann, C.W.; Schlag, P.M. RAGE mediates S100A4-induced cell motility via MAPK/ERK and hypoxia signaling and is a prognostic biomarker for human colorectal cancer metastasis. Oncotarget 2014, 5, 3220–3233. [Google Scholar] [CrossRef]
- Wang, H.; Duan, L.; Zou, Z.; Li, H.; Yuan, S.; Chen, X.; Zhang, Y.; Li, X.; Sun, H.; Zha, H.; et al. Activation of the PI3K/Akt/mTOR/p70S6K pathway is involved in S100A4-induced viability and migration in colorectal cancer cells. Int. J. Med. Sci. 2014, 11, 841–849. [Google Scholar] [CrossRef]
- Duan, L.; Wu, R.; Ye, L.; Wang, H.; Yang, X.; Zhang, Y.; Chen, X.; Zuo, G.; Zhang, Y.; Weng, Y.; et al. S100A8 and S100A9 are associated with colorectal carcinoma progression and contribute to colorectal carcinoma cell survival and migration via Wnt/β-catenin pathway. PLoS ONE 2013, 8, e62092. [Google Scholar] [CrossRef]
- Ichikawa, M.; Williams, R.; Wang, L.; Vogl, T.; Srikrishna, G. S100A8/A9 activate key genes and pathways in colon tumor progression. Mol. Cancer Res. 2011, 9, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Wang, F.; Yin, X.; Chen, L.; Li, G.; Lin, F.; Ni, W.; Wu, J.; Jin, R.; Jiang, L. Overexpression of S100P promotes colorectal cancer metastasis and decreases chemosensitivity to 5-FU in vitro. Mol. Cell. Biochem. 2014, 389, 257–264. [Google Scholar] [CrossRef]
- Jiang, L.; Lai, Y.-K.; Zhang, J.; Wang, H.; Lin, M.C.; He, M.-L.; Kung, H.-F. Targeting S100P inhibits colon cancer growth and metastasis by Lentivirus-mediated RNA interference and proteomic analysis. Mol. Med. 2011, 17, 709–716. [Google Scholar] [CrossRef][Green Version]
- Lin, F.; Zhang, P.; Zuo, Z.; Wang, F.; Bi, R.; Shang, W.; Wu, A.; Ye, J.; Li, S.; Sun, X.; et al. Thioredoxin-1 promotes colorectal cancer invasion and metastasis through crosstalk with S100P. Cancer Lett. 2017, 401, 1–10. [Google Scholar] [CrossRef]
- Leclerc, E.; Vetter, S.W. The role of S100 proteins and their receptor RAGE in pancreatic cancer. Biochim. Biophys. Acta 2015, 1852, 2706–2711. [Google Scholar] [CrossRef]
- Ohuchida, K.; Mizumoto, K.; Miyasaka, Y.; Yu, J.; Cui, L.; Yamaguchi, H.; Toma, H.; Takahata, S.; Sato, N.; Nagai, E.; et al. Over-expression of S100A2 in pancreatic cancer correlates with progression and poor prognosis. J. Pathol. 2007, 213, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Biankin, A.V.; Kench, J.G.; Colvin, E.K.; Segara, D.; Scarlett, C.J.; Nguyen, N.Q.; Chang, D.K.; Morey, A.L.; Lee, C.-S.; Pinese, M.; et al. Expression of S100A2 calcium-binding protein predicts response to pancreatectomy for pancreatic cancer. Gastroenterology 2009, 137, 558–568, 568.e551–511. [Google Scholar] [CrossRef] [PubMed]
- Ohuchida, K.; Mizumoto, K.; Ishikawa, N.; Fujii, K.; Konomi, H.; Nagai, E.; Yamaguchi, K.; Tsuneyoshi, M.; Tanaka, M. The role of S100A6 in pancreatic cancer development and its clinical implication as a diagnostic marker and therapeutic target. Clin. Cancer Res. 2005, 11, 7785–7793. [Google Scholar] [CrossRef]
- Vimalachandran, D.; Greenhalf, W.; Thompson, C.; Luttges, J.; Prime, W.; Campbell, F.; Dodson, A.; Watson, R.; Crnogorac-Jurcevic, T.; Lemoine, N.; et al. High nuclear S100A6 (calcyclin) is significantly associated with poor survival in pancreatic cancer patients. Cancer Res. 2005, 65, 3218–3225. [Google Scholar] [CrossRef]
- Ohuchida, K.; Mizumoto, K.; Ohhashi, S.; Yamaguchi, H.; Konomi, H.; Nagai, E.; Yamaguchi, K.; Tsuneyoshi, M.; Tanaka, M. S100A11, a putative tumor suppressor gene, is overexpressed in pancreatic carcinogenesis. Clin. Cancer Res. 2006, 12, 5417–5422. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.-B.; Jiang, F.; Ni, W.-K.; Chen, B.-Y.; Lu, C.-H.; Li, X.-Y.; Ni, R.-Z. High expression of S100A11 in pancreatic adenocarcinoma is an unfavorable prognostic marker. Med. Oncol. 2012, 29, 1886–1891. [Google Scholar] [CrossRef]
- Ohuchida, K.; Mizumoto, K.; Egami, T.; Yamaguchi, H.; Fujii, K.; Konomi, H.; Nagai, E.; Yamaguchi, K.; Tsuneyoshi, M.; Tanaka, M. S100P is an early developmental marker of pancreatic carcinogenesis. Clin. Cancer Res. 2006, 12, 5411–5416. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhang, Q.; Huang, C.; Shen, Y.; Chen, X.; Shi, X.; Tang, W. Diagnostic value of S100P for pancreatic cancer: A meta-analysis. Tumour Biol. 2014, 35, 9479–9485. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zheng, J.; Huang, Y.; Song, L.; Yin, Y.; Ou, D.; He, S.; Chen, X.; Ouyang, X. Impact of S100A4 expression on clinicopathological characteristics and prognosis in pancreatic cancer: A meta-analysis. Dis. Markers 2016, 2016, 8137378. [Google Scholar] [CrossRef]
- Ikenaga, N.; Ohuchida, K.; Mizumoto, K.; Yu, J.; Fujita, H.; Nakata, K.; Ueda, J.; Sato, N.; Nagai, E.; Tanaka, M. S100A4 mRNA is a diagnostic and prognostic marker in pancreatic carcinoma. J. Gastrointest. Surg. 2009, 13, 1852–1858. [Google Scholar] [CrossRef]
- Tsukamoto, N.; Egawa, S.; Akada, M.; Abe, K.; Saiki, Y.; Kaneko, N.; Yokoyama, S.; Shima, K.; Yamamura, A.; Motoi, F.; et al. The expression of S100A4 in human pancreatic cancer is associated with invasion. Pancreas 2013, 42, 1027–1033. [Google Scholar] [CrossRef]
- Rosty, C.; Ueki, T.; Argani, P.; Jansen, M.; Yeo, C.J.; Cameron, J.L.; Hruban, R.H.; Goggins, M. Overexpression of S100A4 in Pancreatic ductal adenocarcinomas is associated with poor differentiation and DNA hypomethylation. Am. J. Pathol. 2002, 160, 45–50. [Google Scholar] [CrossRef]
- Tabata, T.; Tsukamoto, N.; Fooladi, A.A.I.; Yamanaka, S.; Furukawa, T.; Ishida, M.; Sato, D.; Gu, Z.; Nagase, H.; Egawa, S.; et al. RNA interference targeting against S100A4 suppresses cell growth and motility and induces apoptosis in human pancreatic cancer cells. Biochem. Biophys. Res. Commun. 2009, 390, 475–480. [Google Scholar] [CrossRef]
- Sekine, H.; Chen, N.; Sato, K.; Saiki, Y.; Yoshino, Y.; Umetsu, Y.; Jin, G.; Nagase, H.; Gu, Z.; Fukushige, S.; et al. S100A4, frequently overexpressed in various human cancers, accelerates cell motility in pancreatic cancer cells. Biochem. Biophys. Res. Commun. 2012, 429, 214–219. [Google Scholar] [CrossRef]
- Mahon, P.C.; Baril, P.; Bhakta, V.; Chelala, C.; Caulee, K.; Harada, T.; Lemoine, N.R. S100A4 contributes to the suppression of BNIP3 expression, chemoresistance, and inhibition of apoptosis in pancreatic cancer. Cancer Res. 2007, 67, 6786–6795. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Su, B.; Xie, C.; Wei, S.; Zhou, Y.; Liu, H.; Dai, W.; Cheng, P.; Wang, F.; Xu, X.; et al. Sonic hedgehog-Gli1 signaling pathway regulates the epithelial mesenchymal transition (EMT) by mediating a new target gene, S100A4, in pancreatic cancer cells. PLoS ONE 2014, 9, e96441. [Google Scholar] [CrossRef] [PubMed]
- Che, P.; Yang, Y.; Han, X.; Hu, M.; Sellers, J.C.; Londono-Joshi, A.I.; Cai, G.-Q.; Buchsbaum, D.J.; Christein, J.D.; Tang, Q.; et al. S100A4 promotes pancreatic cancer progression through a dual signaling pathway mediated by Src and focal adhesion kinase. Sci. Rep. 2015, 5, 8453. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.A.; Vimalachandran, D.; Thompson, C.C.; Jenkins, R.E.; Nedjadi, T.; Shekouh, A.; Campbell, F.; Dodson, A.; Prime, W.; Crnogorac-Jurcevic, T.; et al. The expression of S100A8 in pancreatic cancer-associated monocytes is associated with the Smad4 status of pancreatic cancer cells. Proteomics 2007, 7, 1929–1940. [Google Scholar] [CrossRef]
- Ang, C.W.; Nedjadi, T.; Sheikh, A.A.; Tweedle, E.M.; Tonack, S.; Honap, S.; Jenkins, R.E.; Park, B.K.; Schwarte-Waldhoff, I.; Khattak, I.; et al. Smad4 loss is associated with fewer S100A8-positive monocytes in colorectal tumors and attenuated response to S100A8 in colorectal and pancreatic cancer cells. Carcinogenesis 2010, 31, 1541–1551. [Google Scholar] [CrossRef]
- Hermani, A.; Hess, J.; DeServi, B.; Medunjanin, S.; Grobholz, R.; Trojan, L.; Angel, P.; Mayer, D. Calcium-binding proteins S100A8 and S100A9 as novel diagnostic markers in human prostate cancer. Clin. Cancer Res. 2005, 11, 5146–5152. [Google Scholar] [CrossRef]
- Hermani, A.; Servi, B.D.; Medunjanin, S.; Tessier, P.A.; Mayer, D. S100A8 and S100A9 activate MAP kinase and NF-kappaB signaling pathways and trigger translocation of RAGE in human prostate cancer cells. Exp. Cell Res. 2006, 312, 184–197. [Google Scholar] [CrossRef]
- Rehman, I.; Cross, S.S.; Azzouzi, A.-R.; Catto, J.W.F.; Deloulme, J.C.; Larre, S.; Champigneuille, J.; Fromont, G.; Cussenot, O.; Hamdy, F.C. S100A6 (Calcyclin) is a prostate basal cell marker absent in prostate cancer and its precursors. Br. J. Cancer 2004, 91, 739–744. [Google Scholar] [CrossRef]
- Yao, R.; Lopez-Beltran, A.; Maclennan, G.T.; Montironi, R.; Eble, J.N.; Cheng, L. Expression of S100 Protein family members in the pathogenesis of bladder tumors. Anticancer Res. 2007, 27, 3051–3058. [Google Scholar]
- Ha, Y.-S.; Kim, M.-J.; Yoon, H.-Y.; Kang, H.-W.; Kim, Y.-J.; Yun, S.-J.; Lee, S.-C.; Kim, W.-J. mRNA expression of S100A8 as a prognostic marker for progression of non-muscle-invasive bladder cancer. Korean J. Urol. 2010, 51, 15–20. [Google Scholar] [CrossRef]
- Kim, S.-K.; Kim, E.-J.; Leem, S.-H.; Ha, Y.-S.; Kim, Y.-J.; Kim, W.-J. Identification of S100A8-correlated genes for prediction of disease progression in non-muscle invasive bladder cancer. BMC Cancer 2010, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Tsunoda, T.; Song, Y.; Yokomizo, A.; Tada, Y.; Oda, Y.; Naito, S. Enhanced S100 calcium-binding protein P expression sensitizes human bladder cancer cells to cisplatin. BJU Int. 2011, 107, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Ro, S.W.; Seo, S.H.; Jeon, Y.; Moon, H.; Kim, D.Y.; Kim, S.U. Genetically engineered mouse models for liver cancer. Cancers 2019, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Zhu, H.; Wang, W.; Zhang, S.; Zhang, Y.; Mao, G. Abnormal expression of EMT-related proteins, S100A4, vimentin and E-cadherin, is correlated with clinicopathological features and prognosis in HCC. Med. Oncol. 2014, 31, 970. [Google Scholar] [CrossRef]
- Jiao, J.; González, Á.; Stevenson, H.L.; Gagea, M.; Sugimoto, H.; Kalluri, R.; Beretta, L. Depletion of S100A4+ stromal cells does not prevent HCC development but reduces the stem cell-like phenotype of the tumors. Exp. Mol. Med. 2018, 50, 422. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Song, K.; Liu, S.; Zhang, H.; Wang, F.; Ni, C.; Zhai, W.; Liang, J.; Qin, Z.; et al. S100A4 promotes hepatocellular carcinogenesis by intensifying fibrosis-associated cancer cell stemness. Oncoimmunology 2020, 9, 1725355. [Google Scholar] [CrossRef]
- Zhao, F.-T.; Jia, Z.-S.; Yang, Q.; Song, L.; Jiang, X.-J. S100A14 promotes the growth and metastasis of hepatocellular carcinoma. Asian Pac. J. Cancer Prev. 2013, 14, 3831–3836. [Google Scholar] [CrossRef]
- Mohamed, B.F.; Serag, W.M.; Abdelal, R.M.; Elsergany, H.F. S100A14 protein as diagnostic and prognostic marker in hepatocellular carcinoma. Egypt. Liver J. 2019, 9. [Google Scholar] [CrossRef]
- Raffat, M.A.; Hadi, N.I.; Hosein, M.; Mirza, S.; Ikram, S.; Akram, Z. S100 proteins in oral squamous cell carcinoma. Clin. Chim. Acta 2018, 480, 143–149. [Google Scholar] [CrossRef]
- Sapkota, D.; Bruland, O.; Costea, D.E.; Haugen, H.; Vasstrand, E.N.; Ibrahim, S.O. S100A14 regulates the invasive potential of oral squamous cell carcinoma derived cell-lines in vitro by modulating expression of matrix metalloproteinases, MMP1 and MMP9. Eur. J. Cancer 2011, 47, 600–610. [Google Scholar] [CrossRef]
- Sapkota, D.; Costea, D.E.; Blø, M.; Bruland, O.; Lorens, J.B.; Vasstrand, E.N.; Ibrahim, S.O. S100A14 inhibits proliferation of oral carcinoma derived cells through G1-arrest. Oral Oncol. 2012, 48, 219–225. [Google Scholar] [CrossRef]
- White, R.; Pulford, E.; Elliot, D.J.; Thurgood, L.A.; Klebe, S. Quantitative mass spectrometry to identify protein markers for diagnosis of malignant pleural mesothelioma. J. Proteom. 2019, 192, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Nader, J.S.; Guillon, J.; Petit, C.; Boissard, A.; Franconi, F.; Blandin, S.; Lambot, S.; Grégoire, M.; Verrièle, V.; Nawrocki-Raby, B.; et al. S100A4 is a biomarker of tumorigenesis, EMT, invasion, and colonization of host organs in experimental malignant mesothelioma. Cancers 2020, 12, 939. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.F.; Kong, L.Y.; Jordan, J.; Conrad, C.; Madden, T.; Fokt, I.; Priebe, W.; Heimberger, A.B. A novel small molecule inhibitor of signal transducers and activators of transcription 3 reverses immune tolerance in malignant glioma patients. Cancer Res. 2007, 67, 9630–9636. [Google Scholar] [CrossRef] [PubMed]
- Kostianovsky, A.M.; Maier, L.M.; Anderson, R.C.; Bruce, J.N.; Anderson, D.E. Astrocytic regulation of human monocytic/microglial activation. J. Immunol. 2008, 181, 5425–5432. [Google Scholar] [CrossRef]
- Hwang, J.S.; Jung, E.H.; Kwon, M.Y.; Han, I.O. Glioma-secreted soluble factors stimulate microglial activation: The role of interleukin-1β and tumor necrosis factor-α. J. Neuroimmunol. 2016, 298, 165–171. [Google Scholar] [CrossRef]
- Yang, I.; Han, S.J.; Kaur, G.; Crane, C.; Parsa, A.T. The role of microglia in central nervous system immunity and glioma immunology. J. Clin. Neurosci. 2010, 17, 6–10. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, W.; Alizadeh, D.; Zhao, D.; Farrukh, O.; Lin, J.; Badie, S.A.; Badie, B. S100B attenuates microglia activation in gliomas: Possible role of STAT3 pathway. Glia 2011, 59, 486–498. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, L.; Zhang, I.Y.; Chen, X.; Da Fonseca, A.; Wu, S.; Ren, H.; Badie, S.; Sadeghi, S.; Ouyang, M.; et al. S100B promotes glioma growth through chemoattraction of myeloid-derived macrophages. Clin. Cancer Res. 2013, 19, 3764–3775. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, I.Y.; Zhang, L.; Song, Y.; Liu, S.; Ren, H.; Liu, H.; Zhou, H.; Su, Y.; Yang, Y.; et al. S100B suppression alters polarization of infiltrating myeloid-derived cells in gliomas and inhibits tumor growth. Cancer Lett. 2018, 439, 91–100. [Google Scholar] [CrossRef]
- Holla, F.K.; Postma, T.J.; Blankenstein, M.A.; van Mierlo, T.J.M.; Vos, M.J.; Sizoo, E.M.; de Groot, M.; Uitdehaag, B.M.J.; Buter, J.; Klein, M.; et al. Prognostic value of the S100B protein in newly diagnosed and recurrent glioma patients: A serial analysis. J. Neuro-Oncol. 2016, 129, 525–532. [Google Scholar] [CrossRef]
- Gonzalez, L.L.; Garrie, K.; Turner, M.D. Role of S100 proteins in health and disease. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118677. [Google Scholar] [CrossRef]
- Sack, U.; Walther, W.; Scudiero, D.; Selby, M.; Aumann, J.; Lemos, C.; Fichtner, I.; Schlag, P.M.; Shoemakere, R.H.; Stein, U. S100A4-induced cell motility and metastasis is restricted by the Wnt/β-catenin pathway inhibitor calcimycin in colon cancer cells. Mol. Biol. Cell 2011, 22, 3344–3354. [Google Scholar] [CrossRef]
- Stein, U.; Arlt, F.; Smith, J.; Sack, U.; Herrmann, P.; Walther, W.; Lemm, M.; Fichtner, I.; Shoemaker, R.H.; Schlag, P.M. Intervening in β-catenin signaling by sulindac inhibits S100A4-dependent colon cancer metastasis. Neoplasia 2011, 13, 131–144. [Google Scholar] [CrossRef]
- Sack, U.; Walther, W.; Scudiero, D.; Selby, M.; Kobelt, D.; Lemm, M.; Fichtner, I.; Schlag, P.M.; Shoemaker, R.H.; Stein, U. Novel effect of antihelminthic Niclosamide on S100A4-mediated metastatic progression in colon cancer. J. Natl. Cancer Inst. 2011, 103, 1018–1036. [Google Scholar] [CrossRef]
- Burock, S.; Daum, S.; Keilholz, U.; Neumann, K.; Walther, W.; Stein, U. Phase II trial to investigate the safety and efficacy of orally applied niclosamide in patients with metachronous or sychronous metastases of a colorectal cancer progressing after therapy: The NIKOLO trial. BMC Cancer 2018, 18, 297. [Google Scholar] [CrossRef]
- Garrett, S.C.; Hodgson, L.; Rybin, A.; Toutchkine, A.; Hahn, K.M.; Lawrence, D.S.; Bresnick, A.R. A biosensor of S100A4 metastasis factor activation: Inhibitor screening and cellular activation dynamics. Biochemistry 2008, 47, 986–996. [Google Scholar] [CrossRef][Green Version]
- Malashkevich, V.N.; Dulyaninova, N.G.; Ramagopal, U.A.; Liriano, M.A.; Varney, K.M.; Knight, D.; Brenowitz, M.; Weber, D.J.; Almo, S.C.; Bresnick, A.R. Phenothiazines inhibit S100A4 function by inducing protein oligomerization. Proc. Natl. Acad. Sci. USA 2010, 107, 8605–8610. [Google Scholar] [CrossRef]
- Raymond, E.; Dalgleish, A.; Damber, J.-E.; Smith, M.; Pili, R. Mechanisms of action of tasquinimod on the tumour microenvironment. Cancer Chemother. Pharmacol. 2014, 73, 1–8. [Google Scholar] [CrossRef]
- Shen, L.; Sundstedt, A.; Ciesielski, M.; Miles, K.M.; Celander, M.; Adelaiye, R.; Orillion, A.; Ciamporcero, E.; Ramakrishnan, S.; Ellis, L.; et al. Tasquinimod modulates suppressive myeloid cells and enhances cancer immunotherapies in murine models. Cancer Immunol. Res. 2015, 3, 136–148. [Google Scholar] [CrossRef]
- Shen, L.; Pili, R. Tasquinimod targets suppressive myeloid cells in the tumor microenvironment. Oncoimmunology 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, C.; Armstrong, A.; Pili, R.; Ng, S.; Huddart, R.; Agarwal, N.; Khvorostenko, D.; Lyulko, O.; Brize, A.; Vogelzang, N.; et al. Randomized, double-blind, placebo-controlled phase III study of tasquinimod in men with metastatic castration-resistant prostate cancer. J. Clin. Oncol. 2016, 34, 2636–2643. [Google Scholar] [CrossRef]
- Reddy, T.R.K.; Li, C.; Fischer, P.M.; Dekker, L.V. Three-dimensional pharmacophore design and biochemical screening identifies substituted 1,2,4-triazoles as inhibitors of the annexin A2-S100A10 protein interaction. ChemMedChem 2012, 7, 1435–1446. [Google Scholar] [CrossRef]
- Reddy, T.R.K.; Li, C.; Guo, X.; Fischer, P.M.; Dekker, L.V. Design, synthesis and SAR exploration of tri-substituted 1,2,4-triazoles as inhibitors of the annexin A2-S100A10 protein interaction. Bioorganic Med. Chem. 2014, 22, 5378–5391. [Google Scholar] [CrossRef]
- Markowitz, J.; Chen, I.; Gitti, R.; Baldisseri, D.M.; Pan, Y.; Udan, R.; Carrier, F.; MacKerell, A.D.; Weber, D.J. Identification and characterization of small molecule inhibitors of the calcium-dependent S100B-p53 tumor suppressor interaction. J. Med. Chem. 2004, 47, 5085–5093. [Google Scholar] [CrossRef]
- Smith, J.; Stewart, B.J.; Glaysher, S.; Peregrin, K.; Knight, L.A.; Weber, D.J.; Cree, I.A. The effect of pentamidine on melanoma ex vivo. Anti-Cancer Drugs 2010, 21, 181–185. [Google Scholar] [CrossRef]
- Mehnert, J.M.; Hausner, P.F.; Tan, M.; Weber, D.; Sausville, E.A. Treatment of melanoma with wild-type p53 (wtp53) and detectable S100B using pentamidine: A phase II trial with correlative biomarker endpoints. J. Clin. Oncol. 2010, 28, TPS310. [Google Scholar] [CrossRef]
- Arumugam, T.; Ramachandran, V.; Logsdon, C.D. Effect of cromolyn on S100P interactions with RAGE and pancreatic cancer growth and invasion in mouse models. J. Natl. Cancer Inst. 2006, 98, 1806–1818. [Google Scholar] [CrossRef]
- Penumutchu, S.R.; Chou, R.-H.; Yu, C. Interaction between S100P and the anti-allergy drug cromolyn. Biochem. Biophys. Res. Commun. 2014, 454, 404–409. [Google Scholar] [CrossRef]
- Arumugam, T.; Ramachandran, V.; Sun, D.; Peng, Z.; Pal, A.; Maxwell, D.S.; Bornmann, W.G.; Logsdon, C.D. Designing and developing S100P inhibitor 5-methyl cromolyn for pancreatic cancer therapy. Mol. Cancer Ther. 2013, 12, 654–662. [Google Scholar] [CrossRef]
- Klingelhöfer, J.; Grum-Schwensen, B.; Beck, M.K.; Knudsen, R.S.P.; Grigorian, M.; Lukanidin, E.; Ambartsumian, N. Anti-S100A4 antibody suppresses metastasis formation by blocking stroma cell invasion. Neoplasia 2012, 14, 1260–1268. [Google Scholar] [CrossRef] [PubMed]
- Grum-Schwensen, B.; Klingelhöfer, J.; Beck, M.; Bonefeld, C.M.; Hamerlik, P.; Guldberg, P.; Grigorian, M.; Lukanidin, E.; Ambartsumian, N. S100A4-neutralizing antibody suppresses spontaneous tumor progression, pre-metastatic niche formation and alters T-cell polarization balance. BMC Cancer 2015, 15, 44. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.L.; Padilla, L.; Dakhel, S.; Coll, T.; Hervas, R.; Adan, J.; Masa, M.; Mitjans, F.; Martinez, J.M.; Coma, S.; et al. Therapeutic targeting of tumor growth and angiogenesis with a novel anti-S100A4 monoclonal antibody. PLoS ONE 2013, 8, e72480. [Google Scholar] [CrossRef] [PubMed]
- Padilla, L.; Dakhel, S.; Adan, J.; Masa, M.; Martinez, J.M.; Roque, L.; Coll, T.; Hervas, R.; Calvis, C.; Llinas, L.; et al. S100A7: From mechanism to cancer therapy. Oncogene 2017, 36, 6749–6761. [Google Scholar] [CrossRef]
- Kinoshita, R.; Sato, H.; Yamauchi, A.; Takahashi, Y.; Inoue, Y.; Sumardika, I.W.; Chen, Y.; Tomonobu, N.; Araki, K.; Shien, K.; et al. Newly developed anti-S100A8/A9 monoclonal antibody efficiently prevents lung tropic cancer metastasis. Int. J. Cancer 2019, 145, 569–575. [Google Scholar] [CrossRef]
- Dakhel, S.; Padilla, L.; Adan, J.; Masa, M.; Martinez, J.M.; Roque, L.; Coll, T.; Hervas, R.; Calvis, C.; Messeguer, R.; et al. S100P antibody-mediated therapy as a new promising strategy for the treatment of pancreatic cancer. Oncogenesis 2014, 3, 92. [Google Scholar] [CrossRef]
- Dou, C.; Liu, Z.; Xu, M.; Jia, Y.; Wang, Y.; Li, Q.; Yang, W.; Zheng, X.; Tu, K.; Liu, Q. miR-187-3p inhibits the metastasis and epithelial-mesenchymal transition of hepatocellular carcinoma by targeting S100A4. Cancer Lett. 2016, 381, 380–390. [Google Scholar] [CrossRef]
- Yang, D.; Du, G.; Xu, A.; Xi, X.; Li, D. Expression of miR-149-3p inhibits proliferation, migration, and invasion of bladder cancer by targeting S100A4. Am. J. Cancer Res. 2017, 7, 2209–2219. [Google Scholar]
- Li, P.; Lv, X.; Zhang, Z.; Xie, S. S100A6/miR193a regulates the proliferation, invasion, migration and angiogenesis of lung cancer cells through the P53 acetylation. Am. J. Transl. Res. 2019, 11, 4634–4649. [Google Scholar]
- Fan, F.; Lu, J.; Yu, W.; Zhang, Y.; Xu, S.; Pang, L.; Zhu, B. MicroRNA-26b-5p regulates cell proliferation, invasion and metastasis in human intrahepatic cholangiocarcinoma by targeting S100A7. Oncol. Lett. 2018, 15, 386–392. [Google Scholar] [CrossRef]
- Guo, Y.; Fu, W.; Chen, H.; Shang, C.; Zhong, M. miR-24 functions as a tumor suppressor in Hep2 laryngeal carcinoma cells partly through down-regulation of the S100A8 protein. Oncol. Rep. 2012, 27, 1097–1103. [Google Scholar] [CrossRef]
- Lang, B.; Shang, C.; Meng, L. Targeted silencing of S100A8 gene by miR-24 to increase chemotherapy sensitivity of endometrial carcinoma cells to paclitaxel. Med. Sci. Monit. 2016, 22, 1953–1958. [Google Scholar] [CrossRef]
- Lv, H.; Hou, H.; Lei, H.; Nie, C.; Chen, B.; Bie, L.; Han, L.; Chen, X. MicroRNA-6884-5p regulates the proliferation, invasion and EMT of gastric cancer cells by directly targeting S100A16. Oncol. Res. 2019, 28, 225–236. [Google Scholar] [CrossRef]
- Zhong, S.; Jeong, J.-H.; Chen, Z.; Chen, Z.; Luo, J.-L. Targeting tumor microenvironment by small-molecule inhibitors. Transl. Oncol. 2020, 13, 57–69. [Google Scholar] [CrossRef]
- Lavanya, V.; Adil, M.; Ahmed, N.; Rishi, A.K.; Jamal, S. Small molecule inhibitors as emerging cancer therapeutics. Integr. Cancer Sci. Therap. 2014, 1, 39–46. [Google Scholar] [CrossRef]
- Bresnick, A.R. S100 proteins as therapeutic targets. Biophys. Rev. 2018, 10, 1617–1629. [Google Scholar] [CrossRef]
- Gasser, M.; Waaga-Gasser, A.M. Therapeutic antibodies in cancer therapy. Adv. Exp. Med. Biol. 2016, 917, 95–120. [Google Scholar] [CrossRef]
- Lu, R.-M.; Hwang, Y.-C.; Liu, I.-J.; Lee, C.-C.; Tsai, H.-Z.; Li, H.-J.; Wu, H.-C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1. [Google Scholar] [CrossRef]
- Marrocco, I.; Romaniello, D.; Yarden, Y. Cancer immunotherapy: The dawn of antibody cocktails. Methods Mol. Biol. 2019, 1904, 11–51. [Google Scholar] [CrossRef]
- Almagro, J.C.; Daniels-Wells, T.R.; Perez-Tapia, S.M.; Penichet, M.L. Progress and challenges in the design and clinical development of antibodies for cancer therapy. Front. Immunol. 2017, 8, 1751. [Google Scholar] [CrossRef]
- Cruz, E.; Kayser, V. Monoclonal antibody therapy of solid tumors: Clinical limitations and novel strategies to enhance treatment efficacy. Biol. Targets Ther. 2019, 13, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Dufresne, S.; Rébillard, A.; Muti, P.; Friedenreich, C.M.; Brenner, D.R. A review of physical activity and circulating mirna expression: Implications in cancer risk and progression. Cancer Epidemiol. Biomark. Prev. 2018, 27, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Ors-Kumoglu, G.; Gulce-Iz, S.; Biray-Avci, C. Therapeutic microRNAs in human cancer. Cytotechnology 2019, 71, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Ganju, A.; Khan, S.; Hafeez, B.B.; Behrman, S.W.; Yallapu, M.M.; Chauhan, S.C.; Jaggi, M. miRNA nanotherapeutics for cancer. Drug Discov. Today 2017, 22, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Wright, N.T.; Cannon, B.R.; Zimmer, D.B.; Weber, D.J. S100A1: Structure, function, and therapeutic potential. Curr. Chem. Biol. 2009, 3, 138–145. [Google Scholar] [CrossRef]
- Kizawa, K.; Takahara, H.; Unno, M.; Heizmann, C.W. S100 and S100 fused-type protein families in epidermal maturation with special focus on S100A3 in mammalian hair cuticles. Biochimie 2011, 93, 2038–2047. [Google Scholar] [CrossRef]
- Kizawa, K.; Fujimori, T.; Kawai, T. Arachidonate 12-lipoxygenase inhibitors promote S100A3 citrullination in cultured SW480 cells and isolated hair follicles. Biol. Pharm. Bull. 2017, 40, 516–523. [Google Scholar] [CrossRef]
- Bertini, I.; Das Gupta, S.; Hu, X.; Karavelas, T.; Luchinat, C.; Parigi, G.; Yuan, J. Solution structure and dynamics of S100A5 in the apo and Ca2+-bound states. J. Biol. Inorg. Chem. 2009, 14, 1097–1107. [Google Scholar] [CrossRef]
- Jurewicz, E.; Wyroba, E.; Filipek, A. Tubulin-dependent secretion of S100A6 and cellular signaling pathways activated by S100A6-integrin β1 interaction. Cell. Signal. 2018, 42, 21–29. [Google Scholar] [CrossRef]
- Rosenberger, S.; Thorey, I.S.; Werner, S.; Boukamp, P. A novel regulator of telomerase. S100A8 mediates differentiation-dependent and calcium-induced inhibition of telomerase activity in the human epidermal keratinocyte line HaCaT. J. Biol. Chem. 2007, 282, 6126–6135. [Google Scholar] [CrossRef]
- Pietzsch, J.; Hoppmann, S. Human S100A12: A novel key player in inflammation? Amino Acids 2009, 36, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Tao, T.; Raftery, M.J.; Youssef, P.; Girolamo, N.D.; Geczy, C.L. Proinflammatory properties of the human S100 protein S100A12. J. Leukoc. Biol. 2001, 69, 986–994. [Google Scholar] [PubMed]
- Bagheri, V. S100A12: Friend or foe in pulmonary tuberculosis? Cytokine 2017, 92, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Ridinger, K.; Schäfer, B.W.; Durussel, I.; Cox, J.A.; Heizmann, C.W. S100A13–biochemical characterization and subcellular localization in different cell lines. J. Biol. Chem. 2000, 275, 8686–8694. [Google Scholar] [CrossRef] [PubMed]
- Massi, D.; Landriscina, M.; Piscazzi, A.; Cosci, E.; Kirov, A.; Paglierani, M.; Di Serio, C.; Mourmouras, V.; Fumagalli, S.; Biagioli, M.; et al. S100A13 is a new angiogenic marker in human melanoma. Mod. Pathol. 2010, 23, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.K.; Yu, C. The IL1alpha-S100A13 heterotetrameric complex structure: A component in the non-classical pathway for interleukin 1alpha secretion. J. Biol. Chem. 2011, 286, 14608–14617. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Chen, D.; Sung, M.-T.; Chen, C.-J.; Wang, T.-Y.; Lin, Y.-Y.; Lin, M.-C.; Chang, H.-C.; Chen, Y.-M.; Hsiao, C.-C.; et al. Increased S100A15 expression and decreased DNA methylation of its gene promoter are involved in high metastasis potential and poor outcome of lung adenocarcinoma. Oncotarget 2017, 8, 45710–45724. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, R.; Xin, J.; Sun, Y.; Li, J.; Wei, D.; Zhao, A.Z. Identification of S100A16 as a novel adipogenesis promoting factor in 3T3-L1 cells. Endocrinology 2011, 152, 903–911. [Google Scholar] [CrossRef]
- Sturchler, E.; Cox, J.A.; Durussel, I.; Weibel, M.; Heizmann, C.W. S100A16, a novel calcium-binding protein of the EF-hand superfamily. J. Biol. Chem. 2006, 281, 38905–38917. [Google Scholar] [CrossRef]
- Sorci, G.; Riuzzi, F.; Arcuri, C.; Tubaro, C.; Bianchi, R.; Giambanco, I. S100B protein in tissue development, repair and regeneration. World J. Biol. Chem. 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Jiang, H.; Hu, H.; Tong, X.; Jiang, Q.; Zhu, H.; Zhang, S. Calcium-binding protein S100P and cancer: Mechanisms and clinical relevance. J. Cancer Res. Clin. Oncol. 2012, 138, 1–9. [Google Scholar] [CrossRef] [PubMed]



| Inhibitor | S100 Target | Mechanism of Action | Current Status | References |
|---|---|---|---|---|
| Small Molecule Inhibitors | ||||
| Calcimycin | Transcription of S100A4 | Inhibition of β-catenin expression, which inhibits Wnt/β-catenin signalling and therefore expression of S100A4 | Pre-clinical | [214] |
| Sulindac | Transcription of S100A4 | Inhibition of β-catenin expression, which inhibits Wnt/β-catenin signalling and therefore expression of S100A4 | Pre-clinical | [215] |
| Niclosamide | Transcription of S100A4 | Inhibition of β-catenin/TCF complex binding to the S100A4 promoter leading to reduced expression of S100A4 | Phase II for treatment of CRC metastasised patients | [216,217] |
| Trifluoperazine (TFP) | S100A4 | Ca2+-S100A4/TFP dimers form a pentameric ring, disabling interaction between S100A4 and myosinIIA | Pre-clinical | [218,219] |
| Tasqinimod | S100A9 | Binds S100A9 and blocks interaction with RAGE and TLR4 | Phase III completed for treatment of metastatic castration-resistant prostate cancer (mCRPC) | [220,221,222,223] |
| 1,2,4-triazoles | S100A10 | Competes with annexin A2 for the binding to S100A10 | Pre-clinical | [224,225] |
| Pentamidine | S100B | Binds to S100B and inhibits interaction with p53, therefore restoring p53 activity | Phase II for treatment of refractory melanoma | [226,227,228] |
| Cromolyn 5-methyl cromolyn | S100P | Binds to S100P and prevents interaction with RAGE | Pre-clinical | [229,230,231] |
| Antibodies | ||||
| 6B12 | S100A4 | Binds extracellular S100A4 and thereby acts as an immunomodulating agent | Pre-clinical | [232,233] |
| 5C3 | S100A4 | Binds to and neutralises S100A4 | Pre-clinical | [234] |
| 6F5 | S100A7 | Binds to S100A7 and thereby blocks S100A7/RAGE interaction | Pre-clinical | [235] |
| Ab45 | S100A8/S100A9 | Binds extracellular S100A8/S100A9 heterodimer and therefore reduces metastasis | Pre-clinical | [236] |
| 2H8 | S100P | Binds S100P and therefore reduces tumour growth and metastasis | Pre-clinical | [237] |
| miRNA | ||||
| miR-187-3p | S100A4 | S100A4 silencing results in reduced metastasis and EMT in HCC | Pre-clinical | [238] |
| miR-149-3p | S100A4 | S100A4 silencing results in inhibition of invasion and migration in bladder cancer cells | Pre-clinical | [239] |
| miR-193a | S100A6 | S100A6 silencing results in suppression of proliferation, invasion, migration, and angiogenesis within lung cancer models | Pre-clinical | [240] |
| miR-26b-5p | S100A7 | S100A7 silencing inhibits proliferation, invasion, and migration of intrahepatic cholangiocarcinoma cells | Pre-clinical | [241] |
| miR-24 | S100A8 | S100A8 silencing leads to decreased proliferation and invasion of laryngeal carcinoma cells and increases sensitivity of endometrial carcinoma cells for paclitaxel | Pre-clinical | [242,243] |
| miR-6884-5p | S100A16 | S100A16 silencing reduces proliferation, invasion, and EMT in gastric cancer | Pre-clinical | [244] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allgöwer, C.; Kretz, A.-L.; von Karstedt, S.; Wittau, M.; Henne-Bruns, D.; Lemke, J. Friend or Foe: S100 Proteins in Cancer. Cancers 2020, 12, 2037. https://doi.org/10.3390/cancers12082037
Allgöwer C, Kretz A-L, von Karstedt S, Wittau M, Henne-Bruns D, Lemke J. Friend or Foe: S100 Proteins in Cancer. Cancers. 2020; 12(8):2037. https://doi.org/10.3390/cancers12082037
Chicago/Turabian StyleAllgöwer, Chantal, Anna-Laura Kretz, Silvia von Karstedt, Mathias Wittau, Doris Henne-Bruns, and Johannes Lemke. 2020. "Friend or Foe: S100 Proteins in Cancer" Cancers 12, no. 8: 2037. https://doi.org/10.3390/cancers12082037
APA StyleAllgöwer, C., Kretz, A.-L., von Karstedt, S., Wittau, M., Henne-Bruns, D., & Lemke, J. (2020). Friend or Foe: S100 Proteins in Cancer. Cancers, 12(8), 2037. https://doi.org/10.3390/cancers12082037