Radiosensitisation of Hepatocellular Carcinoma Cells by Vandetanib

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
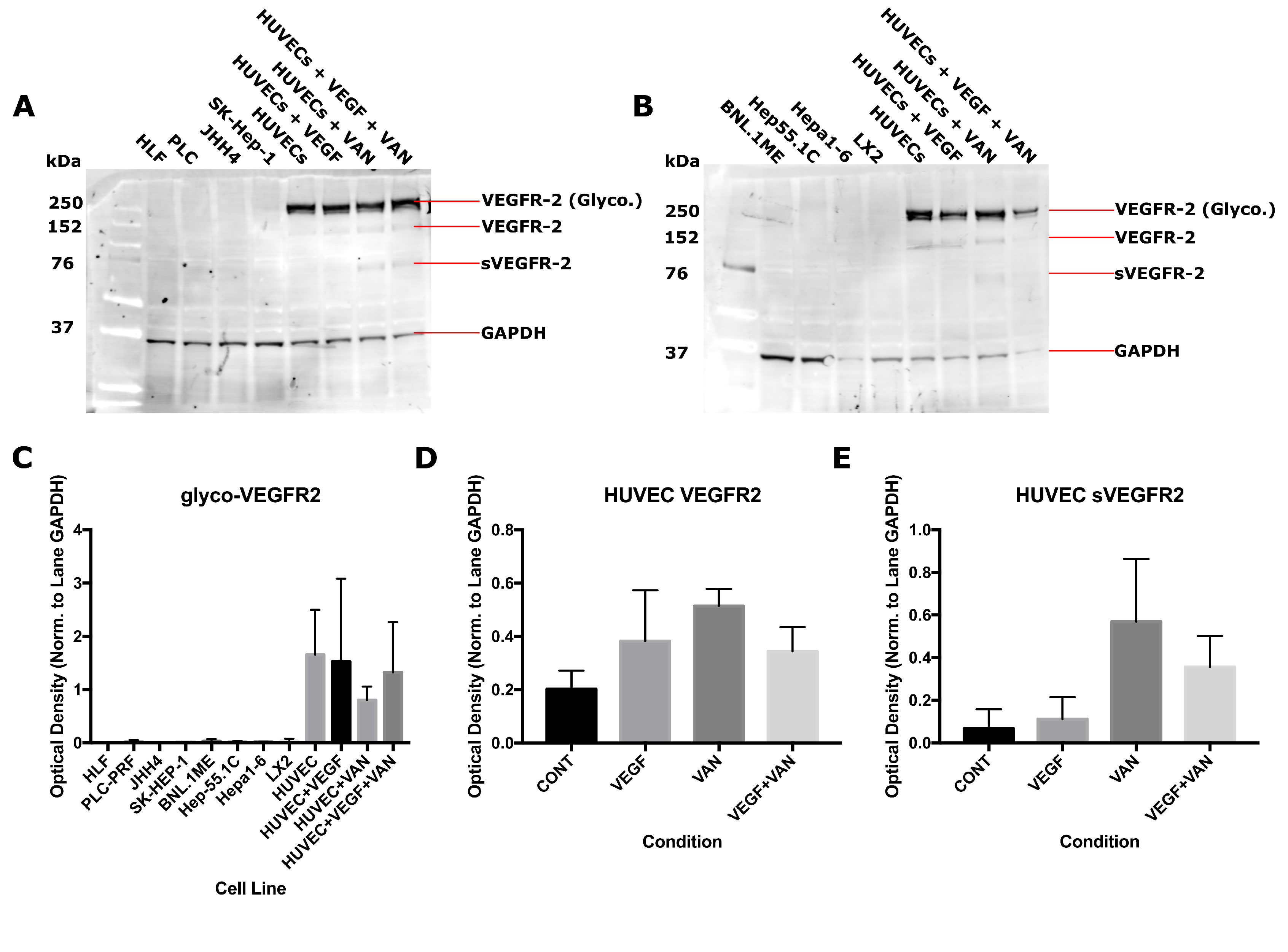
2.1. VEGFR-2 Expression for Human and Mouse Epithelial and Endothelial Cell Lines
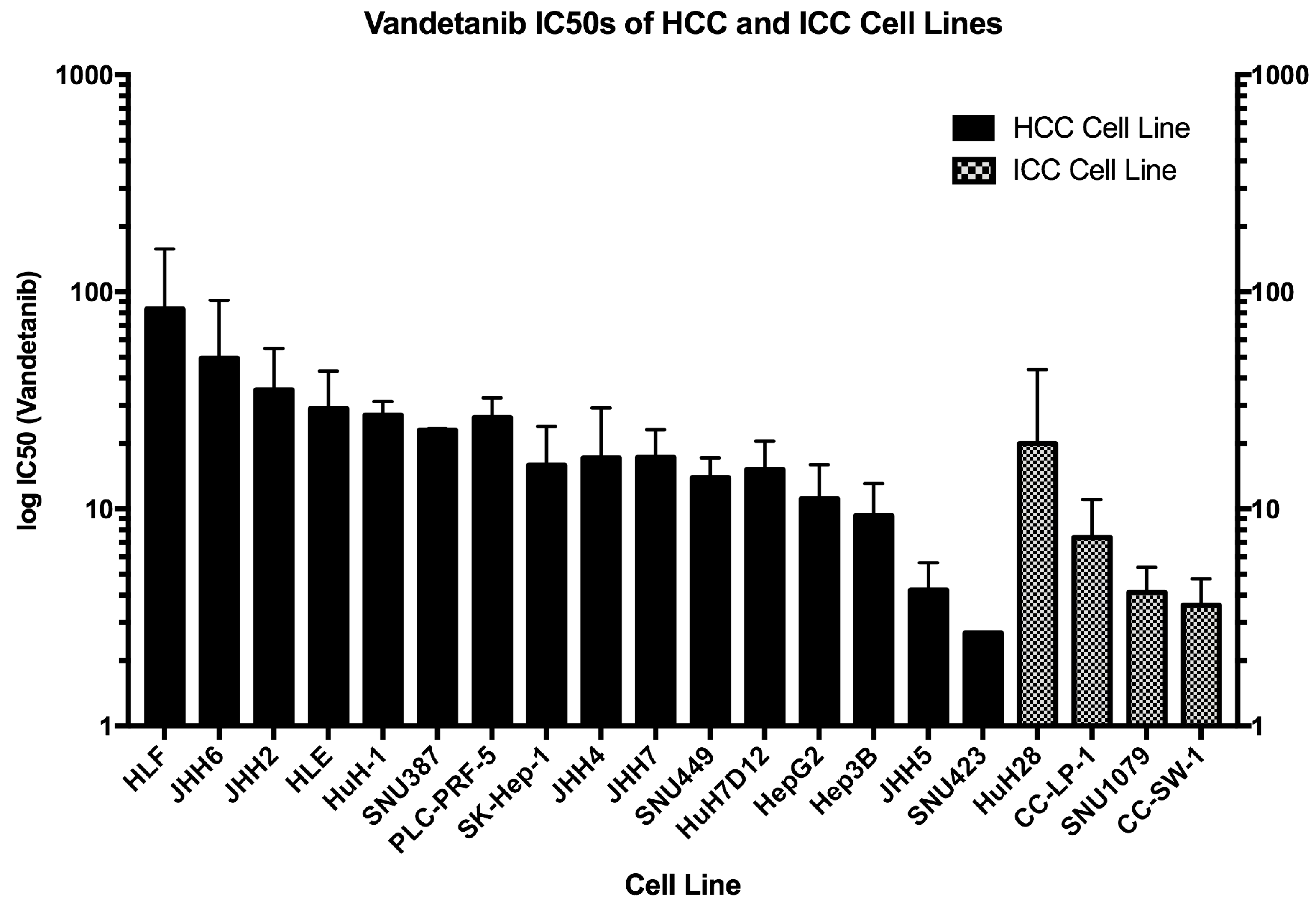
2.2. IC for Vandetanib Across 20 HCC Cell Lines
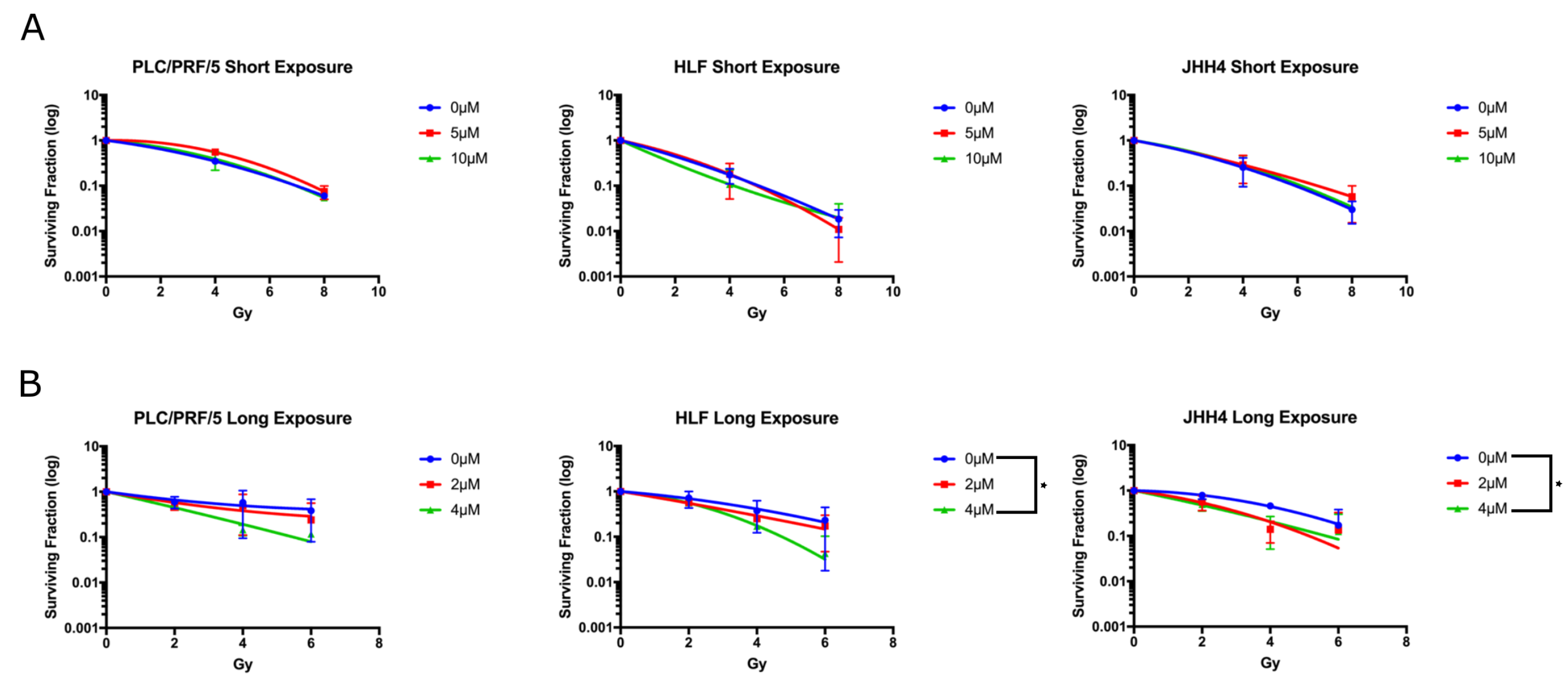
2.3. Long Exposure to Vandetanib Sensitises HCC Cells to Radiation
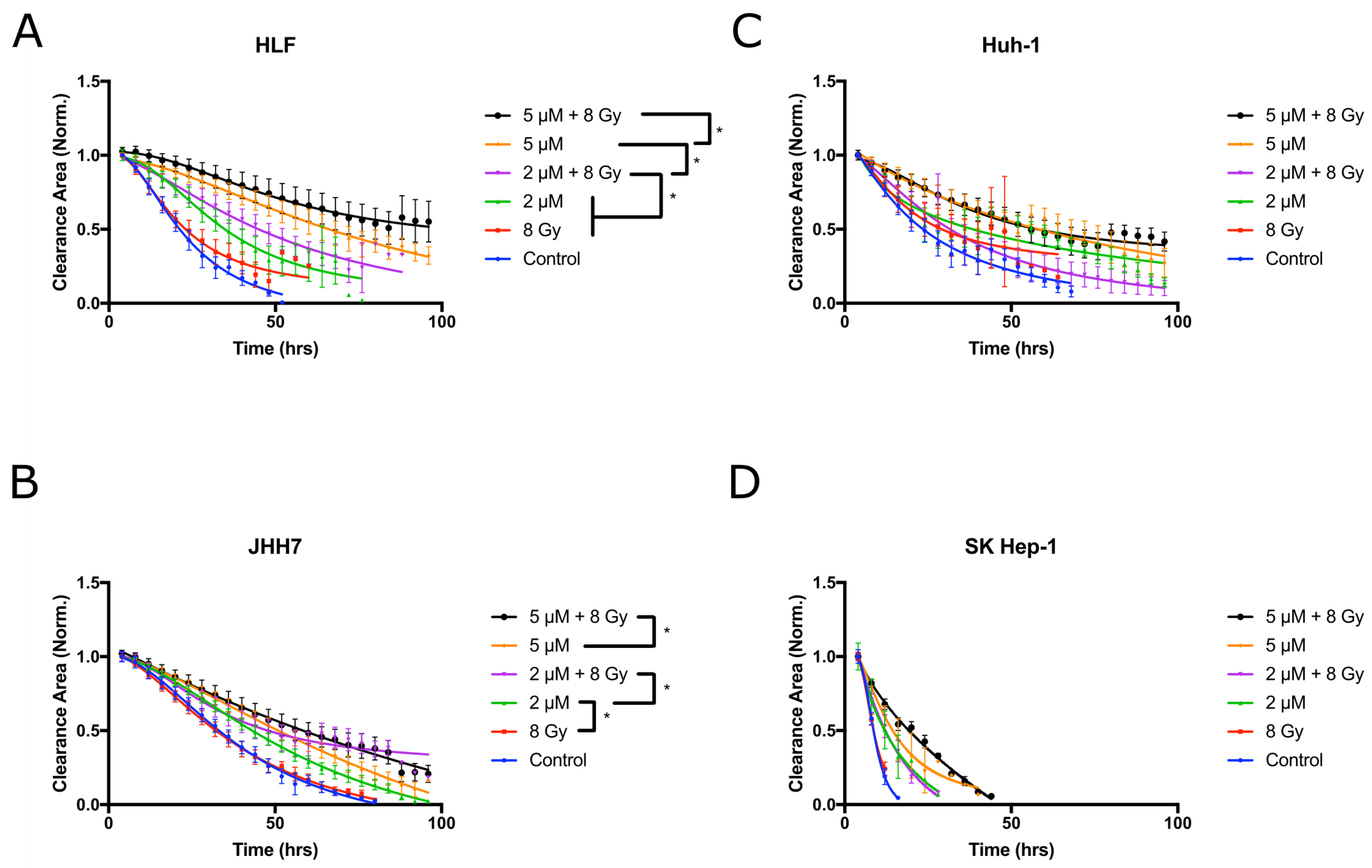
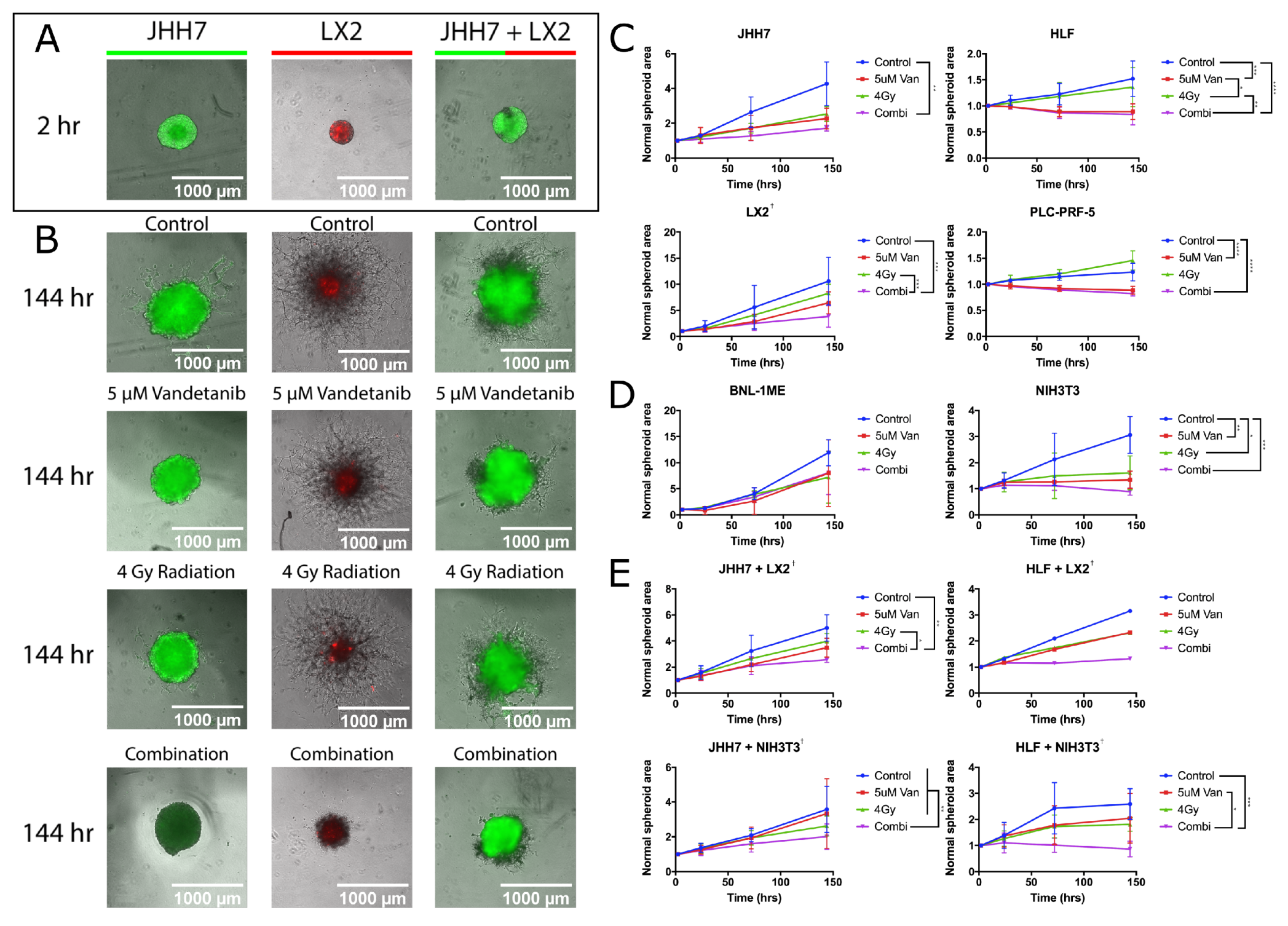
2.4. Vandetanib and Radiation Combine to Inhibit Migration in 2D HCC Cell Culture
2.5. Combined Vandetanib-Radiation Invasion Inhibition of HCC Cells into Matrigel
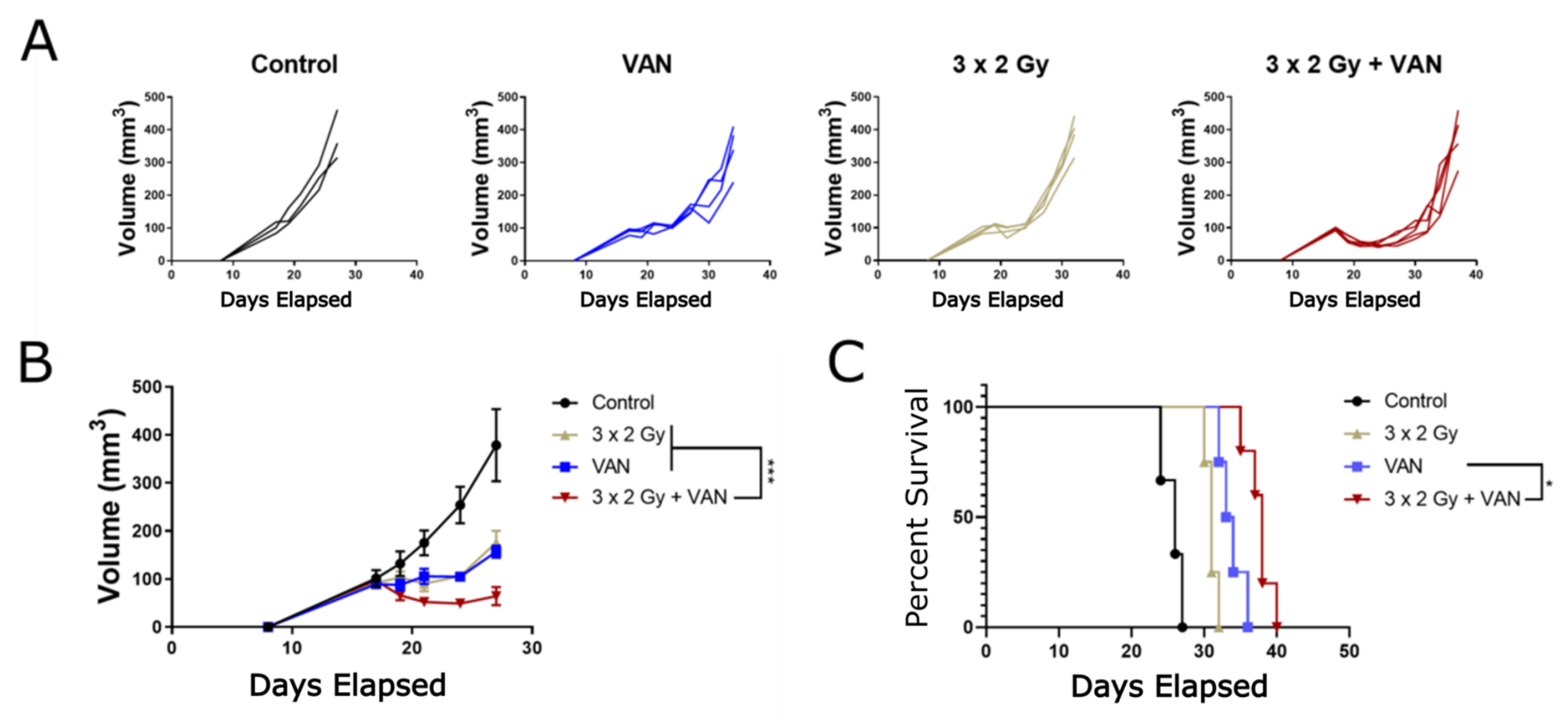
2.6. Vandetanib-Radiation Combination Inhibits Tumour Growth in an Immunocompetent Mouse Model
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Monolayer Cell Culture
4.3. Plasmid Transfection
4.4. X-ray Irradiation of In Vitro Samples
4.5. Western Blotting
4.6. IC Determination
4.7. Clonogenic Survival
4.8. Migration Assay
4.9. 3D Spheroid Cell Culture
4.10. Immunocompetent Animal Model
4.11. BNL.1ME Tumour Engraftment and Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HCC | Hepatocellular Carcinoma | ICC | Intrahepatic Cholangiocarcinoma |
| ECC | Extrahepatic Cholangiocarcinoma | SBRT | Stereotactic Body Radiotherapy |
| TKI | Tyrosine Kinase Inhibitor | PTM | Post-translational Modification |
| VAN | Vandetanib | PARP | Poly ADP Ribose Polymerase |
| DNA | Deoxyribonucleic Acid | VEGF(R) | Vascular Endothelial Growth Factor (Receptor) |
| EGF(R) | Epidermal Growth Factor (Recptor) | RET | REarranged during Transfection Receptor |
| mTOR | Mammalian Target of Rapamycin | GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| 2D | Two Dimensional | 3D | Three Dimensional |
| RER | Radiation Enhancement Ratio | IC | Inhibitory Concentration at 50% Maxima |
| p | probability value | CI | Confidence Interval |
| IQR | Interquartile Range | CDI | Coefficient of Drug Interaction |
| ANOVA | ANalysis of VAriance | MDPI | Multidisciplinary Digital Publishing Institute |
| SI units were used unless otherwise stated. | |||
Appendix A. Mathematical Formulae
Appendix A.1. Coefficient of Drug Interaction
Appendix A.2. Volume Calculation for In Vivo Experiment
References
- McGlynn, K.A.; Tsao, L.; Hsing, A.W.; Devesa, S.S.; Fraumeni, J.F., Jr. International trends and patterns of primary liver cancer. Int. J. Cancer 2001, 94, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Perz, J.F.; Armstrong, G.L.; Farrington, L.A.; Hutin, Y.J.; Bell, B.P. The contributions of hepatitis B virus and hepatitis C virus infections to cirrhosis and primary liver cancer worldwide. J. Hepatol. 2006, 45, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Forner, A.; Reig, M.E.; de Lope, C.R.; Bruix, J. Current strategy for staging and treatment: The BCLC update and future prospects. Semin. Liver Dis. 2010, 30, 61–74. [Google Scholar] [CrossRef]
- Liu, C.Y.; Chen, K.F.; Chen, P.J. Treatment of Liver Cancer. Cold Spring Harb. Perspect. Med. 2015, 5, a021535. [Google Scholar] [CrossRef]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef]
- Sharma, R.A.; Plummer, R.; Stock, J.K.; Greenhalgh, T.A.; Ataman, O.; Kelly, S.; Clay, R.; Adams, R.A.; Baird, R.D.; Billingham, L.; et al. Clinical development of new drug-radiotherapy combinations. Nat. Rev. Clin. Oncol. 2016, 13, 627–642. [Google Scholar] [CrossRef]
- Pignon, J.P.; Bourhis, J.; Domenge, C.; Designé, L. Chemotherapy added to locoregional treatment for head and neck squamous-cell carcinoma: Three meta-analyses of updated individual data. Lancet 2000, 355, 949–955. [Google Scholar] [CrossRef]
- Rose, P.G. Chemoradiotherapy for cervical cancer. Eur. J. Cancer 2001, 38, 270–278. [Google Scholar] [CrossRef]
- van Hagen, P.; Hulshof, M.C.; van Lanschot, J.J.; Steyerberg, E.W.; van Berge Henegouwen, M.I.; Wijnhoven, B.P.; Richel, D.J.; Nieuwenhuijzen, G.A.; Hospers, G.A.; Bonenkamp, J.J.; et al. Preoperative chemoradiotherapy for esophageal or junctional cancer. N. Engl. J. Med. 2012, 366, 2074–2084. [Google Scholar] [CrossRef]
- Lacas, B.; Bourhis, J.; Overgaard, J.; Zhang, Q.; Grégoire, V.; Nankivell, M.; Zackrisson, B.; Szutkowski, Z.; Suwiński, R.; Poulsen, M.; et al. Role of radiotherapy fractionation in head and neck cancers (MARCH): An updated meta-analysis. Lancet Oncol. 2017, 18, 1221–1237. [Google Scholar] [CrossRef]
- Lesueur, P.; Chevalier, F.; Austry, J.B.; Waissi, W.; Burckel, H.; Noel, G.; Habrand, J.L.; Saintigny, Y.; Joly, F. Poly-(ADP-ribose)-polymerase inhibitors as radiosensitizers: A systematic review of pre-clinical and clinical human studies. Oncotarget 2017, 8, 69105–69124. [Google Scholar] [CrossRef] [PubMed]
- Button, R.W.; Vincent, J.H.; Strang, C.J.; Luo, S.Q. Dual PI-3 kinase/mTOR inhibition impairs autophagy flux and induces cell death independent of apoptosis and necroptosis. Oncotarget 2016, 7, 5157–5175. [Google Scholar] [CrossRef] [PubMed]
- Wedge, S.R.; Ogilvie, D.J.; Dukes, M.; Kendrew, J.; Chester, R.; Jackson, J.A.; Boffey, S.J.; Valentine, P.J.; Curwen, J.O.; Musgrove, H.L.; et al. ZD6474 inhibits vascular endothelial growth factor signaling, angiogenesis, and tumor growth following oral administration. Cancer 2002, 62, 4645–4655. [Google Scholar]
- Ryan, A.J.; Wedge, S.R. ZD6474—A novel inhibitor of VEGFR and EGFR tyrosine kinase activity. Br. J. Cancer 2005, 92 (Suppl. S1), S6–S13. [Google Scholar] [CrossRef] [PubMed]
- Motz, G.T.; Coukos, G. The parallel lives of angiogenesis and immunosuppression: Cancer and other tales. Nat. Rev. Immunol. 2011, 11, 702–711. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Principles and mechanisms of vessel normalization for cancer and other angiogenic diseases. Nat. Rev. Drug Discov. 2011, 10, 417–427. [Google Scholar] [CrossRef]
- Morse, M.A.; Sun, W.; Kim, R.; He, A.R.; Abada, P.B.; Mynderse, M.; Finn, R.S. The Role of Angiogenesis in Hepatocellular Carcinoma. Clin. Cancer Res. 2018. [Google Scholar] [CrossRef]
- Finn, R.S.; Aleshin, A.; Dering, J.; Yang, P.; Ginther, C.; Desai, A.; Zhao, D.; von Euw, E.; Busuttil, R.W.; Slamon, D.J. Molecular subtype and response to dasatinib, an Src/Abl small molecule kinase inhibitor, in hepatocellular carcinoma cell lines in vitro. Hepatology 2013, 57, 1838–1846. [Google Scholar] [CrossRef]
- Llovet, J.M.; Villanueva, A.; Lachenmayer, A.; Finn, R.S. Advances in targeted therapies for hepatocellular carcinoma in the genomic era. Nat. Rev. Clin. Oncol. 2015, 12, 408–424. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef]
- Hamzah, J.; Jugold, M.; Kiessling, F.; Rigby, P.; Manzur, M.; Marti, H.H.; Rabie, T.; Kaden, S.; Grone, H.J.; Hammerling, G.J.; et al. Vascular normalization in Rgs5-deficient tumours promotes immune destruction. Nature 2008, 453, 410–414. [Google Scholar] [CrossRef]
- Jain, A.; Jain, A.; Gulbake, A.; Hurkat, P.; Jain, S.K. Solid Tumors A Review. Int. J. Pharm. Pharm. Sci. 2011, 3 (Suppl. 5), 45–51. [Google Scholar]
- Ansiaux, R.; Dewever, J.; Grégoire, V.; Feron, O.; Jordan, B.F.; Gallez, B. Decrease in Tumor Cell Oxygen Consumption after Treatment with Vandetanib (ZACTIMA; ZD6474) and its Effect on Response to Radiotherapy. Cancer Res. 2009, 172, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.J.; Telfer, B.A.; Brave, S.; Kendrew, J.; Whittaker, L.; Stratford, I.J.; Wedge, S.R. ZD6474, a potent inhibitor of vascular endothelial growth factor signaling, combined with radiotherapy: Schedule-dependent enhancement of antitumor activity. Clin. Cancer Res. 2004, 10, 8587–8593. [Google Scholar] [CrossRef] [PubMed]
- Brazelle, W.D.; Shi, W.; Siemann, D.W. VEGF-associated tyrosine kinase inhibition increases the tumor response to single and fractionated dose radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2006, 65, 836–841. [Google Scholar] [CrossRef]
- Frederick, B.; Gustafson, D.; Bianco, C.; Ciardiello, F.; Dimery, I.; Raben, D. ZD6474, an inhibitor of VEGFR and EGFR tyrosine kinase activity in combination with radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2006, 64, 33–37. [Google Scholar] [CrossRef]
- Shibuya, K.; Komaki, R.; Shintani, T.; Itasaka, S.; Ryan, A.; Jurgensmeier, J.M.; Milas, L.; Ang, K.; Herbst, R.S.; O’Reilly, M.S. Targeted therapy against VEGFR and EGFR with ZD6474 enhances the therapeutic efficacy of irradiation in an orthotopic model of human non-small-cell lung cancer. Int. J. Radiat. Oncol. Biol. Phys. 2007, 69, 1534–1543. [Google Scholar] [CrossRef][Green Version]
- Wachsberger, P.; Burd, R.; Ryan, A.; Daskalakis, C.; Dicker, A.P. Combination of vandetanib, radiotherapy, and irinotecan in the LoVo human colorectal cancer xenograft model. Int. J. Radiat. Oncol. Biol. Phys. 2009, 75, 854–861. [Google Scholar] [CrossRef][Green Version]
- Heffelfinger, S.C.; Hawkins, H.H.; Barrish, J.; Taylor, L.; Darlington, G.J. SK HEP-1: A human cell line of endothelial origin. In Vitro Cell. Dev. Biol. 1992, 28, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Castilho-Fernandes, A.; de Almeida, D.C.; Fontes, A.M.; Melo, F.U.; Picanço-Castro, V.; Freitas, M.C.; Orellana, M.D.; Palma, P.V.; Hackett, P.B.; Friedman, S.L.; et al. Human hepatic stellate cell line (LX-2) exhibits characteristics of bone marrow-derived mesenchymal stem cells. Exp. Mol. Pathol. 2011, 91, 664–672. [Google Scholar] [CrossRef]
- Chandler, K.B.; Leon, D.R.; Kuang, J.; Meyer, R.D.; Rahimi, N.; Costello, C.E. N-glycosylation regulates ligand-dependent activation and signaling of vascular endothelial growth factor receptor 2 (VEGFR2). J. Biol. Chem. 2019, 294, 13117–13130. [Google Scholar] [CrossRef] [PubMed]
- Collet, G.; Lamerant-Fayel, N.; Tertil, M.; El Hafny-Rahbi, B.; Stepniewski, J.; Guichard, A.; Foucault-Collet, A.; Klimkiewicz, K.; Petoud, S.; Matejuk, A.; et al. Hypoxia-Regulated Overexpression of Soluble VEGFR2 Controls Angiogenesis and Inhibits Tumor Growth. Mol. Cancer Ther. 2014, 13, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.F.; Shao, M.H.; Gao, P.T.; Ma, J.; Li, H.J.; Li, G.L.; Han, B.H.; Yuan, C.G. The important roles of RET, VEGFR2 and the RAF/MEK/ERK pathway in cancer treatment with sorafenib. Acta Pharmacol. Sin. 2012, 33, 1311–1318. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Y.; Zou, H.; Cai, N.; Chen, X.; Xu, L.; Kong, X.; Liu, P. The multi-targeted tyrosine kinase inhibitor vandetanib plays a bifunctional role in non-small cell lung cancer cells. Sci. Rep. 2015, 5, 8629. [Google Scholar] [CrossRef]
- Giannelli, G.; Azzariti, A.; Sgarra, C.; Porcelli, L.; Antonaci, S.; Paradiso, A. ZD6474 inhibits proliferation and invasion of human hepatocellular carcinoma cells. Biochem. Pharmacol. 2006, 71, 479–485. [Google Scholar] [CrossRef]
- Chou, T.-C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzym. Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Denys, A.; Czuczman, P.; Grey, D.; Bascal, Z.; Whomsley, R.; Kilpatrick, H.; Lewis, A.L. Vandetanib-eluting Radiopaque Beads: In vivo Pharmacokinetics, Safety and Toxicity Evaluation following Swine Liver Embolization. Theranostics 2017, 7, 2164–2176. [Google Scholar] [CrossRef]
- Fuchs, K.; Duran, R.; Denys, A.; Bize, P.E.; Borchard, G.; Jordan, O. Drug-eluting embolic microspheres for local drug delivery—State of the art. J. Control. Release 2017, 262, 127–138. [Google Scholar] [CrossRef]
- Hagan, A.; Phillips, G.J.; Macfarlane, W.M.; Lloyd, A.W.; Czuczman, P.; Lewis, A.L. Preparation and characterisation of vandetanib-eluting radiopaque beads for locoregional treatment of hepatic malignancies. Eur. J. Pharm. Sci. 2017, 101, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Amaoka, N.; Saio, M.; Nonaka, K.; Imai, H.; Tomita, H.; Sakashita, F.; Takahashi, T.; Sugiyama, Y.; Takami, T.; Adachi, Y. Expression of vascular endothelial growth factor receptors is closely related to the histological grade of hepatocellular carcinoma. Oncol. Rep. 2006, 16, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Aucejo, F.; Kim, R.; Zein, N.; Quintini, C.; Uso, T.D.; Lopez, R.; Eghtesad, B.; Fung, J.; Miller, C.; Yerian, L. Vascular endothelial growth factor receptor 2 expression in non-tumorous cirrhotic liver is higher when hepatoma is beyond Milan criteria. Liver Transplant. 2009, 15, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K.; Tong, R.T.; Munn, L.L. Effect of Vascular Normalization by Antiangiogenic Therapy on Interstitial Hypertension, Peritumor Edema, and Lymphatic Metastasis: Insights from a Mathematical Model. Cancer Res. 2007, 67, 2729–2735. [Google Scholar] [CrossRef] [PubMed]
- Staton, C.A.; Reed, M.W.; Brown, N.J. A critical analysis of current in vitro and in vivo angiogenesis assays. Int. J. Exp. Pathol. 2009, 90, 195–221. [Google Scholar] [CrossRef]
- Steel, G.G. Terminology in the description of drug-radiation interactions. Int. J. Radiat. Oncol. Biol. Phys. 1979, 5, 1145–1150. [Google Scholar] [CrossRef]
- Böckelmann, L.C.; Schumacher, U. Targeting tumor interstitial fluid pressure: Will it yield novel successful therapies for solid tumors? Expert Opin. Ther. Targets 2019, 23, 1005–1014. [Google Scholar] [CrossRef]
- Multhoff, G.; Vaupel, P. Radiation-induced changes in microcirculation and interstitial fluid pressure affecting the delivery of macromolecules and nanotherapeutics to tumors. Front. Oncol. 2012, 2, 165. [Google Scholar] [CrossRef]
- Boucher, Y.; Baxter, L.T.; Jain, R.K. Interstitial Pressure Gradients in Tissue-Isolated and Subcutaneous Tumors: Implications for Therapy. Cancer Res. 1990, 50, 4478–4484. [Google Scholar]
- Davies, C.d.L.; Lundstrøm, L.M.; Frengen, J.; Eikenes, L.; Bruland, Ø.S.; Kaalhus, O.; Hjelstuen, M.H.B.; Brekken, C. Radiation Improves the Distribution and Uptake of Liposomal Doxorubicin (Caelyx) in Human Osteosarcoma Xenografts. Cancer Res. 2004, 64, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Hu, C.; Hui, K.; Jiang, X. The role and significance of VEGFR2+ regulatory T cells in tumor immunity. Onco Targets Ther. 2017, 10, 4315–4319. [Google Scholar] [CrossRef] [PubMed]
- Sheen, J.H.; Zoncu, R.; Kim, D.; Sabatini, D.M. Defective regulation of autophagy upon leucine deprivation reveals a targetable liability of human melanoma cells in vitro and in vivo. Cancer Cell 2011, 19, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Bacterial Transformation. Available online: https://www.addgene.org/protocols/bacterial-transformation/ (accessed on 2 July 2018).
- Poirier, Y.; Johnstone, C.D.; Kirkby, C. The potential impact of ultrathin filter design on dosimetry and relative biological effectiveness in modern image-guided small animal irradiators. Br. J. Radiol. 2019, 92, 20180537. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Vinci, M.; Box, C.; Eccles, S.A. Three-dimensional (3D) tumor spheroid invasion assay. J. Vis. Exp. 2015. [Google Scholar] [CrossRef]
- Hou, Y.; Konen, J.; Brat, D.J.; Marcus, A.I.; Cooper, L.A.D. TASI: A software tool for spatial-temporal quantification of tumor spheroid dynamics. Sci. Rep. 2018, 8, 7248. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Source/Access. No. | Donor Patient | Derivation | Cell Type | Etiology |
|---|---|---|---|---|---|
| SK-Hep-1 | ATCC HTB-52 | 52 M | Adenocarcinoma | endothelial/ mesenchymal [31] | |
| Hep G2 | ECACC 85011430 | 15 M Caucasian | HCC | epithelial | |
| Hep3B | ECACC 86062703 | 8 M Black | Hepatocyte carcinoma | epithelial | |
| JHH2 | JCRB1028 | 57 M Japanese | Hepatoma | ||
| JHH4 | JCRB0435 | 51 M Japanese | HCC | ||
| JHH5 | JCRB1029 | 50 M Japanese | HCC | ||
| JHH6 | JCRB1030 | 57 F Japanese | HCC | epithelial | |
| JHH7 | JCRB1031 | 53 M Japanese | HCC | epithelial | HepB |
| HLE | JCRB0404 | 68 M | Hepatoma | ||
| HLF | JCRB0405 | 68 M | Hepatoma | epithelial | |
| PLC-PRF-5 | ATCC CRL-8024 | Hepatoma | epithelial | HepB | |
| Huh1 | JCRB0199 | 53 M Japanese | HCC | epithelial | HepB |
| Huh7-D12 | ECACC 010427 | 12 M | HCC | epithelial | |
| SNU423 | ATCC CRL-2238 | 40 M Asian | pleo. HCC | epithelial | HepB |
| SNU387 | ATCC CRL-2237 | 41 F Asian | pleo. HCC | epithelial | HepB |
| SNU449 | ATCC CRL-2234 | 52 M Asian | HCC | epithelial | HepB |
| SNU1079 | B. Dwyer ‡ | ICC | |||
| Huh28 | B. Dwyer ‡ | ICC | |||
| CC-LP-1 | B. Dwyer ‡ | ICC | |||
| CC-SW-1 | B. Dwyer ‡ | ICC | |||
| LX-2 | K. Rombouts § | Hepatic Stellate Cell [32] | |||
| HUVEC | Lonza CC-2935 | Umbilical Vein | endothelial |
| Cell Line | Source/Accession No. | Mouse Background | Deriv. |
|---|---|---|---|
| Hep-55.1C | CLS-GmbH | C57BL/6 Mouse | HCC |
| Hepa1-6 | CLS-GmbH | C57BL/6J Mouse | HCC |
| BNL1ME A.7R.1 | ATCC TIB-75 | BALB/C Mouse | HCC/ epithelial |
| NIH3T3 | ATCC CRL-1658 | mouse embryo | fibroblast |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Znati, S.; Carter, R.; Vasquez, M.; Westhorpe, A.; Shahbakhti, H.; Prince, J.; Vlckova, P.; De Vellis, C.; Bascal, Z.; Loizidou, M.; et al. Radiosensitisation of Hepatocellular Carcinoma Cells by Vandetanib. Cancers 2020, 12, 1878. https://doi.org/10.3390/cancers12071878
Znati S, Carter R, Vasquez M, Westhorpe A, Shahbakhti H, Prince J, Vlckova P, De Vellis C, Bascal Z, Loizidou M, et al. Radiosensitisation of Hepatocellular Carcinoma Cells by Vandetanib. Cancers. 2020; 12(7):1878. https://doi.org/10.3390/cancers12071878
Chicago/Turabian StyleZnati, Sami, Rebecca Carter, Marcos Vasquez, Adam Westhorpe, Hassan Shahbakhti, Jessica Prince, Petra Vlckova, Chiara De Vellis, Zainab Bascal, Marilena Loizidou, and et al. 2020. "Radiosensitisation of Hepatocellular Carcinoma Cells by Vandetanib" Cancers 12, no. 7: 1878. https://doi.org/10.3390/cancers12071878
APA StyleZnati, S., Carter, R., Vasquez, M., Westhorpe, A., Shahbakhti, H., Prince, J., Vlckova, P., De Vellis, C., Bascal, Z., Loizidou, M., & Sharma, R. A. (2020). Radiosensitisation of Hepatocellular Carcinoma Cells by Vandetanib. Cancers, 12(7), 1878. https://doi.org/10.3390/cancers12071878